
Abstract
The lateral occipital cortex (LOC), a visual area known to be involved in object recognition, was dynamically coupled with each of two distributed patterns of neural activity depending upon the percept (default or alternative) elicited by a bistable figure. The two distributed patterns included core nodes of the default-mode and frontoparietal networks (FPN), and they were most highly coupled to each other during the alternative percept, whereas they were less coupled during the default percept. Surprisingly, the regions associated with the nonengaged percept exhibited the highest connectivity to the LOC. Together, these findings reveal a dynamic organization between the default mode and the FPNs, and the incoming bottom-up visual stream during perceptual binding of visual images.
Introduction
The mechanism by which the neural correlates of human vision segment and bind features to form unified percepts from a complex visual world is a long-standing central question that has also been linked to more general questions related to the neural correlates of awareness and consciousness (Leopold and Logothetis, 1999; Rees et al., 2002; Sterzer et al., 2009). Image segmentation is a complex process by which stimulus elements are perceptually arranged into a unified whole. A bistable figure presents a unique opportunity to investigate mechanisms involved in segmentation of visual input, because one stimulus elicits two mutually exclusive percepts representing alternative organizations of the same visual input. Although neuroimaging studies have previously confirmed the involvement of the parietal and frontal brain regions in high-level visual processes, including bistable perception (Kleinschmidt et al., 1998), there is no established framework to describe the underlying neural mechanisms of image segmentation. We envision that this complex process of alternating visual perceptions will employ large-scale distributed neural systems.
The default-mode network (DMN), sometimes referred to as the task-negative network, has been defined by task-induced deactivations as well as higher energy consumption during rest, and consists of temporal and midline structures that are known to be more active during rest than during a task (Buckner et al., 2008; Greicius et al., 2003; Gusnard et al., 2001; Raichle et al., 2001). It has also been associated with internal stimuli or self-reflection as well as memory of past events (Andrews-Hanna et al., 2010). The frontoparietal network (FPN), sometimes referred to as the task-positive network, is classically defined by task-induced activations, and consists of the dorsal, frontal, and parietal regions associated with volitional tasks that require attention to external stimuli (Corbetta and Shulman, 2002; Dosenbach et al., 2007; Kastner and Ungerleider, 2000). These two networks have also been identified on the basis of spontaneous correlations during resting states characterized by anticorrelations between them (Anderson et al., 2011; Fox et al., 2005), suggesting an intrinsic oppositional functional organization of neural processes that mediate cognitive tasks. Despite a general consensus regarding the regions comprising these networks, there is a lack of consensus regarding their functions.
In this study, functional magnetic resonance imaging (fMRI) and functional connectivity methods were employed to identify neural substrates and dynamics engaged during each of the mutually exclusive percepts elicited by a common bistable figure, the Schröder Staircase (Schröder, 1858). Subjects viewed alternating 15-sec blocks of rest and stimulus, and were instructed on each stimulus block that percept to maintain, that is, default or alternative. All subjects were well practiced, and they demonstrated competence with the task before scanning. This paradigm differs from previous studies of bistable perception (Kleinschmidt et al., 1998; Tong et al., 1998) by providing a targeted percept with instructions to maintain a percept for each 15-sec block. This paradigm was chosen to provide a structured focus on each percept, which permitted an experimental approach to investigate the underlying neural circuitry associated with each percept.
In the default condition, the figure was readily perceived as a familiar staircase, whereas in the alternative condition, the figure was perceived as an inverted staircase. Although prior investigations of bistable perception have considered the role that attention plays in forming each of the percepts (Meng and Tong, 2004; Slotnick and Yantis, 2005), the attention network and its relation to the DMN have not been previously implicated in this process. As is typical with bistable figures, the two percepts differed with respect to the volitional effort and attention required for their realization, suggesting a putative role for both the attentional control and the DMNs. In this study, we test the hypothesis that mutually exclusive visual image segmentations, as in the case of two bistable percepts, are associated with neural processes that engage both the default-mode and the frontoparietal attention networks. Further, we compare the coupling between the networks and the bottom-up visual stream during the two perceptual states to investigate the intrinsic dynamic organization associated with these percepts.
Materials and Methods
Subjects
A total of 12 healthy volunteers participated in the functional imaging study (8 men and 4 women; ages 18–27 years of age; mean=22.8 years of age), as approved by the institutional review board of the Columbia University Medical Center. All subjects were informed about possible risks of MRI and provided consent according to the established guidelines.
Stimulus
The stimulus was a black-and-white line drawing of a common bistable figure (Supplementary Fig. S1; Supplementary Data are available online at
Functional imaging procedures
The functional study was run as a block design in which the stimulus was presented for 12 fifteen-sec epochs, each of which was preceded by a 15-sec baseline epoch that featured a black screen with a crosshair (+). Before scanning, the default and the alternative percepts were determined for each subject based on the percept that the subject reported as seen first and most automatically. For all subjects, the default percept was the ascending staircase most resembling a familiar staircase, and the alternative percept was the upside-down staircase that appeared to be suspended in midair. The subject was instructed to hold the default percept for the first 15-sec stimulus epoch, and then, following a rest epoch, instructed to hold the alternative percept for the following 15-sec stimulus epoch, and to continue this alternation for the duration of the 6.0-min run. The target percepts were cued by the written words alternative or default above the image, and the subjects indicated on a keypad the actual engaged percept and whenever a perceptual switch (voluntary or otherwise) occurred. Subjects practiced outside the scanner until they could perform this perceptual task. Button-press indications of the engaged percept confirmed that on average, the default percept was sustained for a total of 92.36±6.48 sec, whereas the alternative percept was sustained for a total of 75.7±6.16 sec, and are consistent with known difficulty and attentional differences between the two percepts. The average total time that the default percept occurred spontaneously during the target alternative condition was 20.34±6.29 sec, whereas the average total time that the alternative percept occurred spontaneously during the target default condition was 12.01±4.92 sec, and also consistent with the default percept as the more natural and less effortful of the two.
Image acquisition and analysis
Functional images were acquired on a 1.5T GE MRI scanner located in the Columbia University fMRI Research Center, New York, NY. Whole-brain eco planar functional images (EPI) were collected with an 8-channel GE head coil in 25 contiguous axial slices obtained parallel to the AC/PC line (TR=3000 ms, TE=35 ms, flip angle=84 degrees, FoV=19.2×19.2 cm2, array size=128×128, spatial resolution of acquisition=1.5×1.5×4.5 mm, voxel size after spatial normalization=2×2×2 mm). One hundred twenty whole-brain images were acquired during each of two identical 6-min runs. High-resolution 3-D anatomical scans were also acquired with a T1-weighed SPGR sequence (TR=19 ms, TE=5 ms, flip angle=20 degrees) FoV=220×200 mm, a slice thickness of 1.5 mm, in-plane resolution of 0.86×0.86 mm, and 124 slices per image.
Image preprocessing and statistical analysis were completed using SPM8 software (Wellcome Department of Cognitive Neurology, University College London, UK). Functional T2*-images were slice-timing corrected and spatially realigned to the first volume of the first run. Finally, images were smoothed with a Gaussian kernel of 8.0×8.0×8.0-mm full-width half-maximum, and a 128-s temporal high-pass filter was applied.
General linear model analysis
Statistical analysis of the blood oxygen level-dependent (BOLD) signal was modeled using a single-factor percept, with two levels: alternative and default. The analysis aimed to detect activity associated with each perspective. Perceptual durations (according to button presses) for the default and alternative percepts were convolved with the canonical hemodynamic response function (HRF). Additional nuisance regressors, that is, six motion parameters, mean white-matter, and mean CSF signal, were included to remove unnecessary noise from the data. Contrasts of resulting beta-estimates (Default>Alternative and Alternative>Default) for each run separately were averaged across both runs, and were passed to 2nd-level random-effect analyses (one-sample t-tests). Beta-estimates from each condition were also passed to a 2nd-level random-effect analysis (paired t-test) to determine conjoined activation and deactivation common to both percepts in run 1, used for independent region of interest (ROI) analyses (see below).
Psychophysiological interaction analysis
The psychophysiological interaction (PPI) analysis measures the extent to which regions are differentially correlated between conditions (Friston et al., 1997), and is strictly correlative and not indicative of directional causation. While there are various approaches regarding the removal of task-associated variance in PPI analysis (McLaren et al., 2012; O'Reilly et al., 2012), we have adopted the long-standing standard approach as described in the current version of SPM8 (
Areas Active During the Default Percept as Defined by the Contrast Default>Alternative and Also Identified as Elements of the Default-Mode Network
ROI abbreviations, peak voxel MNI coordinates (x, y, z), t-values, and cluster sizes are shown for each region.
Asterisks indicate the regions that survive cluster correction thresholding at p<0.005 and a cluster size of 150.
DMN, default-mode network; GLM, general linear model.
Areas Active During the Alternative Percept as Defined by the Contrast Alternative>Default and Also Identified as Elements of the Frontoparietal Network
ROI abbreviations, peak voxel MNI coordinates (x, y, z), t-values, and cluster sizes are shown for each region.
FPN, frontoparietal network.
Independent ROI analysis
To test whether the DMN and FPN were significantly more active and functionally connected with the visual cortex during one percept versus the other, we conducted an independent ROI analysis using the Marsbar Toolbox (
Effective connectivity analysis
Effective connectivity analysis was carried out using dynamic causal modeling, DCM (Friston et al., 2003), as implemented in SPM8 (Wellcome Department of Cognitive Neurology, University College London, UK). Predictions based on the observed data consist of the combination of driving inputs, intrinsic connection activity, and bilinear modulation, which reflects the effects of experimental variables. In this case, the default and alternative percept conditions served as both the driving input (on individual regions) and the modulatory input (on connections between regions). These effects are modeled by the equation, dz1/dt=(A+umB)z2+Cui, in which dz1/dt is the state vector per unit time for the target region; z2 corresponds to timeseries data from the source region; ui indicates the direct input to the model; and um indicates input from the modulatory variable onto intrinsic pathways specified by the model. Activity in the target region is therefore determined by an additive effect of the intrinsic connectivity with the source region (Az2), the bilinear variable (umBz2, corresponding to the modulatory experimental manipulation), and the effect of direct input into the model (Cui).
Given our specific hypotheses, a fully specified model was estimated (i.e., intrinsic bilateral connections between the LOC, DMN, and FPN, with both conditions modulating all regions and connections). In each subject, the contrast (Default>Alternative) was calculated for each connectivity parameter and submitted to a one-sample t-test over all the subjects. Unless otherwise indicated, there were 11 degrees of freedom for all reported t-values.
Results
Functional magnetic resonance imaging
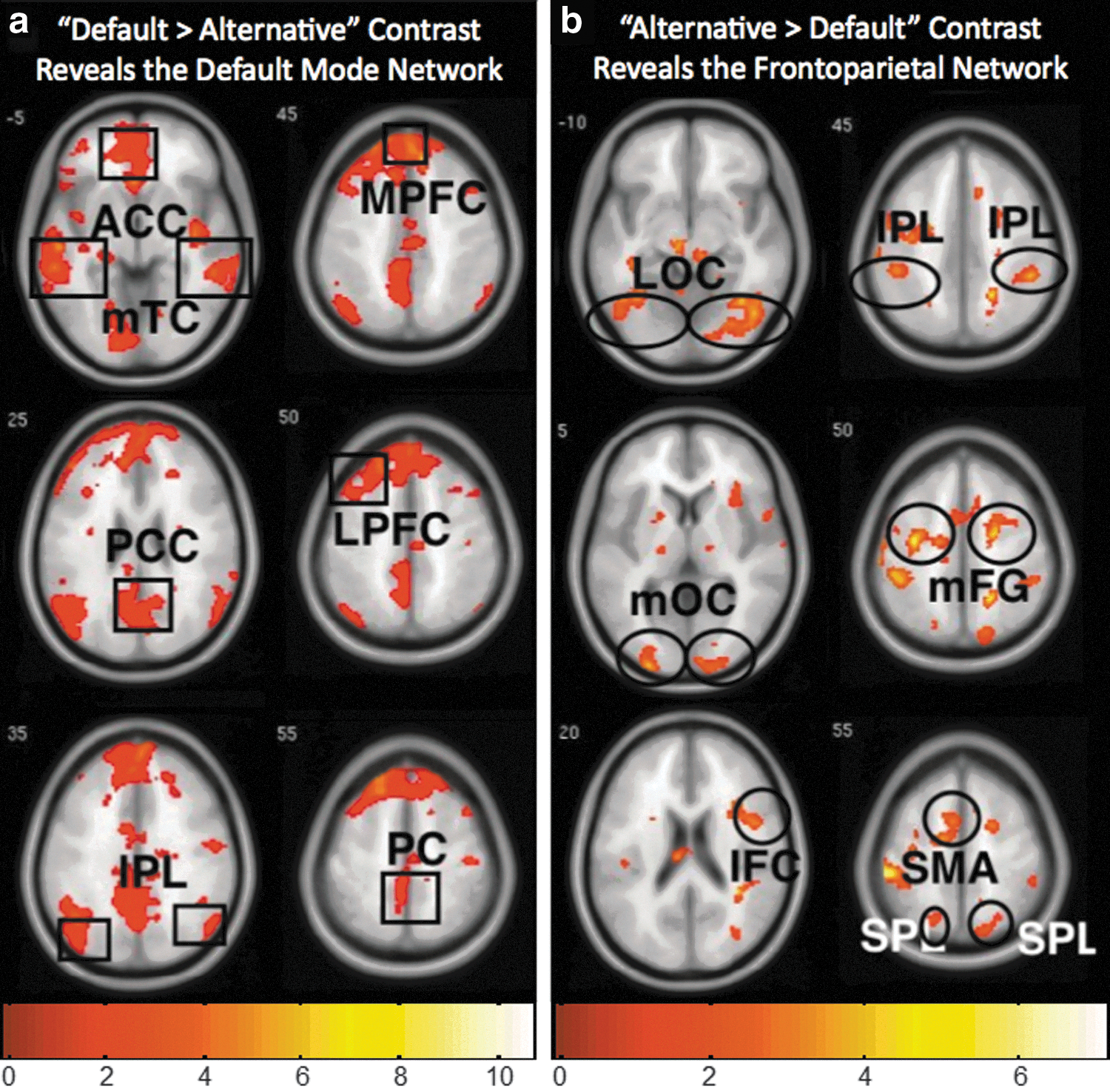
Patterns of whole-brain fMRI activity based on the BOLD response observed during the default>alternative contrast (Fig. 1a) and alternative>default contrast (Fig. 1b) corresponded to known activity patterns previously associated with the DMN (Anderson et al., 2011; Buckner et al., 2008; Greicius et al., 2003; Raichle et al., 2001) and FPN (Anderson et al., 2011; Corbetta and Shulman, 2002; Dosenbach et al., 2007; Kastner and Ungerleider, 2000), respectively. In particular, the default perspective activity (as defined by the contrast default>alternative; Table 1A) included the middle temporal cortex, anterior cingulate cortex, posterior cingulate cortex, inferior parietal lobule (IPL), medial prefrontal cortex, lateral prefrontal cortex, and precuneus (PC), which have been previously associated with the DMN (Buckner et al., 2008; Greicius et al., 2003; Raichle et al., 2001). In comparison, the alternative perspective activity (as defined by the contrast alternative>default; Table 1B) included the LOC, middle occipital cortex, inferior frontal cortex, IPL, superior parietal lobule, middle frontal gyrus, and supplementary motor area, which have previously been associated with the FPN (Corbetta and Shulman, 2002; Dosenbach et al., 2007; Kastner and Ungerleider, 2000). An independent ROI analysis confirmed that activation of the DMN, as a whole, was significantly greater during the default perspective (default>alternative, t=2.29, p<0.05), while activation of the FPN, as a whole, was significantly greater during the alternative perspective (alternative>default, t=2.01, p<0.05) (see the Materials and Methods section).
Functional connectivity with the LOC
The functional roles of the DMN and FPN in bistable image segmentation were explored in relation to the incoming bottom-up visual stream. PPI analysis of functional connectivity between the LOC, which was active during both percepts, and all other brain regions revealed that a higher connectivity was observed between the LOC and the network associated with the unconscious percept. For example, during the default percept (default>alternative contrast), LOC connectivity increased specifically with the FPN regions (Fig. 2a), whereas connectivity during the alternative percept (alternative>default contrast) increased specifically with the DMN regions (Fig. 2b). An independent ROI analysis confirmed that connectivity with the FPN was significantly greater during the default perspective (default>alternative, t=4.80, p<0.05), while connectivity with the DMN was greater during the alternative perspective (alternative>default, t=5.39, p<0.05) (see the materials and Methods section).
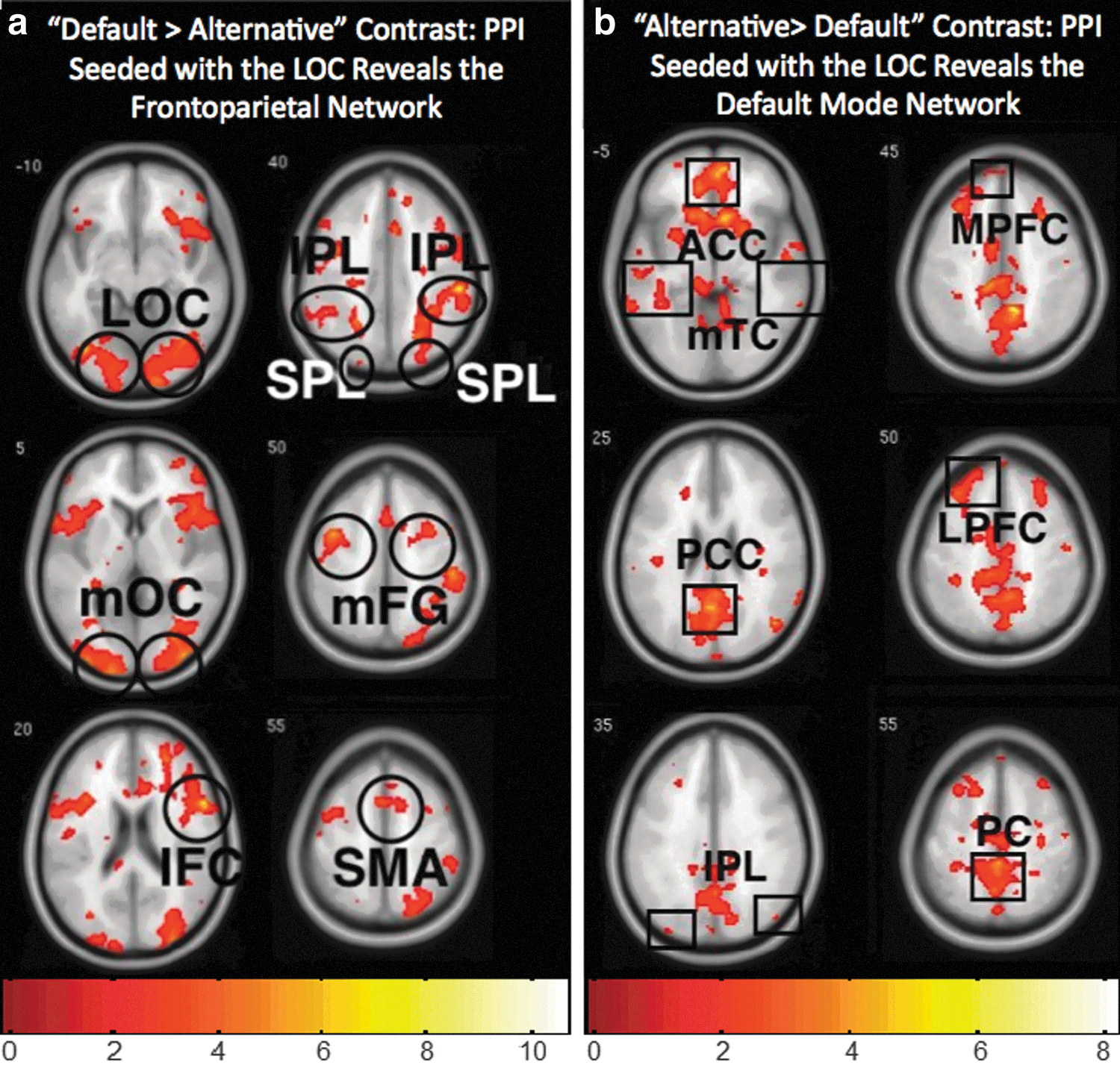
Functional connectivity (PPI) between the lateral occipital cortex (LOC) and regions where connectivity during one percept (as defined by the contrasts) exceeds connectivity in comparison to the other. The same global networks observed in the fMRI analysis (Fig 1) were also observed in the PPI analysis, however,
Connectivity between the FPN and DMN
In addition to the dynamic connectivity between the LOC and the two networks, the connectivity between the DMN and FPN was also measured using PPI analysis to investigate possible cross-network connectivity in association with connectivity with the incoming visual stream. During the default contrast, both the DMN (Supplementary Fig. S2a) and the FPN (Supplementary Fig. S2b) exhibited higher connectivity within their respective networks. Independent ROI analyses confirmed that the connectivity within each network was significantly greater during the default perspective (default>alternative, DMN t=4.45, p<0.05 and FPN t=6.58, p<0.05). During the alternative contrast, however, the two networks increased their connectivity to each other, such that the DMN was more connected to the FPN (Supplementary Fig. S3a), and the FPN was more connected to the DMN (Supplementary Fig. S3b) as shown by the PPI results. This cross-network connectivity that was observed most prominently during the alternative perspective was also confirmed by independent ROI analyses (alternative>default, DMN connectivity with FPN seed, t=3.63, p<0.05 and FPN connectivity with DMN seed t=5.33, p<0.05). In general, the PPI analysis indicates that during the default contrast, the individual networks tended to be more connected within themselves, whereas during the alternative percept, the cross-network functional connectivity was increased.
Effective connectivity
The PPI findings were also confirmed by a dynamic causal model (Penny et al., 2004), where effective connectivity between the LOC, DMN, and FPN was estimated during both conditions using a fully specified model. In accordance with our model, significant contrasts of connectivity parameters (Alternative>Default) were observed for the connectivity from the DMN to the LOC (t=1.91 and p<0.04), and from the FPN to the DMN (t=1.79 and p<0.05). Thus, these two approaches, PPI and DCM, provide convergent findings, indicating that during the alternative percept, the connection was increased between the LOC and the DMN, and also between the FPN and DMN.
Discussion
Differences in connectivity between the cortical regions have previously been reported depending upon volitional (top-down) goals (Chadick and Gazzaley, 2011), as well as interactions between the DMN and the FPN (Fox et al., 2005; Uddin et al., 2009). Here we extend these findings and show that volitional image segmentation tasks also engage distributed neural patterns consistent with the DMN and the FPN. Further, functional connectivity reveals a mechanism of oppositional coupling and decoupling between the incoming visual stream and these networks that is associated with the bistable percepts.
Recent EEG findings reporting that neural activity precedes the perceptual emergence of the hidden percept (Britz et al., 2009) are consistent with our finding that the nonengaged percept is associated with an active process correlated with the incoming visual stream. Further, previously proposed models for bistable perception suggest that fatigue or satiation of the neural correlates associated with conscious percept contribute to the emergence of the suppressed percept (Toppino and Long, 1987). Our data are also consistent with the notion that active stages of percept construction involve neuronal suppression between the levels of visual information processing. For example, our finding that the DMN and the FPN are more internally correlated during the default percept, that is, increased intranetwork connectivity, and more cross-correlated during the alternative, that is, increased internetwork connectivity, is consistent with the previously reported competitive and suppressive interactions between these networks (Kelly et al., 2008; Uddin et al., 2009).
A framework proposed by (Spreng et al., 2010) can be applied to our findings where a model of interactive top-down neural processes originating from the FPN mediates between the two networks (Fig. 3). During the default percept, the distributed BOLD response was consistent with DMN activity (the DMN was less deactivated during the default percept relative to the alternative percept), whereas during the alternative percept, the distributed BOLD response was consistent with FPN activity, indicating that when one network was active (Fig. 3- yellow), the other was relatively less active.
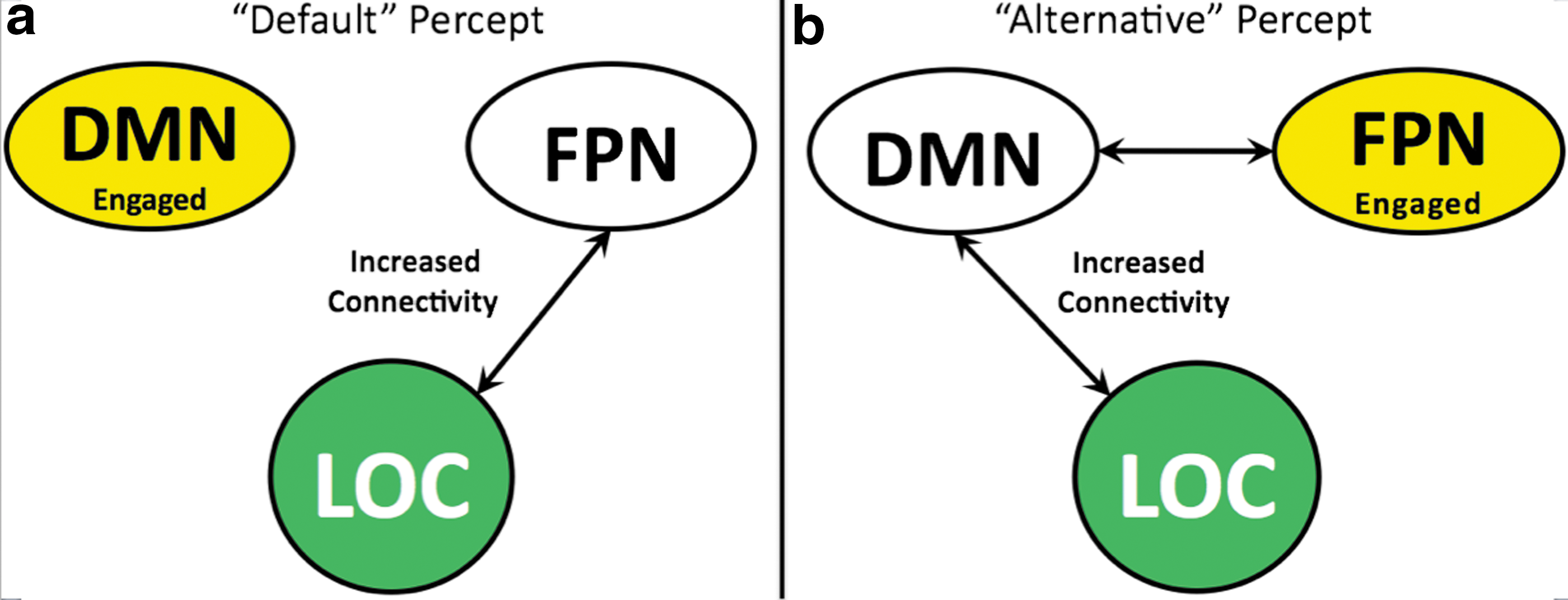
A conceptual summary of findings.
The functional connectivity between the bottom-up visual stream, originating from the LOC (Fig. 3–green), was highly correlated with the less-engaged network. Variations in concurrent deactivations of irrelevant sensory input have been associated with a suppressive mechanism (Amedi et al., 2005; Shmuel et al., 2002; Wade, 2002). Accordingly, our finding of the increased connectivity between the FPN and the deactivated DMN suggests that the FPN may suppress DMN activity during the alternative percept. Additionally, during the default percept, the less-engaged FPN was internally connected suggestive of a regulation of this suppressive mechanism. These findings lead to the novel interpretation that increased connectivity between the visual stream and the deactivated network reveals a suppressive mechanism associated with the conscious percept possibly mediated by oppositional long-range networks that interact with the incoming visual information (Fig. 3).
The discovery that the bottom-up visual stream was anticorrelated with the network associated with the ongoing conscious percept is surprising. However, working with the framework put forth, these new findings can be interpreted as reflecting a balance between the suppressive and excitatory interactions between the networks that are associated with the unconscious and conscious percepts and the bottom-up visual stream. Together, these findings are consistent with a model where active image segmentation, as observed in bistable figures, is mediated by top-down mechanisms that influence incoming visual information.
Conclusion
Bistable percepts provide a unique opportunity to investigate the neural mechanism of image segmentation, because a single visual figure gives rise to two mutually exclusive perceptual constructs. In this study, percepts elicited by the Schroder Staircase differentially gave rise to distributed patterns of neural activity consistent with the DMN and an FPN, during fMRI. In particular, the DMN was observed during the default percept, while the FPN was observed during the alternative percept. Additionally, the functional connectivity revealed that the incoming visual stream was more coupled with the DMN during the more effortful, alternative percept, and that the DMN and FPN were most interconnected during the alternative percept. These findings suggest that the process of binding image segments into perceptual units engages oppositional and interacting long-range neural networks.
Author Contributions
JH supervised the study. AK and DK performed the experiments. AK analyzed the activation data. SPP analyzed the functional connectivity data. JH and AK drafted the manuscript. XZ assisted with data analysis and provided technical advice.
Footnotes
Acknowledgments
Authors are grateful for significant contributions by students and subjects who have participated in the development of this project, including Grace Lai (Columbia University Program in Neuroscience), graduate and undergraduate educational support for science education, Macaulay Honors College [AK], and the Intel Science Competition for high school students [AK, DK]. Current funding for this project includes NIAAA-09-07 (NIH) HHSN27500900019C (subcontract to JH, PI Jon Morgenstern); a predoctoral fellowship (NRSA) F31MH088104-02 (SP, mentor: JH); U.S. Army RDECOM-TARDEC W56H2V-04-P-L (JH); and NIH RO1NS056274 (subcontract to JH, PI Nicholas Schiff ).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.