
Abstract
Recognizing emotional facial expressions is a part of perceptual decision-making processes in the brain. Arriving at a decision for the brain becomes more difficult when available sensory information is limited or ambiguous. We used clear and noisy pictures with happy and angry emotional expressions and asked 32 participants to categorize these pictures based on emotions. There were significant differences in behavioral accuracy and reaction time between the decisions of clear and noisy images. The functional magnetic resonance imaging activations showed that the inferior occipital gyrus (IOG), fusiform gyrus (FG), amygdala (AMG) and ventrolateral prefrontal cortex (VPFC) along with other regions were active during the perceptual decision-making process. Using dynamic causal modeling analysis, we obtained three important results. First, from Bayesian model selection (BMS) approach, we found that the feed-forward network activity was enhanced during the processing of clear and noisy happy faces more than during the processing of clear angry faces. The AMG mediated this feed-forward connectivity in processing of clear and noisy happy faces, whereas the AMG mediation was absent in case of clear angry faces. However, this network activity was enhanced in case of noisy angry faces. Second, connectivity parameters obtained from Bayesian model averaging (BMA) suggested that the forward connectivity dominated over the backward connectivity during such processes. Third, based on the BMA parameters, we found that the easier tasks modulated effective connectivity from IOG to FG, AMG, and VPFC more than the difficult tasks did. These findings suggest that both parallel and hierarchical brain processes are at work during perceptual decision-making of negative, positive, unambiguous and ambiguous emotional expressions, but the AMG-mediated feed-forward network plays a dominant role in such decisions.
Introduction
Making perceptual decisions about external stimuli depends on available sensory information and is known to involve a widely distributed cortical network. Identifying emotions from facial expressions is a part of such perceptual decision-making processes. These processes underlie coordinated neural activity among a number of cortical and subcortical brain regions, which along with the amygdala (AMG) includes the inferior occipital gyrus (IOG) that receives input from retina (Britton et al., 2006; Haxby et al., 2000), the fusiform gyrus (FG) that identifies each individual face (Fairhall and Ishai, 2007; Kanwisher et al., 1997) and the ventrolateral prefrontal cortex (VPFC) that evaluates emotional stimuli (Iidaka et al., 2001). Visual regions like the lateral occipital region plays a significant role, while processing emotional stimuli and providing inputs to the fusiform areas and the frontal gyrus (Haxby et al., 2000). The fusiform represents static features of faces, whereas temporal gyrus represents dynamic facial expressions like direction of gaze (Adolphs, 2002; Hoffman and Haxby, 2000). Pessoa and colleagues (2002) suggested that a critical pathway proceeds from primary visual cortex to AMG, including FG and superior temporal sulcus. Further, Fairhall and Ishai in 2007 examined directionality of the connection between FG and AMG using dynamic causal modeling (DCM). The FG activity was found to be influenced by AMG activation (Fairhall and Ishai, 2007). Herrington and colleagues (2011) suggested the bidirectional connectivity between AMG and FG during facial recognition. A few studies have used effective connectivity approaches like Granger causality, DCM, structural equation modeling to look at the role of the AMG-involved networks (Dima et al., 2011; Fairhall and Ishai, 2007; Foley et al., 2011; Herrington et al., 2011). In the current study, using DCM (Friston et al., 2003), we investigated how the AMG coordinates activity among various structures during decisions about happy and angry emotional faces, and how degraded facial contents modulate the coordinated activity.
Previous neuroimaging studies suggested the involvement of several brain areas, including the AMG during perceptual decision-making of emotions (Damasio et al., 1995; Habel et al., 2007; LaBar et al., 2003; Phan et al., 2004; Shin et al., 2005). Traditionally, the AMG is known to be associated with negative emotions, such as threat and fear (Adolphs et al., 1995; Fusar-Poli et al., 2009). Consistent AMG activation was observed during processing of negative emotions (anger, fear and threat) but not only specific to fear related events (Sander et al., 2003; Scott et al., 1997; Siegle et al., 2002). A study by Sander and colleagues (2003) mentioned that consistent involvement of a brain region during processing of certain stimuli does not specify the role of that particular region for those stimuli. However, the AMG activity is also related to positive events, such as processing of happy facial expressions (Breiter et al., 1996; Sergerie et al., 2008; Somerville et al., 2004; Williams et al., 2004), positive words (Hamann and Mao, 2002), pleasant tastes (Small, 2002) and reward (O'Doherty et al., 2002). Anatomically, the AMG is well placed that it receives direct thalamic projections from pulvinar and medial geniculate nucleus (Jones and Burton, 1976). It integrates exteroceptive as well as interoceptive stimuli and modulates sensory, motor and autonomic processing (Morris et al., 1998). Inputs to AMG through olfactory bulb, the nucleus of solitary tract and other subcortical structures are considered to provide information about the stimuli properties (Amaral et al., 1992; Morris et al., 1998). For ambiguous stimuli, depending on emotional content, the AMG biases top-down attentional signal, which may in turn help to increase or decrease the influence of sensory inputs promoting salience, vigilance, and attention (Davis and Whalen, 2001; Whalen, 1998). The AMG influence is preferentially invoked in ambiguous learning situations. It is now generally accepted that the AMG may respond to a wide range of stimuli regardless of positive or negative (Habel et al., 2007; Sander et al., 2003) and clear or ambiguous stimuli and it also acts in a functionally integrated way to act as a neuromodulator (Amaral et al., 1992; Morris et al., 1998).
Face perception is mediated by a distributed neural systems consisting of bilateral regions like IOG and FG. Its anatomical configuration suggests an organization where IOG provides input to FG and AMG plays main role in processing information gleaned from faces (Fairhall and Ishai, 2007; Haxby et al., 2000, 2002). In addition, it has also been suggested that ventral prefrontal cortex modulates activity of the AMG during evaluation of emotional facial stimuli. Using structural equation modeling, a functional association between right AMG and right prefrontal cortex has been observed during processing of angry facial expressions (Nomura et al., 2004). A significant negative influence from AMG to FG is also reported that might be related to the potentiation of AMG activity through a route, including prefrontal cortex and FG during positive as well as negative feedback projections (Amaral et al., 1992; Morris et al., 1998; Nomura et al., 2004; Tabert et al., 2001).
Bilinear and nonlinear approximation was used to analyze the effective connectivity during facial affect processing for angry, sad and fearful cases where the significant modulation from IOG to VPFC was observed for angry and fear expressions (Dima et al., 2011). The feed-forward connections were observed to dominate over feed-backward connections causing unidirectional flow of information from occipital to prefrontal areas. This along with other studies (Critchley et al., 2000; Lane et al., 1999; Vuilleumier et al., 2004) signifies the role of the prefrontal cortex in emotion processing. Our study involves emotional expressions- happy and angry with and without noise resulting into four different conditions. Involvement of two opposite emotions (happy and angry) in the task reflects the spectrum of emotions to some extent, whereas inclusion of noisy pictures is motivated by the idea to keep the participants attentive and to check whether ambiguity highlights any particular connectivity in the activated networks. Our hypotheses are (1) that a factor, such as noise in ambiguous stimuli makes the outcome uncertain and vigilance is required to discern the stimulus contents, and that (2) during the processing of faces and facial expressions, positive or negative and easier or difficult, emotional stimuli enhance the connectivity from the early sensory region to the prefrontal cortex.
We applied the bilinear approximation of DCM (Friston et al., 2003; Stephan et al., 2010) to functional magnetic resonance imaging (fMRI) data. In addition to the Bayesian model selection (BMS) approach in DCM, we also used Bayesian model averaging (BMA) approach (Friston, 2009; Stephan et al., 2009). Our aims were to understand and differentiate (i) effective connectivity during positive (happy) versus negative (angry) and clear versus noisy face and facial affect processing as we go from an easier task (recognizing emotions from clear images) to difficult one (emotions from noisy images), and (ii) how the intrinsic and modulatory connections differ among four (happy, angry, happy noisy, angry noisy) conditions.
Materials and Methods
Participants
Thirty-two healthy human volunteers from diverse social backgrounds (16 males, 16 females; mean age, 27.6 (±4.7) years) participated in the experiment. All participants had normal or were corrected to normal vision and were interviewed in person. All participants had clean and normal neurological history. None of them reported the use of medication known to affect any neurological function. A written informed consent was obtained from each participant before the experiment and all the participants were compensated for their participation. The Institutional Review Board for Georgia State University and Georgia Institute of Technology Center for Advanced Brain Imaging (CABI), Atlanta, GA, approved the experimental procedure.
Design and procedure
Stimuli
We used two sets of human face images (happy set and angry set) as stimuli, each of eight images (4 males and 4 females). All the presented pictures were downloaded from F.A.C.E. Training–an interactive training by Dr. Paul Ekman (
Tasks
The experimental tasks were divided into two sessions. In the first session, participants did behavioral tasks outside the MRI scanner where reaction time and accuracy were measured and considered into behavioral analysis. They were instructed to respond as soon and as correctly as possible. In the second session, participants did the tasks inside the scanner during fMRI data acquisition. The task paradigm is shown in Figure 1. They performed two functional runs; each of 674 sec long. The total number of trials was 80, that is, 20 trials for each condition. They were forced to respond only after a delay, so reaction times were not available inside the scanner. In both sessions, participants perceived emotional faces (clear and noisy) and indicated their decisions whether the face reflects happy or angry expressions. In a task trial, a stimulus (picture of a face, clear or noisy) was presented for 500 ms followed by 8 sec-long display of fixation cross, and a question mark was presented for 500 ms at the end of this 8 sec interval (Fig. 1). Participants were instructed to make and hold their decisions during 8 sec, and indicate their decisions by pressing either a left or right button on a response box only when a question mark appeared on the screen at the end of 8 sec. They were instructed to prepare and make their motor responses in the next 6 sec. Stimulus onset times were extracted for each stimulus and the timing sequence was convolved with hemodynamic response function (hrf). The presentation software (

Experiment design.
Behavioral experiments
The behavioral experiment outside the scanner consisted of only one behavioral run. It was done to assess the task-related behavioral performance accuracy and reaction (response) time (time between the stimulus onset and button press). All four conditions were presented 30 times, in a random order with a total number of 120 trials. Participants sitting in front of a computer were asked to perceive the emotions of the pictures displayed as quickly and as accurately as they could by clicking the right or left button on mouse. They indicated their decisions about the presented stimuli by clicking the right button for angry faces and clicking the left button for happy faces. They were instructed to press the space bar on the keyboard to go to the next trial. Participants repeated these tasks inside the scanner according to the scheme shown in Figure 1. The behavioral performance accuracy was calculated from these behavioral responses recorded inside and outside the scanner. Pairwise t-test was used to compare the performance accuracy and the response time outside the scanner for both happy and angry cases separately (Fig. 2). Inside the scanner, the accuracy rates were 97.88 (±1.07)% for happy clear, 72.50 (±3.44)% for happy noisy, 97.61 (±1.06)% for angry clear, and 80.04 (±2.34)% for angry noisy trials.
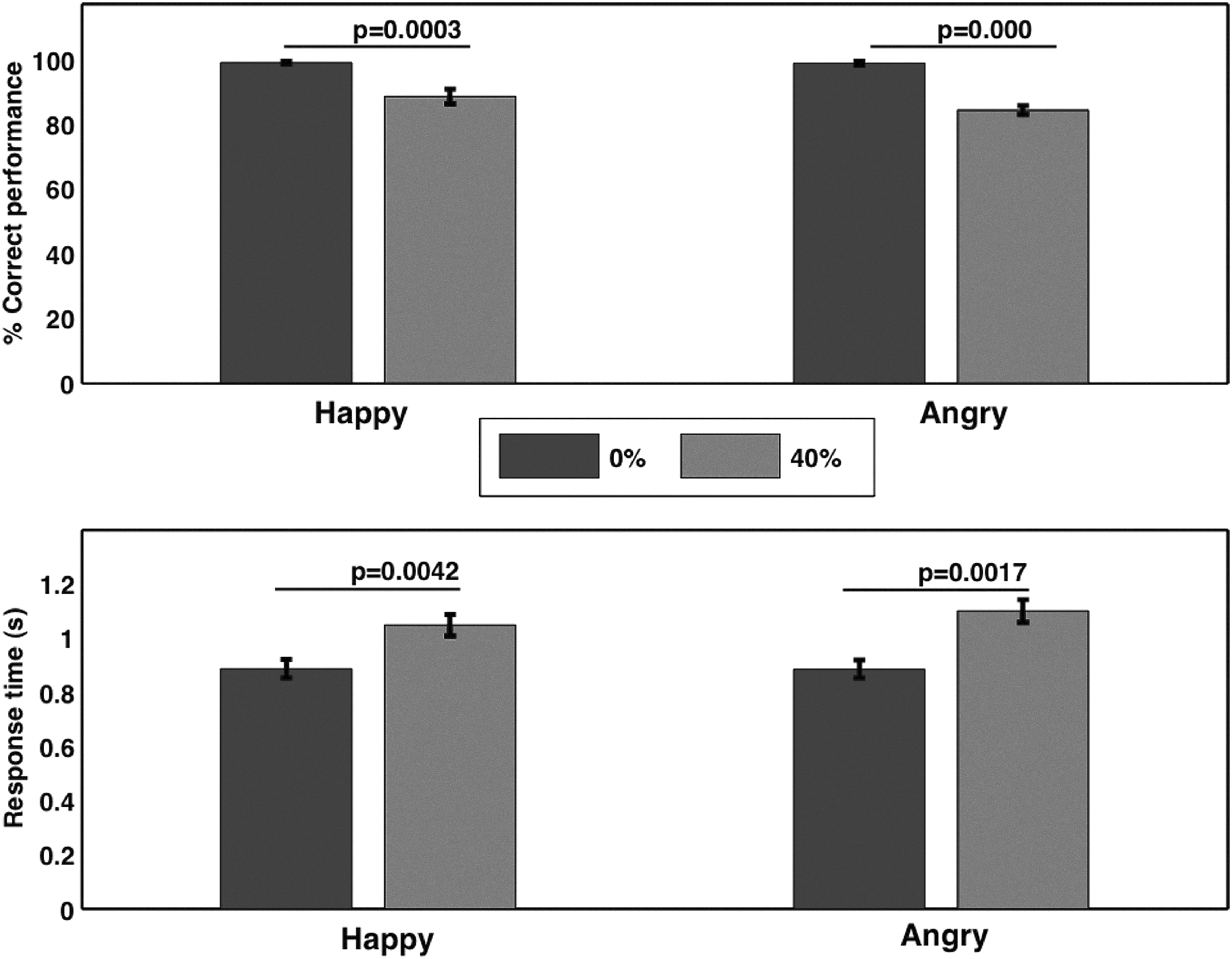
Behavioral response (outside the scanner). For both clear happy and clear angry emotional expressions, the behavioral accuracy significantly increased and the response time significantly decreased, whereas behavioral accuracy significantly decreased and the response time significantly increased with noise in the images.
Data acquisition and analysis
Image acquisition
The imaging was done on a 3-T Siemens MRI scanner, available at CABI (Georgia State and Georgia Tech CABI, Atlanta). After acquiring a high-resolution T1-weighted anatomical image, two whole-brain functional runs were performed with 337 scans in each run with the following parameters: echo-planar imaging, gradient recalled echo sequence; repetition time (TR)=2000 ms; echo time (TE)=30 ms; flip angle=90°; 68×68 matrix, voxel dimensions=3×3×3 mm3, 37 axial slices each of 3 mm thickness acquired in interleaved fashion with orientation parallel to the anterior-posterior commissural line for the measurement of the T2*-weighted blood oxygenation level-dependent effect. High-resolution anatomical T1-weighted images were acquired for anatomical references using an magnetization-prepared rapid acquisition gradient echo (MPRAGE) sequence with TR=2250 ms; TE=4.18 ms; inversion time=900 ms, flip angle=9° and voxel dimensions=1×1×1 mm3.
Conventional image analysis
fMRI data was preprocessed using statistical parametric mapping software (SPM8) (Wellcome Department of Cognitive Neurology, London, UK,
First level (within-participant) activation analysis
For each participant and each run, data collected from the experiment was modeled with a general linear model (GLM). Stimulus onset times in each run corresponding to the correctly identified happy and angry faces (clear and noisy) were convolved with canonical hrf. Total 15 parameters with two parameters (one correct and one incorrect) for each of the four conditions, one for rest period and six motion parameters were included in GLM analysis. Here six motion parameters were entered as nuisance covariates and were regressed out of the data. Two rest durations, each of 30 sec from the beginning and the end of each functional run were used as contrast to obtain images of brain activations associated with each condition in each participant.
Second level (between-participant) activation analysis
The individual contrast images generated in the first-level analysis were then entered into a second-level analysis. We used one-sample t tests to investigate the main effect of task (correctly identified emotional faces>rest period) for face and facial affect processing. We did not include the incorrect trials in the activation analysis. The resulting summary statistical maps were then subjected to a t-threshold at p<0.005 (false discovery rate [FDR] correction for multiple comparisons) and at cluster threshold of k>10 voxels. FDR correction controls the expected proportion of incorrectly rejected null hypotheses (“false discoveries”). These maps were overlaid on a high-resolution structural image in MNI orientation for displaying fMRI activations. Further, we combined all the four conditions (happy clear, happy noisy, angry clear and angry noisy) to consider a set of activated regions as regions/volumes of interest (ROIs/VOIs).
Volumes of interest
VOIs were defined in SPM8 using the first eigen-variate of activations within a sphere of radius 6 mm in MNI co-ordinate system. VOIs comprised four regions-IOG (39, −73, −8), FG (33, −43, −23), AMG (24, −1, −26) and inferior frontal gyrus within the VPFC (45, 5, 28). The co-ordinates were obtained from the second level analysis at p<0.005 (FDR) using xjView8 (
Brain Activations Associated with the Perceptual Decisions of Emotional Faces (32 Participants)
p < 10−5 (FDR corrected).
p < 10−3 (FDR corrected).
MNI, Montreal Neurological Institute; FDR, false discovery rate.
Dynamic causal modeling
DCM (Friston et al., 2003) treats the brain as nonlinear but dynamic system. It differs from other approaches like structural equation modeling (Büchel and Friston, 1997), which is based on the assumptions of interactions being linear, inputs unknown and without any external perturbations.
The aim of DCM approach is to estimate the connectivity structure among functionally distinct connected brain areas and the intrinsic parameters influenced by external experimental perturbations (Friston et al., 2003; Stephan et al., 2010). This is done by defining and constructing different models where each model has specific intrinsic connections that are modulated by different external changes. Best model that explains the observed data is obtained when all the models are supplemented with corresponding forward model of how the activity is transformed into a measured response. Hence, it is a BMS procedure that determines how the data is generated from the models (Friston et al., 2003; Penny et al., 2004; Stephan et al., 2009). The DCM involves the use of a set of ordinary differential equations. There are three sets of estimated parameters, including (1) direct influence of an external input on a region, (2) intrinsic connectivity between regions representing influence without any external perturbation, and (3) modulation affects representing changes in intrinsic connection strength induced by external experimental perturbation (Friston et al., 2003).
In this DCM study, we defined a network consisting of four areas that included-IOG, FG, AMG and VPFC with bidirectional connections among each other as a base model (Fig. 3a). Base model was further elaborated using external experimental changes as a function of emotional contents: happy or angry emotional expressions with or without noise, represented by ‘face with emotional expressions’ as shown in Figure 3b. We had a set of four bilinear families of the four conditions (happy-without noise, angry-without noise, happy-with noise and angry-with noise), each family having seven models. Each model included all the possible combinations of how facial as well as facial affect could modulate forward connections to VPFC. Models were distinguishable based on the modulations of the intrinsic connections. Bidirectional connections along with modulatory effects were tested for each of the four conditions to check if the information passed to VPFC was direct or mediated. We limited the number of models defined by bilinear approximation (Friston et al., 2003). We did not consider nonlinear approach and any other complex models since simple models are preferred to complex models (Friston, 2011; Penny et al., 2004) to maintain the balance between accuracy and complexity (Dima et al., 2011; Herrington et al., 2011; Li et al., 2010; Pitt and Myung, 2002; Stephan et al., 2010).

Defining models.
BMS and BMA
Exceedance probability is the measure used to compare posterior probabilities of different models. It is a degree of belief about a model having higher posterior probability than other models (Stephan et al., 2009; Wasserman, 2000). In our study, we use RFX BMS (random effects BMS) available in DCM12 to compute expected and exceedance probabilities of each model that gives us the optimal model among all seven models for each condition. Here we assume the model structure to be identical across participants. Further, we obtain BMA parameters by averaging over optimal models. For computational efficiency, this employs Occam's window, which discards all the models with probability ratio <0.05 compared to the best model (Penny et al., 2010; Stephan et al., 2010). Further t-tests are applied over the parameters obtained from all the optimal models over all the participants.
Results
Behavioral responses
Accuracy rates and reaction times were computed from the behavioral experiments done outside the scanner. Participants identified happy and angry emotions with an accuracy of 99.06 (±0.42)% and 98.85 (±0.53)%, respectively for clear images and 88.46 (±2.31)% and 84.36 (±1.43)%, respectively for noisy images. Reaction times during identification of happy and angry emotions were 0.88 (±0.03) sec and 0.88 (±0.03) sec, respectively for clear images and 1.04 (±0.04) sec and 1.10 (±0.04) sec, respectively for noisy images. Behavioral accuracy decreased and the reaction time (response time) increased significantly with noise (Fig. 2). The noise-degraded performance and increased reaction time showed that the task became more difficult with noise in those images.
Brain activations
Table 1 shows all the significantly activated brain areas in correct trials for two emotions happy (clear and noisy) and angry (clear and noisy) relative to the rest period at significance threshold of p<0.005 (FDR correction) and at cluster threshold of k>10 voxels. A set of significantly and commonly activated regions associated with emotional faces versus rest is the inferior frontal gyrus within VPFC, FG, AMG, and IOG as shown in Figure 4a. The contrast of noisy (happy and angry) versus clear (happy and angry) emotions shows activations in the insula, intraparietal lobule and supplementary eye field as shown in Figure 4b. These activations are consistent with the previous study by Heekeren and colleagues in 2004 (Heekeren et al., 2004).
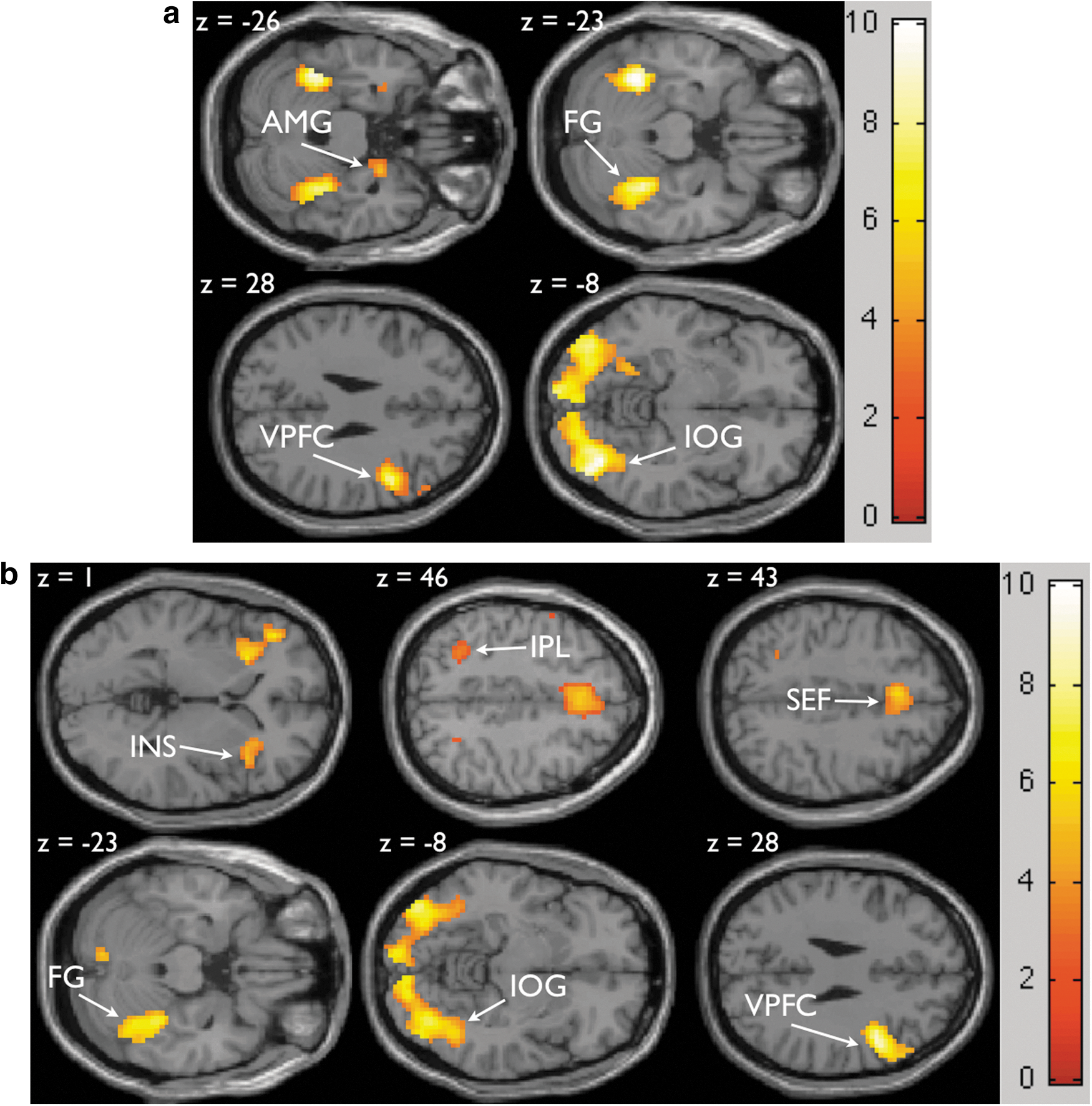
Task related brain activations from in-group (32 participants) analysis of correct trials for
DCM results
Optimal model selection
Comparing the exceedance probabilities of all the predefined seven models of bilinear family for each condition identifies model 4 as the best model for both happy clear and happy noisy facial expressions with an exceedance probability of 47% and 29%, respectively. Model 5 turns out to be the best model for angry clear with an exceedance probability of 32% and model 4 for angry noisy faces with an exceedance probability of 23% (Fig. 5). Both models-model 4 and model 5 involve the bidirectional connections among four regions with the modulations of the forward connections but no modulation from AMG to the VPFC in model 5.
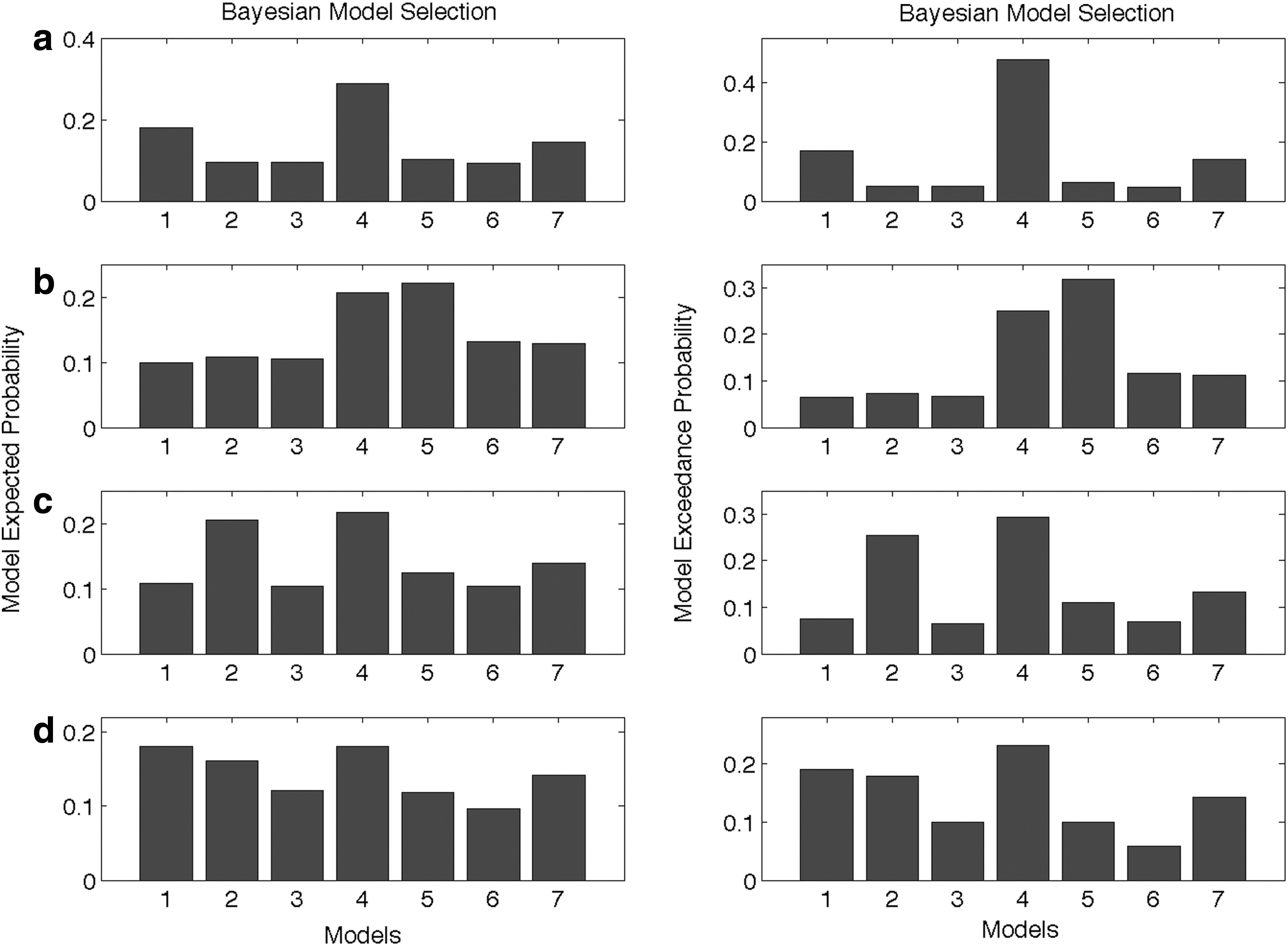
Expected probability and exceedance probability for 7 models during
Bayesian parameters and significance test
Since the optimal model remained the same, as the task becomes difficult in case of happy expressions but not for angry expressions, we obtain the parameters by averaging over optimal models. From intrinsic parameters obtained from BMA, we find that the connections from IOG to FG, AMG and VPFC dominate over all other connections for clear happy case (Table 2a, Fig. 6a), whereas connections from IOG to FG and AMG dominate over all other connections for clear angry case (Table 2a, Fig. 6c). As the task becomes difficult, significant intrinsic connections either become weaker or the strength remains the same in comparison to the easier task in case of both noisy happy (Fig. 6b) and noisy angry (Fig. 6c) facial affect processing (Table 2b). These BMA parameters are summarized in Table 2.
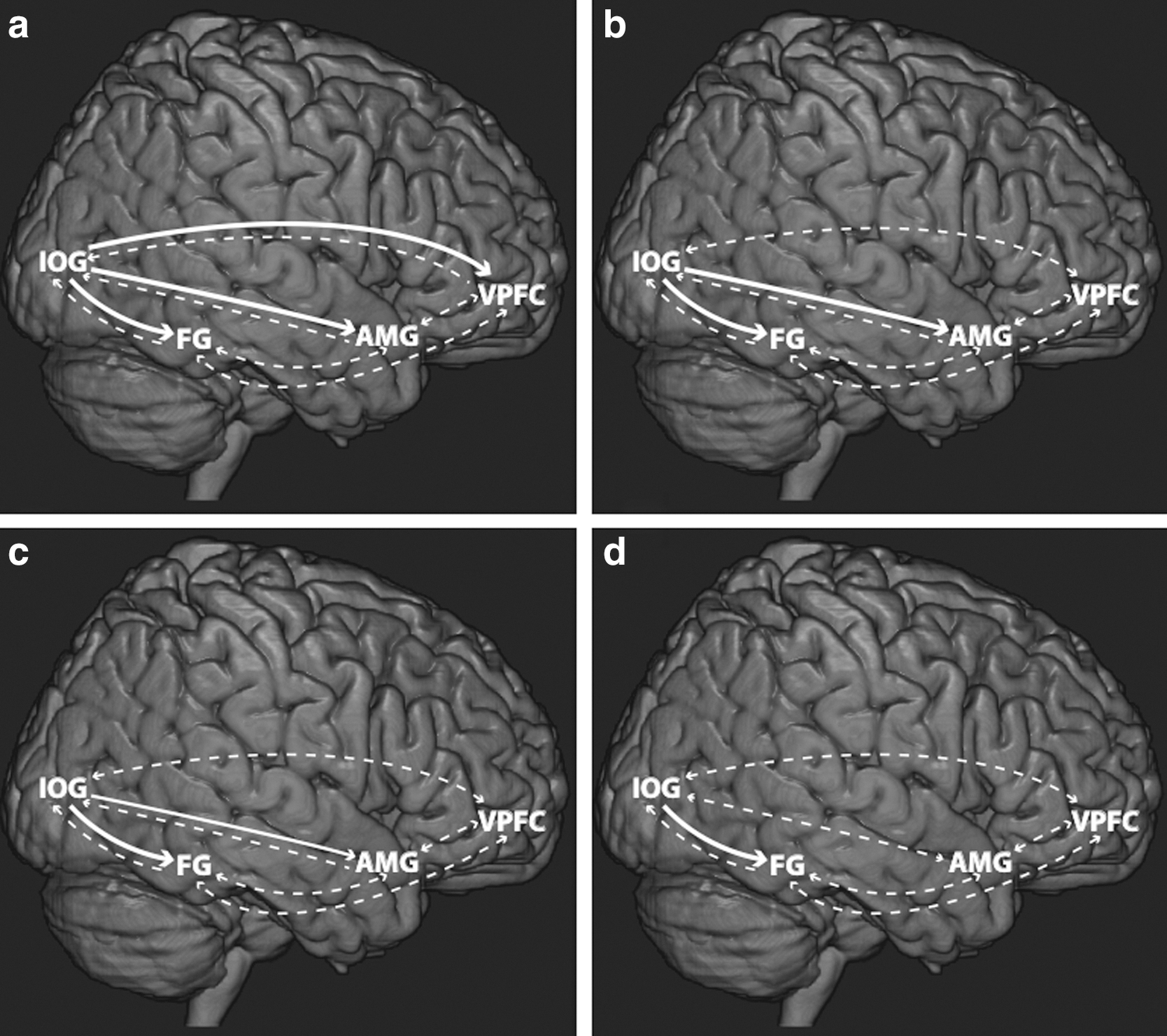
Effective connectivity network resulting from Bayesian model averaging over all the participants (N=30) and over all the models considered (n=7) during facial affect processing for
Dynamic Causal Modeling Intrinsic Connectivity Parameters Across All Possible Connections and All the Models from Bayesian Model Averaging for Clear Happy and Clear Angry Cases
Hc, clear happy; Ac, clear angry; SD, standard deviation.
Dynamic Causal Modeling Intrinsic Connectivity Parameters Across All Possible Connections and All the Models from Bayesian Model Averaging for Noisy Happy and Noisy Angry Cases
Hn, noisy happy; An, noisy angry.
Discussion
In this study, we employed SPM8 (Wellcome Department of Cognitive Neurology, London, UK,
The fusiform area is known to be active during recognizing facial expressions (Kanwisher et al., 1999; Kanwisher and Yovel, 2006). It is also involved in discriminating individual identity (Kanwisher et al., 1997; Kanwisher and Yovel, 2006). Anatomically, the AMG sends projections to the prefrontal cortex (Kim and Whalen, 2009; Kim et al., 2011). Many studies of brain activity (Dima et al., 2011; Dolan, 2002; Foley et al., 2011; Ohman, 2002; Pessoa and Adolphs, 2010) are consistent with the AMG-prefrontal cortex connectivity. Perception of emotions is known to recruit the AMG (Dolan, 2002; Ohman, 2002; Pessoa et al., 2006; Vytal and Hamann, 2010).
The role of prefrontal cortex is not specific to a particular task (Banich et al., 2000; Miller and Cohen, 2001). It is known to send a biasing signal to lower order regions (Davis and Whalen, 2001; Ledoux, 1998; Miller and Cohen, 2001). We find that it responds to the information received from different brain areas involved during the processing of faces with emotional content. It is activated during perceptual decision making when stimuli are ambiguous. These findings are consistent with previous studies (Gur et al., 2002; Heekeren et al., 2008; Miller and Cohen, 2001; Noppeney et al., 2010; Summerfield et al., 2006). A study by Miller and Cohen (2001) report that the prefrontal cortex is an important component for emotional evaluation and when the sensory information is available in excess, for example, to decide among multiple possibilities, to curtail confusions and when the task performed is weak or rapidly changing. Consistent with this, we find that the connectivity from the IOG and FG to VPFC is modulated during processing of faces with ambiguous emotional content.
In our study, pictures with positive (happy) emotions activate the AMG-mediated pathways to the prefrontal cortex. The attention in this case is either the emotional content present in the faces or the ambiguity of presented pictures (Miller and Cohen, 2001). Thus, the involvement of AMG during facial affect processing may be because of demand for sufficient attentional resources, which are required more in case of noisy stimuli than in clear stimuli for both happy and angry cases. Davis and Whalen (2001) suggested that the AMG is involved for increasing vigilance and attention when stimuli are ambiguous. Facial expressions that are ambiguous and behaviorally relevant activate AMG (Sander et al., 2003; Wright et al., 2002; Yang et al., 2002). Consistent with these results, our results show that the AMG-mediated network is involved during processing of noisy happy and noisy angry facial expressions. A contrasting possibility is that the task difficulty may also divert the participant's attention and contribution to avoid the stimulus. But it has been found in a study of threat related processes that the AMG activation does not get reduced when attention to faces was reduced (Anderson et al., 2003). Along with this, coupling of IOG and FG with prefrontal areas was also increased during all the four conditions representing facial and facial affect processing. Attention enhances the functional association and neural synchrony between prefrontal and visual cortices at a wide range of frequency (Gregoriou et al., 2009). Emotional stimuli capture additional attention (Vuilleumier, 2005; Vuilleumier and Driver, 2007), which may ultimately influence the feed-forward sweep of sensory inputs. Since attention gates the visual processing (Moran and Desimone, 1985), the connectivity from IOG to VPFC gets modulated because of visual attention where attention toward emotional content may further strengthen the connectivity. Fusiform area is also well known to respond strongly to a wide variety of face stimuli than objects and it extracts perceptual information to recognize them (Kanwisher et al., 1997; Kanwisher and Yovel, 2006).
The connectivity from AMG to VPFC was not modulated in case of clear angry since this did not involve any ambiguity or complex facial expressions. AMG-mediated network was found to be involved during processing of positive emotions like clear happy even in absence of any ambiguity or complex expressions. This is consistent with previous findings on processing of positive emotions like happy facial expressions as well as reward conditions in animals and humans (Breiter et al., 1996; Spiegler and Mishkin, 1981; Williams et al., 2005) but not during clear angry facial expressions, which demands immediate action but engages VPFC to avert potential overreaction (Dima et al., 2011; Vytal and Hamann, 2010). Thus, the functional significance of the AMG can be broadly understood for the detection of emotions (Morris et al., 1999; Rolls, 1999). These emotions can be basic as well as complex processed by a common set of neural systems (Sander and Koenig, 2002). It has a key role to modulate the information, to process the ambiguity in the presented tasks, to predict the unpredictability in the tasks, and to significantly highlight the processing of activated network. Agren and colleagues (2012) have confirmed the modulatory role of AMG in a study of fear related network. Similar studies, for example, a study by Adolphs and colleagues (1999) on processing of negative emotions like fear and anger for a rare participant with bilateral damaged AMG have shown the partial ability to recognize valence, and inability to recognize arousal (Dolan, 2002). A study by Dolan in 2002 also suggests that the psychological states of emotions are embodied as automatic arousal fashion (Dolan, 2002). In a face-processing study of fearful faces, Vuilleumier and colleagues in 2004 report that lesions in AMG can change the functional pattern of activations for the regions that are distant from the AMG implying connectivity with the AMG (Vuilleumier et al., 2004). Anatomical and functional structure of AMG also suggests that it is designed to act as a relevance detector. It consists of 13 heterogeneous nuclei that do not result into one functional system (Sander et al., 2003; Swanson and Petrovich, 1998) but integrates multiple functions within itself. It may be inherently structured to process perception of various emotions, emotional expressions and uncertainty in emotions.
Our BMA results, obtained by averaging the parameters over optimal models, show that the feed-forward connectivity dominates over feed-backward connectivity. This implies that backward connectivity plays a definite but minor role in comparison to the forward connectivity.
To summarize, our BMS results from the DCM analysis revealed that the AMG-mediated feed-forward network is enhanced during perceptual decision-making processes when clear and noisy happy faces are presented. It is also enhanced in case of noisy angry faces, but the AMG-mediated network was not modulated in case of clear angry expression. This suggests that the task difficulty leads to an overall weaker forward connectivity but involves the AMG-mediated network for both positive and negative emotions. BMA results also suggest that the network activity strength depends upon the type of emotions and the available information content about the emotion. BMA results also show that the feed-forward connections dominate over feed-backward connections during perceptual decision-making of emotional expressions.
Footnotes
Acknowledgments
An NSF CAREER Award (BCS 0955037) to M. Dhamala supported this work. We would like to thank Dr. Karl Friston, Dr. William Penny, Dr. Guillaume Flandin, Dr. Klaas E. Stephan, and Dr. Danai Dima for their replies to our questions on numerous occasions and for providing us the latest version of DCM (DCM12). We would also like to thank Eli Goshorn for writing Matlab programs to make noisy pictures and Crystal Smith for writing Presentation programs for stimuli.
Author Disclosure Statement
There is no conflict of interest for any of the authors.