
Abstract
Introduction
In neuroimaging research, the term “functional connectivity” (FC) has been used to refer to the synchronous, low-frequency fluctuations across nonadjacent parts of the brain. Recent reports on the concordance of resting-state functional connectivity magnetic resonance imaging (fcMRI) findings with those from other modalities have generated a surge in fcMRI research (e.g., Bettus et al., 2011; Negishi et al., 2011; Pizoli et al., 2011 for EEG-fcMRI concordance; Greicius et al., 2009 for DTI-fcMRI concordance; Brookes et al., 2011 for MEG-fcMRI concordance). Remote changes in FC measured by fcMRI also coincide with reports on the network-level impairment that is thought to accompany various epilepsy syndromes (Bell et al., 2011; Bernhardt et al., 2009; Bonilha et al., 2007; Laufs, 2012; Riley et al., 2010), which makes fcMRI a useful research tool for evaluating network dysfunction in epilepsy.
In recent years, several resting state studies (Laufs et al., 2007; McGill et al., 2012; Waites et al., 2006; Zhang et al., 2010) have independently shown a decrease in default mode network (DMN) activity predominantly in the anterior and posterior cingulate cortex (PCC) in patients with temporal lobe epilepsy (TLE). This network is important in TLE because of its inclusion of the hippocampus (HC) and possible association with the episodic memory impairments observed in patients with refractory TLE (Holmes et al., 2012; McCormick et al., 2013). These diffuse, extratemporal reductions in the DMN agree with the diffuse structural impairments observed in the gray and white matter of patients with refractory TLE (Bernhardt et al., 2010; McDonald et al., 2008; Pittau et al., 2012). Thus, the DMN reductions may provide important information as to the overall degree of network damage and memory impairment in TLE.
However, both the nature and extent of the disease-related changes observed in medial temporal networks in TLE are a matter of ongoing debate. Pereira et al. (2010) reported decreased FC in hippocampal networks in patients with unilateral mesial TLE (mTLE) comorbid with ipsilateral hippocampal sclerosis (HS), and noted a more pronounced ipsilateral decrease in those with left mTLE compared to right. These findings parallel recent MRI studies that indicate greater gray and white matter atrophy in patients with left compared to right TLE (Kemmotsu et al., 2011; Kucukboyaci et al., 2011). However, other studies have shown reductions in hippocampal FC ipsilateral to right hippocampal seed (Morgan et al., 2011) that differentiate right versus left TLE patients. Another shows decreased FC between hippocampi and the DMN in patients with TLE (Pittau et al., 2012) that are similar for patients with right and left TLE. Furthermore, an increase in hippocampal FC in the hemisphere spared by TLE has also been reported by multiple groups (Bettus et al., 2010; Zhang et al., 2010).
To date, resting state fcMRI has been considered the gold standard for probing FC changes in neurological populations, including TLE. However, it has been shown that small but significant biases can be introduced to resting state data by choosing slightly different data acquisition procedures (e.g., eyes open vs. closed) (Jao et al., 2013; Patriat et al., 2013). For research groups who are interested in FC analyses that already have task-based functional MRI (fMRI) data, a viable alternative is to use task-regressed fMRI data, that is, to regress-out the activations from task-based data (i.e., task-regressed fcMRI)—an alternative method that may permit more control over cognitive processes than the presumed “resting state.” This task-regressed approach is gaining support in the literature: some data show that the correlation values obtained via task-regressed methods may be approximately equal to the values obtained from resting state studies (Arfanakis et al., 2000; Fox et al., 2006), while others posit that the convergence may be more qualitative than quantitative (Fair et al., 2007). This approach has yet to be applied to patients with refractory TLE, where task-based fMRI data may be readily available. Thus, task-regressed methods may be underutilized in FC research and results generated by this method could enrich our understanding of network dysfunction in TLE.
In this study, we apply a seed-based task-regressed FC analysis to explore FC changes in patients with TLE and to investigate whether or not these changes depend on the side of the seizure focus or other clinical variables. This method has been previously validated in healthy controls (Fair et al., 2007) and patients with autistic disorder (Noonan et al., 2009; Shih et al., 2010; Villalobos et al., 2005), but has yet to be applied to patients with TLE. We predict that the FC pattern resulting from our task-regressed analysis will match those found in the existing resting-state studies in TLE, providing further validation of this method. Specifically, we predict that patients with TLE will show reduced hippocampal FC primarily ipsilateral to their seizure focus. Given the involvement of the HC in the DMN, we expect the spatial pattern of the observed FC reductions to include key portions of the DMN (e.g., the posterior cingulate region). Furthermore, based on existing structural imaging findings, we predict that these FC reductions will be greater in patients with left compared to right TLE, perhaps resulting from the greater structural compromise.
Materials and Methods
Participants
This study was approved by the ethical standards committee on human experimentation at University of California, San Diego (UCSD) and completed according to the standards established in the Helsinki Declaration. Written informed consent was obtained from all participants. Seventeen patients with refractory TLE (ages 16–53) and 17 controls (ages 20–55) were included in the study. Control subjects were recruited through open advertisement and screened for past or present neurological or psychiatric conditions; those with a history of neurological or psychiatric illness were excluded from the study. All TLE patients were either undergoing or had previously completed presurgical evaluation at the UCSD Epilepsy Center and diagnosed by a board-certified neurologist (E.S.T. and V.J.I.) with expertise in epileptology. Seizure laterality was identified based on ictal recordings in a video-EEG monitoring unit (with scalp and/or foramen ovale electrodes, where available), seizure semiology, and neuroimaging results. In particular, patients exhibiting dual pathology on MRI (e.g., tumors) were excluded from the sample. Clinical MRI scans were available on all patients, and were visually inspected by a board-certified neuroradiologist for detection of unilateral HS, which was observed in 10 patients (6 patients with left TLE; 4 patients with right TLE; none with bilateral HS). Patients with contralateral temporal lobe structural abnormalities, and structural abnormalities other than HS were excluded. Demographic information for controls and patients with TLE are presented in Table 1. There were no significant group differences in terms of age, education or sex distribution, although the left TLE group included more female patients than the others (all ps>0.05; Table 1). The absence of significant age differences was particularly important given the possibility of age-related changes in FC (Steffener et al., 2012). In addition, Mann-Whitney U-tests revealed no significant distribution differences between patients with left and right TLE in terms of clinical variables, that is, ipsilateral hippocampal volume, age of seizure onset, seizure frequency, disease duration, and number of medications (all ps>0.1). However, there was a significant group difference in hippocampal volumes between controls and the TLE patients for the ipsilateral hemisphere (both ps<0.05). Moreover, disease variables did not exhibit a statistically significant correlation with the hippocampal volume in either hemisphere. Chi-square tests revealed a significant difference between the patient groups in their history of status epilepticus, but not in the diagnosis of co-morbid HS or history of febrile seizures (Table 1). All patients were right handed.
Sample Demographics and Clinical Characteristics
Both volumetric measures for the hippocampi were significantly different (p<0.05) between patients and controls for the affected hemisphere but were not significantly different between the patient groups.
TLE, temporal lobe epilepsy; RTLE, right temporal lobe epilepsy; LTLE, left temporal lobe epilepsy; HC, hippocampus; Sz, seizure; MTS, mesial temporal sclerosis.
Procedures
Structural image acquisition/processing
MRI was performed on a General Electric Discovery MR750 3.0 Tesla scanner with an in vivo eight-channel phased-array head coil. Image acquisitions included a conventional three-plane localizer, a T1-weighted three-dimensional (3D) structural sequence (TE=3.16 ms, TR=8.08 ms, TI=600 ms, flip angle=8°, FOV=25.6 cm, matrix=256×192, slice thickness=1.2 mm), and a diffusion-weighted sequence. The imaging protocol was identical for all participants, and all patients were seizure-free for a minimum of 24-h before the MRI scan.
Functional data acquisition and processing
Acquisition: Functional BOLD data images were acquired using a T2*-sensitive echo planar imaging sequence (TR=3000 ms, TE=30 ms, flip angle 90°, FOV=220 mm, matrix 64×64, slice thickness 2.5 mm). Forty-seven axial slices were obtained during each TR, covering the entire cortex. The first five volumes were discarded, and a total of 171 volumes were obtained for data analyses. The image files in DICOM format were transferred to a Linux workstation for functional BOLD analysis.
Processing and analysis: We employ a relatively novel method that relies on a task-regressed fcMRI dataset, where we analyze the temporal correlation of the residual fluctuations after removing the task response modeled separately for each task condition with an orthogonal regressor. Equivalent regression-based fcMRI methods have been employed in autism research with success (Noonan et al., 2009; Shih et al., 2010; Villalobos et al., 2005).
fMRI data analysis was carried out by using Analysis of Functional NeuroImages (AFNI) (Cox, 1996), Surface Mapping (SUMA) software (Saad and Reynolds, 2012) and MatLab (MathWorks, Natick, MA). Event related fMRI data was obtained from two counterbalanced, back-to-back runs. The underlying task followed an event-related, semantic judgment task where participants pressed a button with their left index finger each time a target word (i.e., name of an animal; n=24 target words per run), interspersed with new words, repeating words, and false font sequences, appeared on the screen. A similar, block-design implementation of this task is detailed in McDonald et al. (2010). In the event-related design, a stimulus appeared on the screen each 1.5–3 sec (M
inter-stimulus interval=1.9 sec). Participant wakefulness was tracked auditorily using the intercom and visually using the operator facing lights on the participant response box. Forty-seven axial slices were obtained during each TR, covering the entire cortex. The first five volumes were discarded, and a total of 172 volumes were obtained per each run. Sample stimuli and events design can be found in the Supplementary Figure S1; Supplementary Data are available online at
FC analysis
Task-regressed data (residuals of the fMRI analysis) and the motion files were first filtered using a bandpass filter of 0.008–0.08 Hz. Cerebral parcellations and subcortical volume segmentation were accomplished using FreeSurfer and imported, aligned, and applied to the EPI data, using AFNI. Masks for the whole brain, white-matter, and ventricles were created based on this segmentation in each patient's native volume. Three-dimensional, spherical hippocampal seeds (radius: 3 voxels) were drawn by the same image analyst trained in neuroanatomy in each subject's original structural volumetric image. Special attention was given to maximize the seed's coverage of the Cornu Ammonis (CA) subregions of each HC. Random checks on seed locations were performed bilaterally by a board-certified neuroradiologist. All confirmed seeds and regions of interest (ROIs) were then projected to native space of the functional images and time-series data were extracted for the selected ROIs and the hippocampal seeds. Resulting data were used to perform a subject-level, voxelwise FC analysis that generated brain-wide correlation coefficients separately for each seed, where motion parameters (one for each of the six translational and rotational axes), global signal level and scanner drift, measured via CSF and white-matter signal fluctuations, were regressed out as nuisance variables. Obtained voxelwise correlation coefficients were then converted into Fisher's Z (i.e., z-prime); projected into the Talairach space for standardization (TT_N27), where 3D two sample t-tests were performed separately in AFNI for each seed. Results were corrected for multiple comparisons using voxelwise p<0.05 and stringent cluster cutoffs (i.e., more than 50 first nearest neighbor voxels) whose conservatism was vetted using AFNI's built-in Monte Carlo simulator (3dClustSim) and were projected on to the Talairach TT_N27 standard brain for easy visualization. Two sets of thresholded statistical findings were interpreted: for healthy controls, correlation maps showing the degree of each voxel's hippocampal FC compared to the average cortex-wide hippocampal FC observed for the group; for patients, the degree with which each voxel's hippocampal FC in each patient group differed from its healthy control counterpart.
Results
FC in healthy controls
Consistent with previous research on the DMN connectivity in healthy individuals (Buckner et al., 2008), we observed a strong positive correlation between the signals in the left HC seed and left inferior parietal lobule (IPL), and between the left HC seed and the dorsal and the medial aspects of the superior frontal cortex, compared to the average correlation across the entire cortex. In addition, we observed a strong positive correlation between the left HC seed and the left paracentral and pre/postcentral cortex, mesial temporal, and superior temporal cortices (Fig. 1A). In the right hemisphere, the parahippocampal, superior temporal, pre/postcentral, anterior cingulate and medial prefrontal cortices exhibited strong positive FC with the left HC seed. In contrast, we observed a strong negative correlation between the left HC seed and the right IPL, and the lateral middle frontal gyrus.
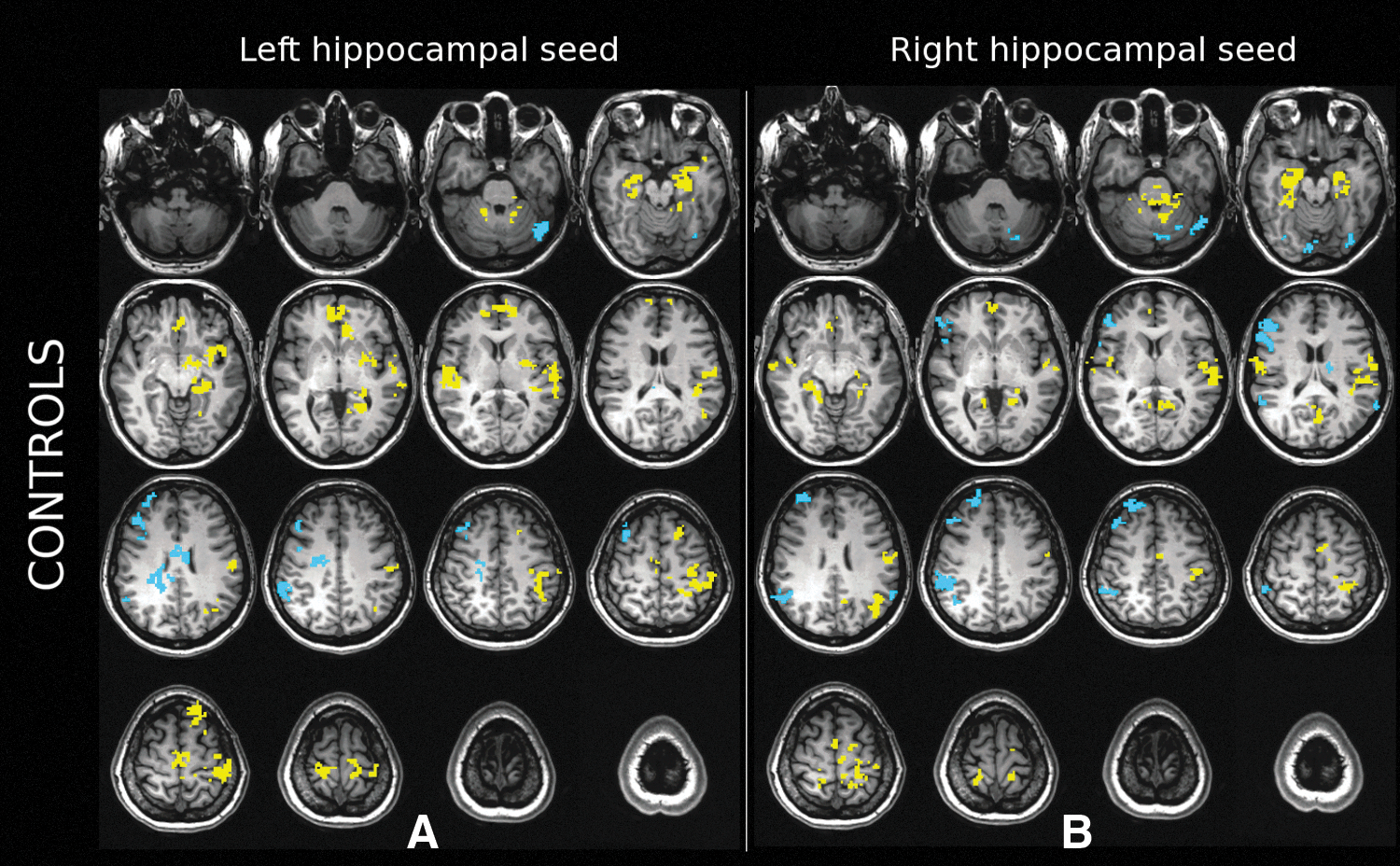
Panels show task-regressed hippocampal FC in healthy controls.
For the right HC seed, positive FC was observed in the parahippocampal gyrus, postcentral gyrus, precuneus, superior temporal, PCC and medial prefrontal regions in the right hemisphere (Fig. 1B). Anterior portions of the cerebellum near the vermis also showed significantly increased right hippocampal FC. Significantly, negative FC was observed in the right IPL, right supramarginal cortex, and along the lateral frontal cortex (superior, middle and inferior frontal gyri). In the left hemisphere, superior temporal gyrus, pre/post/paracentral cortex, precuneus, anterior and posterior cingulate and IPL showed increased FC with the right HC seed. A portion of the supramarginal gyrus, fusiform gyrus, and lateral frontal cortex (superior, middle and inferior frontal gyri) showed significant negative correlation with the right FC seed.
FC differences in left TLE patients
Compared to healthy controls, we observed significantly reduced FC between the left HC seed and the ipsilateral PCC/precuneus, medial frontal gyrus, and IPL. Additionally, patients with left TLE showed significantly lower FC in regions adjacent to these DMN areas, including the ipsilateral superior frontal gyrus, anterior cingulate gyrus, pre-/post- and para-central cortices (Fig. 2A). Increased FC was observed near the left lingual gyrus, posterior to the parahippocampal region. In the right (contralateral) hemisphere, the reductions were limited to the superior and medial frontal gyri, inferolateral parietal and paracentral cortices and the PCC/precuneus. Increased FC was also observed in patients with left TLE in regions of the right parahippocampal, supramarginal/IPL, and around the right caudate near the internal capsule.
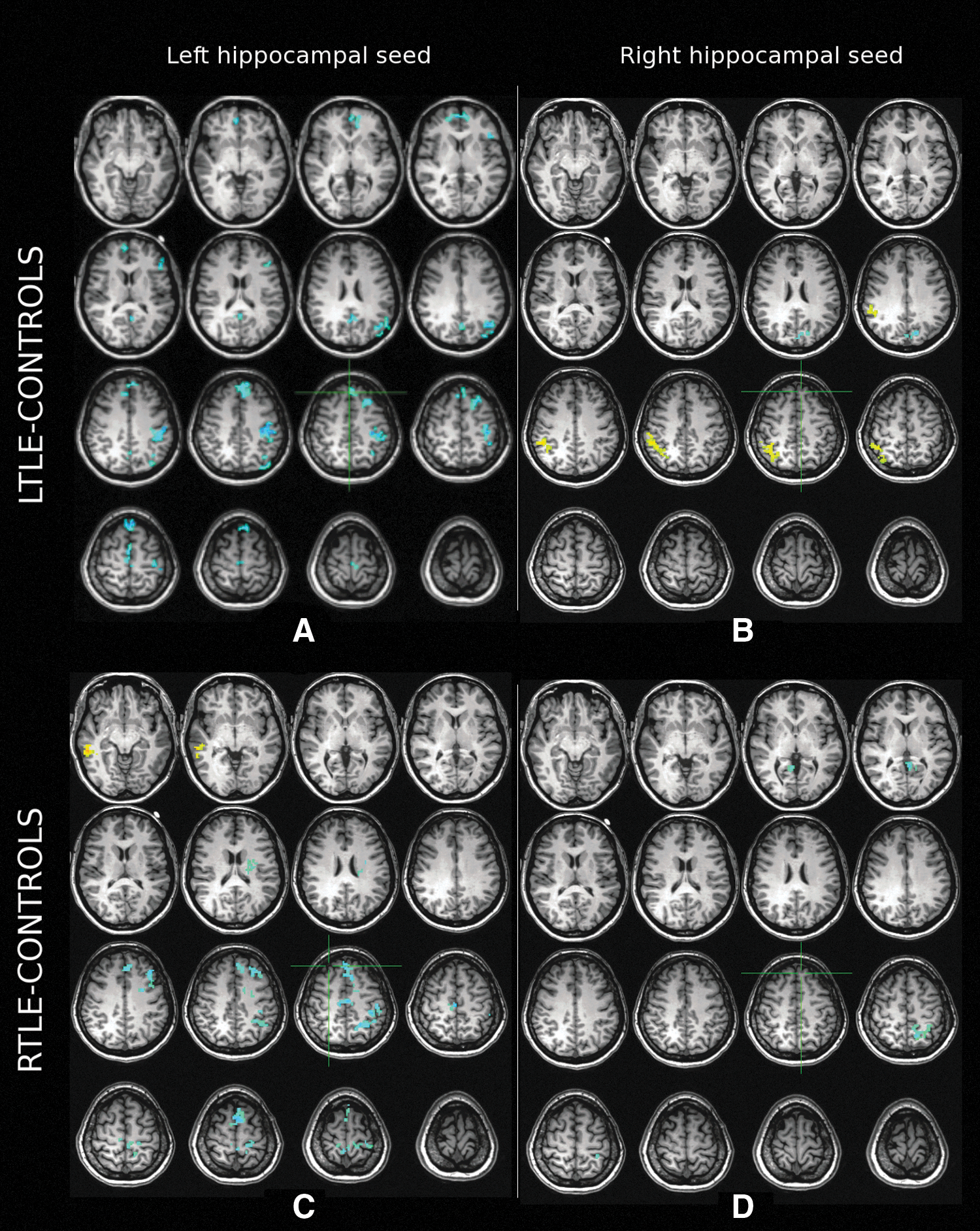
Middle
For the right (spared) HC seed, a decrease in FC was found in the left PCC/precuneus region, while subthreshold (cluster size less than 50 but larger than 40) decreases were also noted in the left anterior cingulate, middle frontal, middle and superior temporal gyri (Fig. 2B). Significantly, increased FC was observed in the right IPL, while a subthreshold increase was noted in the left lingual gyrus around the calcarine sulcus. No significant reductions were noted in the right hemisphere.
FC differences in right TLE patients
For the right HC seed, FC was decreased bilaterally in the PCC-precuneus region and ipsilaterally in the right hemisphere near the posterior portion of the parahippocampal gyrus (Fig. 2D). A subthreshold increase in FC was observed in the right IPL region. Significant decreases in FC were also observed contralaterally in the left precuneus, left IPL and in the left cerebellum near the vermis.
For the left (spared) HC seed, reduced FC was observed along the left superior, middle, inferior and medial frontal gyri, in the pre/post/paracentral gyri, IPL, PCC/precuneus and anterior and middle cingulate cortices (Fig. 2C). The left caudate nucleus also showed decreased FC with the HC on the left side. Decreased FC was found in the right pre-, post- and paracentral cortex, medial superior frontal gyrus. A significant left hippocampal FC increase was observed in the right middle temporal cortex and the parahippocampal gyrus.
FC correlations with disease-related variables
Given statistically nonsignificant yet highly variable distribution of seizure duration (in years) and seizure frequency (defined as number of seizures in the past month) we also performed a post hoc correlational analysis to observe any association of the FC differences within the TLE groups with these clinical variables. Spearman's rho was selected as the criterion measure given our small sample size and the robustness of this measure to the effects of any outlier data (Schwarzkopf et al., 2012). When all patients were analyzed together, we found no significant correlations between FC differences in either hemisphere and the clinical variables (i.e., duration and seizure frequency), making it difficult to attribute the group-level FC differences to these clinical variables. Results remained nonsignificant when tabulated for ipsi/contralateral laterality with respect to TLE foci across both groups.
Discussion
The goal of this study was to examine network dysfunction in patients with left and right TLE using a novel, task-regressed seed-based FC analysis whose application is rare in epilepsy research. In doing so, we first explore findings in healthy controls and compare them to previous resting-state studies. We then examine hippocampal FC changes in patients with TLE and explore whether our findings showed convergence with existing FC findings in epilepsy literature. Specifically, we tested the hypotheses brought forward by the existing literature that (1) patients with TLE exhibit reductions in hippocampal FC compared to healthy controls, (2) the reductions are more pronounced ipsilaterally to the seizure focus (3), and these reductions include areas associated with the DMN. We also tested the hypothesis that patients with left TLE would show more widespread reductions than those with right TLE given literature of greater structural damage in patients with left relative to right TLE (McDonald et al., 2008).
For healthy controls, hippocampal FC patterns matched the pattern highlighted in the extant literature in multiple ways. First, we were able to observe the hippocampal FC of the regions that are associated with portions of the DMN. Namely, the significant hippocampal connectivity of the superior temporal, inferior parietal, medial frontal and PCC/precuneus regions observed in our healthy controls overlapped with previous delineations of the DMN (Buckner et al., 2008), also known as task-negative networks (Fox and Raichle, 2007). Similar to past research, our results showed left>right laterality pattern for the IPL and the lateral temporal gyrus (Swanson et al., 2011), and a right>left laterality pattern for the medial frontal gyrus (Saenger et al., 2012). Overall, DMN appeared better delineated in the left hemisphere with the ipsilateral HC seed compared to the right hemisphere with the ipsilateral HC seed (Fig. 1). We acknowledge and discuss possible methodological reasons for this hemispheric difference further below.
For the patient population, our results suggest that the pattern of FC exhibited by TLE patients is significantly different from that of healthy controls, and both the nature and the extent of the difference vary by brain region. We found that patients with TLE exhibit reductions in hippocampal FC in areas that are consistent with previous reports (i.e., along the medial components of the DMN). Similar to past research, we also found that the reductions in FC appear to colocalize in nearby regions with purported involvement in the medial DMN (Table 2). In the discussion that follows we describe (1) FC reductions generated by our task-regressed approach within regions of key interest, and (2) how the areas of FC reduction we find coincide with previous resting state studies of reduced network connectivity in TLE.
Distribution of the Patient Findings in the RAI Coordinate Space
Table summarizes the RAI coordinates of the peak differences between patients and healthy controls for each seed where cluster size>40. Bolded items meet significance (p cluster<0.008). Entries with x>0 are located in the left hemisphere.
PCC/precuneus (RAI:±10, 54, 14;±14, 61, 41)
Both left and right TLE patients exhibited small but significant bilateral reductions in FC in the PCC and precuneus regions of the DMN mostly when the HC seed was found ipsilateral to seizure focus. In both cases, the results were more pronounced for the ipsilateral hemisphere. These results are concordant with the majority of the past DMN findings in TLE (Laufs et al., 2007; Luo et al., 2011b; McGill et al., 2012; Pereira et al., 2010; Pittau et al., 2012; Waites et al., 2006). Our results also agree with Doucet et al. (2012), who find more extensive FC reductions in left mTLE patients. Luo et al.'s (2011a) report of similar results in patients with absence seizures and Kim et al.'s (2012) report of precuneus FC changes in early Alzheimer's Disease (AD); however, suggests that our findings may capture a broader disease process not specific to TLE. Given how PCC is considered a key hub of the DMN (Buckner et al., 2008), with potential functional involvement in HC-related episodic memory, these findings are especially noteworthy for investigating the cognitive changes that accompany decreased PCC/HC FC in patients with TLE.
Inferior parietal lobule (RAI:±48, 41, 39)
Reductions in the FC between the HC and the IPL were observed in the left hemisphere with the ipsilateral seed for both patient groups. A concomitant FC increase observed in the right IPL area with the right (spared) hippocampal seed in the left TLE patients was absent in the right TLE patients with either seed. These differences appear in line with Swanson et al. (2011), who report spatial pattern asymmetries in this region in healthy controls that may be enhanced in the IPL of patients with TLE. It is possible that this spatial pattern of increased FC partially follows the localization of the nearby task-positive networks (intraparietal sulcus), which are known to correlate inversely with network patterns obtained by DMN-based seeds (Fox and Raichle, 2007). However, this increase was specific to the right hemisphere for all patients, regardless of seizure laterality. The opposite pattern of the HC-IPL relationship observed for ipsilateral and contralateral seeds has been discussed in detail by Holmes et al. (2012), and the larger hemispheric differences have been implicated in improved memory performance (Holmes et al., 2012; McCormick et al. 2013). The regions of increased FC we found differed from those reported by Bettus et al. (2010), who reported increases specific to the HC and the amygdala contralateral to seizure focus. We believe that these differences can be partially attributed to sampling and methodological differences (ROI-based analyses with a majority of HS+TLE patients by Bettus' group compared to our whole-brain analyses with TLE patients where only 60% HS+) and in part be attributed to the specific phase-encoding direction (Mathiak et al., 2012). It's also worth noting that our task-regressed findings otherwise showed an encouraging overlap with Bettus et al's (2010) resting state results in the left hemisphere, as Bettus et al.'s data revealed bilateral reductions that were more pronounced ipsilateral to seizure laterality (Table 2). One area of concern on the task-independence of our findings in this region is raised by Fair et al. (2007), who note larger differences between pure-resting state versus event-based task-regressed methods in this region, and suspects that the consistency of activation patterns in this region before and after task-regression may be associated with the incomplete removal of task effects.
Pre/post/paracentral regions (RAI:±44, 8, 38;±43, 25, 43;±7, 32, 53)
The left sensorimotor areas surrounding the central sulcus both in the lateral and medial aspects (i.e., pre, post and paracentral cortex) exhibited lower hippocampal FC in both right and left TLE patients. For each patient group, the FC reductions appeared largest in the left hemisphere with ipsilateral seed location. Despite the differences in methodology (i.e., resting-state vs. task-regressed data), these results were in agreement with Zhang et al. (2009), who observed decreased FC in the auditory and sensorimotor cortex of patients with TLE. They also agree with Luo et al. (2011b), who report FC reductions within the sensorimotor network of patients with TLE. Voets et al. (2012) have similarly reported similar FC reductions in the ipsilateral sensorimotor cortices of right TLE patients that co-localized with reductions in underlying white-matter integrity. Although the direct relationship of this finding to clinical symptoms remains unknown (Voets et al., 2012), it has been proposed that patients with TLE have impairments in tactile discrimination (Grant, 2005) and egocentric memory (Weniger et al., 2012), both of which rely on sensorimotor networks that may be adversely affected in TLE. Furthermore, structural imaging studies have shown pronounced cortical thinning in sensorimotor cortex in TLE (Lin et al., 2007; McDonald et al., 2008) that co-localize with the hippocampal connectivity reductions observed in the sensorimotor areas. Despite the replication of similar results in other FC research in epilepsy that coincide with structural compromise, it is of note that left sensorimotor cortex was also activated by our semantic task in the left TLE group, despite the left-handed response to target-stimuli. Therefore, given the qualitative local overlap between task-related and task-regressed activity around the left central sulcus, it is difficult to attribute the TLE-control FC differences observed solely to disease-related FC reductions.
In conclusion, our findings provide further support for the FC changes observed in healthy controls and patients with TLE using a novel method that relies on task-regressed fMRI data. The convergence of our patient results with past resting-state studies (Doucet et al., 2012; Holmes et al. 2012, Pereira et al., 2010) in terms of the variation in the extent, strength and the laterality of findings is encouraging for the employment of similar task-regressed fcMRI approaches in epilepsy research. By seeding the hippocampal region, which is traditionally associated with the DMN, (Buckner et al., 2008) we were able to validate existing findings in resting-state DMN connectivity in TLE with task-regressed MRI data. The most compelling overlap in findings was that ipsilateral hippocampal FC reductions were observed in the ventral PFC, PCC and the IPL in left TLE patients. Consistent with Doucet et al. (2012), right TLE patients exhibited smaller and more localized DMN reductions compared to left TLE patients, which were mainly in the PCC of the affected hemisphere. Consistency of the PCC reductions across both patient groups support Greicius et al.'s (2003) view that the PCC may be a “critical node” of the DMN, which contributes towards an autonomic capacity in episodic memory (McCormick et al., 2013). It now appears that, despite some variation in findings, most of the existing research has implicated midline structures, for example, medial frontal cortex, PCC, precuneus, and the mesial temporal lobes. These structures also appear involved in the neuropathology of other neurodegenerative disorders, including AD (Greicius et al., 2004) and dementia with Lewy bodies (Kenny et al., 2012), which makes it likely that the FC changes observed in patients with TLE may be related to network-level changes not specific to epilepsy.
We relied on a relatively novel approach, task-regressed fMRI, whose reliability and validity has been supported in past MRI research. This method removes the task-related BOLD activation of the fMRI data using computational techniques (e.g., linear regression, component analysis) and treats the residual, task-regressed fMRI data as a substitute for resting state MRI data. Because of its potential diagnostic utility (e.g., seizure lateralization and surgical outcome prediction in TLE; Morgan et al., 2012), FC research has the potential to provide important milestones in our understanding of the disease processes and the mechanisms that underlie network impairment in epilepsy. The convergence of task-regressed fcMRI with resting-state findings are encouraging and could permit fMRI researchers to utilize task-based functional data to perform FC analyses in the absence of pure resting state data. Equivalent regression-based fcMRI methods have been employed in healthy controls (Fair et al., 2007) and those with autistic disorder (Jones et al., 2010; Noonan et al., 2009; Shih et al., 2010; Villalobos et al., 2005) with success, showing that results obtained using a task-regressed method strongly correlate with those obtained using a traditional resting-state analysis. Jones et al. (2010) posit that the task-regressed approach increases the likelihood that observed correlations are much more likely to be driven by task-independent fluctuations. Arfanakis et al. (2000) has shown that independent component analysis (ICA) can be reliably used to generate task-regressed data that is not only clear of task-related BOLD response, but also sensitive to signal increases as low as 0.3% and convergent with resting state data in terms of FC analysis in the low frequency domain. The same group also suggests that multiple interleaved tasks, as is the case with our data, may facilitate the optimal removal of task-effects. Erhardt et al.'s (2011) discussion of cross-modal results across various methodologies, which especially highlights the robustness of the DMN across various FC methods, along with Langers and Van Dijk (2011) report of convergent results from seed-based and ICA-driven approaches, supports the accuracy and the reliability of DMN findings from seed-based task-regressed approaches, similar to the one employed in our analysis.
Despite the promising use of this task-regressed approach in neurological populations, such as TLE, there are several limitations that should be discussed. First and foremost, it is possible that the effects of task-related fMRI signal changes in our regions of interest may have attenuated or masked real findings. Comparing results from 10 subjects across pure resting-state, block-design derived and event-related design resting state data, Fair et al. (2007) raise concerns on the quantitative differences between different methodologies, and point out that the task-regression method assumes a discrete, linear additivity and superposition of resting state and task-activated signal in observed fMRI data. Task-independence of “temporally coherent networks” is also questioned by Calhoun et al. (2008). Both groups point out that the strength and the pattern of correlations observed between different fcMRI approaches do not overlap perfectly, and that despite their qualitative similarity, quantitative differences may exist between these methods. They posit that the GLM's inability to remove nonlinear effects, or task-dependent attenuation in resting state signal, which undermines the assumption made on the linear additivity of the fMRI signal between resting-state and task-based components, may be responsible for the divergence of quantitative results across methods. Therefore, it is critical to ensure that the task-regression residuals are indeed orthogonal to the underlying task activation, which can be difficult to establish in most event-related designs since commonly used inter-stimuli intervals often fall inside the frequency range explored in FC analysis (e.g., 0.1–0.01 Hz). This may be easier to ensure for simpler tasks with more frames, similar to the one we employ with 342 frames. By this token, it's noteworthy that Fair et al.'s (2007) findings were based on the comparison of 124–128 frames of resting state data with 73 frames of event-related data, although it's often advisable for the latter method to include more frames for better signal-to-noise ratio (SNR). Furthermore, the authors discuss that results differences are smaller for nontask seeds, and that the event-related task-regressed data may find fewer statistically significant areas than pure resting state data, which is in line with the higher SNR of shorter-duration event-related designs. By that token, findings from event-related task-regressed methods may also be statistically more conservative (i.e., lower false positive rates). Similarly, there is also evidence that the task-regressed approaches adequately filter out nuisance parameters (e.g., motion, high-frequency noise) and that task-related activation can yield to unbiased estimates for FC of the DMN when following certain methodological guidelines (Muller, 2011). Erhardt et al.'s (2011) report on the high cross-methodological robustness of DMN findings is also worth noting in favor of task-regressed methods achieving comparable results with resting-state analyses especially for regions involved in the DMN.
Several other limitations of our study should be noted. Although our sample size is comparable to those of other fcMRI studies, larger patient samples are needed to increase the power of statistical analyses and allow more detailed left TLE versus right TLE comparisons. When most of the FC reductions we detect appear close to the midline, more conservative interpretations that de-emphasize hemispheric specificity are needed, as fMRI data can be subject to a directional bias in the phase-encoding direction (Mathiak et al., 2012). We utilize a-priori correction to minimize this bias (i.e., combine A-P and P-A phase-encoded data for distortion correction). Some of the DMN signal correlations observed in our healthy controls (i.e., between the left HC and left PCC) also appear weaker compared to existing literature. This may be a by-product of our reliance on hippocampal seeds for clinical reasons, despite the fact that HC seeds are known to generate less robust correlations compared to the hub regions of the DMN (i.e., PCC, IPL and the ventromedial PFC). This may also be due to task-specific idiosyncracies that we are not able to accurately model and regress out with our seed-based approach. Moreover, we are not currently able to vet our task-regressed findings against resting-state findings for the same patients. A direct comparison between task-regressed and resting state data would provide the most compelling evidence that the two methods produce similar results in patients with TLE. Data collection to make this comparison is currently underway in our lab. Finally, future research that combines structural and functional imaging data with neuropsychological and neurobiological findings would contribute towards explanatory models that would provide additional insight into the determinants and consequences of network-level dysfunction in patients with TLE and related neurological disease.
Footnotes
Acknowledgments
This work was supported by the National Institute of Health (C.R.M., grant number R01NS065838-01A1), the Epilepsy Foundation Health Science Student Fellowship (N.E.K.) and the Epilepsy Foundation Behavioral Sciences Post-Doctoral Fellowship (N.K.). We also greatly acknowledge support from GE Healthcare. We would like to thank R.A. Muller and his graduate students Chris Keown and Aarti Nair for the early methodological validation provided, and Alison K. Noda and Mark Fiecas for their input for the revision.
Author Disclosure Statement
Authors report no conflicts of interest and confirm that no competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.