
Abstract
Neurobiological models of obsessive–compulsive disorder (OCD) assume abnormalities in corticostriatal networks involving cingulate and orbitofrontal cortices, but the connectivity within these systems is rarely addressed in experimental imaging studies in this patient group. Using an established monetary reinforcement paradigm known to involve the cingulate cortex and the ventral striatum, the present study sought to test for altered corticostriatal coupling in OCD patients anticipating potential punishment. The anterior midcingulate cortex (aMCC), a region integrating negative emotion, pain, and cognitive control, was chosen as a seed region due to its particular relevance in OCD, representing the neurosurgical target for cingulotomy, and showing increased responses to errors in OCD patients. Results from psychophysiological interaction analyses revealed that significantly altered, inverse coupling occurs between the aMCC and the ventral striatum when OCD patients anticipate potential punishment. This abnormality links the two major contemporary neurosurgical OCD target sites, and provides direct experimental evidence of altered corticostriatal connectivity in OCD. Noteworthy, an abnormal aMCC coupling with cortical areas outside of traditional corticostriatal circuitry was identified besides the alteration in the cingulostriatal pathway. In conclusion, these findings support the importance of applying connectivity methods to study corticostriatal networks in OCD, and favor the application of effective connectivity methods to study corticostriatal abnormalities in OCD patients performing tasks that involve symptom provocation and reinforcement learning.
Introduction
N
PPI analyses compute whole-brain connectivity maps for a given seed region (Stephan, 2004) and identify areas in which the connectivity with the seed significantly changes due to an experimental factor, that is, the psychological context (Friston et al., 1997). In healthy individuals, PPIs have been applied to study amygdala–prefrontal interactions in the context of fear (Banks et al., 2007; Das et al., 2005; Williams et al., 2006), and periaqueductal gray connectivity during pain (Linnman et al., 2012) and defensive fear (Mobbs et al., 2009). Despite the simplicity of PPI, testing for task-specific connectivity changes of one single region in the absence of particular model assumptions, its applications in the context of group comparisons involving neuropsychiatric populations have been relatively rare. However, PPI has been used previously to investigate altered cingulate connectivity in post-traumatic stress disorder (Lanius et al., 2004), amygdala–prefrontal coupling in social anxiety disorder (Guyer et al., 2008), the connectivity of motor areas in traumatic brain injury (Kasahara et al., 2010), and self-other source monitoring in schizophrenia (Wang et al., 2011).
In OCD, the anterior midcingulate cortex (aMCC) (Vogt, 2005), a brain region involved in multiple functions, such as pain, fear avoidance (Vogt, 2005), error avoidance (Magno et al., 2006), and reinforcement-guided decision-making (Rushworth and Behrens, 2008) is of particular relevance, representing the neurosurgical target for cingulotomy (Dougherty et al., 2002). This procedure is known to reduce striatal volume (Rauch et al., 2000). Further, the aMCC harbors the rostral cingulate zone (Shackman et al., 2011), that is, the source of the error-related negativity (Debener et al., 2005), an electrophysiological correlate of performance monitoring, which has consistently been shown to be enhanced in OCD patients (Endrass et al., 2008, 2010; Riesel et al., 2011). In addition to electrophysiological data, neuroimaging studies of OCD patients have also linked the aMCC with overactive error processing (Fitzgerald et al., 2010). Considering its anatomical location and connections, the aMCC has been suggested to represent a hub region that is ideally suited to integrate information and to refine behaviors motivated by negative feedback (e.g., pain or punishment) via its direct connectivity with limbic, motor, and dopaminergic midbrain areas (Shackman et al., 2011). In particular, the anatomical connection of the aMCC with the ventral striatum, including the nucleus accumbens [the cingulostriatal pathway (Kunishio and Haber, 1994)], has been linked with the coordination of aversively motivated instrumental behaviors (Shackman et al., 2011).
The present study sought to ask whether altered coupling occurs between the aMCC and other CBTGC circuit components, and the ventral striatum in particular, when OCD patients experience contexts that involve errors and negative feedback. Therefore, we tested for altered functional coupling between the aMCC and the ventral striatum in OCD patients in an experimental context that involves both regions of interest, applying the monetary incentive delay (MID) task (Knutson et al., 2000, 2001) in conjunction with PPI analysis. Noteworthy, a previous univariate GLM analysis of this dataset had indicated altered activation in medial prefrontal and cingulate cortices in OCD patients, but no group differences with respect to the regional activity of the striatum (Kaufmann et al., in revision). As a seed region for PPI, the present study used an aMCC region located in the cingulotomy target area (Rauch et al., 2000), approaching areas of the aMCC white matter abnormalities in OCD (Szeszko et al., 2005). Based on CBTGC circuit models in OCD (Kopell and Greenberg, 2008), we predicted altered coupling in the cingulostriatal pathway in OCD patients during anticipation of potential punishment, a condition that is relevant to both OCD symptomatology due to potentially negative consequences in the case of erroneous performance (Alonso et al., 2008; Salkovskis, 1985) and aMCC function (Shackman et al., 2011).
Materials and Methods
Participants
Nineteen adult outpatients with a DSM-IV diagnosis of OCD and 19 healthy controls matched for age, gender, intelligence, and handedness were recruited from the OCD outpatient clinic at the Humboldt-Universität zu Berlin. Patients were interviewed by a licensed clinical psychologist and diagnosed using the German version of the Structured Clinical Interview for DSM-IV (First et al., 1996). Ten patients had cormorbid diagnoses (affective disorder, n=7; phobic disorders, n=3; impulse control disorder, n=3). Three out of nineteen patients of the analysis sample were receiving medications to treat OCD at the time of the functional magnetic resonance imaging (fMRI) examination (one patient took clomipramine 10 mg/day, the second a combination of paroxetine 30 mg/day and clomipramine 75 mg/day, respectively, and the third took venlafaxine 75 mg/day). No patient received benzodiazepines within 4 weeks before the scanning session. Exclusion criteria for patients and healthy controls included cardiac pacemakers or other metallic implants or artifacts, and pregnancy. Patients had no significant neurological illness, no previous psychosurgery, no current or past substance abuse or dependence, dementia, delirium, schizophrenia, delusional disorder, or other psychotic disorder. Healthy controls had neither DSM-IV diagnosis (as indicated by a negative patient report in a SCID screening questionnaire) nor a first-degree relative with a reported history of psychiatric disorders. All participants provided written informed consent according to the institutional guidelines before enrollment. t-Tests for independent samples were used to compare groups regarding age, IQ, handedness, earnings in the MID task, and STAI test scores. t Values, means and standard deviations of questionnaire data are available in Table 1. The study was approved by the local ethics committee.
Comparisons are based on two-sample t tests. More information regarding symptom dimensions is available in Supplementary Tables S1a and b. (Supplementary Data are available online at
OCD, obsessive–compulsive disorder; SD, standard deviation; STAI, State and Trait Anxiety Inventory; Y-BOCS, Yale-Brown Obsessive–Compulsive Scale; OCI-R, Obsessive–Compulsive Inventory Revised; BDI, Beck Depression Inventory.
MID task
The MID task (Knutson et al., 2001) was adapted in the present study (Fig. 1). There were seven different types of trials: three trial-types with the possibility of winning money on a correct (i.e., timely) button press (potential reward trials), three trial-types with the possibility to avoid losing money on a correct button press (potential punishment trials), and the remaining trial-type in the absence of any monetary consequences (neutral trials). At the beginning of each trial, one of seven different cues was shown to indicate trial type. Participants were asked to press a button as soon as the target stimulus (gray colored square) appeared. Depending on the performance (i.e., timely motor response), participants received feedback about winning or losing money. Each run consisted of 72 trials, that is, 27 indicating potential reward, 27 indicating potential punishment, and 18 neutral cues with each trial lasting for 11.6 sec on average (see Fig. 1 for timing details). The interstimulus interval (from target offset to cue onset) ranged from 6570 to 7370 msec. The experiment lasted for 14 min (depending on the subject's response time). Task difficulty was continuously adapted to achieve a 66% success level in each subject across a task run, which was derived from individually tailored reaction times from the training session, and by adapting the response deadline as a result of the averaged reaction times in previous trials and the correctness of the immediately preceding trial during the test runs.

Monetary incentive delay task.
Clinical scales
Severity of OCD symptoms was evaluated using the German versions of the Yale-Brown Obsessive–Compulsive Scale (Goodman et al., 1989) and the Obsessive–Compulsive Inventory Revised (Foa et al., 2002). In addition, the State and Trait Anxiety Inventory-Revised (Spielberger et al., 1970) and the Beck Depression Inventory (Beck et al., 1961) were administered before scanning. The Edinburgh Handedness Inventory (Oldfield, 1971) was used to classify handedness. For matching purposes, all participants completed a German vocabulary test (Schmidt and Metzler, 1992) as a brief measure of verbal intelligence.
fMRI acquisition and statistical analysis
Stimuli were generated using Presentation (Neurobehavioral Systems) and were projected by means of a mirror system attached to the head coil. To reduce head motion, the subjects' head was immobilized by a vacuum head cushion. Earplugs were used to attenuate background noise. Headphones were used to communicate with subjects. Before functional runs, 176 anatomical Modified Driven Equilibrium Fourier Transform (MDEFT) slices were acquired (spatial resolution 1×1×1 mm, repetition time (TR)=12.24 msec, echo time (TE)=3.56 msec, flip angle=23°, 256×224 matrix) during the training session of the MID task on a 1.5 T Siemens Sonata scanner. A total of 450 volumes (T2*-weighted single-shot gradient echo planar imaging sequence) were acquired in each of the two test runs using the following parameters: TR=1870 msec, TE=40 msec, 33 consecutive slices, 3×3×3.5-mm voxel, flip angle=90°, field of view (FOV)=192 mm, 64×64 matrix.
fMRI data analysis was performed using SPM5 (Version 1782, Statistical Parametric Mapping,
PPI analysis
PPI analyses test whether one region, the seed time series, changes its coupling with other regions due to a specific experimental manipulation (Friston et al., 1997; Gitelman et al., 2003). To perform PPI analysis, seed voxels were derived by extracting the first eigenvariate time series (volumes of interest within SPM5) from a 5-mm sphere centered around three aMCC regions. Volumes of interest were adjusted for effects of interest. In the present study, seed voxels were extracted from the aMCC gray matter regions encompassing both the cingulotomy target region in Brodmann area 24 (Dougherty et al., 2002; Rauch et al., 2000) and regions where functional anisotropy decreases have previously been described in OCD (Szeszko et al., 2005). Since these coordinates were located in the white matter, the nearest gray matter locus displaying individual task effects was used for seed voxel extraction. Three control steps were conducted to ensure validity of seed voxel extraction (see Supplementary Methods for details; Supplementary Data are available online at
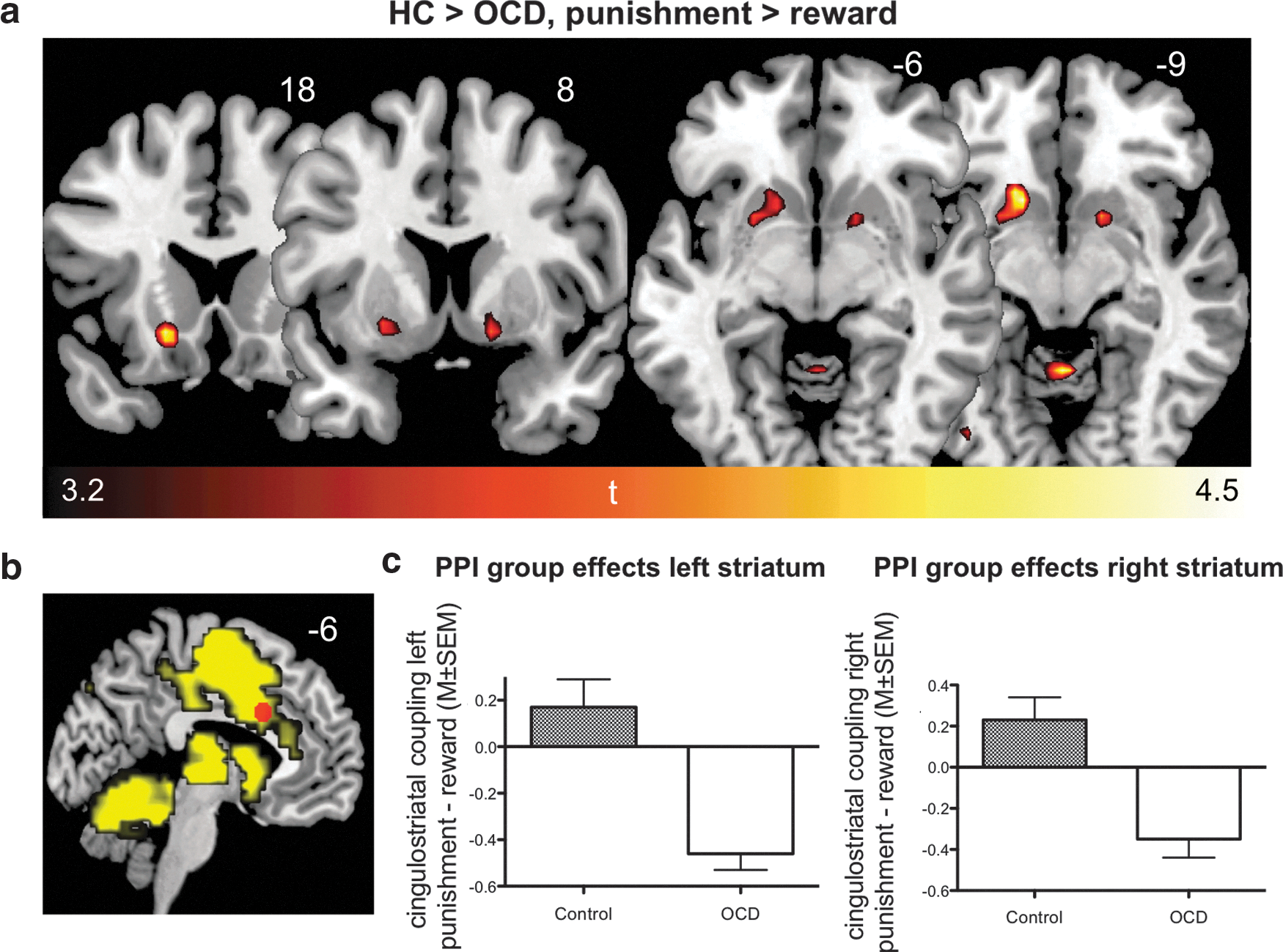
PPI group effects in the bilateral ventral striatum, displayed coronally and axially
PPI analyses regress the interaction between a BOLD seed voxel time series (here: aMCC seed) and a psychological parameter (anticipation of punishment minus anticipation of reward) on all voxel time series in the brain (Friston et al., 1997; Gitelman et al., 2003). Therefore, the seed time series and the psychological factor are multiplied (element-by-element multiplication), resulting in the PPI interaction term regressor. Accordingly, the design matrix for PPI analyses includes three columns: the psychological parameter (onsets of anticipated punishment and reward), the seed voxel time series (physiological parameter), and the PPI interaction term. Modeling the interaction term in the first column, and using the contrast [1 0 0] allowed to ask for voxel time series that are predicted by the PPI interaction term regressor. Importantly, the extracted time series was deconvolved with a canonical hemodynamic response function before creating the interaction term, to account for the fact that interaction of brain regions are not expressed on the level of hemodynamic responses, but on a neuronal level (Gitelman et al., 2003). In addition, six motion correction parameters were included into the PPI design matrix representing effects of no interest.
PPI group analysis
For each subject, PPI effects were estimated at each voxel, and statistical parametric maps (SPMs) were calculated for the psychological parameter contrast of interest (anticipated punishment minus anticipated reward). These individual SPMs were submitted to a random effects analysis, and a two-sample t test was performed to compare PPIs between groups. Due to the fact that the psychological parameter represents a term subtracting punishment (i.e., [1]) and reward (i.e., [−1]) conditions, the PPI regressor asks for voxel time series that are predicted by the seed during punishment, whereas signaling inverse to the seed time series is predicted in the reward condition. Therefore, the PPI regressor identifies voxels that show a difference in regression slopes when comparing regressions of the seed time series between the two conditions, indicating that a change in task conditions has an effect on the connectivity among the seed and the identified region. This difference in regression slopes is subsequently compared between two groups, and accordingly, regions showing significant PPI group effects (two-sample t test) reveal significant group differences with respect to the differences in regression slopes. For this reason, explanatory PPI analyses are needed for reasonable interpretation of PPI group effects, allowing one to attribute the PPI group effects to one group, and ideally, one condition driving it. To characterize PPI differences between groups in detail, effects are provided using an uncorrected threshold of p<0.001 with an extent of 5 voxels (Tables 2 and 3). In accordance with previous PPI studies (Das et al., 2005; Mobbs et al., 2009), we performed small volume corrections for a priori regions of interest, using a threshold of p<0.05 (false discovery rate corrected for multiple comparisons). These were applied using one combined, bilateral mask of the following regions in all analyses: caudate, putamen, pallidum (as implemented in the Automatic Anatomic Labeling tool integrated in the WFU PickAtlas (
Cluster sizes indicate the number of contiguous voxels for each cluster.
Between-group effects thresholded at p<0.001 (uncorrected; minimum cluster extent, five contiguous voxels).
PPI effect coordinates (x, y, z) are given in MNI space.
Magnitude statistics correspond to a minimum (whole-brain) uncorrected threshold of p<0.001 with a minimum cluster extent of five contiguous voxels.
Direction of the PPI: negative indicating a more negative relationship between seed and PPI effect region during punishment relative to the reward condition.
Significant between-group main effect differences in a priori regions of interest (one combined mask of bilateral caudate, putamen, and pallidum masks generated from the AAL tool of the Wake Forest University WFU PickAtlas [
BA, Brodmann area; HC, healthy controls; CS, cluster size, PPI, psychophysiological interaction; MNI, Montreal Neurological Institute; AAL, Automatic Anatomic Labeling; FDR, false discovery rate.
Cluster sizes indicate the number of contiguous voxels for each cluster.
Between-group effects thresholded at p<0.001 (uncorrected; minimum cluster extent, five contiguous voxels).
PPI effect coordinates (x, y, z) are given in MNI space.
Magnitude statistics correspond to a minimum (whole-brain) uncorrected threshold of p<0.001 with a minimum cluster extent of five contiguous voxels.
Direction of the PPI: negative indicating a negative interaction between seed and PPI effect region during potential punishment.
Significant between-group main effect differences in a priori regions of interest (one combined mask of bilateral caudate, putamen, and pallidum masks generated from the AAL tool of the Wake Forest University WFU PickAtlas [
Explanatory PPI analyses
Within one group, positive beta values indicate a positive relationship between the seed and an identified region in one condition relative to the other (positive PPI), whereas negative beta values index a negative relationship in one condition relative to the other (negative PPI). Accordingly, comparing effects in the OCD group to the control group reveals more positive (OCD>Control) or more negative (Control>OCD) beta values in the OCD group. However, the interpretation of PPI group effects is hindered by the fact that difference values of two groups are subtracted from each other. As for univariate GLM group comparisons, a second level group effect using individual first level contrast images (already including a subtraction of two conditions) as an input cannot be directly attributed to a single group or condition driving it. To overcome this problem, two additional explanatory PPI analyses were conducted. Thus, regressions of the original aMCC seed time series for both conditions (punishment, reward) were calculated separately, comparing one condition at a time to the implicit baseline, instead of directly subtracting them from one another as in the original PPI analysis. This resulted in two new first level PPI contrasts in each subject (anticipated punishment>implicit baseline, anticipated reward>implicit baseline). From a technical perspective, the only difference between the original PPI first level analysis and the explanatory PPI first level analysis was that task parameters (punishment or reward) were removed from the psychological parameter (i.e., [1 0] and [0 1] instead of [1 −1]) in the original design matrix) one at a time before calculating the PPI interaction term. Thus, two new interaction term regressors were built and accordingly, two new first level models were estimated, resulting in two separate SPMs (i.e., anticipation of punishment>implicit baseline, anticipation of potential reward>implicit baseline) for each participant. Subsequently, these new individual contrast images were fed into random effects analyses. This allowed to perform group comparisons (two-sample t tests as before) with respect to the potential punishment condition. The attempt to decompose PPI effects might be complicated by the fact that PPI group analyses compare regression slopes between conditions, whereas explanatory PPI analyses compare the regression slope of single conditions to an implicit baseline (grand mean). Therefore, we validated the explanatory PPI procedure by correlating PPI beta estimates from the original PPI analysis with difference (i.e., subsequently subtracted) values of beta estimates from explanatory PPI analyses, assuming that a high correlation between these two variables would indicate successful decomposition of the PPI group effect. To validate successful decomposition of the PPI group effects using explanatory PPI analysis, the combined sample (n=38) was used to calculate Pearson correlation coefficients between the PPI beta estimates of the left (MNIXYZ=−15, 18, −9) and right (MNIXYZ=15, 9, −6) ventral striatal peak voxels of significant PPI group effects and difference values of the PPI beta estimates of the same exact coordinate from the explanatory PPI analyses (PPI beta estimates potential loss of reward>implicit baseline minus PPI beta estimates potential reward>implicit baseline), revealing a highly significant correlation (left ventral striatum: r=0.97, R 2=0.94; right ventral striatum: r=0.84, R 2=0.71), assuring accuracy and validity of the explanatory PPI results.
Results
PPI results
PPI group analysis revealed significant group differences (two-sample t-test, Controls>OCD, potential punishment minus potential reward) for the ventral striatum bilaterally (Fig. 2a), that is, ventromedial left caudate head and right putamen. Additionally, PPI group differences were observed between the aMCC seed and inferior, postcentral, and parietal areas, and further medial prefrontal and cerebellar areas in the patient group compared with healthy controls (Controls>OCD; Table 2). These PPI effects for Controls>OCD indicate that the relationship between the aMCC and PPI effect regions is more positive in the reward compared with the punishment condition in Controls in relation to patients, or vice versa, a more negative relationship between the aMCC and PPI effect regions in the punishment compared with the reward condition in patients in relation to controls. Therefore, reasonable interpretation of PPI group differences requires consideration of within-group (main) effects (Table 2) and additional explanatory PPI analyses (see below). PPI group differences for OCD>Controls were observed for a region in the middle temporal gyrus. Analyses considering medication status and potential partial white matter signal extraction as potential confounds are available as Supplementary Materials (Supplementary Tables S2–S5 and Supplementary Figs. S1 and S2). There was no significant correlation between PPI effects and OCD symptom severity or duration of OCD in years (see Supplementary Methods and Results).
Explanatory PPI analyses
Comparisons between groups regarding aMCC connectivity in the potential punishment condition (relative to implicit baseline, derived using explanatory PPI analyses revealed significant group differences (two-sample t-tests) for cingulostriatal coupling (Controls>OCD), that is, right ventral putamen extending into nucleus accumbens (Fig. 3a and Table 3) and left ventral caudate head (Fig. 3a). Besides group differences in basal ganglia regions, inverse coupling between the aMCC and orbitofrontal, inferior parietal, lingual and cerebellar areas and the red nucleus was evident. All the above effects were observed for Controls>OCD, indicating negative beta values in patients compared with controls. A post hoc search for group differences in aMCC connectivity with the midbrain during potential punishment was conducted, since dopaminergic centers are critical in modulating prediction error responses in the aMCC and the ventral striatum (Rushworth and Behrens, 2008). This analysis revealed that negative coupling effects similar to the ones observed in the cingulostriatal pathway were also present between the aMCC and the substantia nigra, given that the red nucleus cluster extended into this region when using a liberal threshold of p<0.005, uncorrected (Fig. 4).

Explanatory PPI effects. Regressions of the aMCC seed time series during potential punishment (minus implicit baseline) compared between both groups
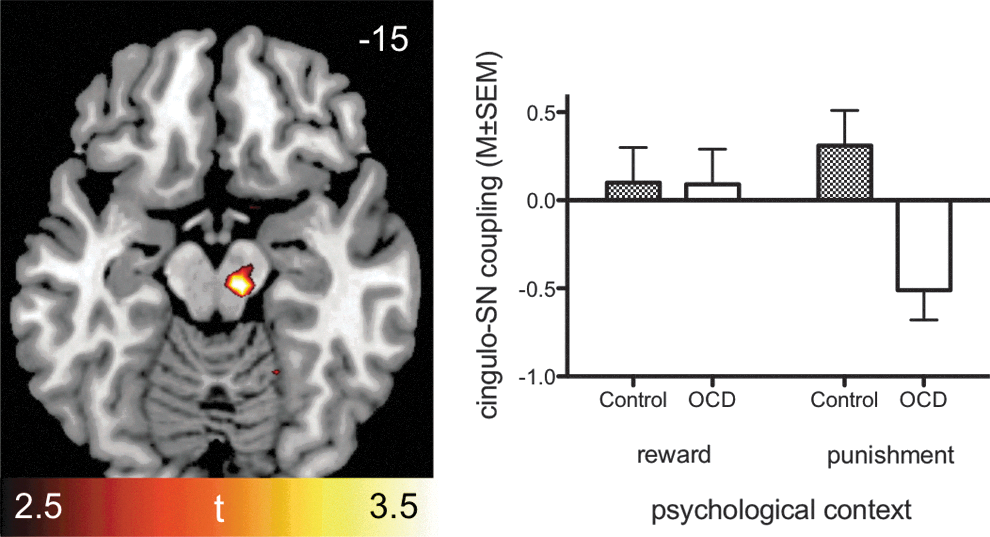
Results of a post hoc small volume search (p<0.005, uncorrected) for the midbrain revealing differences (Controls>OCD) with respect to coupling between aMCC seed and SN (MNIXYZ=12, −15, −15, t=3.13) and the red nucleus due to inverse coupling between these two regions in OCD patients anticipating potential punishment. SN, substantia nigra.
Discussion
This study tested for aMCC connectivity abnormalities in OCD patients anticipating potential punishment. PPI analyses revealed significantly altered, inverse coupling of the aMCC with the bilateral ventral striatum and inferior parietal regions known to exhibit direct anatomical connections with the aMCC (Kunishio and Haber, 1994; Vogt, 2005; Vogt and Pandya, 1987). More precisely, cingulostriatal coupling was identified to be specifically driven by an inverse relationship of these two areas in the potential punishment condition, as revealed by explanatory PPI analyses. With respect to CBTGC model assumptions, these data extend the existing literature by showing abnormal connectivity among CBTGC components known to conjointly exhibit regional hyperactivity during symptom provocation (Breiter et al., 1996; Rauch et al., 1994), while at the same time showing abnormal connectivity between the aMCC and regions showing functional abnormalities in OCD outside traditional CBTGC circuitry (Menzies et al., 2008a). In summary, these findings suggest that dysfunctional functional coupling in CBTGC circuits affects cingulostriatal pathways when OCD patients anticipate potential punishment, while at the same time providing evidence for the view that the aMCC interacts not only with striatal, but rather with a more distributed cortical set of areas in OCD patients experiencing symptom-relevant contexts.
In addition to the observation of altered coupling in the cingulostriatal pathway, it is worth noting that cortical areas showing altered inverse coupling with the aMCC during anticipation of potential punishment included the orbitofrontal cortex (OFC). This area is of particular relevance for the pathophysiology of OCD because it is known to reveal increases in activity during symptom provocation (Breiter et al., 1996) and decreased activity during reversal learning in OCD (Chamberlain et al., 2008). Consistent with CBTGC circuit considerations involving the OFC (Kopell and Greenberg, 2008), one of the few OCD studies applying connectivity methods on fMRI data found increased resting-state functional connectivity between the ventral striatum and the OFC (Harrison et al., 2009). Crucially, the ventral striatal/nucleus accumbens coordinates used as seeds by Harrison et al. (2009) highly correspond to the areas showing abnormal connectivity with the aMCC in the present study. Further indication for specific interplay of the aMCC, the ventral striatum, and OFC in OCD is provided by the fact that all three regions have repeatedly been shown to respond during symptom-provocation (Breiter et al., 1996; Rauch et al., 1994). In addition, structural MRI analyses have previously pointed at abnormalities in the ventral striatum (Pujol et al., 2004), and volume reductions exclusively in the bilateral striatum following cingulotomy (Rauch et al., 2000), and a meta-analytic study (Rotge et al., 2009) confirmed volume reductions of the left aMCC and bilateral OFC in OCD. Considering anatomical evidence indicating that limbic loops transfer information among each other (Haber, 2003), these findings might suggest interactions among CBTGC areas, namely, the aMCC, OFC, and ventral striatum in symptom-relevant contexts in OCD.
In addition to altered coupling among CBTGC regions traditionally involved in the pathophysiology of OCD, altered aMCC connectivity with parietal and lingual occipital areas, thus regions outside of CBTGC circuitry was evident. These areas partially overlap with locations of white matter reductions in OCD, that is, inferior parietal (Menzies et al., 2008b; Szeszko et al., 2005) and lingual (Szeszko et al., 2005) areas (Tables 2 and 3), and also converge with meta-analytic evidence of functional alterations outside of traditional CBTGC circuitry, predominantly found in parietal, medial occipital, and cerebellar areas (Menzies et al., 2008a).
From a methodological perspective, this is one of the few studies (Guyer et al., 2008; Kasahara et al., 2010; Lanius et al., 2004) that used PPI to compare diagnostic groups in the neuropsychiatric field, in contrast to a majority of studies applying univariate GLM analyses on experimental fMRI data. As already indicated above, it is crucial to apply connectivity methods, such as PPI, in the case of theoretical assumptions of altered networks (Stephan, 2004). More precisely, univariate GLM and connectivity analyses are based upon two fundamentally different concepts of brain functioning, the first reflecting the idea of functional segregation of brain areas (specific regional responses to a task component), and the latter reflecting functional integration (specific interregional interaction to a task component) (Friston, 2011). Following the notion that identification of candidate elements of a neural system by means of GLM-based analyses requires subsequent analysis of their functional integration to provide a model for the underlying neural system (Stephan, 2004), the present study tested for altered corticostriatal coupling in OCD patients using a seed in the aMCC. The fact that the observed effects concern regions sharing direct anatomical connections with the aMCC (i.e., inferior parietal areas, ventral striatum, substantia nigra) advocates for the view that PPI, despite its simplicity, is a useful approach to ask for whole-brain connectivity changes (Friston et al., 1997) of one particular, extensively connected functional hub region as it is the case for the aMCC (Shackman et al., 2011). Taken together, previous univariate GLM analyses have provided evidence for regional functional abnormalities of the cingulate, including the aMCC, and the present data suggest that the anticipation of potential punishment evokes abnormal functional integration among the aMCC and areas within (the ventral striatum) and outside (lingual and inferior parietal gyri) of CBTGC circuitry. While these findings of abnormal cingulate connectivity might suggest that regional functional abnormalities in occipital and parietal areas (Menzies et al., 2008a) are influenced by activity in the aMCC, this question requires application of effective connectivity approaches, such as dynamic causal modeling [DCM (Friston et al., 2003)], which directly address causal interregional interaction in response to task manipulations. Noteworthy, the present PPI data suggest to consider the inclusion of areas outside of traditional CBTGC circuitry in the DCM modeling, particularly in the case that the applied task is known to involve medial occipital and parietal areas.
From a clinical perspective, the finding of altered connectivity in the cingulostriatal pathway has implications for neurosurgical and stimulation treatment in OCD. This becomes evident considering that an aMCC area located in the cingulotomy target region (Dougherty et al., 2002; Rauch et al., 2000) was used as a seed, whereas the ventral striatum, the main PPI effect region, represents a primary target for deep brain stimulation (DBS) in OCD (Denys et al., 2010; Rauch et al., 2006). Remarkably, recent studies emphasize particular efficiency of using the anterior internal capsule, that is, the anatomical connection between the aMCC and the ventral striatum (Kopell and Greenberg, 2008), as a target for stimulation and surgery (Ruck et al., 2008). Thus, the present findings might provide potential explanations for the observation that targeting of different neuroanatomical locations has been shown to be beneficial for refractory OCD patients, since these treatments all influence locations within the cingulostriatal pathway. Future research on DBS stimulation in OCD might benefit from the observation of altered connectivity between two different target locations, and from examinations of how DBS affects cingulostriatal connectivity. Similarly, studying how cingulostriatal stimulation affects the OFC and the above-mentioned areas outside CBTGC circuitry revealing functional alterations (Menzies et al., 2008a) could help to further specify neuroanatomical models of OCD.
Limitations of the present findings concern the small sample size, and the fact that PPI analyses involve pair-wise regressions of a single brain region, thus limiting conclusions about complex dynamic interactions encompassing multiple brain regions. Further, PPI do not inform about causality among influences between brain regions, favoring the application of DCM analyses (Stephan et al., 2010) in future examinations of corticostriatal connectivity in OCD. Further limitations of the present study concern the application of fMRI as a method that does not address neurochemical mechanisms that might underly abnormal cingulostriatal coupling. While the experimental context of this study (Knutson et al., 2000, 2001) and post hoc analyses revealing negative aMCC-substantia nigra coupling suggest dopaminergic involvement, it is known that direct, reciprocal glutamatergic connections exist between the aMCC and the ventral striatum (Kopell and Greenberg, 2008), which might also contribute to the abnormalities found here. Depletion and neurochemical tracer studies could potentially reveal specific transmitter interplay in the cingulostriatal pathway in relation to obsessive–compulsive behavior.
In summary, our results reveal specific aMCC connectivity abnormalities in OCD patients in an experimental context that challenges aMCC function. Our finding of inverse cingulostriatal coupling demonstrates direct evidence of a dysfunctional pathway within CBGTC circuits, while at the same time identifying altered aMCC coupling with cortical areas outside of traditional CBTGC circuitry. Therefore, the present findings relate to neurobiological models of OCD. Further, these data are clinically relevant considering that contemporary neurosurgical OCD treatments target components of the cingulostriatal pathway. Finally, the present data favor the application of effective connectivity approaches to study corticostriatal connectivity in OCD.
Footnotes
Acknowledgments
Jan Beucke is supported by a Ph.D. scholarship from Evangelisches Studienwerk e.V. Villigst (Schwerte, Germany), and is also an ERP scholar of the German National Academic Foundation. The authors thank Eva Kischkel, Ph.D., and Rüdiger Spielberg, Ph.D. for clinical assessments, Rainer Kniesche for technical assistance, and acknowledge access to the MR scanner kindly provided by Charité Universitätsmedizin Berlin.
Author Contributions
C.K., T.E. and N.K. designed the experiment, and C.K. collected the data. J.C.B. and C.K. analyzed the data, D.D.D., C.L., T.D., R.G. and N.K. contributed to data interpretation and discussion, and J.C.B. and C.K. wrote the paper. All of the authors contributed to the preparation of the manuscript.
Author Disclosure Statement
The authors declare no competing financial interests.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.