
Abstract
Temporal lobe epilepsy (TLE), affecting the medial temporal lobe, is a disorder that affects not just episodic memory but also working memory (WM). However, the exact nature of hippocampal-related network activity in visuospatial WM remains unclear. To clarify this, we utilized a functional connectivity (FC) methodology to investigate hippocampal network involvement during the encoding phase of a functional magnetic resonance imaging (fMRI) visuospatial WM task in right and left TLE patients. Specifically, we assessed the relation between FC within right and left hippocampus-seeded networks, and patient performance (rate of correct responses) during the encoding phase of a block span WM task. Results revealed that both TLE groups displayed a negative relation between WM performance and FC between the left hippocampus and ipsilateral parahippocampal gyrus. We also found a positive relationship between performance and FC between the left hippocampus seed and the precuneus, in the right TLE group. Lastly, the left TLE specifically demonstrated a negative relationship between performance and FC between both hippocampi and ipsilateral cerebellar clusters. Our findings indicate that right and left TLE groups may develop different patterns of FC to implement visuospatial WM. Indeed, the present result suggests that FC provides a unique means of identifying abnormalities in brain networks, which cannot be discerned at the level of behavioral output through neuropsychological testing. More broadly, our findings demonstrate that FC methods applied to task-based fMRI provide the opportunity to define specific task-related networks.
Introduction
While episodic memory has been commonly described as one of the most common cognitive impairments reported by temporal lobe epilepsy (TLE) patients, evidence of working memory (WM) impairment in TLE has also been observed (Corcoran and Upton, 1993; Stretton and Thompson, 2012). More generally, different WM effects have been reported to be dependent on the side of the TLE. Using functional magnetic resonance imaging (fMRI), van Asselen et al. (2006) described bilateral “hippocampal formations as having a fundamental role in maintaining spatial information in WM over an extended time period.” Thus, functional data have suggested that both medial temporal lobes (MTLs) are critical for visuospatial WM functionality.
One method for testing the role of each MTL in visuospatial WM in TLE patients involves functional connectivity (FC). Indeed, FC-based analysis has been highly useful for clarifying both the brain regional involvement and the functional integrity of specific cognitive networks either at rest (Fox and Greicius, 2010; Shirer et al., 2012) or during cognitive tasks (He et al., 2007; Ranganath et al., 2005). Furthermore, FC has been shown to be correlated with cognitive performance (e.g., Doucet et al., 2012; Welchew et al., 2005). It is now largely accepted that FC can be an efficient means of revealing functional impairments in populations suffering from a neurologic disease (Fox and Greicius, 2010; Greicius, 2008). Ranganath et al. (2005) were among the first to apply this FC-based method on data acquired during a visual WM task. Using an event-related design, they tested for differential FC between the hippocampus and the rest of the brain in 15 healthy controls during the processing of novel objects. They found that FC between the hippocampus and a large network involving perirhinal, orbitofrontal, and posterior cingulate cortices was enhanced for the objects successfully recognized on a subsequent postscan task. These authors suggested that successful memory formation was associated with transient increases in cortico-hippocampal interaction. In the case of the epilepsy pathology, Voets et al. (2009) explored the extent of functional reorganization within memory networks in nine LTLE patients. The authors used a complex visual memory-encoding task, involving familiar color photographs, and demonstrated significant reductions in FC between bilateral MTL, occipital and left orbitofrontal regions in the patients during complex scene encoding. These authors suggested that “FC analyses may offer improved sensitivity to subtle changes in the distribution of memory functions.” In addition, Wagner et al. (2007) showed that stronger FC between the hippocampus and neocortical regions, such as inferior frontal and superior temporal cortices, was associated with better performance in 11 right and 10 left TLE patients during a verbal encoding and recognition memory task composed of concrete and highly imageable word pairs. In contrast, Frings et al. (2009) did not observe a significant relation between interhippocampal FC and performance during a spatial memory task, in 14 TLE patients. The task required the encoding and recognition of object locations in a virtual three-dimensional environment.
Overall, these studies investigated whether FC between the MTL and the rest of the brain can be transiently modulated by memory processes. These studies highlight that FC-based methodology may, indeed, be particularly efficient in revealing the effects of neurologic pathology on brain activity during a cognitive task.
In this context, we investigated correlations of signal fluctuations between each hippocampus and the rest of the brain in unilateral right and left TLE patients, during the encoding phase of a visuospatial WM task. More precisely, we sought to determine whether hippocampal FC during visuospatial WM was related to subsequent performance, and whether such associations varied as a function of right or left TLE. For this purpose, we used a version of the Corsi task (Corsi, 1972; Milner, 1971), which was adapted for the fMRI environment. This task is a well-recognized method for assessing visuospatial WM, and it has been found sensitive to the effects of TLE compared with controls (Milner, 1971). None of the previously cited studies investigated brain FC modulation during this task. We hypothesized that an FC-based approach will clarify the contribution of each hippocampus during this visuospatial WM in TLE. Furthermore, we postulated that the left and right TLE patients would show quite different patterns of FC between the epileptogenic and healthy hippocampi.
Materials and Methods
Participants
A total of 36 patients with refractory unilateral TLE (18 left and 18 right) were recruited from the Thomas Jefferson University Comprehensive Epilepsy Center. A combination of electroencephalography (EEG), MRI, positron emission tomography (PET), and neuropsychological testing was used to lateralize the side of seizure focus (Sperling et al., 1992). All participants met the following inclusion criteria: unilateral temporal lobe seizure onset through surface video/EEG recordings (i.e., a single unilateral temporal lobe focus); MRI evidence of temporal lobe pathology confirming the presence of unilateral temporal lobe pathology in the epileptogenic temporal lobe and no extra-temporal abnormalities; concordant PET finding of hypometabolism in the temporal lobe (available for most patients, with no patient having a nonconcordant PET); and full-scale intelligence quotient of at least 75. TLE patients were excluded from the study for any of the following reasons: medical illness with central nervous system impact other than epilepsy; head trauma; prior or current alcohol or illicit drug abuse; extratemporal or multifocal epilepsy; contraindications to MRI; psychiatric diagnosis other than an Axis-I Depressive Disorder; or hospitalization for any Axis I disorder listed in the Diagnostic and Statistical Manual of Mental Disorders, IV. Depressive disorders were allowed given the high co-morbidity of depression and epilepsy (Tracy et al., 2007). Participants provided written informed consent. The study was approved by the Institutional Review Board for Research with Human Subjects at Thomas Jefferson University. Table 1 outlines the demographic and clinical characteristics of the patients. The Edinburgh handedness scale was used as a measure of handedness (Oldfield, 1971).
Demographic Data
CPS as primary type with rare secondary generalized seizures.
CPS as primary type with rare generalized tonic-clonic seizures.
MTS, mesial temporal sclerosis; LTLE, left temporal lobe epilepsy; RTLE, right temporal lobe epilepsy; CPS, complex partial seizures; SPS, simple partial seizures; IQ, intelligence quotient.
Block span task
Assessment of patients' visuospatial span was determined before the scan using the Corsi Block Span (BS) task. The span was determined by the ascending number of items until the patient had two recall failures at the same span length. The BS task was adapted for the fMRI environment. Instead of using blocks, a picture of 10 squares was presented (Fig. 1). Each square corresponded to 1 of 10 keys (one for each finger) on a response pad.
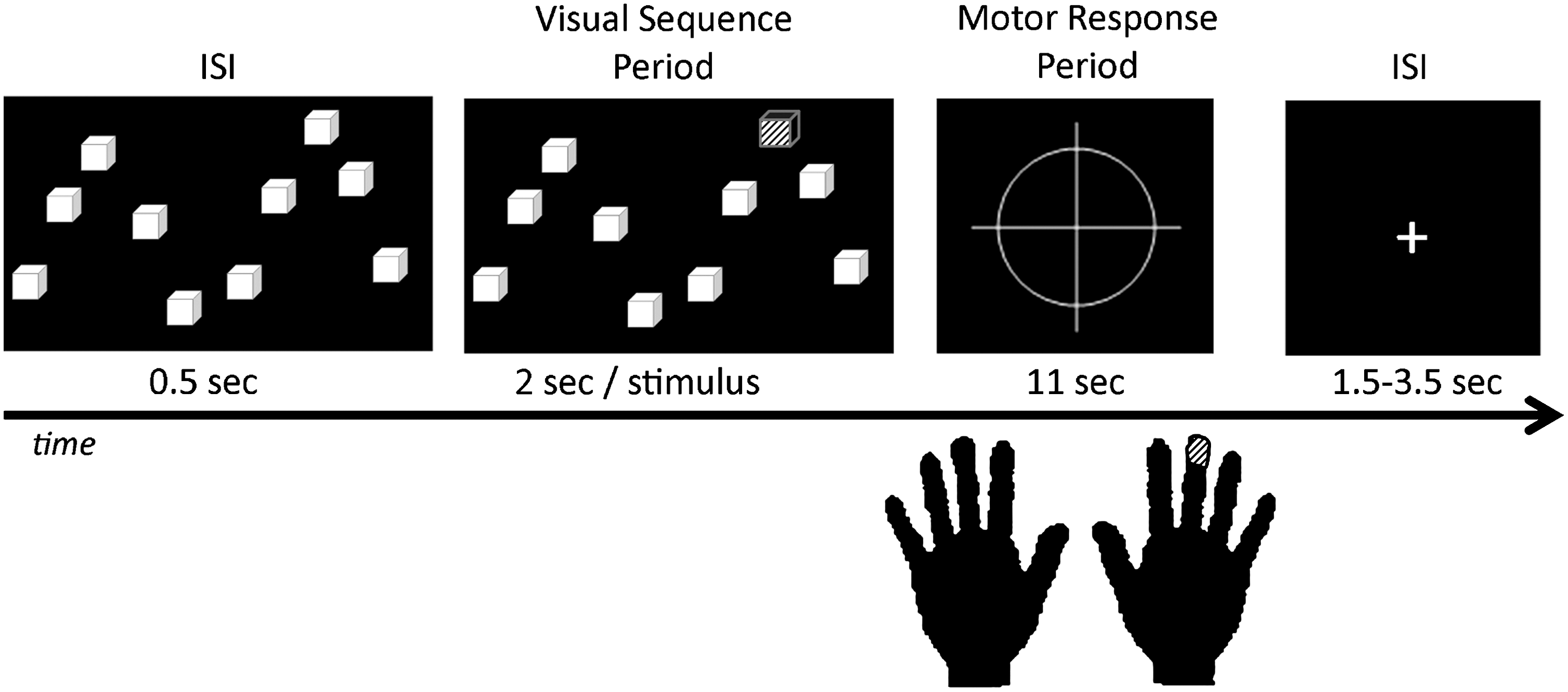
Block span task design. The patient was instructed to remember the position and sequence of successively illuminated squares, and to press the corresponding buttons on the keypad in the same order. ISI, inter-stimuli interval.
The experiment was composed of 24 visual sequence periods. A sequence consisted of the presentation of n randomized stimuli, with each stimulus being a single yellow block presented for 2 sec (note only one stimulus became yellow at a time). The length of the sequence was customized to each individual by adding one element/stimulus to their baseline BS performance (n=BS+1) collected outside the scanner before scanning. This “span plus one” strategy was utilized to make the WM task more challenging, and more likely to evoke the full network of connections needed to implement effective WM. The patient was instructed to remember the location of the yellow squares as they appeared, with no button press during this visual sequence period. This phase was considered the encoding phase. At the end of the visual sequence period (e.g., encoding phase), the patient had to recall the sequence by pressing his/her fingers on the response pad in the same order as the previously presented squares. The duration of the recall phase was 11 sec. Between two visual sequences, an inter-stimulus interval was presented with a jittered duration between 1.5 and 3.5 sec. No patient had motor impairments that could prevent them from pressing buttons. Before scanning, all patients completed at least one training session to learn the pairing of each visual stimulus with the appropriate button response. Patients were presented the same stimuli as during the BS task, but were instructed to press the corresponding button immediately, and were given feedback until they were able to quickly and accurately match each stimulus with the appropriate response. This training accounted for the paired associative learning demands of the motor response during BS task execution. Of note, on a postscan interview of the patients, we had each patient record the method or strategy they used to recall the location of the stimuli. In no case did the patient describe using a verbal or labeling strategy related to the ordinal position of items along a 1-D axis, (e.g., as items “1” through “10”), or another strategy that would minimize visuospatial WM processes. Based on the fact that patients had different spans, the experiment's duration was not similar for all the patients (mean±SD=10.2±0.9 min, [min-max]=[8–12]min).
The software E-prime v1.1 was used to run the BS paradigm, record, and save the patients' answers. Due to the high level of difficulty of the task, one sequence was considered correct when at least 70% of the spatial stimuli were recalled correctly. Using this method of computation, the rate of correct responses (CR) was computed for each patient.
Patient MRI data acquisition
All patients underwent Magnetic Resonance Imaging on a 3-T X-series Philips Achieva clinical MRI scanner (Amsterdam, the Netherlands) using an eight-channel head coil. Anatomical and functional acquisitions were similar for all patients. Single-shot echoplanar gradient echo imaging sequence acquiring T2* signal was used for the BS task, with the following parameters: n volumes (depending on the patient's span, min-max=[192–288], median=240), 36 axial slices acquired parallel to the AC-PC line, TR=2.5 s, TE=35 ms, FOV=256 mm, 128×128 data matrix isotropic voxels, and flip angle=90°. In addition, after the BS task, patients underwent a 5 min resting-state condition, with the following parameters: 120 volumes, 34 axial slices acquired parallel to the AC-PC line, TR=2.5 s, TE=35 ms, FOV=256 mm, 128×128 data matrix isotropic voxels, flip angle=90°, and bandwidth=1.802 (±241.1 kHz). The patients were instructed to rest with eyes closed and refrain from any voluntary motion. For both functional tasks, the in-plane resolution was 2×2 mm2, and the slice thickness was 4 mm. Before collection of the T2* images, T1-weighted images (180 slices) were collected using an MPRAGE sequence (256×256 isotropic voxels; TR=640 ms, TE=3.2 ms, FOV=256 mm, and flip angle=8°) in positions identical to the functional scans to provide an anatomical reference. The in-plane resolution for each T1 slice was 1 mm2. Each EPI imaging series started with three discarded scans to allow for T1 signal stabilization.
Preprocessing analyses
BS and resting-state fMRI data from the TLE patients were preprocessed using SPM8 (
Definition of the seed regions
Seed regions were restricted to voxels within the left and the right hippocampus, respectively. Their spatial standardized definitions were taken from the normalized template available in the automated anatomical labeling atlas (Tzourio-Mazoyer et al., 2002), provided as a toolbox in SPM8. Data analyses within each patient included calculation of the mean signal time course over the whole experiment in each seed. This step was done for both the BS and the resting-state tasks.
We considered the pathologic hemisphere as the one including the epileptogenic source, based on the EEG recordings. Accordingly, analyses involving the right hippocampus seed for the RTLE group will be referred to as the pathologic seed, regardless of the presence of MTS. Conversely, the analyses involving the left hippocampus seed for the LTLE group will be referred to as the pathologic seed, regardless of the presence of MTS.
Statistical analyses for the BS task
We only considered the encoding phases for each subject, excluding the recalling/response phases. For this, we concatenated the volumes associated with the 24 encoding periods in one single sequence and realized the next analyses (described below) on it.
At the individual level and for each seed region, a correlation map was produced by computing the correlation between the mean signal time course in the seed region and the time course from all other brain voxels. Next, this matrix of correlation coefficients was submitted to a Fisher r-to-z transformation [Z(r)], yielding an approximate normal distribution for the sampled data. All second-level statistical analyses were conducted on these transformed data. This aim of this step was to extract the network emergent from the hippocampus seed during the encoding phase, at the individual level.
In order to reveal this network at the group level, individual Z(r) values maps were entered into a second-level random-effects analyses. We first determine whether differences in FC existed between the right and left TLE groups. For this, we performed two-sample t-tests to detect these differences, independently for each seed. We fixed a height threshold at p<0.001 (uncorrected), and the cluster-level threshold was set at a corrected level of p<0.01.
Next, we investigated whether the hippocampus-seeded network connectivity was modulated by the patient's performance (e.g., the rate of CR). To accomplish this, we explored the regions of the hippocampus-seeded network that covaried with the BS performance within each of our patient groups (LTLE, RTLE) during the encoding phase. To do this, these individual Z(r) values maps were entered into a GLM, using SPM8. The (unthresholded) Z(r) maps were entered into a one-sample t-test design with two additional covariates: the BS performance as the covariate of interest and the patient's span value (e.g., reflecting the task duration) as a covariate of no-interest. Both covariates were continuous values. The covariate of no-interest was only used to correct for the different task durations between the patients.
Finally, for each patient group and each seed, we computed two contrasts highlighting the regions of the encoding-related network that are either positively or negatively modulated by the patients' BS performance, respectively. Overall, within each patient group, this analysis resulted in spatial maps, highlighting the regions of the hippocampal network that have their FC significantly covarying with patients' performance. The height threshold was fixed at p<0.001 (uncorrected; i.e., T>3.4), and the cluster-level threshold was set at a corrected level of p<0.01. It should be noted that we excluded the hippocampal seed from these analyses.
Due to the nonhomogeneity of our patient samples regarding the presence of MTS, we also checked that the significant effects were not caused by the presence of MTS using two-sample t-tests (patients with MTS versus patients without MTS, within LTLE or RTLE group).
Statistical analyses for the resting state data
The resting-state data were used as a control condition of the BS task. In other words, using the same statistical model in SPM, we checked to see whether a hippocampus-seeded resting-state network was modulated by BS performance. Any such modulation would have indicated that the FC network which we had observed during the encoding phase of the BS task was not specific to that task, representing perhaps a more general effect. The only difference within the statistical model was the noninclusion of the patient's span value as a covariate, because the duration of the resting state was similar for all patients.
All other statistics were realized using the software IBM® SPSS® v19.
Results
Behavioral data
RTLE and LTLE patient groups did not differ by age or gender (p>0.2). With regard to the BS task, there was also no significant difference for either the span or the rate of CR between the groups (span: p>0.6; CR: p>0.5). Across all the patients, the average span was 5±1 (Table 1). In addition, the two TLE groups did not differ by medication type or number of medications.
BS functional data
Seed in the right hippocampus
A direct comparison of the network emerging from the right hippocampus between the TLE groups did not reveal FC significant differences. The major regions of this network (FWE, p<0.05) involved the ipsilateral MTL, including the parahippocampal gyrus, hippocampus, and fusiform gyrus. The contralateral parahippocampal gyrus was also a part of this network. At a less stringent threshold (uncorrected, p<0.0001), this network extended to the bilateral fusiform gyri, the bilateral MTLs, the ipsilateral cerebellum, and temporal pole.
The investigation of the FC modulation by the BS performance within this right-sided network revealed significant results for the LTLE group only (Table 2). In detail, the FC of one specific region located in the crus I of the right ipsilateral cerebellum was negatively correlated with the rate of CR, for the LTLE group (T=5.8, Ke=161; Fig. 2). In other words, the LTLE patients with better performance in the BS task showed decreased FC between the right hippocampus and the right cerebellar cluster during the encoding phase. No differences were found when comparing LTLE patients with and without MTS.
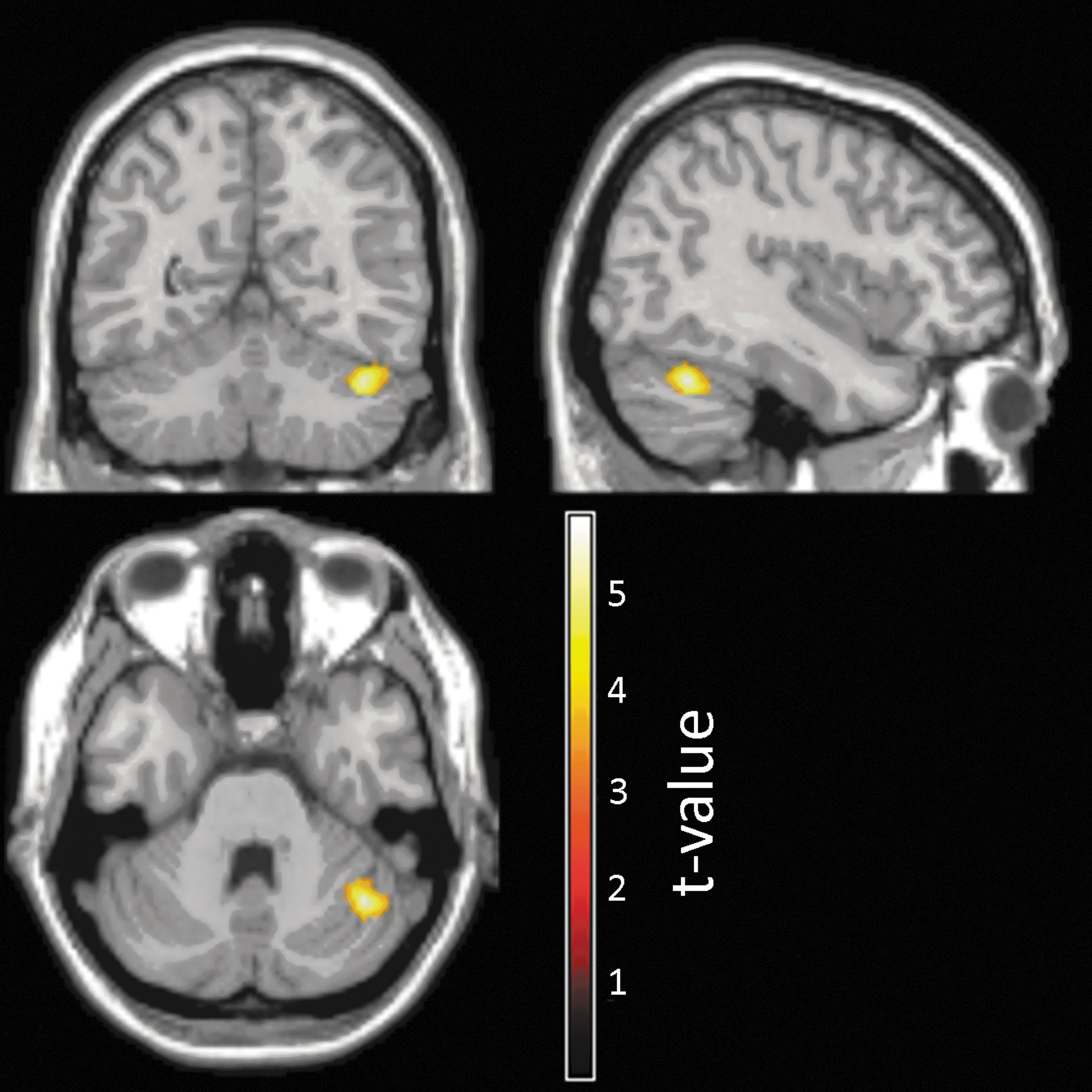
Mapping of the cerebellar cluster demonstrating a negative relation between their FC values with the right seed and patient performance in the LTLE group. FC, functional connectivity; LTLE, left temporal lobe epilepsy.
Regions Showing a Positive Functional Connectivity with the Right Hippocampal Seed and Correlated with the Rate of Correct Responses During the Encoding Phase of the Block Span Task
FWE, family wise error; CR, correct responses; R, right.
We did not find regions that were also positively correlated with the rate of CR for either patient group.
Seed in the left hippocampus
For the left hippocampus seed, a direct comparison of the spatial maps did not reveal significant differences between the TLE groups. The major regions that correlated with the seed involved the ipsilateral and contralateral MTLs, including the parahippocampal gyrus and hippocampus (FWE, p<0.05). At a less stringent threshold (uncorrected, p<0.0001), this network extended to the bilateral fusiform gyri, the ipsilateral cerebellum, and temporal pole.
We found several modulations of the network by the rate of CR, for each patient group (Fig. 3, Table 3). For the RTLE group, a bilateral cluster, including the precuneus, had increased positive FC with the left (nonpathologic) seed in association with a better rate of CR (T=4.9, Ke=332; Fig. 3A, left panel). In contrast, for the LTLE group, no positive modulations with the rate of CR were evident.
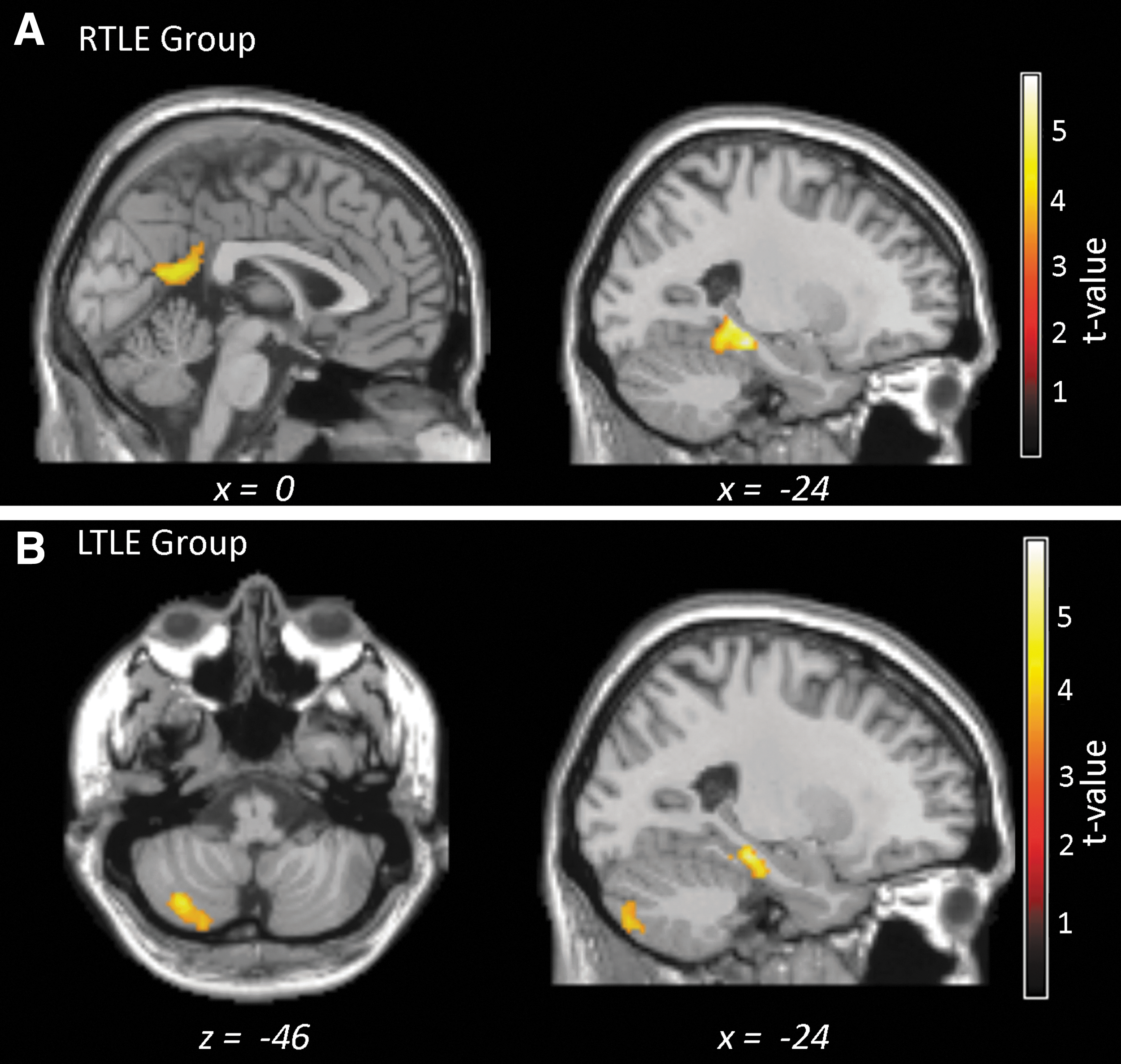
Mapping of the clusters demonstrating a significant relation between their FC values with the left hippocampal seed and patient performance (CR).
Regions Showing a Positive Functional Connectivity with the Left Hippocampal Seed and Correlated with the Rate of Correct Responses During the Encoding Phase of the Block Span Task
L, left; R, right; gy, gyrus.
In both TLE groups, we found that one region located in the left parahippocampal cortex was associated with decreased FC with the left hippocampus and a better rate of CR for both groups (RTLE: T=6.4, Ke=345; LTLE: T=5.7, Ke=239; Table 3). It should be noted that the clusters for the two patient groups do not overlap (Fig. 3, right panel). In the RTLE group, the cluster was more posterior than the one revealed for the LTLE group. Furthermore, as for the right seed, a cerebellar cluster (Lobule VII B) was revealed ipsilateral to the seed, showing a negative relation between its FC with the left hippocampus and the rates of CR in the LTLE group (T=4.6, Ke=239; Fig. 3B).
No difference was found between the patients with and without MTS.
In summary, regardless of the seed, our data showed that in the LTLE group the FC between both seeds and the ipsilateral cerebellum bore a negative relation with BS performance. Both right and left TLE patient groups showed a significant negative association between FC and BS performance, with this effect involving the left posterior MTL (parahippocampal gyrus, extended to the fusiform gyrus) and the left hippocampus. Finally, only one positive modulation with the performance was highlighted, including the left (nonpathologic) hippocampus and the precuneus in the RTLE.
Resting-state functional data
The resting-state data, collected separately from the BS task, were analyzed and used as a control condition. No significant correlations were observed between the rate of CR and the regions noted earlier (i.e., found to have their FC values significantly correlated with the CR rate during the encoding phase of the BS task). This analysis reinforces the idea that the FC findings observed during encoding of the BS task are quite specific to the cognitive processes present at that time.
Discussion
This study demonstrates that computing FC emerging from each hippocampus during the encoding phase of a WM task is an efficient method for investigating the WM system in TLE, particularly with regard to the differing effects of lateralized seizure pathology. Although the task generally produced low performance levels in the patients, we showed that FC values between the hippocampus and specific cortical and cerebellar regions were modulated by visuospatial WM performance during the encoding phase of the task.
Although the global functional networks emerging from hippocampi were not significantly different between the TLE groups, we revealed that distinct parts of these networks were modulated, in both different locations and directions, by the visuospatial performance in each group. In greater detail, the LTLE had only negative effects that mostly involved the FC between each seed and ipsilateral cerebellar cluster; while the RTLE showed increased FC between regions that were highly involved in memory circuits, the left nonpathologic hippocampus, and the precuneus, as well as decreased FC with the nonpathologic hippocampus with improved memory performance. In contrast, most of the negative effects in the LTLE involved the (left) pathologic hippocampus, suggesting that shutting down a pathologic network may help memory encoding. However, we also revealed a common effect in both patient groups: The FC between the left hippocampus and the ipsilateral parahippocampal gyrus was negatively modulated with the performance.
Overall, we highlighted that the functional modulation during WM performance only involved regions located in the MTL, medial posterior parietal cortex, and the cerebellum. Therefore, our results support previous neuropsychological and functional studies describing the hippocampus (Lee and Rudebeck, 2010; Stretton et al., 2012; Wagner et al., 2009) as modulating spatial WM function through a network involving parahippocampal gyrus (Shankar et al., 2011) and precuneus [see review (Kravitz et al., 2011)]. Importantly, the present findings confirm that analyses of FC emerging from hippocampi during cognitive tasks can provide a valuable and precise complement to standard analyses of subsequent memory effects.
More specifically, we found that the left hippocampus and parahippocampal gyrus were less synchronized when the RTLE or the LTLE patients had better performance. This finding was not significant for the right hippocampus. In other words, our data suggest that the left hippocampal and parahippocampal gyri had a tendency to work in opposite directions in order to support better visuospatial WM process in TLE patients, regardless of the side of the pathology. Thus, the present results suggest that right and left TLE negatively impact the FC between the left MTL regions in similar manner, perhaps related to the fact that the left MTL system may well represent the side not dedicated, in a material-specific sense, to visuospatial memory processing in both patient groups. However, our results also point out that different regions of the left parahippocampal gyrus were involved in such processing for the RTLE and LTLE groups, with the RTLE patients demonstrating a more posterior involvement. This difference may be related to the pathology side, but the exact reason for this anterior/posterior difference is unclear. It is important to note that even though all the LTLE patients did not present a left MTS, it is likely that the left hippocampus remains more pathologic in the LTLE than in the RTLE patients, given its proximity to seizure activity. Overall, one direct consequence of the association between reduced FC and better performance is that it may illustrate how reduced engagement of the medial temporal regions may be needed in order to optimize cognitive task performance in TLE.
Other additional cognitive mechanisms may be at work in the patient groups, highlighting differences between them. In fact, in RTLE we found the only positive relationship between FC and better performance. This was between the precuneus and the left (nonpathologic) hippocampus. This result is concordant with both functional studies describing the involvement of the precuneus during visual memory processing (Mason et al., 2007; Ranganath et al., 2005) and anatomical studies showing that precuneus and hippocampi have direct anatomical connections (Greicius et al., 2009). The same results have been described in healthy participants by Ranganath et al. (2005), indicating that successful memory formation is associated with transient increases in cortico-hippocampal interaction. Overall, our results further confirm the modulatory role of the precuneus as a part of a functional network supporting spatial WM processing in the right TLE patients. Furthermore, the hippocampus and precuneus have been described as a part of the well-known default-mode network (DMN) (Buckner et al., 2008), making our results consistent with previous studies describing the modulation of the DMN activity with WM processing (Esposito et al., 2009). Thus, the present data suggest that the functional connection between hippocampus and precuneus may not be impaired in right TLE, implying that other cognitive functions or processes dependent on these linked areas may have integrity.
Overall, we suggest that the coupling between the reduction of FC between left MTL regions and the increased FC between the left hippocampus and the precuneus with performance may be a sign that a complex coordinated network is necessary to support optimal visuospatial WM in the face of right temporal lobe pathology. Indeed, the right hippocampus may be impaired in these patients, and the left hemisphere may be needed to generate compensatory activity to support high-level cognitive tasks. In contrast, optimized WM in the setting of LTLE may involve solely decreased FC with better performance.
The LTLE showed that their functional connections between both hippocampi and ipsilateral cerebellum were reduced with better performance during the BS task, regardless of the side of the seed. This suggests that cerebellar involvement represents reorganization of WM functions in left but not right TLE, though this is speculative and would require comparisons with normal controls in order to be confirmed. The cerebellum has been described as involved in motor activities such as motor control, including motor inhibition and motor learning (Tracy et al., 2001). Thus, based on earlier literature, we suspect that the cerebellar involvement we observed is a sign of motor control, as the patients did have to refrain from pressing the buttons during the encoding phase. This explanation is consistent with the absence of such cerebellar modulation at rest.
Finally, it is worth noting that despite distinct functional patterns of connectivity between right and left TLE, the patients did not significantly differ in their performance in the BS task. This suggests that different cognitive mechanisms, involving different regional implementation, may be similarly efficient (or inefficient, as both showed relatively weak performance on the task) between the patient groups. In this sense, our project contributes to the literature showing that FC-based methods may be an alternative for investigating brain function (or dysfunction) during a cognitive task. Indeed, exploring FC may be a particularly valuable and efficient technique for investigating cognitive networks in patients demonstrating cognitive difficulties and weak performance levels.
The major limitation of this study is the lack of a healthy control group. Nevertheless, the present results highlighted statistical differences in FC patterns within the MTL system between right and left TLE patients. Such comparisons remain valid without a control group, and to the best of our knowledge, no previous studies have described such results in TLE. The weak performance level of the patients in the BS task is another concern. Our results reflect a network response during visuospatial encoding and WM that was not highly accurate. It remains for future projects to determine how FC during such WM processes varies as a function of different performance levels. Finally, the fact that we concatenated, for each subject, the functional volumes associated with the encoding phase, a concern might arise regarding false FC values related more strictly to each, unique encoding phase rather than the point-by-point changes common during encoding across all the encoding phases. To better understand the role played by low-frequency changes in our data, that is, isolated encoding phase effects, we checked the concatenated time series for each subject and were not able to distinguish between unique and unusual phase changes. We should also note that temporal concatenation is a common method used for independent component analyses when subjects have had several fMRI sessions (similar to the present study with several encoding periods for each subject). Such session changes have not been reported (e.g., Erhardt et al., 2011; Guo et al., 2012). In addition, using resting-state data, Fair et al. (2007) showed that FC computed on concatenated data are both qualitatively and quantitatively very similar to FC computed on one “continuous” dataset, suggesting that our results remain valid despite this temporal concatenation.
We reiterate that we believe the value of this study is in showing that FC can help elucidate the networks implementing important cognitive processes, which, regardless of accuracy for the task at hand, accurately reflect the brain's response to specific types of cognitive challenge. As such, FC holds great promise for identifying the underpinnings of flawed and deficient cognition in impaired individuals.
Conclusion
To the best of our knowledge, we are among the first to highlight a direct relation between visuospatial WM performance and hippocampal activity during the encoding phase, in TLE patients using an FC-based method. We demonstrated that distinct patterns of hippocampal FC may support visuospatial WM processing in unilateral TLE patients. Importantly, this finding points to the possibility that unilateral TLE causes distinct patterns of functional network responses according to the side of epileptic seizures. We believe our data raise the possibility that patients with temporal lobe seizures implement distinct neuroplastic responses in the face of differently lateralized pathology. The end result is that right and left TLE take on different patterns of FC in order to execute spatial WM. Our patient groups demonstrated no behavioral differences in the visuospatial WM task, yet exhibited different brain response modularity patterns in association with performance. Accordingly, our results indicate that FC may capture potential or accruing problems in visuospatial WM in TLE. Indeed, our findings suggest that FC may provide a unique means of identifying abnormalities in brain networks, which cannot be discerned at the level of behavioral output through techniques such as neuropsychological testing. In contrast to resting-state studies which assume that FC reflects functional integration (or segregation) and, more generally, cognitive activity, the present study has investigated and captured a more direct relation between FC and cognitive performance. Indeed, it is important to note that our analyses of the resting-state condition did not reveal the key performance-related FC relationships which we discussed earlier, suggesting that there is value added when investigating FC during a task-based condition.
Footnotes
Acknowledgment
This work was supported, in part, by the National Institute for Neurological Disorders and Stroke (NINDS) [grant number R21 NS056071-01A1] to Joseph I. Tracy.
Ethical Publication Statement
The authors confirm that they have read the journal's position on issues involved in ethical publication and affirm that this article is in line with those guidelines.
Author Disclosure Statement
None of the authors have any conflict of interest to disclose.