
Abstract
One of the greatest challenges involved in studying the brain mechanisms of fear is capturing the individual's unique instantaneous experience. Brain imaging studies to date commonly sacrifice valuable information regarding the individual real-time conscious experience, especially when focusing on elucidating the amygdala's activity. Here, we assumed that by using a minimally intrusive cue along with applying a robust clustering approach to probe the amygdala, it would be possible to rate fear in real time and to derive the related network of activation. During functional magnetic resonance imaging scanning, healthy volunteers viewed two excerpts from horror movies and were periodically auditory cued to rate their instantaneous experience of “I'm scared.” Using graph theory and community mathematical concepts, data-driven clustering of the fear-related functional cliques in the amygdala was performed guided by the individually marked periods of heightened fear. Individually tailored functions derived from these amygdala activation cliques were subsequently applied as general linear model predictors to a whole-brain analysis to reveal the correlated networks. Our results suggest that by using a localized robust clustering approach, it is possible to probe activation in the right dorsal amygdala that is directly related to individual real-time emotional experience. Moreover, this fear-evoked amygdala revealed two opposing networks of co-activation and co-deactivation, which correspond to vigilance and rest-related circuits, respectively.
Introduction
O
Several imaging studies aimed at determining whether real-time emotional rating would affect limbic/paralimbic neural activation. For example, (Hariri et al., 2000) showed that when processing emotional faces, the act of labeling the emotional expression reduced activation of the amygdala, compared with visually matching facial expressions. Furthermore, (Taylor et al., 2003) demonstrated that real-time emotional appraisal of pictures was related to less activation of the insula and amygdala compared with passive viewing of the same pictures. In contrast, however, when (Hutcherson et al., 2005) used a method involving a continuous self-reporting dial and asked subjects to rate their emotional experience while watching a movie, it decreased neither the self-reported experience of emotion nor the amount of neural activations compared with passive viewing. This suggests that choosing a rating method with minimally intrusive characteristics may not interfere with ongoing emotional responses.
Another difficulty involved in studying the neural correlates of the authentic aspect of emotional experience is the need to capture its emerging nature. Although static emotional stimuli that are prefixed in time of onset and duration are well controlled, they are hampered by the lack of dynamics. Although presenting an experimental challenge due to their noncontrolled nature (Gross and Levenson, 1995), movies more closely resemble real-life scenarios; thus, free viewing of movies has received increased attention in neuroimaging studies on emotions (Bartels and Zeki, 2004; Hutcherson et al., 2005), mostly along a data-driven analysis approach (Goldin et al., 2005; Hasson et al., 2004). These studies showed that functional brain specialization is preserved even with a naturally occurring time course produced by the free viewing of movies. This cross-correlation approach is, however, based on an a priori assumption of the sources of information outside the individual's brain. A data-driven analysis approach, on the other hand, based on clustering algorithms of each individual's brain responses, precludes such previous assumptions. This latter approach has been applied to functional magnetic resonance imaging (fMRI) fine voxel analysis for obtaining distinct brain activity states, which can later be used as regressors for external/behavior state inference (Dodel et al., 2002; Eguiluz et al., 2005; Jacob et al., 2010; Sporns et al., 2000, 2007; Stam and Reijneveld, 2007).
The prevailing assumption of a data-driven analysis approach in neuroimaging is that neurons in close vicinity tend to be co-activated in times of need for rapid generation and transfer of information, thus forming functional cliques (Dodel et al., 2002; Eguiluz et al., 2005; Stam and Reijneveld, 2007). Notably, clustering based on functional cliques has so far been applied in neuroimaging studies involving mostly motor areas where the activity is anatomically well confined. The aim of the current fMRI study was to develop a robust data-driven analysis that is localized to the amygdala and is guided by individual reports on the emergence of fear experience. We chose the amygdala for localized focused clustering, as it is a key structure in processing the fear response in animals and humans (Bishop et al., 2004; LeDoux, 2000; Murphy et al., 2003). Ongoing changes in subjectively reported experiences of fear were measured while individuals were viewing two different horror movies, and they were used to define periods of high conscious fear (HCF). It was assumed that the range of affect during movie viewing can vary considerably among individuals, and that the specific time course of emotional appraisal during that activity would reflect the authentic real-time subjective feeling toward the viewed material. A semi-supervised clustering approach was then applied to precisely localize the brain activation related to these HCF periods in each participant. It is assumed that the unified unfolded cluster is constructed under the condition and/or for eliciting a fear reaction.
We expected that the individualized data-driven clustering would reveal areas of activation within the amygdala comprising a few voxels that might not be detected by the common paradigm-driven analysis, especially under the condition of performing real-time emotional appraisal. If so, then it will enable capturing activated parts of the amygdala that are directly related to ongoing modulations in an authentic experience of the feeling of “I'm scared.”
Materials and Methods
Participants
Twenty healthy individuals participated in the behavioral experiment (right-handed, age 36±12, 10 women). Twenty other healthy individuals participated in the fMRI experiment, of which the data of 15 (right-handed, age 31±6, 8 women) were fully analyzed due to excessive head movement in the other five subjects. The enrollees had no history of neurological or psychiatric illness. All participants had normal or corrected-to-normal vision and provided written informed consent to participate in this study that had been approved by the Tel Aviv Sourasky Medical Center Ethics Committee.
Behavioral experiment
Ten video clips from 10 commercial, well-known scary movies, including “The Saw,” “Halloween,” “Mystic River,” “Audition,” and “The Birds,” were chosen for their fear-inducing reputation. We chose 5 min movie clips that gradually induce a high level of fear within 5 min and maintain the fear feeling throughout the majority of the clip. The participants were instructed to rate the experience that evoked the conscious feeling of “I'm scared.” Participants watched 2–4 5-min movie clips (each clip was watched six to seven times overall in a counter-balanced manner) on a computer screen and were asked to mark their experiencing of fear in real time by pressing a button. A soft beeping sound marked the grading intervals. The 20 sec time interval between reports followed the repetition time (TR) timing. Participants were instructed to avoid planning their next rating but rather to instantaneously rate their fear when they heard the beeping sound as follows: 1—“I'm not scared,” 2—“I'm a little scared,” 3—“I'm very scared.” The three-point scale rating was chosen in order to limit the degree of contemplation over the rating level. After the movies ended, the participants were asked to complete a 16-item self-report emotion inventory adapted from a procedure described by Gross and Levenson (1995) for identifying a movie's discrete emotional category.
Preliminary evaluation of 10 movie clips revealed that two movie clips, “The Birds” and “Halloween” elicited the highest scores of fear (“The Birds” average of 6.0±2.04, “Halloween” average of 6.1±0.7) as well as the lowest scores for other emotions. The participants' real-time self-reporting of fear showed that these two movie clips also elicited more often instantaneous intense fear in comparison to the other clips.
fMRI experiment
Stimulation
While lying in the scanner, participants viewed 5 min of each selected scary movie. At the beginning and end of the clip, the participants viewed 1 min of blank gray screen with a fixation point. As for the behavioral experiment procedure, the participants were instructed to rate their fear on a scale of 1–3 by pressing a key following a soft beeping sound that cued the response time every 20 sec.
fMRI acquisition
Brain scanning was done by a 3T GE scanner with an eight-channel head coil. fMRI was performed with a gradient echo-planar imaging sequence of functional T2*-weighted images (TR/time to echo [TE]/flip angle: 3000/35/90; FOV: 20×20 cm2; matrix size: 128×128) divided into 39 axial slices (thickness: 3 mm; gap: 0 mm) covering the whole cerebrum. Anatomical three-dimensional (3D) sequence spoiled gradient echo sequences were obtained with a high-resolution 1-mm slice thickness (FOV: 250 mm; matrix: 256×256; TR/TE: 6.7/1.9 msec).
Image analysis
Imaging data were preprocessed and analyzed using BrainVoyager QX software package version 1.7 (Brain Innovation, Maastricht, The Netherlands) and homemade add-on computational tools for advanced clustering analysis. Raw functional images were superimposed and incorporated into the 3D data sets through trilinear interpolation. The complete data set was transformed into Talairach space (Talairach and Tournoux, 1988). Preprocessing of the functional scans included head movement assessment (scans with head movement >2 mm were rejected) and a temporal smoothing process that included linear trend removal and usage of a high-pass filter of 0.005 Hz. To resolve a distinction of the sub-regions of the amygdala, the neuroimaging data were acquired at a relatively high spatial resolution (1.5×1.5×3 mm voxels) and then resampled to 2×2×2 mm voxels. The first eight volumes of the scan were discarded to allow for T1* equilibration effects. The 3D group statistical parametric maps were calculated using a general linear model (GLM) analysis.
Individually tailored functional clustering
To demonstrate the relation between the fear experience and cliques of activation in the amygdale, we ran a series of analyses following the outlined rationale (see Fig. 1; method flow chart, A–E): first, we looked for the link between the periods of reported HCF during film viewing and a voxel's number of connections (i.e., degree) in the amygdala, showing that higher rating corresponded with a higher degree. Then, we applied a data-driven approach in order to reveal the time course of changes in degree along the movie in each amygdala's voxel, thus marking the time point with the greatest degree, which also reflects the highest fear experience. To find the functional clique within the amygdala that is related to the highest fear experience, we applied cross-correlation and graph theory techniques. We subsequently examined each clique using a distance-based analysis, to find the most unified fear-related cluster within the amygdala.

Method flowchart. The chart is divided into two parts. The first part describes the steps for the construction of the amygdala conscious-fear-related functional cliques, and the second part describes the steps toward revealing the amygdala conscious-fear-related brain networks. Blue pointers refer to the relevant figures.
The following are detailed descriptions of each step in the clustering procedure: (1) Linking between a voxel's number of connections (voxel's max degree) within the amygdala and individual real-time fear experience. This procedure correlates between participants' real-time self-reporting of no conscious fear (NCF) or HCF and the max degree values. The interval depicting the NCF was determined by repeated reports of “I'm not scared” marked as 1. The other interval of interest depicted the periods of “I'm very scared” (HCF), marked as repeated reports of 3 (Fig. 2). Repeated reports of 2 were used for subjects who did not report any 3 ratings. The NCF and HCF experience time intervals were confined to 7 TR (21 sec), which were determined for each participant based on the real-time ratings of the movie during scanning. The time window was defined as including the minimum range of reported fear (20 sec). For subjects with a longer range of reported fear (>20 sec), the middle of the time range was arbitrarily selected. The calculation of max voxel degree was performed on a bilateral amygdala that was manually segmented for each participant, defining the region of interest (ROI). The extracted functional data were unsmoothed and had high voxel resolution to ensure that the signal originated from the identified ROIs. A cross-correlation analysis was conducted between all the voxels' signal time course in the ROI restricted to the two experience-based time intervals (i.e., HCF and NCF). Both intervals were shifted (Δt=6 sec) to correct for hemodynamic delay in the fMRI responses. We applied a threshold of p<0.001 to the two resulting correlation matrices, converting them into binary matrices (i.e., if the p value of the correlation between two voxels was >0.001, the correlation value was set to 0; otherwise, it was set to 1). We calculated the sum of each voxel's connections (i.e., degree) individually and used a criterion for minimum connections (i.e., fewer than four voxel connections were considered as 0). We chose this value presuming that 1–3 connections can be created due to spatial proximity. The voxel with the max number of connections (i.e., max voxel degree) was identified, and this number was recorded (Fig. 3). (2) Tracing the data-driven continuous measuring of a voxel's max degree to exhibit the relation between changes in the number of voxel's degree in the amygdala and the responder's HCF modulations during move viewing. This procedure calculates the changing max degree (i.e., the max connections of one voxel per time interval) using the time course of an ROI in the amygdala throughout the two movies for each participant. Consequently, here, we did not restrict ourselves to the participants' reporting but instead adopted a full data-driven approach. A sliding window technique was applied to calculate the changing max degree of the voxels in each participant's extracted two amygdalas. We followed the same procedure as described in step number 1. With a window step of 2 TRs and a length of 7 TRs that is about the interval between two points of self-rating, we ran a cross-correlation, filtered with a threshold of p<0.001, and found the voxel with the max number of connections. This number was recorded for every 7 TR steps throughout the whole time course for each participant (Fig. 4A). For subjects who produced more than half of the largest max degree (i.e., 25 connections), we increased the window of degree calculations up to 9 TR's for reducing overlapping clusters (Newman, 2006). We did not increase the number of TR's above this level in case the next report interfered with the current report. (3) Applying cross-correlation and functional clustering to find the individual's HCF-related activation in the amygdala. This was done in the following way: First, for each participant, we defined a unique time window corresponding to the period representing the highest fear experience (taking into account the hemodynamic delay in the fMRI responses, Δt=6 sec) and eliciting the highest degree throughout the run (as calculated in step 2). Following the same procedure as described in step number 2, this time window lasted for 21–27 sec across participants (i.e., 7–9 TRs). Then, for each participant, we executed Pearson-based cross-correlations between all pair-wise combinations of all the voxels extracted from the bilateral amygdala's signal time courses (see Fig. 5A). Following Newman (2006), we used p<0.001 as our criteria threshold in order to convert the data to a binary adjacency correlation matrix. The strict threshold enabled focusing on the “core” voxels that respond to fear in the amygdala and also reducing the overlapping clusters. Next, we computationally applied graph theory concepts and community mathematical ideas in order to compose a functional clique in the amygdala in which every member is fully connected to the other members (implemented in MATLAB; Mathworks, South Natick, MA). In order to find the connected components of five to seven voxels, we calculated all possible five to seven voxels combinations and checked, for each combination, whether all the voxels were connected in the adjacency matrix. This process was repeated for several different thresholds. The connected component that was found using the highest threshold was chosen as the “core” functional cluster which corresponds with the HCF period. (4) Forming distance-based clustering to assemble the voxels as a joint nucleus. The cluster of voxels produced by the functional connectivity-based method is not restricted by location; thus, the correlated voxels may be spread around the amygdala and do not necessarily form one area. We further applied Euclidean distance <2 restriction between all members of the cluster, excluding the voxels that were spatially far from the neighboring group, forming a united HCF ROI.
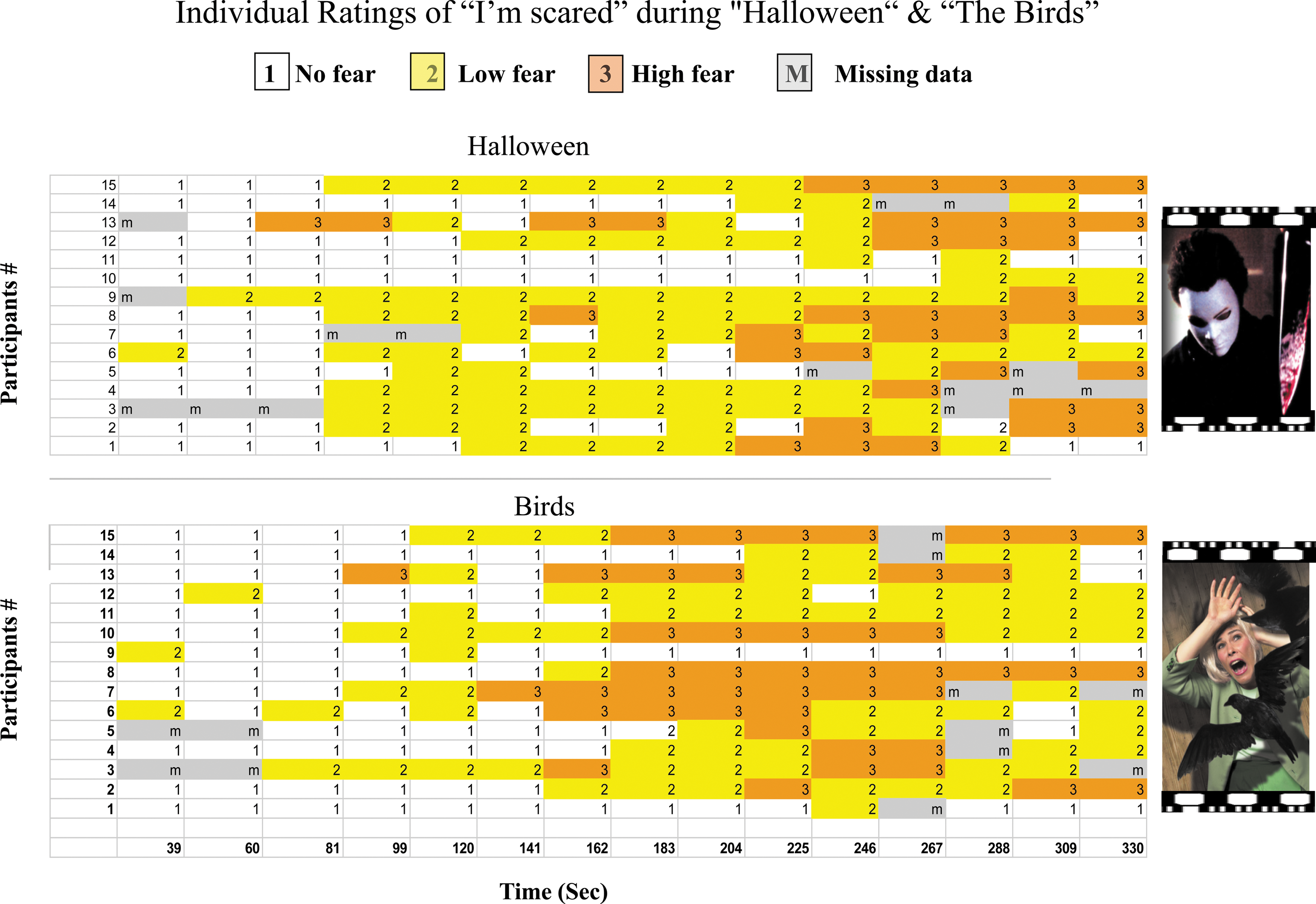
Summary of real-time emotional ratings of the “I'm scared” level obtained from 15 participants during brain scanning while viewing 5 min movie clips of “The Birds”and “Halloween.” The distribution of individual rating intensity (1–3) throughout the movie clips is illustrated by periods of different colors (white, yellow, and orange respectively).



Whole-Brain GLM-Based Mapping
Whole-brain GLM was calculated on the fMRI data obtained from each participant while viewing scary movies. The averaged HCF amygdala nuclei activity was used as a seed region. Rather than relying solely on the standard hemodynamic response function (HRF) GLM, we adopted a more individually tailored GLM method that considered the individual's emotional progression (Fig. 5C, D). The commonly used GLM model (Friston et al., 2000) consists of a boxcar function convolved with the hemodynamic response function for each condition. Previous studies attempted to optimize the GLM by using several different base functions, which may be more appropriate for certain types of experimental designs, for example, half sine, discrete cosine, gamma, modified gamma, Gaussian function, and so on. We introduced a novel design of GLM base function, which may improve the validity of the GLM in terms of subjective experience and the individual ROI signals. A comparison between the new individual base GLM predictor and the standard HRF for similar subjects is described next. (see Fig. 1; method flow chart, F, G)
The analysis included the following two steps: first, calculating an individually tailored base function (ITBF) from the bilateral amygdala's time course. The GLM seed-region analysis is based on subject-specific time courses (confounded here to times of high fear) that were individually used as explanatory variables. For each movie, we defined two HCF intervals of 7–9 TRs around the two highest-degree time points for each participant's signal time course obtained from the bilateral amygdala (Birds movie—74% of the second-degree peaks overlapped with fear reports; Halloween movie—80% of the second-degree peaks overlapped with fear reports).We extracted the shape of the signal time course (averaged across the HCF-based amygdala clusters) and used this as a predictive function instead of the HRF in a GLM calculation. Both the onset function time and its temporal shape were different for each participant and film (see example Fig. 5C, D). Second, performing a multi-subject GLM using the ITBF or the HRF as predictors. The HRF base function was applied as a predictor using the same time intervals defined for the individual subject in the ITBF procedure. For each participant, the onset of the predictor was different, but the shape was the same (i.e., HRF function). Using the GLM HRF with the same time intervals as the ITBF function enabled a comparison of the impact of the individual function while the emotional experience remained the same. For each predictor, we also performed a multi-subject whole-brain analysis using the Brainvoyager GLM package. This process includes the individual Talairach files with the individual predictors functioning as just described from the two movies. The negative function for both base function styles was the basis for the amygdala whole-brain deactivation network.
Results
Real-time emotional rating
Figure 2 shows the time course of cued emotional rating ranging from 1 to 3 for each participant (see methods). It is evident that although several time intervals showed similar fear level reports among participants, there was inter-individual variability as well. Importantly, for the validity of experiencing fear, all participants repeatedly rated the level of their feeling as 2 or 3 at various points during the viewing of the horror film clips. Discrete time intervals derived from individuals' emotional intervals of HCF and NCF were defined for each participant and for each movie. Figure 3B shows the time course of one participant's rating during the viewing of a clip from “The Birds.” The max voxel degree of both amygdalas was then calculated separately for the HCF and the NCF periods for each participant (see methods).
HCF periods and max voxel degree of the amygdala
Figure 3A demonstrates a clear difference in voxel max degree of the amygdala as revealed by the emotional intervals of HCF versus NCF in each movie. Specifically, while they were watching the movie “The Birds” or “Halloween,” the max degree of a voxel in the amygdala during HCF is significantly higher across subjects than the degree during NCF intervals. These results link on an individual level between a computational neural feature of the connectivity within a region and the subjective emotional experience, thus pointing to the voxel degree as a neural fingerprint for this behavior. We further investigated the amygdala's functional organization throughout the movie by using data-driven, whole-movie analysis and reverse engineering during periods of emotional experience. The analysis of the temporal change in the voxel max degree enabled a blinded reverse comparison with the participant's self-report on the experience of feeling increased fear. Figure 4 depicts the relation between the max number of voxel degree in the amygdala (Fig. 4A) and the individual's heightened emotional state of fear (Fig. 4B). It is evident that the max number of a voxel's connections (green dot) has appeared for all the participants during the period marked by each individual as HCF (orange lines). Figure 4C verifies via reverse engineering for two participants that the max degree indeed corresponded to a horror scene in the movie clip.
Networks driven by amygdala's HCF cluster
The average signal time course from the amygdala's HCF cluster was used for ITBF whole-brain GLM analysis. The high resolution of data from the amygdala (i.e., small voxel sizes, no smoothing, and no averaging across participants) enabled localizing of a functional cluster in the amygdala tagged by the individual's fear experience (two identified amygdala functional clusters for participants #13 and #15 in Fig. 5 and an illustration in Fig. 6). Table 1 shows the results of whole-brain GLM analysis using the HRF versus the ITBF as the prediction model. There was a marked difference in the statistical power between the two base function styles, pointing to a higher power for the ITBF approach, whose brain network includes the areas revealed by the HRF approach and more (see also Supplementary Data; Supplementary Data are available online at

The amygdala's functional clique networks obtained from 15 participants while viewing two movie clips. Maps depicting the amygdala's individually tailored base function (ITBF) co-activation (left views) or co-deactivation (right views) revealed by GLM group analysis. Note that the ITBF represents the time windows in which participants reported “I am scared.” (p<0.01 for demonstration purposes): The areas included (1) bilateral amygdala, (2) dorsolateral prefrontal cortex, (3) precuneus, (4) dorsomedial prefrontal cortex, (5) brainstem, (6) bilateral globus pallidus, (7) nucleus accumbens, and (8) insula. View complete details in Table 1.
Positive: p<0.05 FDR corrected, Negative: p<0.05 uncorrected.
HRF and ITBF brain network of activation. See Figure 6 for explanation. A clear advantage is evident for the ITBF approach in power and sensitivity, as shown in B.
HRF, hemodynamic response function; FDR, false discovery rate; ITBF, individually tailored base function.
Figure 6 demonstrates the amygdala-related brain co-activated and co-deactivated networks including limbic, prefrontal, and brainstem areas. (Fig. 6 and Table 1) No significant results were obtained with the negative HRF function.
Discussion
The present fMRI study investigates the amygdala-related network activation underlying the self-rated emotional experience of “I'm scared.” By using fear-inducing movie clips, we were able to examine the onset and dynamics of brain reactions to frightening moments as indicated online by the viewer. The periods in which the viewer experienced “I'm scared” were used as the reference to derive activity clusters of the amygdala. The results indicated that an individual-based approach driven by real-time emotional appraisal is a credible means for exposing amygdala activation, thus, exemplifying the ability to record the amygdala's response to fear even under the condition of real-time self-reporting. Furthermore, this individually tailored function-based approach for clustering of the amygdala enhanced the sensitivity of a second-level whole-brain functional calculation that revealed widespread co-activated and co-deactivated networks which may have remained hidden with a standard whole-brain approach (see Table 1 and Supplementary Data).
Functional clustering of the amygdala corresponding to the individual's real-time experience of “I'm scared”
To pinpoint the area within the amygdala that is most closely linked with the online emotional experience of “I'm scared,” we employed unique experimental and data analysis approaches. Experimentally, the use of fear-evoking movies along with minimally intrusive real-time cues for rating enabled the detection of authentic reporting of experiencing fear and did not alter the subjects' ability to experience fear (Fig. 2). This corresponds to previous demonstrations that real-time requests for an emotional rating did not attenuate a conscious experience of intense emotions (Hutcherson et al., 2005; Mauss et al., 2005). The real-time emotional rating guided our newly developed regional clustering approach demonstrating both (1) increased inter-voxel co-activation in a relatively small brain region such as the amygdala during distinct time points of reported high fear (Figs. 3 and 4), and (2) the fully connected functional cluster in the amygdala showed a clear phasic increase in signal amplitude during the individually determined moments of enhanced fear (see Fig. 5C, D and Supplementary Fig. S1).
Taken together, these results suggest that the activated cluster in the amygdala can be considered the neuronal representation of the online reported “I'm scared” experience.
The whole-brain group analysis derived from the individual amygdala's cliques locates the functional cluster as a relatively dorsal aspect of the right amygdala.
The dorsal area is anatomically consistent with the central nucleus of the amygdala (CeA), which is found to be an output region that densely projects to the brain stem, hypothalamic, and basal forebrain targets in rodents (Paxinos and Tork, 1990). Additionally, studies in rodents found that the central nucleus is essential for the basic species-specific defensive responses associated with the valued fear (Davis and Whalen, 2001). In humans, corresponding with our current results, the dorsal aspect of the amygdala was selectively more activated during negative stimuli compared with neutral stimuli (Davis and Whalen, 2001; Whalen et al., 2001) and more so, for conscious than unconscious stimuli (Etkin et al., 2004), in addition to being involved in modulating the attention toward salient events (Mackiewicz et al., 2006). Our work adds novel information to these accumulating insights about the dorsal amygdala, suggesting that it is also involved in the actual aware appreciation that a stimulus or an event is emotional; thus, it is relevant and may, therefore, warrant a call for action. In the setting of fear, this result corresponds to the recent fMRI finding (Mobbs et al., 2007) that the dorsal right amygdala in humans was more activated when threat was proximal than distal, and the prospect of punishment was high. Similarly, a previous work from our lab demonstrated increased dorsal amygdala activation to scary music only when the negative emotion was grounded in a real-world scenario (Eldar et al., 2007).
Amygdala-driven distributed activation corresponding to online fear experience
Whole-brain group analysis based on the ITBF GLM approach revealed what can be considered the online fear-related brain activation. Several regions showed a positive correlation with the fear cluster of the right dorsal amygdala, including the bilateral amygdala, dorsomedial and dorsolateral prefrontal cortex (PFC), periaqueductal gray matter, hippocampus, cerebellum, and fusiform gyrus, while other regions showed a negative correlation with the fear-related activation in the right dorsal amygdala, including the subgenual cingulate, lateral globus pallidus, and ventral striatum (Fig. 6 and Table 1).
The amygdala's related functional networks were previously studied by fMRI in humans, but most of these studies used the complete amygdala nucleus' signal as a seed for whole-brain correlation (Lerner et al., 2009). However, more recent imaging studies have pursued a functional connectivity approach in which this small brain structure was subdivided according to its functional selectivity (Lerner et al. 2012). Of special interest to the current work is an fMRI functional connectivity study of the amygdala during rest (Roy et al., 2009) using seed activations from ventral, dorsal, and lateral aspects of the nucleus, corresponding to the basolateral, centromedial, and superficial subdivisions, respectively. Interestingly, the dorsal amygdala activation was negatively correlated with a set of regions that correspond to our positive co-activation network, and was positively correlated to a set of regions corresponding to our negative co-activation network (see Fig. 6 and Table 1). The reverse relationship between Roy's and our findings support the idea that networks of spontaneous and task-related activity might involve similar brain areas, though exhibiting opposite states of activation. These observations are also consistent with studies that found an increase in limbic-paralimbic activation (e.g., subgenual cingulate, anterior insula) and a decrease in neocortical activation (e.g., dorsolateral PFC, inferior parietal cortex) during induced sadness in healthy participants, but an opposite pattern in depressed patients (Bewernick et al., 2010; Mayberg, 2003; Mayberg et al., 1999). To note, a normalization of this pattern was seen with recovery from depression (Mayberg, 2003). Overall, these previous findings and our results relate to a recent idea that the brain reallocates processing resources when moving from one mental state to another (Gusnard et al., 2001; McKiernan et al., 2003). Specific to the current work, during an increased emotional state of “I'm scared,” we found two opposite patterns of co-activation with the amygdala in two different sets of regions (Fig. 6). It is possible that the co-deactivation with regard to the amygdala serves as an exchange of processing resources over the immediate need for attention and action in times of evoked intense fear. As far as we know, this is the first demonstration of such a reverse profile of amygdala-driven network activation that can be directly related to the distinct emotional experience of “I'm scared.” One can speculate that this co-deactivation may act as a state-dependent response of an “amygdala-related emotional default network.” The “default mode network” usually refers to a set of regions characterized by decreased neural activity during varied goal-oriented tasks (Raichle and Snyder, 2007). Our results add to recent findings demonstrating that the default mode network is not one entity but rather can be subdivided into a set of sub-networks which deactivate indiscriminately in response to different mental demands (Mayer et al., 2010). Here, we suggest that amygdala fear-related co-deactivation in areas such as the subgenual cortex might represent the brain's resource allocation during fear reaction processing.
Conclusion
The use of free viewing of horror movies enabled the elicitation of authentic fear feelings in a laboratory setting. A robust data-driven clustering approach enhanced the sensitivity and specificity for detecting involvement of the right dorsal amygdala in this evoked fear experience, as depicted by the subject's reporting “I'm scared.” In addition, the amygdala cluster-driven whole-brain ITBF GLM analysis presented herein distributed co-activation or co-deactivation in the context of a subjective increase in negative emotional feelings. These networks closely correspond to previously demonstrated brain organization related to mood modulation. Although fMRI results do not imply excitation or inhibition of activation, they may shed light on the relationship between the amygdala and its neural partners during real-time evoked conscious fear experience, and, thus, may have implications for the neural characterization of human anxiety-related disorders and the quest for their emotional regulation.
Footnotes
Acknowledgments
This research was supported by U.S. Department of Defense award number W81XWH-11-2-0008. We thank Vicky Myers for editorial assistance and Liviu Carmely for movies selection.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.