
Abstract
Great effort has been dedicated to mapping the functional architecture of the brain in health and disease. The neural centers that support cognition and behavior are the “hubs” defining the salient geographic landmarks of the cerebral topography. Similar to urban cartography, however, the functionality of these hubs is critically dependent on the infrastructure permitting the transfer of relevant information from site to site, and this infrastructure is susceptible to deterioration. The groundwork of the brain lies in the form of the complexly organized myelinated nerve fibers responsible for the inter-regional transmission of electrical impulses among distinct neural areas. Damage to the myelin sheath and reduction in the total number of nerve fibers with aging are thought to result in a degradation in the efficiency of communication among neural regions and to contribute to the decline of function in older adults. This article describes selected studies that are relevant to understanding the deterioration in structural connectivity of the aging brain with a focus on potential consequences to functional network activity. First, the neural substrates of connectivity and techniques used in the study of connectivity are described with a focus on neuroimaging methodologies. This is followed with discussion of the negative effects of age on connective integrity, and the possible mechanisms and neural and cognitive consequences of this progressive disconnection. Given the potential for natural repair of certain elements of the connective network, understanding the basis of age-associated decline in connectivity could have important implications with regard to the amelioration of neural dysfunction and the restoration of the infrastructure necessary for optimal function in older adults.
The Broad Spectrum of Function in Older Adults
T
The literature on the associated topics of this article is vast. The focus here is on a selected portion of this work, describing subtle effects of aging on the connective integrity of neural fibers and the potential etiologies and consequences of this damage. Altered connectivity in AD is additionally reviewed as a sole contrast to non-demented aging; however, it should be acknowledged that several other conditions including advanced cerebrovascular disease are of similar importance. Additionally, in the context of neuroimaging, connectivity is discussed here with reference to properties of the cerebral white matter, as structural connectivity at finer scales (e.g., cortical laminar connectivity) is not readily available through standard imaging procedures. It should be noted, however, that a portion of the histological findings discussed refer to effects in myelinated nerve fibers outside the cerebral white matter.
Neuroimaging Structural Connectivity
Structural connectivity refers to the relatively static properties that govern the degree of the direct anatomical linkage across brain regions. Classically, such information was gained through histological dissection and tract tracing techniques, thus allowing the mapping of fine interregional structural connectivity [for historic review of this research and techniques, see (Lanciego and Wouterlood, 2011; Schmahmann and Pandya, 2007)]. Adaptations of these procedures are now noninvasively performed with diffusion imaging as well as through manganese contrast magnetic resonance imaging (MRI)-based tract tracing in animals. Variation in structural connectivity is associated with native behavioral performance, and there is trophic modulation in connectivity with development, environmental enrichment, and learning and plasticity (Fields, 2005; Lebel and Beaulieu, 2011). Neuroimaging procedures allow unprecedented opportunity to examine these phenomena in large human participant samples. The cellular architecture of these links is also susceptible to degenerative abnormalities with aging and disease, and neuroimaging measures of structural connectivity hold promise for assessing specific histologic and pathologic properties of degeneration (Budde et al., 2007); however, etiologic specificity of MRI measures is still lacking.
Assessment of the connective anatomy and integrity, or the health of brain white matter via neuroimaging can be divided into three broad technical categories; macrostructural morphometry, microstructural integrity, and tract-based anatomy. Within each of these domains are a host of data acquisition and analysis procedures differentially sensitive to various underlying neural tissue properties. Morphometry of the aging brain via neuroimaging procedures dates back to early computed tomography research (Thaler et al., 1978), and has been performed using MRI techniques for more than two decades (Jernigan et al., 1991). Macrostructural measures thought to reflect the connective integrity of the brain include the total amount of white matter tissue (the “volume” of the white matter) as well as the total amount of abnormal white matter, referred to as white matter hyperintensities or white matter signal abnormalities (WMSAs; Fig. 1) (Fazekas et al., 1988). There are global and regional decreases in white matter volume with aging and dementia (Guttmann et al., 1998; Raz et al., 1997; Salat et al., 1999, 2009a). In an appreciable subset of older adults, but not all, there is an increase in WMSA. Fazekas and colleagues referred to WMSA as “hyperintense white matter foci on T2-weighted images” (Fazekas et al., 1987, 1988). The presence of WMSA implies damage to the tissue; however, there are many phenomena that can contribute to signal change (Schmidt et al., 2011). White matter signal changes are most often measured in the literature as increased signal on T2 and fluid attenuated inversion recovery (FLAIR) imaging due to the sensitivity of these modalities. However, damage to the white matter results in altered signal on other imaging modalities as well (e.g., hypointense signal on T1). Therefore, a more general use of the term WMSA to refer to any abnormal signal on MRI within the white matter may be appropriate and is used as such throughout this article. Estimates of the prevalence of WMSA are broad and depend on the sample studied. For example, Soderlund and colleagues (2003) found a high 90% prevalence of subcortical hyperintensities and 67% periventricular hyperintensities in a sample of individuals 64–74 years of age initially randomly selected from the community dwelling population. Given that in even the highest prevalence estimates a substantial portion of older adults do not have such markers is of particular importance, as this demonstrates that certain tissue changes are not a necessary consequence of aging. Volumetric measures of white matter damage are associated with several age-related conditions, and early work demonstrated the potential for these lesions to affect neural function (DeCarli et al., 1995); however, such measures are generally lacking in specificity to any particular condition. This may be due to the fact that many studies examine these measures as a composite volume across the entire white matter mass. Examination of macrostructure based on subdivisions of the white matter, for example, based on lobar or gyral distinctions, enhances sensitivity to detect regional distinctions among groups and associations with clinical parameters [e.g., (Murray et al., 2010; Salat et al., 2009a)]; however, difficulties in obtaining neuranatomically principled regional measurements within this tissue from standard structural MRI have limited such application. An additional constraint of volumetric measurements is that they do not typically provide information about the integrity of the tissue within the overall volumetric compartment, regardless of whether it is deemed normal or abnormal tissue. Thus, it is possible that, although two individuals have similar total normal or abnormal white matter volumes, the health of the tissue within those volumes, for example, the degree of myelination, may differ substantially between them.

Macrostructural measures of white matter integrity. Top panel: white matter signal abnormalities (WMSAs) are found in a substantial portion of scans of older adults and in several disease states. Although properties of the abnormal signal can vary, a typical WMSA is dark on T1 (left; red arrow), bright on T2 (second from left), and bright on proton density imaging (third from left). Segmentation algorithms can be utilized to distinguish between normal white matter and WMSA in the volumetric assessment of aging. Bottom panel: demonstration of whole-brain segmentation that labels white matter based on a gyral labeling procedure in a younger adult (top) and an older adult (bottom). Differences in regional white matter volume are visually apparent between the younger and older adult, for example, in the superior temporal white matter (yellow arrows).
Compared with macrostructural measurements, microstructural procedures provide a quantitative value of tissue integrity at each sampled location (e.g., at the level of the imaging voxel or region/tract of interest; Fig. 2). Various imaging procedures can be utilized to obtain quantitative microstructural values that are somewhat selectively sensitive to different histologic and pathologic properties. For example, magnetization transfer imaging (MTI) has been used in the quantitative assessment of age-associated white matter deterioration and the differential characterization of WMSA (Spilt et al., 2005, 2006). Magnetic resonance spectroscopy (MRS) has similarly been used to characterize the metabolic profile of WMSA (Constans et al., 1995). We focus here on the application of diffusion tensor imaging (DTI) to map microstructural deterioration in aging and dementia (O'Sullivan et al., 2001; Pfefferbaum et al., 2000; Salat et al., 2005a), as this procedure has been utilized in a rapidly expanding literature on this topic (Salat et al., 2009c). Diffusion properties obtained from MRI are used in the generation of maps differing in their microstructural contrasts. The anisotropy (often termed “fractional anisotropy”; FA) is a measure of the directional coherence of diffusion within a voxel and has been utilized as in indicator of white matter health in much previous work (Fig. 2).
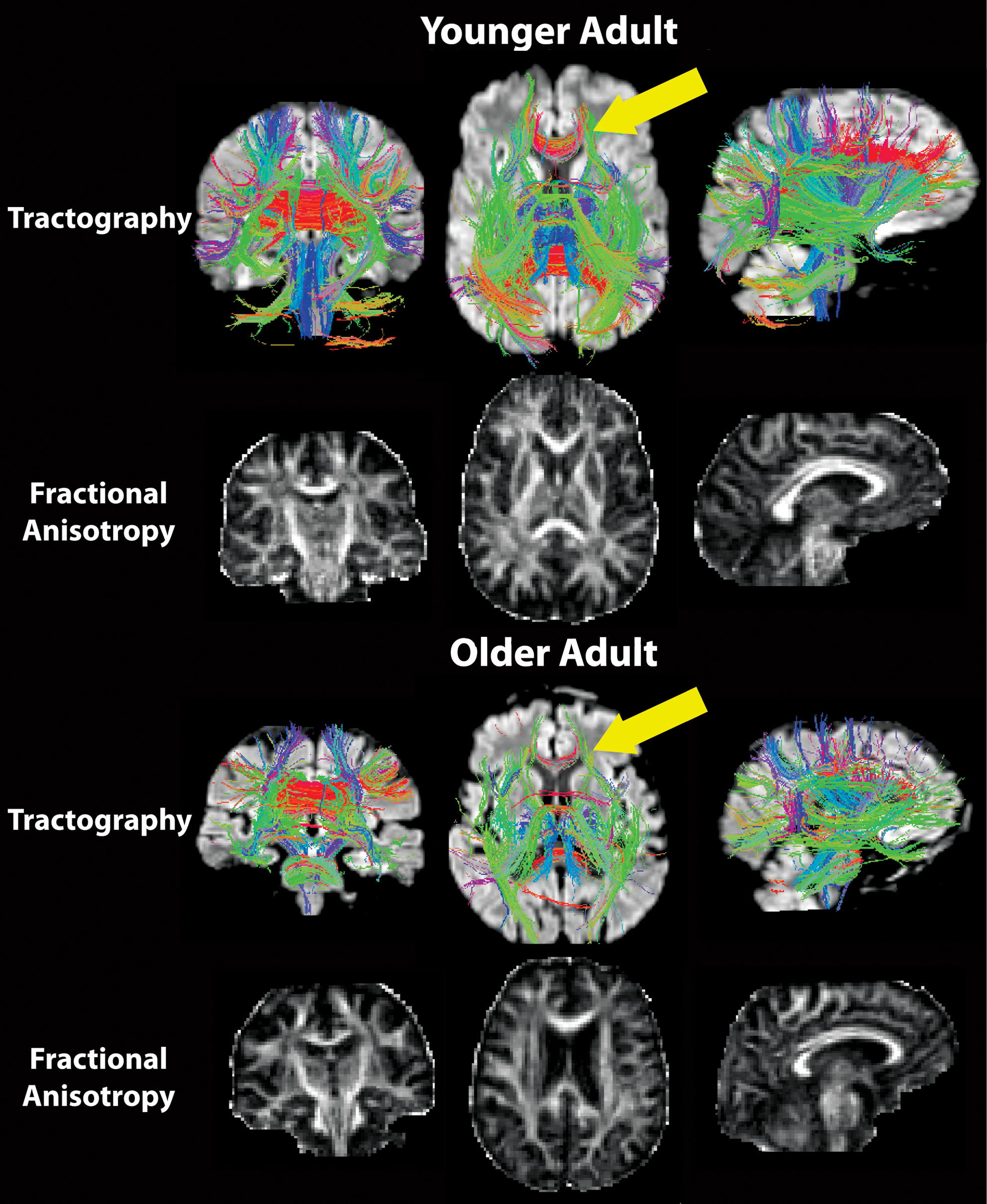
Diffusion imaging of white matter anatomy and microstructure. Diffusion tractography allows for the visualization of the gross fiber anatomy of the brain through the mapping of the primary directionality of water diffusion in the brain. This directionality is dependent on the restrictive environment of diffusion, notably, occurring along the myelinated nerve fibers in the cerebral white matter. Tractography procedures are influenced by changes in the microstructural properties of the tissue. In this case, changes in the white matter integrity in the genu of the corpus callosum with aging result in tractography results, thus suggesting a reduced fiber density in the older adult (bottom yellow arrow; red fibers) compared with the younger adult (top yellow arrow); however, the ability to infer specific histological interpretations from magnetic resonance imaging-based microstructural data is currently limited. The bottom row for each individual shows their fractional anisotropy map, a microstructural marker influenced by the directional properties of water diffusion within white matter. Darkened regional signal in the older adult compared with the younger adult suggests a reduction in the white matter integrity of the older adult.
The procedures just described provide quantitative macrostructural and microstructural metrics of tissue integrity, whereas diffusion tractography instead is used to examine the fascicular anatomy of the brain. Much of the attractiveness of diffusion imaging as a research technique is generated from this previously unavailable contrast for mapping the directionality of large fiber bundles throughout the white matter [for recent technical and applications reviews of DTI, see (Chanraud et al., 2010; Tournier et al., 2011)]. Techniques for diffusion tractography can provide information about the existence or lack of major connections in the brain, and have been used to examine the connectivity between cytoarchitectonically, morphometrically, and functionally distinct neural areas. The connectivity of brain regions can be probed through an evolving set of computational procedures that assess the likelihood and degree of structural links between those regions. Such techniques have a range of applications. In the study of aging and age-associated disease, tractography procedures are typically used to map regions of white matter to be utilized as “tracts of interest” in the examination of the microstructural integrity of specific fiber bundles or neural systems (Davis et al., 2009; Sullivan et al., 2006). Information about the orientation of fibers from the diffusion tensor allows for a more careful regional comparison of white matter properties across individuals than was previously possible. However, it is important that such procedures are based on the tissue diffusion properties, and therefore, their application can be challenging with degenerative alterations in a microstrucutral environment.
Efficiency of Neurotransmission
The efficiency of the transmission of neural signals is dependent on the integrity of interregional connections in the brain. The cerebral white matter consists, to a large degree, of the axonal projections encased in myelin (Fig. 3). The number of nerve fibers comprising the connection, the diameter of the fibers, the degree of myelination, and the specific myelin configuration along the fiber (e.g., node size and spacing) will contribute to the amount to which distinct areas of the brain can communicate, and these divergent parameters are susceptible to changes with aging (Luebke et al., 2010; Peters, 2007). In the central nervous system, the myelin sheaths are outgrowths of oligodendrocytes that form an electrically insulating coil around the axonal membrane. Myelin-based insulation of axonal fibers prevents the loss of current, promotes greater energy efficiency, and increases propagation velocity compared with nonmyelinated fibers due to their use of saltatory conduction mechanisms (Tasaki, 1939). Additionally, the fine tuning of transmission properties that occurs as a result of myelination is a critical factor determining synchronous activity among neurons (Lang and Rosenbluth, 2003). Although myelinated fibers comprise a large portion of the total white matter, additional components of this tissue class include support cells (e.g., oligodendrocytes, astrocytes, and microglia) and blood vessels. Thus, a range of elements may be altered within the white matter with aging and disease, and this should be considered in the interpretation of neuroimaging results. The multifactorial decline in structural connectivity with aging is, therefore, expected to have a substantial impact on the effectiveness of neural communication, and in turn, have a negative influence on cognition and behavior. At more extreme ranges of degeneration, or when affecting specific locations of the brain, deterioration of structural connectivity may even form the basis of the profound symptoms of age-associated neurological disorders such as AD (Bartzokis, 2011). It is, therefore, critical to understand the mechanisms by which neuroanatomical connectivity is compromised with aging, and in turn, how this altered state negatively influences optimal brain function and cognition. It is important to note that, although damage to myelin and loss of fibers has been a major focus in the study of altered connectivity, several other changes occur in this machinery with aging including a reduction in dendritic spines and synapses (Duan et al., 2003), although it is possible that these processes could be, to some degree, inter-related.

Simplified substrates of structural connectivity. The representation illustrates the cellular components of structural connectivity that promote communication across brain regions. Nerve impulses are transmitted from the neuron soma along the axon, and the conduction characteristics of this impulse are influenced by properties of the myelinated fiber including axonal diameter, degree of myelination, and spacing of the nodes between myelin segments. Aging compromises this system with normal patterns of myelination being altered, nerve fibers becoming damaged, and there is a reduction in the total number of myelinated fibers. Such changes could reduce the quality and consistency of nerve impulses, alter critical timing, and degrade the communication of information throughout affected areas. Although much imaging work has focused on age-associated changes in structural connectivity within the white matter, it is important to note that several additional changes impact connectivity including reduction in the number of dendritic spines and total number of synapses.
From Localization to Interaction
Previous models of brain aging have emphasized the selective vulnerability of specific neural regions. In particular, accumulation of a wealth evidence from neuropsychological, neuropathological, and structural and functional brain imaging have supported a “frontal aging” hypothesis, thus suggesting that the frontal lobe is substantially more vulnerable to the detrimental effects of aging than other portions of the brain [for review, see (Greenwood, 2000)]. Although it is clear that the frontal lobe exhibits considerable deterioration with aging, limitations on this model led Greenwood to call instead for a paradigm of brain aging that emphasized disruption in the interactions among brain regions, as opposed to the selective vulnerability of any particular region (Greenwood, 2000). Localization and network-based models of brain aging are not mutually exclusive, as known from the early work on disconnection by Norman Geschwind and the thesis work of Carl Wernicke before him (Catani and ffytche, 2005). The salient argument of Greenwood's critique, however, was that there are a myriad of changes, from cellular and metabolic to morphometric and functional, which occur as a consequence of advancing age throughout the brain, and that the impact of degenerative processes on cognition would, therefore, be best understood through network-based approaches; concluding that,
“Use of such a network approach does not minimize the processing role of the frontal lobes, but serves to question the explanatory power of a localizationist approach such as the frontal aging hypothesis to questions of cognitive and brain aging.”
(Greenwood, 2000)
In this regard, recent trends in neuroimaging research have shifted from analyses focused on regional localization to “connectomics”; the study of the connective anatomy of the brain across a range of anatomical and neurophysiological scales (Sporns, 2011). The utilization of network approaches to brain function and cognition permit a vastly expanded range of possible models of age-associated decline; however, this framework also greatly increases the complexity of the problem. Thus, continued development of sophisticated procedures for the integration of information from the structural and functional domains will benefit our current understandings about neural aging and cognitive decline.
Functional Connectivity
Continuing evolution in MRI image acquisition and analysis procedures has resulted in this technique being a primary tool for the measurement of the structural and functional connectivity of the brain. This more recent work is based in a rich scientific exploration of the structural and functional connective anatomy of the brain through histologic and invasive anatomic and neurophysiologic techniques in animal models and certain patient populations. With regard to function, electrophysiological studies have referred to the “functional connectivity” of individual neurons and neural circuits measured by the cross-correlation of neuronal spike trains for quite some time (Feldt et al., 2011). Coherence procedures were adapted for the noninvasive measurement of connectivity of neural signals through electroencephalography (EEG) in human studies (Sauseng and Klimesch, 2008). Although limited in temporal resolution compared with EEG, the versatility of MRI for data acquisition and evolution in the fields of data processing and image analysis have greatly broadened the collection of tools available to examine the connectome. Correlated activity in the functional MRI (fMRI) blood oxygenation level dependent signal across regions provides an estimate of the communication, or functional connectivity, between those regions. Strikingly, covariance of neural signal exists across brain regions even in the absence of an experimental paradigm, when the brain is in a “resting” state (functional connectivity MRI; fcMRI) (Biswal et al., 1995; Cole et al., 2010; Raichle, 2011; Van Dijk et al., 2010). Although the origins of measured signal covariance are yet to be fully determined, particularly in the study of aging (Kannurpatti et al., 2011), a wealth of information supports that a substantial portion of this phenomenon is due to regional neural interactions driven by anatomical connectivity and by common energy demands with a functional purpose [e.g., (Quigley et al., 2003)]. Thus, it is of great interest to understand the impact of age-associated decline in connective integrity on the synchronized neural processes measured in the brain via MRI.
Histologic and Pathologic Properties
Histologic studies in humans and animal models have demonstrated the vulnerability of white matter, as well as the myelinated fibers within gray matter, to degenerative changes with aging and age-associated disease. Histological changes span a range of phenomena, including a reduction in the total number of myelinated nerve fibers and alterations in the myelin sheath (Meier-Ruge et al., 1992; Peters, 2007; Tang et al., 1997). The consequence of changes in certain tissue constituents, and subsequent atrophy of the tissue includes a relative increase in the capillary density within similar measuring fields [up to 20% in the corpus callosum; (Meier-Ruge et al., 1992)], and this is despite the fact that there may be an overall preservation or decrease in total capillary density with aging [for review see (Brown and Thore, 2011)]. Degenerative changes in the myelin sheaths of existing fibers have been described through electron microscopy studies of nonhuman primates and include myelin splitting as well as abnormalities including separations in the sheath and abnormal fluid-filled expansions, or “balloons”, dense axoplasm, and sheaths that abnormally fit their corresponding axonal projection (redundant sheaths) (Bowley et al., 2010; Peters, 2007; Peters and Rosene, 2003). It is possible that the reduction in myelinated nerve fibers reported in postmortem human studies (Marner et al., 2003; Meier-Ruge et al., 1992; Tang et al., 1997) is due to a vulnerability of the smaller-diameter fibers (Tang et al., 1997). This preferential susceptibility may provide one explanation for accelerated changes in certain regions of the brain with aging. Other detectable alterations in connective integrity in human postmortem tissue include evidence of microglial and endothelial activation, enlarged perivascular spaces (Virchow-Robin spaces), vasogenic edema, and granular ependymitis, a form of gliosis in the cellular layer bordering the lateral ventricles. The finding of loss of myelinated nerve fibers in the relative (but potentially not complete) absence of neuronal death led Peters and Rosene to surmise that with nondiseased aging,
“It is possible that the loss of nerve fibers preferentially effects long range connections, as neuronal loss is reported to be minimal, and therefore the loss of fibers is due to a selective degeneration of the long projecting axons with relative preservation of the soma.” (Peters and Rosene, 2003)
This is in contrast to neurodegenerative diseases such as AD, which impacts fibers (Englund and Brun, 1990) as well as promotes death of the neuron, thus effectively isolating the hippocampal formation from cortical input (Geula, 1998; Hyman et al., 1984). This brief discussion of pathologies highlights the complexity in trying to understand aging based on a specific type of degenerative tissue change. A number of the noted properties affected by aging would be expected to modulate tissue integrity measured by MRI, thus making a simple interpretation of results unlikely. However, it is of great interest that with degeneration, there is a range of possible tissue contrasts to be exploited with neuroimaging to better understand these phenomena.
Alterations in white matter tissue properties and disruption of myelin is expected to have functional consequences on the physiology of altered fibers (Aston-Jones et al., 1980), and the efficiency of the functional connectivity of neural regions would be affected by reduction in overall connectivity (loss of fibers) as well as decreased efficiency and timing precision of remaining connections (Fig. 4). Conduction velocity of neural signal declines with aging in animals (Aston-Jones et al., 1985) and humans (Kimura, 1984), and data support that damage to the myelin sheath contributes to such changes. For example, Adinolfi and colleagues (1991) examined the ultrastructure of the sciatic nerve in aged cats and found a range of focal abnormalities of the myelin sheath, including lipid-like droplets, granulo-vacuolar debris in the myelinating glial cells, and disruption of segments of the myelin sheath by interlamellar splitting and ballooning. An accompanying decrease in conduction velocity of similar nerve fibers was found in older animals, and the authors posited that this was due to the myelin abnormalities (Adinolfi et al., 1991). Sato and colleagues (1985) demonstrated a decrease in conduction velocity in myelinated fibers but a relative preservation of velocity in nonmyelinated fibers of aged rats, thus suggesting that changes in myelin with aging contribute to altered function. Reduced conduction velocity with damage to the white matter could decrease the fidelity of neural synchrony and have consequences on cognition and behavior.

Function is dependent on structure. Network models of neural function emphasize interactions among brain regions that have fine spatiotemporal tuning to support optimal cognitive and behavioral function. Performance of this system is dependent on the structural integrity of the connective substrates linking otherwise distinct brain regions.
Neuroimaging White Matter in Aging
A growing neuroimaging literature has documented and reviewed the effects of aging and age-associated disease on white matter integrity [recently reviewed in: (Brickman et al., 2009; Gunning-Dixon et al., 2009; Madden et al., 2011; Salat et al., 2009c)]. There is little disagreement about the vulnerability of white matter tissue structure to age-associated pathologies; however, the regional and pathologic specificity of this vulnerability and the resulting clinical significance of degenerative changes is much less clear. To summarize some major findings to date, imaging evidence of degenerative changes in white matter are found throughout a substantial portion of the brain with some predilection for frontal tissue (Head et al., 2004; Salat et al., 2005a, 2005b, 2009a; Sullivan et al., 2006; Pfefferbaum et al., 2000, 2005). Microstructural measures can be used to detect differences by middle age (Salat et al., 2005a), and both microstructural and volumetric white matter measures show accelerated alterations in late mid-age (Guttmann et al., 1998; Hasan et al., 2010; Hopkins et al., 2006; Salat et al., 2009a). These finding are supported by effects of age on myelin which are detectable by middle age in the rhesus monkey, and that the frequency of degenerating fibers may substantially increase after this time (Bowley et al., 2010). It is unclear why changes become so prominent with mid to later aging, but it is of interest that factors related to vascular health in midlife are associated with white-matter changes in later life (Debette et al., 2011b). Continued changes in vascular health with increasing age may compound the effects of aging (Kennedy and Raz, 2009; Leritz et al., 2010; Salat et al., 2011b) and the risk for AD, which also affects white matter microstructure (Salat et al., 2010; Zhang et al., 2009), increases past middle age.
It is an open question as to the pathological basis of the subtle microstructural alterations in older adults detectable by diffusion imaging. Interpretations of diffusion-based microstructural measures from studies of animal models should be made with caution; however, prominent changes in the radial component of diffusivity with aging (diffusion perpendicular to the primary fiber orientation) suggests that much of the effect of age on white matter may be a consequence of degradation of the myelin (Bennett et al., 2010; Davis et al., 2009; Zhang et al., 2010).
Although the more prominent, visually apparent WMSA have been associated with a range of clinical phenomena, specific links between this presumed damage and unique clinical correlates are lacking. Given the high prevalence of WMSA across a variety of age-associated conditions with very different associated dysfunction (or lack thereof), including nondiseased aging, previous investigators have noted the difficulty in ascribing a more substantial clinical significance to such changes (Zimmerman et al., 1986). For example, although changes in periventricular white matter signal abnormalities are highly prevalent in patients with demyelinating disease and with hydrocephalus, they are also found across a range of patients and older adults, regardless of diagnosis (Zimmerman et al., 1986). More recent meta-reviews on the topic have noted that WMSA are indeed associated with stroke risk, dementia, and death, and described various study considerations that may account for conflicting data (Brickman et al., 2009; Debette et al., 2011a). From an imaging methodology perspective, there are several additional factors related to procedures for the quantitation of WMSA that may contribute to the difficulty in assigning clinical relevance to such changes. For example, hyperintense signal within the white matter on T2/FLAIR imaging provides a visually evident measure of tissue change that can be quantified as a volume (Pantoni and Garcia, 1997); however, it is likely that a binary classification of change alone is not enough for clinically sensitive measurement. Given the heterogeneous pathology within WMSA, it is clear that there is a need for additional information to complement measures of normal and abnormal white matter volume, and procedures such as MTI may be useful in characterizing this heterogeneity (Spilt et al., 2006). Additionally, it is not entirely obvious whether or how this measure should be normalized. The total volume of WMSA is typically corrected for total intracranial volume, a procedure adapted from standard volumetric measurements of brain structures; however, this correction assumes that a scaling relationship between WMSA volume and head size exists, and this is not evident, as many individuals have little or no WMSA at all. Advances in the routine measurement of white matter damage may include, alternatively, examining the amount of abnormal tissue relative to total white matter (a metric of the relative damage to white matter overall) and quantifying the degree and quality of tissue abnormality (utilizing microstructural characterization procedures). Additionally, a more detailed regional localization of the white matter abnormalities based on gyral or fiber anatomy would increase the potential to understand the consequences of this damage. Previous work has examined the differential impact of WMSA within specific lobar regions, and has contrasted changes in the periventricular region (bordering the ventricles) to those in the deep white matter (distal from the ventricles). These different types of WMSA that differ in their etiologies, and the cognitive consequences of WMSA are demonstrated to be regionally dependent (Brickman et al., 2009; Bunce et al., 2010). Within both normal and abnormal appearing white matter, there is a range of tissue integrity at a microstructural level, measurable by quantitative MRI techniques including DTI and MTI. Indeed, microstructural studies have demonstrated substantial heterogeneity across WMSA (Spilt et al., 2006). With regard to regional effects, examination of the integrity of U-fibers has not been readily possible due to technical limitations. However, it is possible that some combination of advanced DTI methods (Oishi et al., 2011) and procedures for surface sampling of microstructural properties (Salat et al., 2009b; 2011a) may allow for a needed analysis of this short-range fiber system.
Regional Notions
Understanding the regional nature of age-associated brain changes has been an important goal of studies to date, as this information could provide clues about the pathologic origins of white matter damage. Trends have been noted in the regional profiles of white matter damage. Among these, three ideas have been advanced and are generally supported by existing research. First, it seems that frontal white matter is, to some degree, more vulnerable to damage than the remainder of the brain. This idea is based on the fact that frontal white matter shows effects of age across a wide range of studies, and the frontal white matter may show statistical effects in a greater proportion of tissue of the lobe overall compared with other areas of the brain (Pfefferbaum et al., 2005; Salat et al., 2005a), and may be supported by a mechanism of smaller diameter fibers being vulnerable to degeneration, at least for commissural fibers (Tang et al., 1997). Contrary to this idea, however, is the fact that there is considerable heterogeneity within the frontal white matter with certain areas showing limited microstructural associations with age (Salat et al., 2005a, 2005b). It is possible that the frontal vulnerability is more centralized to the frontal periventricular regions (Fig. 5). Thus, frontal disconnection alone is not sufficient to describe the effects of age on neural connectivity and cognition. Second, an anterior to posterior gradient of degeneration has been suggested, and this idea has support from several studies (Bennett et al., 2010; Head et al., 2004; Pfefferbaum et al., 2005; Salat et al., 2005a), at least for certain fiber systems [e.g., commissural fibers (Sullivan et al., 2010a, 2010b) or fronto-parietal fibers (Davis et al., 2009)]. Evidence against a simple anterior-posterior gradient comes from the fact that certain posterior regions demonstrate a stronger decline in white matter microstructure than even frontal regions, and, thus, a gradient of this sort would likely be with regard to specific vulnerable fiber systems. It will be important to describe whole-brain patterns of tissue change to determine whether the gradient effect is due to a selective deterioration of fiber systems, or alternatively, it will also be key to determine whether this effect is due to relatively nonspecific damage, for example, in the frontal periventricular white matter. Third, a retrogenesis model (Reisberg et al., 1999) of age-associated white matter degeneration has been proposed, where the developmentally late developing regions of the brain are the most vulnerable to degeneration with aging and/or AD. Similarly, it is suggested that association regions of the brain may be more vulnerable than primary cortical regions. There is some limited support for such patterns in white matter deterioration in nondemented aging with white matter in certain primary regions such as pericalcarine being preserved to microstructural loss relative to frontal regions (Salat et al., 2005a). Similarly, although showing very different patterns of degeneration, there is some limited support for retrogenesis in AD (Brickman et al., 2011; Stricker et al., 2009). It may be the case that, although primary systems are somewhat resistant to age-associated decline in structural connectivity, association, or later myelinating systems are selectively vulnerable depending on the condition examined. In non-demented aging, there is a preferential loss of commissural and fronto-parietal fibers with an anterior-posterior gradient. Similarly, there is a degeneration of periventricular white matter with a preponderance of damage to frontal and parietal white matter (Fig. 5). In contrast, the effects of AD on white matter may somewhat mirror the well-defined cortical pathology of the disease (Braak and Braak, 1991). Specifically, several studies have noted alterations in white matter microstructure in regions including the limbic, parahippocampal, posterior cingulate, and precuneus and parietal white matter (Salat et al., 2010; Zhang et al., 2007). Thus, AD may be characterized by more limbic and parietal microstructural deterioration, and by WMSA formation with a prominence in parietal tissue (Brickman et al., 2009). Much of the work describing regional patterns to date has been qualitative in nature, and therefore, more formal testing of spatial patterns will be a welcome addition to the literature. Such comparisons can be difficult given the various available contrasts with neuroimaging. Although developmental models for the study of retrogenesis are typically considered with regard to what might be expected from Flechsig's myelogenetic maps from 1901 (Flechsig, 1901), the use of imaging techniques to map development suggests that patterns of growth can differ depending on the metric used for assessment (Lebel and Beaulieu, 2011). Similar differential effects are found when using different diffusion contrasts for measuring degenerative change. The challenge, therefore, becomes one of understanding the potential pathophysiologic basis of the different measurements obtained by MRI and attempting to link specific neural and physiologic properties to the systems in maximum decline.

Ranking of the association between age and white matter microstructure. The image demonstrates results from a voxel-based regression of age on diffusion tensor imaging measures of fractional anisotropy (FA) across the adult age span. Statistical values for each voxel were divided into five equal bins, ranked by the magnitude of the statistical effect and color coded, to determine whether age-related effects showed patterns suggestive of current conceptualizations of age-associated decline. Although there was a strong association between age and FA in frontal white matter (highest bin color; charcoal), these effects were only found in selected portions of total frontal region, and other frontal areas were relatively spared. FA in anterior regions showed substantial associations with age; however, the strongest class of effects extended to parietal and anterior medial occipital white matter as well. These data provide a rudimentary example of potential methods for modeling regional effects.
Pathophysiology
Understanding the biological environment promoting white matter degeneration may provide a better comprehension of key mechanisms of neural alterations and cognitive decline with aging, and may hold the key to the development of novel therapeutics to ameliorate such decline (Brown and Thore, 2011). It has been known for some time that macrostructural white matter damage is associated, in a somewhat complex manner, with cerebrovascular health and risk [see substantial relevant work by the Rotterdam Study (
These findings collectively suggest that simple inter-individual variation in vascular health may have an important influence on the integrity of the connective anatomy of the brain in the general population. How more extreme vascular insult contributes to overt lesions and the development of dementia is still open to debate. Given the significant increase in WMSA in individuals with AD and compelling evidence from epidemiological and other data, it is reasonable that there is speculation that vascular dysfunction is a primary component of the pathophysiology of AD (de la Torre, 2002). Similarly, given the potential influence of ischemia on myelin, a vascular etiology would also support the idea that dementia begins a homeostatic cascade to this degeneration (Bartzokis, 2011). Alternatively, it is possible that vascular insult promotes tissue degeneration that is consistent with a non-demented aging process, but those individuals who have both types of damage have a more rapid conversion rate due to a simple compounding of effects. Finally, it is possible that there are interactions between independent AD-associated and vascular-associated insult that produces a syndrome which is more severe than the simple combination of the two conditions (Buckner, 2004; Burns et al., 2005) (Fig. 6).

Models of vascular influences on brain aging. Accumulating evidence points to an important role for altered vascular health in neural and cognitive decline with aging, and there is speculation that vascular health could be a primary etiology of Alzheimer's disease (de la Torre, 2010). Given associations between measures of vascular function and cerebral white matter integrity, this idea would be in accord with the proposition of Alzheimer's disease arising from homeostatic processes associated with myelin degradation (Bartzokis, 2011). Thus, vascular health may influence the aging brain in a manner that contributes to typical, age-associated neural decline and cognitive loss. Alternatively, vascular health may initiate a series of events that result in the atypical syndrome of Alzheimer's disease. Neuroimaging can be an important tool in testing such theories by linking epidemiologic information with measures of systemic physiology to examine the influence of vascular health on neural health and cognition and to determine whether patterns of neural decline with vascular risk resemble more those of typical aging (bottom right; frontal pathology and decline in executive functions and speeded processing) or those of Alzheimer's disease (top right; temporal pathology and decline in episodic memory). Tests of this type will prove to be more complex, as it is also possible that vascular health influences both systems, and/or interacts with additional ongoing degenerative processes associated with aging and Alzheimer's disease.
Connectivity and Cognition
Measures of white matter integrity in humans show variable associations with cognitive function [as reviewed in (Gunning-Dixon et al., 2009; Madden et al., 2011)]; however, consistent associations with performance on tests of executive function and speeded processing support these cognitive domains as being preferentially affected by white matter deterioration. A similar finding of associations between diffusion measures and executive function with aging is additionally found in nonhuman primates (Makris et al., 2007). Of the diffusion microstructural measures, associations between cognition and DTI measures are dominated by variation in radial diffusivity [e.g., (Jacobs et al., 2011)]. The association between white matter integrity and cognition may be more complex, however, as diffusion measures also show regionally differentiated associations with cognitive control and episodic memory (Ziegler et al., 2010) as well as implicit sequence learning (Bennett et al., 2011). As just noted, it has been further hypothesized that white matter deterioration plays a primary role in the etiology of AD through the disruption of neural transmission and disconnection in the form of altered temporal synchrony. Indeed, previous studies have suggested that risk for conversion from mild cognitive impairment to dementia is greater in individuals with white matter damage and that regional measures of damage provide information about degree of impairment; however, it is much less clear whether WMSA contribute to a specific aspect of the cognitive decay accompanying AD. It might be expected that information about connective integrity would have a greater prognostic significance for predicting conversion to AD given the fact that the finding of increased WMSA volume in clinically diagnosed AD is consistently reported (Carmichael et al., 2010; Salat et al., 2010), that there is a regional decrease in white matter volume in patients with AD (Salat et al., 2009a), and that there is a decrease in the integrity of specific regions of white matter in this neurodegenerative disorder (Zhang et al., 2007), which is, in part, independent of hippocampal atrophy (Salat et al., 2010). It is likely that more advanced procedures for the measurement of the connective integrity of the brain will advance our understanding of the role of white matter damage in the clinical profile of individuals with AD. For example, Wen and colleagues (2011) recently employed innovative DTI tractography procedures and graph theory-based analyses in a large sample of older adults and related connective efficiency to cognitive domains including processing speed, memory, language, visuospatial, and executive functions. The investigators found that whole-brain cortical connectivity was associated with processing speed and visuospatial and executive functions, whereas specific regional connectivity in superior frontal gyrus and posterior cingulate cortex was more selectively associated with executive function. This study exemplifies the shift to connectomics as opposed to the region of interest or even whole-brain voxel/tract-based studies, and such analyses will likely be fruitful for understanding brain function and dysfunction in future work, and for a more comprehensive understanding of how degeneration in the connective matrix may contribute to dementia.
From Structure to Function
Previous work has attempted to link structural connective integrity to functional activity in the study of aging through examining associations between paradigm-based fMRI and diffusion tensor or WMSA measures (Chen et al., 2009; Davis et al., 2011; Gold et al., 2010; Hedden et al., 2011; Li et al., 2009; Madden et al., 2007, 2010; Nordahl et al., 2006; Persson et al., 2006). Not surprisingly, this body of work generally demonstrates that the functional integrity of the brain is, in part, mediated by the structural integrity of the white matter architecture. Similar approaches have only recently been applied to the study of network activity with functional connectivity procedures (e.g., seed-based cross-correlation and independent component analysis based studies). A review of the literature combining structural and functional connectivity was presented by Damoiseaux and Greicius (2009). With regard to aging, white matter deterioration measured by DTI is associated with decline in network activity in the default mode network in older adults (Andrews-Hanna et al., 2007). Specifically, white matter microstructure in the centrum semiovale was associated with a diminished coherence between posterior cingulate and medial frontal fMRI activity, thus suggesting that generally deteriorated structural connectivity promotes abnormal network function. Madden and colleagues found that functional connectivity of specific task-associated regions was greater in younger compared with older adults during certain portions of an executive control task (cue processing) and not others (target processing), and that this effect was not mediated by white matter integrity in task-related regions (Madden et al., 2010). The authors concluded that white matter integrity and functional connectivity may have independent influences on cognitive performance in this context. The integrity of the corpus callosum, which deteriorates with aging, is associated with functional connectivity parameters (Chen et al., 2009) and potentially supports interhemispheric connectivity between contralateral brain regions, thereby contributing to a task performance gain (Davis et al., 2011). These studies highlight the promise of bridging the structural and functional domains in the study of aging. Other findings linking structural and functional connectivity include correlations between executive function and fiber bundles connecting the putamen to the dorsal attention network (Ystad et al., 2011), and white matter deterioration is associated with a decline in functional connectivity in the posterior cingulate cortex and hippocampus in early AD (Zhou et al., 2008). Thus, the modest number of studies to date combining measures of structural and functional connectivity in aging suggests that the multimodal integration of techniques to optimally assess these two domains of connectivity will be of significant value for understanding aging of the brain. The specificity of the reported structure function associations is still unclear. It will be important to distinguish between the effects of deterioration of a precise fiber system versus a more generalized effect of attenuated neural health in future work employing comprehensive analysis procedures. In particular, it is of interest to determine whether specific systems are most vulnerable to decline and provide clues as to the biological basis of any susceptibility.
Caveats and Recommendations
Several challenges will need to be overcome to increase our ability to understand how deteriorating structural connectivity affects functional connectivity and cognition with aging. Given the prominence of DTI in studies of structural connectivity, it is noteworthy that previous work has described the potential corruption of diffusion data by partial volume contamination and by suboptimal procedures for spatial normalization (Smith et al., 2006). These issues remain challenges in current diffusion work. Such limitations will be partially overcome by high-resolution techniques and by careful procedures for spatial normalization (Smith et al., 2006). Although challenging for in vivo studies, high-resolution DTI may be an important tool in the postmortem analysis of the aging brain (McNab et al., 2009). In addition to necessary advances in acquisition and analysis procedures, interpretation of changes in white matter properties measured by MRI remains challenging. Atrophy-associated changes in tissue composition per unit, such as reported increase in capillary density, could have an important effect on tissue microstructural measurements and may confound typical interpretations as microstructural changes being due to myelin degeneration or loss of myelinated nerve fibers. Additionally, it should be considered that functional connectivity is typically measured based on the hemodynamic response using fcMRI procedures, and there is a known effect of aging on vascular function which influences signal measured by fMRI (Kannurpatti et al., 2011). This consideration is compounded by the fact that vascular health may be the primary factor determining structural connective integrity. It will, therefore, be important to distinguish the co-occurring effects of vascular alterations on structural connectivity and on vascular reactivity from those due to effects of changes in structural connectivity on neural metabolism and physiology. Similarly, it is unclear whether alterations in conduction velocity with damage to myelin as a result of aging, measurable by neuroimaging, would be expected to have a detectable effect on signal co-variation at the timescale of the hemodynamic response; however, it can be speculated that accumulating damage to a collection of fibers could have a cumulative effect on this measure. It is significant to note that since there is such support for vascular pathophysiologic origins to white matter damage, this may be expected to influence the white matter based on the anatomy of the vascular tree, and not with a tract-based specificity per se, as has been the focus of much work. In fact, associations between blood pressure and white matter integrity may be somewhat widespread, intersecting several major fascicles, yet not particularly selective to the anatomy of any particular fiber bundle (Salat et al., 2011b). Thus, it will be important to explore a range of potential mechanisms and patterns by which changes in structural connectivity may influence functional connectivity and cognition. Specific details about the temporal expectations (within the age-span), regional expectations, and the types of degenerative changes expected will be needed to augment current models of brain aging, and formal statistical comparison across regions will be necessary to demonstrate selective or preferential spatial effects. It will be important that interpretations of regional changes consider the potential influence of differential sensitivity and reliability of the particular measure utilized, which will be influenced by specifics of the biology as well as the data acquisition and analysis procedures. For example, certain regions showing substantial age-associated change, such as medial frontal white matter, are also those affected by MRI susceptibility artifacts (Yang et al., 1997). Thus, a range of properties about the data acquisition and analysis should be quantified and considered in any formalized theoretical regional tests.
Future Directions
There are several areas that would benefit from continuing development of neuroimaging procedures in the study of white matter and connective integrity of the brain. Three immediate domains of focus would be (1) enhancing anatomical scale of analysis; (2) determining the degree of abnormality; and (3) determining the category of abnormality. With regard to anatomical scale, increased resolution acquisitions will be essential for controlling for the continuing influence of partial volume contamination. Additionally, a finer detail of the locational prevalence of lesions with regard to cortical and tract anatomy will be important for understanding the clinical consequences of such damage. Continuing development of automated tractography procedures and with high angular resolution diffusion imaging techniques will be important toward better defining the full anatomy of specific age and disease effects on neural systems; whereas new procedures for voxel-based analysis of WMSA (Smith et al., 2011) using novel anatomic matching procedures for inter-subject spatial normalization would provide the exploratory statistical framework for understanding the true anatomical nature of age-associated white matter deterioration. Recent studies have demonstrated substantial progress in mapping the anatomy of U-fibers (Oishi et al., 2011), and surface-based procedures for sampling white matter integrity subjacent to the gray/white border may provide important information about the integrity of these superficial fibers (Salat et al., 2009b, 2011a). Additionally, an important structural parameter contributing to neural conduction is axon diameter, and this property is potentially measured through specialized diffusion acquisitions (Assaf et al., 2008). With regard to degree of abnormality, there are now several quantitative microstructural procedures that can be utilized in conjunction with more classical WMSA segmentation procedures to explore the domain of lesion severity. Examination of the degree of signal change across a range of available quantitative MRI sequences, including (e.g., DTI, MTI, and MRS) for the assessment of white matter, may contribute to these goals. Defining the specific type of abnormality is potentially the most ambitious of current goals. Although little work has examined the potential for automated classification of lesion types, it is possible that information in available MRI tissue contrast parameters provide information about the pathologic specificity of changes measured. Several studies have interpreted findings from diffusion imaging based on results in animal models, and pathologic correlates of imaging data have been explored in human aging and disease (Chen et al., 2011a; Klawiter et al., 2011). Alternate factors should be considered as potential confounds to specific interpretations of MRI signals; however, with a growing spectrum of acquisition procedures with unique contrast [e.g., (Falangola et al., 2008)], there is reason for optimism that multispectral signal profiles may provide the information necessary to distinguish pathologies at this scale, and future research will be highly informative in this domain. Such studies would greatly benefit from longitudinal design. Much of the work reviewed here is based on cross-sectional observational research, and, therefore, only describe associated conditions and cannot imply any direct influence. Several studies have examined the longitudinal change in WMSA volume. Individuals with greater amounts of WMSA at baseline (Gouw et al., 2008) and greater vascular risk (Raz et al., 2007) are most likely to show progression in WMSA over time. In contrast, relatively few studies have examined the progressive decline in DTI measures such as FA with advancing age (Barrick et al., 2010; Sullivan et al., 2010a). Findings from this work are conflicting with suggestions of nonspecific, or minimal progressive decline in white matter microstructure within approximately 2 years (Barrick et al., 2010; Sullivan et al., 2010a). Future work will clarify the full spatiotemporal and longitudinal patterns of microstructural change in the aging brain. There are, therefore, several promising lines of investigation to be explored to advance understanding of the influence of age-associated structural decline on the functional capacity of older adults.
Final Considerations
Advances in technologies for mapping the structural connective anatomy and integrity of integrated neural systems will provide great opportunity for understanding brain network function, and the underpinnings of cognition and behavior. Individual differences in the connective infrastructure have an important role in variation in neural processing and cognition in the general population. Although there is degradation in the construction necessary for optimal neural function in aging, several hopeful considerations accompany this. First, it is of note that in the general population, a number of older adults do not have appreciable amounts of WMSA, thereby demonstrating that accumulation of this type of damage is not a necessary consequence of aging. Additionally, there is a wide range in the function of older adults with individuals often performing similar to their younger counterparts on cognitive testing. It is important that several factors associated with white matter degeneration, such as vascular risk, can be significantly modified by lifestyle and dietary alterations. In the healthy brain, there is a normal cycle of dynamic myelin turnover (Ando et al., 2003; Franklin and Ffrench-Constant, 2008), and there is, notably, a continued yet attenuated remyelination of damaged nerve fibers with aging (Bowley et al., 2010). Interventional studies to prevent age-associated white matter degradation are of pronounced importance, and data from macaques suggest that dietary modification and caloric restriction can attenuate white matter degeneration with aging, at least in select regions of the brain (Bendlin et al., 2011). Thus, strategies to restore this cerebral infrastructure may be fundamental toward the development of therapeutics for the amelioration of age-associated white matter degeneration, deficits in functional connectivity, and the enhancement of cognitive function in older adults.
Footnotes
Acknowledgments
This work was supported by the National Institutes of Health and National Institute of Nursing Research R010827. The authors gratefully acknowledges Jean Augustinack and Betsy Leritz for providing guidance with this article; Jean-Philippe Coutu, Juli Shepel, and Doug Greve for assistance with data presented; and the peer reviewers of this article for their constructive feedback.
Author Disclosure Statement
No competing financial interests exist.