
Abstract
The decade-long endeavor to link brain structure with brain function has met with varying degrees of success. Recent advances in imaging techniques allow a fresh look at the issue, however. In the current investigation, functional magnetic resonance imaging and diffusion tensor imaging (DTI) were employed to directly investigate the link between functional connectivity during a verb generation task and the underlying structural substrate. Nineteen participants performed a verb generation task during functional scanning and also underwent DTI. A psychophysiological interaction analysis was used to map the functional networks recruited during the task and as an index of functional connectivity. The degree to which this correlated with diffusion measures was used to explore the existence of anatomofunctional relationships within the identified connections. A significant correlation was seen for the middle frontal (MFG) to precentral gyrus pathway, where a pattern of low fractional anisotropy and high perpendicular diffusion was associated with low functional connectivity. A second pathway between the MFG and inferior temporal gyrus did not show a significant correlation, which may be attributed to two independent factors that might be influencing the structural properties of this pathway. The variation in structure function relationships within this network may relate to each pathways involvement in different cognitive functions.
Introduction
The relationship between the structure and function of the brain still remains a fundamental question in neuroscience. However, recent advances in neuroimaging have enabled the exploration of anatomofunctional relationships in greater depth. This has been seen in studies of language networks in particular (e.g., Catani and Thiebaut de Schotten, 2008; Catani et al., 2005, 2007; Glasser and Rilling, 2008; Nucifora et al., 2005; Powell et al., 2006, 2007; Saur et al., 2008). For example, the superior longitudinal fasciculus (SLF) was suggested to be more structurally asymmetrical [as measured by fractional anisotropy (FA)], the more strongly the participants were functionally lateralized during language tasks (Powell et al., 2006). Using similar methods, Saur and colleagues (2008) identified white matter pathways that followed both a dorsal as well as a lesser known ventral route to connect the frontal and temporal language regions. They suggested that the dorsal and ventral pathways are thought to be involved in the mapping of sound to articulation and sound to meaning, respectively. These approaches, however, are limited by the fact that comparisons are based on group average data, resulting in a more indirect and general structure–function relationship. In a recent comprehensive evaluation of methods used to assess functional connectivity, Smith and colleagues (2011) cautioned the use of functionally inaccurate regions of interest (ROI; such as those from structural atlases), as they can lead to unreliable and erroneous estimations of a functional network. Instead, individual subject-based ROIs can be used to minimize the confounding effects of anatomical variability across subjects, and thus be more certain that we are investigating the same functional regions across individuals (Fedorenko et al., 2010). This is particularly important for cortical regions where the identification of anatomical boundaries remains a challenge. In such cases, a functional landmark approach can be employed whereby patterns of functional response are used to locate the same functional region across individuals (Saxe et al., 2006). Therefore, we sought to employ a functional landmark approach for defining anatomical regions in each participant, in an endeavor to map a specific network in terms of both function and structure.
Recently, structural connectivity has been measured noninvasively with diffusion tensor imaging (DTI) and tractography. Most commonly, the measure of FA has been reported as an index of the degree of white matter integrity. Far less common are studies using the measures of parallel and perpendicular diffusion. Whereas we have yet to ascertain the exact underpinning of these indices, they can provide useful information regarding the possible cytoarchitecture of the white matter pathways as has been previously illustrated (Iwabuchi et al., 2011). Exploring these measures (including FA) might offer a more detailed, although complex, description of the association between the structure and function of a task-related network.
Assessing functional connectivity on the other hand, appears to be a more complex endeavor. Various approaches to quantify connectivity have been developed, including dynamic causal modeling (DCM) (Friston et al., 2003), structural equation modeling (SEM) (McIntosh and Gonzalez-Lima, 1994), and psychophysiological interaction (PPI) (Friston et al., 1997), each with their own limitations. In the current study, an exploratory and data-driven approach was preferred over one that is model driven (as in DCM and SEM); therefore, functional connectivity was investigated using a PPI analysis. The blood-oxygen-level-dependent time series for a given seed region is correlated with the activity within the whole brain for a given cognitive task. Any area that shows correlating activity is deemed to be functionally connected. Using this method, combined with probabilistic tractography, it is possible to explore the neuroanatomical basis of a network and the way in which this may relate to the functional coupling of these brain regions.
Taken together, our main objective of the current study is to provide a more in-depth analysis of white matter structure of a specific task-related pathway identified using the verb generation task, and investigate to what degree functional connectivity corresponds with these measures.
Materials and Methods
Participants
Twenty-one healthy right-handed participants (11 males) with no known history of neurological disease participated in the study. The age of participants ranged from 19 to 37 years (mean age 26.05 years, standard deviation 4.9). All participants provided informed consent with approval from the University of Auckland Human Participants Ethics Committee and had normal or corrected-to-normal vision.
Scanning procedure
Functional, diffusion-weighted and T1-weighted structural images were acquired on a 1.5 Tesla Siemens Avanto scanner (Erlangen, Germany) in a single session per participant. Total acquisition time was ∼50 min. Functional magnetic resonance imaging (fMRI) data were analyzed using SPM8 (Wellcome Department of Imaging Neuroscience, London, United Kingdom;
Experimental paradigm
The verb generation task was programmed using E-Prime software (
fMRI data acquisition and preprocessing
For each participant, T2*-weighted echo-planar imaging (EPI) scans were acquired, which included two dummy scans to allow for signal saturation, parallel to the AC/PC line using the following parameters: repetition time (TR)=2400 ms; echo time (TE)=30 ms; field of view (FOV)=192 mm2; 35 axial slices; matrix size=64×64 mm; and voxel size=3×3×3 mm. A total of 270 T2*-weighted volumes were acquired during the verb generation task. T1-weighted structural volumes were also acquired for each participant using a 3D magnetization prepared rapidly acquired gradient echo sequence with the following parameters: TR=11 ms; TE=4.94 ms; FOV=256 mm2; 176 sagittal slices; matrix size=256×256 mm; voxel size=1×1×1 mm. The data were processed using SPM8 software following the standard preprocessing protocol (realignment, coregistration, normalization, and smoothing). Realignment involved realigning the first volume of each session with the rest of the volumes and also generating a mean of the functional volumes. This mean of the functional scans was used for coregistration of the T1-weighted structural scan. Both structural and functional images were normalized to the standard Montreal Neurological Institute (MNI) space, and functional images were spatially smoothed used a Gaussian filter of 9×9×9 mm at full-width half maximum. First-level analyses were performed for each participant using the general linear model (GLM). The conditions were modeled as a boxcar function and convolved with a canonical hemodynamic response function. Movement regressors were also included in the model. One-sample t-tests were used on verb generation versus noun repeat and verb generation versus X words contrast images in a second-level random-effects analyses to see the general patterns of activation during the language task. To correct for multiple comparisons, a familywise error correction at the cluster level was applied at p<0.05 with a cluster extent threshold of 30 voxels. The lower level contrast (verb generation vs. X words) was used to ensure that typical language regions were activated. However, since our interest was focused on targeting a more functionally specific network, we decided to use the verb generation versus noun repeat contrast for the PPI analyses. All voxel coordinates reported here were transformed from the MNI space to Talairach and Tournoux space (Talairach and Tournoux, 1988) using nonlinear transformations.
DTI data acquisition and analysis
Data were acquired on a 1.5 Tesla Siemens Avanto scanner (Erlangen, Germany) using a single shot spin-EPI sequence (45 slices; TR=6600 ms; TE=101; FOV=230 mm2; matrix size=128×128 mm; and voxel size=1.8×1.8×3 mm) in 30 noncollinear directions with a diffusion weighting of b=1000 sec/mm−2 with one scan at b=0 sec/mm−2. The sequence was applied three times and averaged to increase the signal to noise ratio. Data were analyzed using tools from FSL's (Behrens et al., 2003; Woolrich et al., 2009) Diffusion Toolbox (FDT, version 2.0). For each subject, the eddy current correction tool was used to correct for eddy current distortions and motion artefacts. Diffusion-weighted images were registered to a standard MNI brain image using the FMRIB Linear Image Registration Tool (FLIRT). FA, mean diffusion (MD) and eigenvector maps were computed, which were then transformed to standard space using the FMRIB Non-Linear Image Registration Tool (FNIRT). The data were then run through the program BEDPOSTX to build probability distributions on diffusion parameters and model for crossing fibers at each voxel (Behrens et al., 2007), in preparation for tractography.
The Freesurfer image analysis suite (
Psychophysiological interaction
A PPI analysis (Friston et al., 1997) was conducted to investigate the influence of language areas on other regions across the brain. The GLM analysis for the verb generation>noun contrast showed a large activation cluster at Talairach and Tournoux coordinates −46 21 25 [left middle frontal gyrus (MFG), Brodmann area (BA) 9]. This is shown in Figure 1. A spherical seed ROI with a radius of 10 mm was created for each participant by selecting the peak maximum from the verb generation>noun contrast that was located closest to the group coordinate above. The time course was extracted for the voxels that were significant within each ROI, and subjected to a PPI analysis to find regions that showed a higher correlation with the ROI in the language condition (i.e., verb generation) compared with the control condition (i.e., repeating nouns). A Monte Carlo simulation was employed for the PPI group analyses to determine a minimum cluster extent of 33 voxels (at a voxelwise threshold of p<0.001) to achieve an alpha level of p<0.01, corrected for multiple comparisons. The PPI analyses showed an interaction between the seed region (left MFG) and the left inferior temporal gyrus (ITG; BA 37; peak at −43 −41 −8) and also the left precentral gyrus (BA 4; peak at −52 −4 44; Fig. 2). Maxima for the functional activation and the PPI analyses are reported in Table 1. For each participant's PPI analysis, spherical ROIs with a radius of 10 mm were generated by locating activation peaks closest to the significant group PPI coordinates. Mean parameter estimates were then extracted from each of the connected regions. Parameter estimates provide a measure of the strength of effective connectivity or functional coupling of brain regions. All ROIs generated were binarized to be used as masks for the tractography component.
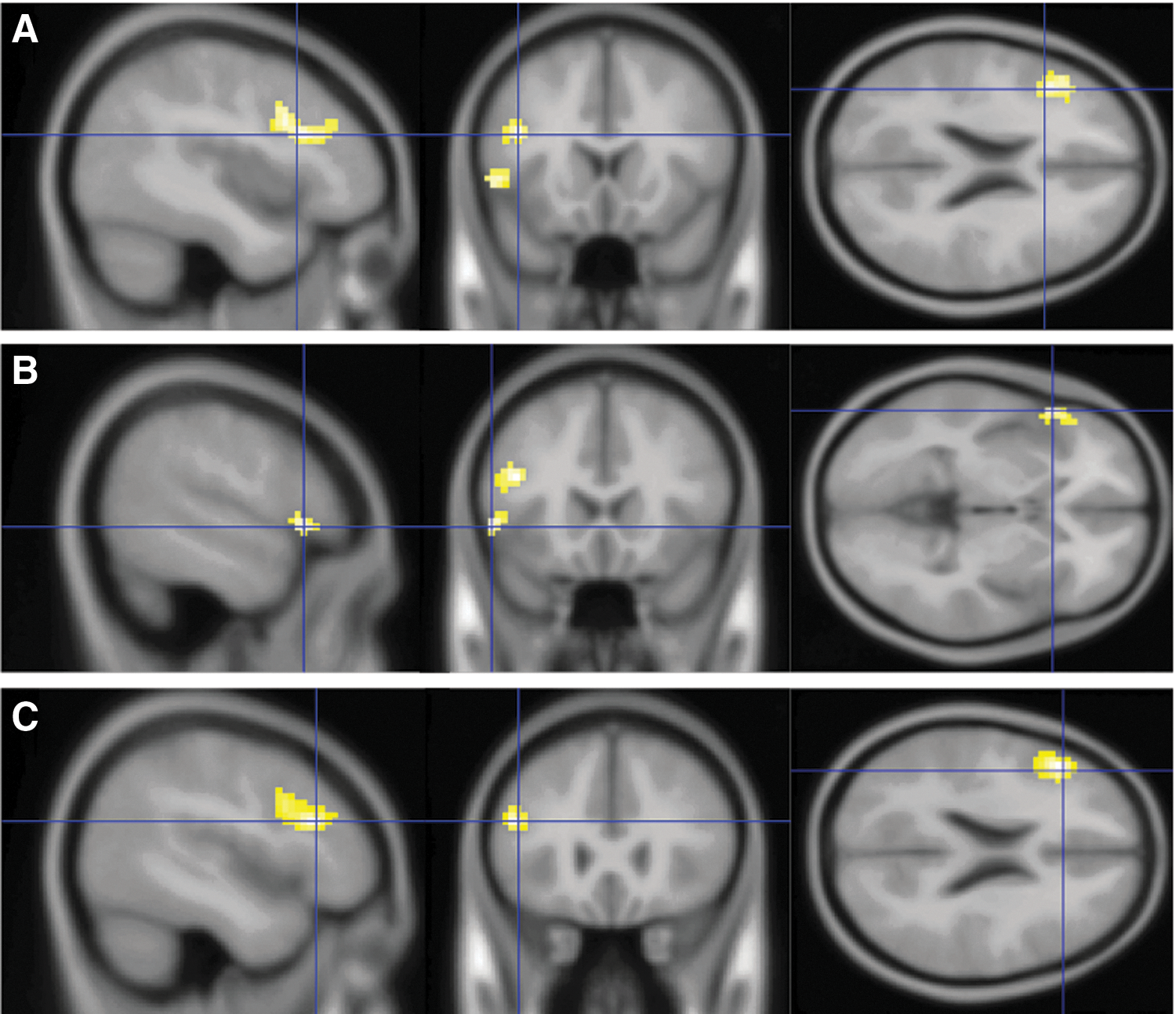
Group activations for the verb generation task.
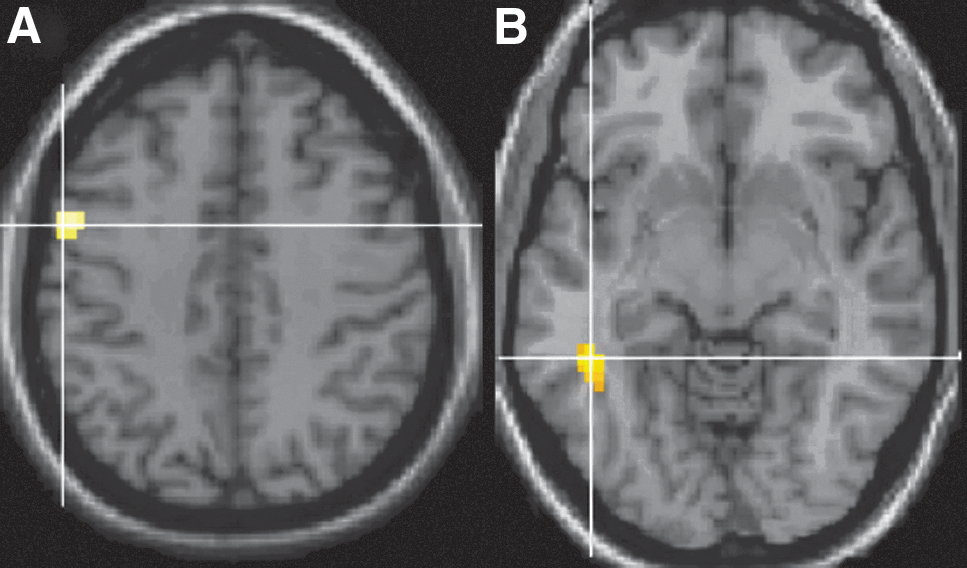
Significant activation from psychophysiological interaction (PPI) analyses. Regions showing correlating activity with the seed MFG region [
Functional Activation and PPI Analysis Results for the Verb Generation Task
Activation peaks with their locations, coordinates in Talairach and Tournoux (1988) atlas space and Z scores. BA is the Brodmann area nearest to the coordinate and should be considered approximate. All regions from GLM analysis were significant at p<0.05, FWE corrected, cluster threshold of 30 voxels. PPI regions were significant at p<0.01, corrected using Monte Carlo simulation for voxelwise threshold of 33 voxels.
FWE, familywise error; GLM, general linear model; ITG, inferior temporal gyrus; MFG, middle frontal gyrus; PPI, psychophysiological interaction.
Tractography
The masks produced from the fMRI analyses were used as seed and waypoint masks for probabilistic tractography (Behrens et al., 2003). Specifically, the functional activation masks were used to seed tractography, while the masks derived from the PPI analyses were used as waypoint masks. To minimize the risk of generating spurious pathways that cross over gyri, the seed masks were projected onto a Freesurfer surface at the gray matter–white matter boundary to create surface labels, thus, allowing tractography to be run from surface vertices (rather than voxels) toward the brain. Tractography was seeded from the left MFG, with left ITG and left precentral gyrus masks as separate waypoints to generate two pathways for each participant. Fiber tracking drew 5000 samples from each seed voxel with a step length of 0.5 mm and curvature threshold of 0.2. This generated a connectivity map where each voxel represents a connectivity value, where the higher the number, the greater the probability of the pathway passing through that voxel. The connectivity maps were thresholded at 20 samples (out of 5000) to remove voxels that showed extremely low connectivity. The pathways were then binarized and masked onto the transformed FA, MD, and eigenvalue maps to derive mean values for each generated tract. Figure 3 shows an example of pathways generated for one participant. The secondary (λ2) and tertiary (λ3) eigenvalues were averaged to provide a value for perpendicular diffusion, while the primary eigenvector (λ1) was used as an index of parallel diffusion.
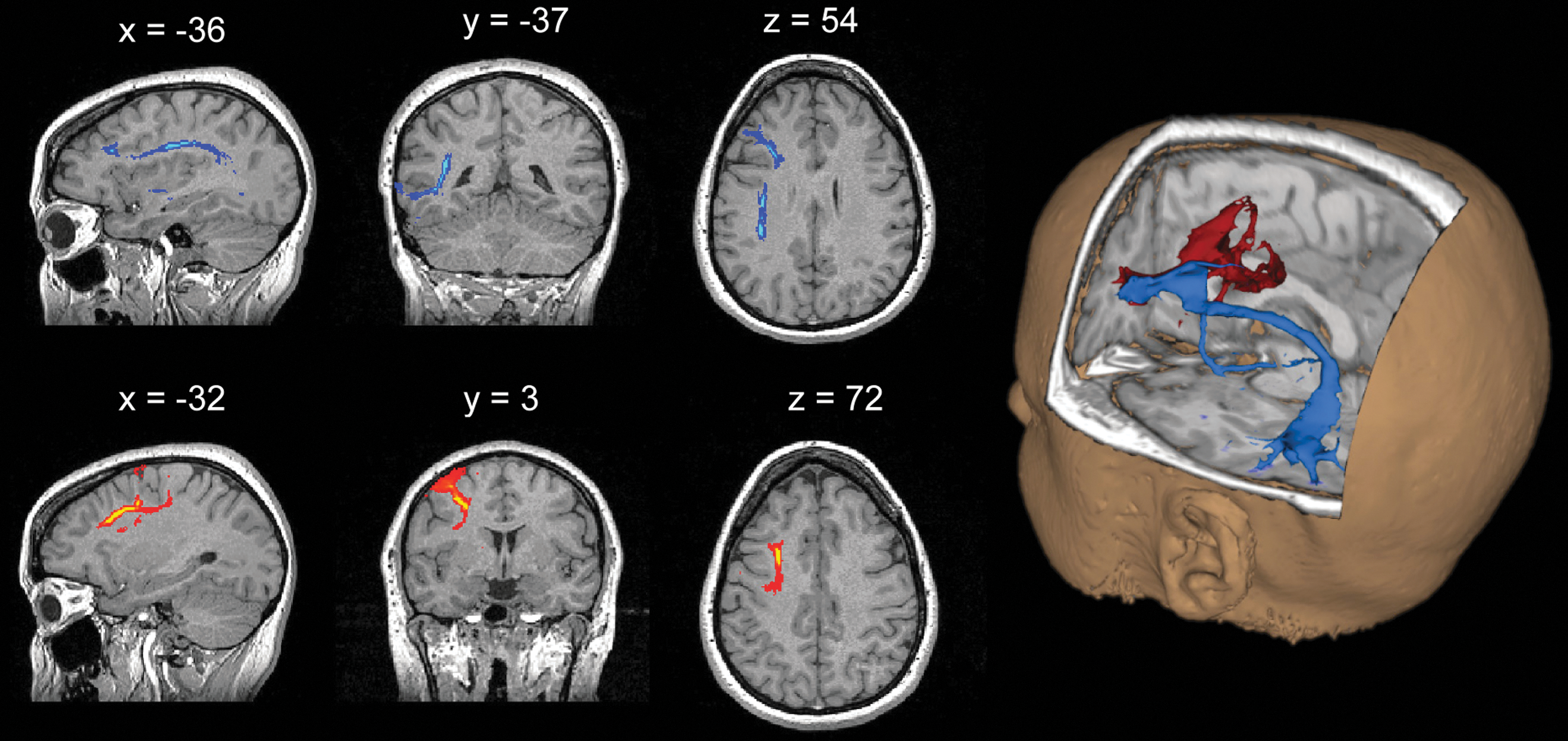
Tractography for the verb generation pathways. Pathways generated in one participant for verb generation. The pathway in blue shows the arcuate fasciculus tract connecting the MFG to the ITG, while the pathway in red connects the MFG to the precentral gyrus.
Statistical analysis
A bivariate Pearson correlation was used to investigate the codependence of all four diffusion measures (i.e., FA, MD, parallel, and perpendicular diffusion) for each pathway. Both showed a high degree of correlation between the four diffusion measures, where each measure significantly correlated with a minimum of two other measures (mean absolute Pearson's correlation value |r|=0.66 and 0.54 for the MFG to precentral and MFG to ITG pathways, respectively). To account for this codependence between diffusion measures, we therefore used a principal component analysis (PCA) to extract principal components for each pathway. As a general rule of thumb, factor loadings above 0.6 are considered high, while below 0.4 would be considered low. The factor scores were then entered into a Pearson correlation to look for associations with PPI parameter estimates. All statistical analyses were performed on SPSS 19.0 (SPSS, Inc., Chicago, IL) for Windows and used an α level of .05.
Results
The GLM analysis for the verb generation>X word contrast showed two large clusters of activation at Talairach and Tournoux coordinates −40 16 22 (left MFG, BA 46) and −52 16 1 [left inferior frontal gyrus, BA 9], showing typical regions of activation for the verb generation task (Fig. 1). The noun>X word contrast did not show any significant clusters of activations at corrected thresholds. We attribute this to the use of Xs as the control words, which may be too similar to the passive viewing of nouns.
For the PCA analysis of the diffusion measures, a single principal component was extracted for the MFG to precentral pathway that explained 76.04% of the variance. This loaded negatively on FA and positively with the three other diffusion measures. The factor scores for this component represent shared white matter profile across all subjects. Specifically, subjects had a high factor score if they had low FA, but high MD, parallel, and perpendicular diffusion. The MFG to ITG pathway revealed two components with comparable eigenvalues. The first component extracted explained 58.18% of the variance, while the second explained 41.69%. The first component loaded negatively on FA, but positively on MD and perpendicular diffusion, while the second component loaded positively on FA, MD, and parallel diffusion. The two factors showed differing patterns of loading where subjects had high scores for component one if they had low FA and high MD and perpendicular diffusion, while those with high scores for component two indicated high MD and parallel diffusion. These results are summarized in Table 2 with high factor loadings shown in bold. The Pearson correlations between the factor scores and PPI values for the MFG to precentral pathway showed a significant negative correlation (r=−0.54, p<0.05). Neither component for the MFG to ITG pathway correlated significantly with the PPI values (component 1: r=0.19, p=0.44; component 2: r=−0.114, p=0.64). The correlation results are illustrated in Figure 4.
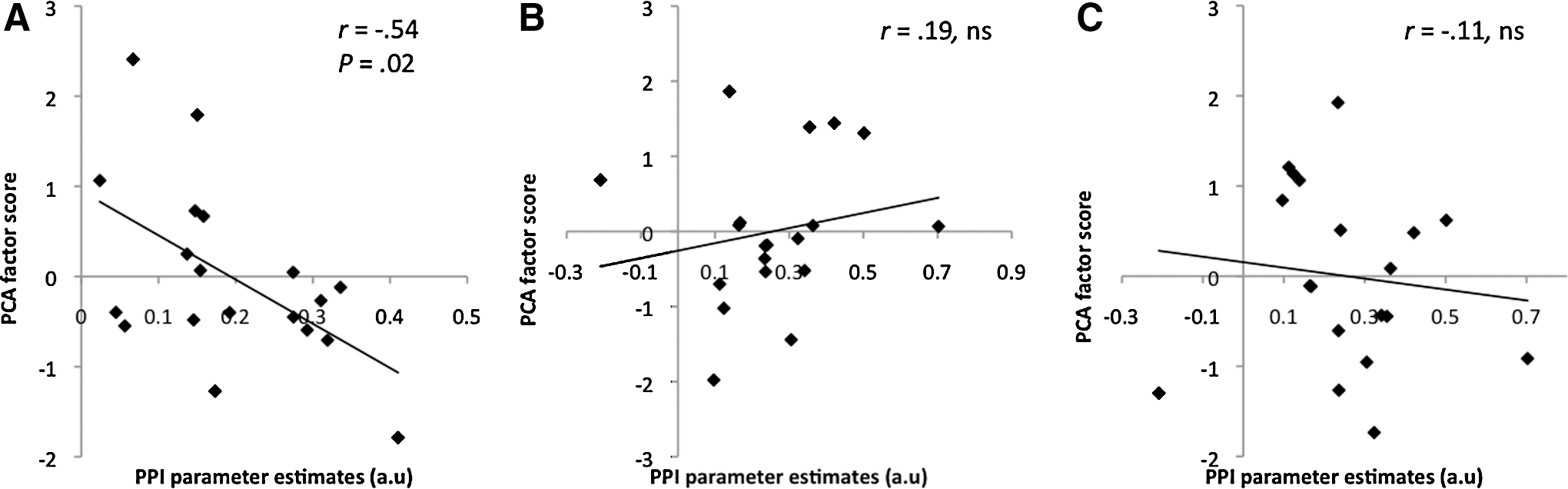
Correlations between PPI parameter estimates and factor scores from principal component analysis (PCA).
Factor Loadings on the Principal Components Extracted from the Diffusion Measures for Each Pathway
FA, fractional anisotropy; MD, mean diffusion.
Discussion
The current study explored the degree to which functional and structural connectivity are related by combining two MR techniques. PPI analyses were employed to identify functional connections relating to verb generation processes, which were combined with tractography to determine the structural connectivity between these regions. Significant clusters of activation in the left MFG were found that are typical for the verb generation task (e.g., Lehéricy et al., 2000; Martin et al., 1995; Papathanassiou et al., 2000; Warburton et al., 1996; Wise et al., 1991). This region was shown to be functionally connected to the primary motor and inferior temporal cortices. However, only the MFG to primary motor pathway showed a significant structure–function correlation.
The involvement of motor regions in the generation of verbs is not an uncommon proposal, although findings lack consistency. Earlier studies using positron emission tomography have shown activity in motor regions such as the presupplementary motor area (Papathanassiou et al., 2000), premotor (Papathanassiou et al., 2000; Péran et al., 2010; Tettamanti et al., 2005; Warburton et al., 1996; Wise et al., 1991), and supplementary motor area (Warburton et al., 1996; Wise et al., 1991). More recently, fMRI studies using similar language tasks involving action-related words have also yielded similar activation in premotor (Hauk et al., 2004, 2008; Tettamanti et al., 2005) as well as motor (Hauk et al., 2004) cortices. It has been suggested that this may be attributed to the strengthening of the connections between the frontal and temporal language regions and motor regions during development, when action-related words are mapped onto the physical action during learning (Pulvermüller et al., 1999). According to the motor theories of language comprehension, accurate perception and understanding of the language requires the participation of regions involved in the production of speech (Liberman et al., 1967). Given that we used a contrast that subtracted some aspects of language, it is possible that the connectivity with the motor region may be due to cognitive processes specific to processing verbs. Although this isolates some language functions to a certain degree, activation regions could nevertheless be contributing to a multitude of processes, such as the processing of action-related words (which could include motor simulation) (Willems et al., 2010) and executive functions involved in word selection. Interestingly, the strength of connectivity of this pathway showed a significant correlation with measures of white matter integrity derived from PCA. The underlying factor represented a structural profile whereby individuals with high factor scores had low FA values for this pathway associated with high levels of overall diffusion. This factor correlated negatively with the PPI parameter estimate values, indicating that those with a high degree of the above structural connectivity pattern showed lower functional connectivity between the MFG and precentral gyrus. Indeed, one would expect that tracts, which contain low levels of FA (and therefore low white matter integrity) along with high levels of perpendicular diffusion (therefore a greater axonal spacing) (Schwartz et al., 2005), would show a lower functional connectivity. Furthermore, Harsan and colleagues (2007) have demonstrated that opposing effects of FA and perpendicular together, are indicative of the presence of myelin. Applied to the current findings, low FA coupled with high perpendicular diffusion might suggest lower levels of myelin, which would lead to reduced functional connectivity as was observed here. This also shows that there is a strong relationship between the cognitive functions associated with generating a verb and the underlying structural connectivity patterns within the connecting white matter. We speculate that this significant association may be due to this pathway playing a specific role in verb generation, although the current data cannot fully ascertain this notion. Consistent with our finding, a recent study similarly reported structure associations between functional connectivity and the mean fiber bundle radius in the pathway connecting the supplementary motor area to Broca's area derived from a word generation task (Morgan et al., 2009).
In support of the involvement for motor regions in linguistic processing, recent evidence suggests a link from the Wernicke's area to the motor/premotor regions rather than directly to Broca's area, as previously hypothesized (Ardila, 2010; Barrick et al., 2007; Bernal and Altman, 2010; Bernal and Ardila, 2009). For example, tractography studies have identified the arcuate fasciculus (AF) directly connecting the temporal language areas with frontal language regions, as well as to the precentral gyrus, and this tract has also been shown to be strongly asymmetrical due to language lateralization (Catani and Thiebaut de Schotten, 2008; Catani et al., 2005, 2007; Glasser and Rilling, 2008; Nucifora et al., 2005; Powell et al., 2006), suggesting a role in language processes. Recently, a study using tractography in conjunction with fiber dissection has shown the existence of the classical AF, as well as separate anterior and posterior segments of the SLF (Martino et al., 2013). The anterior portion connects the superior temporal regions with the precentral gyrus, while the posterior portion follows a pathway between the middle temporal gyrus and angular gyrus. On visual inspection, the present data revealed a continuous pathway from the MFG seed region to the ITG with additional projections to the precentral gyrus, which is consistent with the AF reported in Martino and colleagues' (2013) fiber dissection study. It is quite possible that the tracts generated in the current study from the MFG to precentral gyrus, may also encompass the anterior connections described above, since our waypoint mask within the precentral gyrus was located on an individual by individual basis based on PPI localizations. Martino and colleagues (2013) reported that the anterior segment of the SLF projects to the ventral precentral gyrus, while the long AF connects to the dorsal region. Therefore, it is quite possible that the generated pathways may not differentiate between the AF and anterior SLF. Future work could explore these separate pathways and their relationships with functional connectivity, which may provide insight into the specific roles of each pathway.
The structural measures of the classical AF pathway identified in the current study did not show any significant associations with functional connectivity, despite previous reports of a link between white matter integrity and functional lateralization (Catani and Thiebaut de Schotten, 2008; Catani et al., 2005, 2007; Glasser and Rilling, 2008; Nucifora et al., 2005; Powell et al., 2006). In contrast to the MFG to precentral pathway, the AF pathway revealed two distinct factors underlying the pattern of white matter structure. It appears that there are two independent influences on diffusion parameters with comparable magnitude. The first has a strong negative influence on FA and strong positive influence on perpendicular diffusion, while the second had a particularly strong positive influence on parallel diffusion. We suggest these underlying factors represent structural integrity for the first factor, based on the opposing relationship between FA and perpendicular diffusion, and axonal alignment for the second factor, due to its strong association with parallel diffusion. We theorize that the unrelated influence of these two factors on white matter structure may attribute to the lack of correlation with functional connectivity in this pathway. Although we are not able to comment on the exact function of this pathway, we speculate that two underlying factors may relate to the diverse functions that this pathway might be involved in. Whereas the AF involvement in linguistic functions is not doubted, there is still much controversy surrounding the precise role of the AF and the type of signals it carries between the major language regions (Yeatman et al., 2011). Furthermore, a bidirectional transmission of information through the AF has been detected using a method called corticocortical evoked potentials in epilepsy patients (Matsumoto et al., 2004). Such a finding is a challenge to the long-standing Wernicke–Geschwind model of language, whereby language is processed in a unidirectional fashion from the temporal to frontal language areas. Not only was there an opposing transmission direction, there was greater excitation from the Broca's area to Wernicke's than in the other direction. Matsumoto and colleagues (2004) speculate that this could be due to a more dispersed arrangement of neurons in the posterior language regions, and therefore stimulation reaching only a subset of the neurons encompassing the posterior language area. Given that in the current study, only excitatory connections originating from the frontal language regions were explored for the verb generation task, mere speculations can be put forth in regard to the above notion. Indeed, the investigation into inhibitory and bidirectional connections would no doubt be of considerable value in understanding the communication between the anterior and posterior language sites, and perhaps, this may even contribute to the independent influences on the white matter structure of the AF observed in the current study.
One limitation of the study is that the lateralization of language was not considered. Due to the nature of the methodology, it was not possible to explore this specific element as the PPI analysis did not reveal any right hemisphere connections with the functional seed points. The specific objective of this study was to test for a direct relationship between structure and function, but also avoid the use of arbitrary (contralateral) or atlas-based ROI for reasons outlined by Smith and colleagues (2011).
Conclusions
By combining DTI-derived tractography with PPI, we identified a structure–function association for a pathway involved in verb generation. Specifically, a clear association was found between structure and function for the pathway connecting the MFG to the precentral gyrus. Using PCA, we determined that a structural profile with low FA and high perpendicular diffusion was associated with low functional connectivity. We suggest that this association may relate to this pathway being specifically involved in verb generation. In contrast, the AF pathway connecting the MFG with the ITG did not show a structure–function relationship in one of the pathways, between the frontal region and the motor cortex, it appears that this relationship, which we suggest may be related to two independent underlying latent factors that determine the structural properties of this pathway. We conjecture that these factors may relate to the diverse nature of cognitive functions of which the AF is involved.
Footnotes
Acknowledgments
This research was supported by the University of Auckland, Faculty of Science Research Development Fund, an NZ Royal Society Marsden Fund to I.J.K. The authors would also like to thank the reviewers for their helpful comments and suggestions.
Author Disclosure Statement
No competing financial interests exist for any author.