
Abstract
Implicit probabilistic sequence learning (IPSL) involves extracting statistical regularities from sequences of events without awareness, and is thought to underlie learning of language and behavioral repertoires of everyday life. We examined whether resting-state functional connectivity networks of the caudate predicted individual differences in IPSL performance measured on a separate day. Whole-brain connectivity maps of a bilateral dorsal caudate (DC) seed were created for each subject and examined for voxelwise correlations with sequence learning performance, as well as with overall response speed. Higher learning scores (but not overall response speed) were associated with stronger resting-state connectivity between the DC and right medial temporal lobe, as well as with lower resting-state connectivity between the DC and premotor regions involved in motor planning. Thus, how well one learns probabilistic regularities without awareness is predicted by the strength of a striato-cortical network in the resting brain.
Introduction
Implicit learning refers to the effortless acquisition of information without explicit knowledge of what has been learned (Reber, 1989). Implicit probabilistic sequence learning (IPSL) is a specific type of implicit learning that involves extracting statistical regularities from sequences of events, a skill which is crucial for important life skills such as learning languages or developing new routines (Lewicki et al., 1987; Newport and Aslin, 2004). Here, we focus on how the intrinsic functional communication among neural regions known to underlie IPSL relates to individual differences in learning performance.
To accomplish this, we used the Triplets Learning Task (TLT) (Howard et al., 2008). In the TLT, as in more traditional sequence learning (SL) tasks such as the serial reaction time (RT) (Nissen and Bullemer, 1987) and Alternating serial RT (Howard and Howard, 1997) tasks, participants are exposed to a series of stimuli in which a probabilistic regularity is embedded, such that some events are more predictable from earlier events than others. During the task, subjects learn the regularity, responding increasingly faster to more versus less predictable events, even though they are unable to distinguish between them in subsequent recognition tests (Howard et al., 2008; Simon et al., 2011a, 2011b). The TLT allows for precise control of event timing and reduces motor-response sequencing, enabling us to examine implicit learning uninfluenced by motor fluency.
Evidence from studies using a variety of methods (including patient groups, diffusion tensor imaging [DTI], task-related functional magnetic resonance imaging [fMRI], and genetics) indicates that IPSL in the TLT and related SL tasks depends on the functional and structural integrity of a subcortical-cortical network, including the caudate, frontal cortices, and, potentially, the medial temporal lobes (MTL) (Bennett et al., 2011; Gheysen et al., 2011; Rieckmann et al., 2010; Rose et al., 2002; Schendan et al., 2003; Simon et al., 2011b, 2012; Smith and McDowall, 2004; Wilkinson, et al., 2009). While frontal regions are thought to support the attentional and motor-planning demands of IPSL tasks, the caudate and, more controversially, the MTL may underlie the formation of associations that are necessary for learning in the tasks (Rose et al., 2002; Schendan et al., 2003; Simon et al., 2012). These latter regions are the focus of the present study.
The fact that the caudate and MTL, regions once thought to subserve distinct and functionally incompatible learning systems (e.g., Robbins, 1996; Squire, 1987), are sometimes coactivated in implicit SL tasks suggests that optimal SL performance may depend on the efficiency of their interaction (Henke, 2010). This would be the case even if such interactions are competitive, as a number of task-related fMRI studies suggest (e.g., Albouy et al., 2008). Efficient caudate/MTL interactions—whether competitive or cooperative—may be facilitated by their ongoing communication not only during IPSL tasks, but also during a task-free state. Patterns of temporally correlated brain activity at rest (termed intrinsic, or resting-state functional connectivity [rsFC]) often mirror the brain activation patterns observed during cognitive tasks (Smith et al., 2009), suggesting that functional networks during cognitive tasks maintain their network configuration during rest [for review, see (Fox and Raichle, 2007)].
Close correspondence between task-evoked and resting functional networks also suggests that individual differences in network functions at rest should be associated with learning performance. Indeed, rsFC of task-relevant networks relates to individual differences in the consolidation of learned information, such that individuals with superior performance on a preceding learning task display the greatest changes in rsFC from a pre- to postlearning resting scan (e.g., Albert et al., 2009; Vahdat et al., 2011; Wegman and Janzen, 2011). In addition, the baseline (i.e., pretask) strength of rsFC in task-relevant networks can predict individual differences in subsequent performance, by which individual differences in the strength of selective resting-state networks relate to learning and memory ability (Baldassarre et al., 2012; Gordon et al., 2013; Wang et al., 2010a, 2010b; Ystad et al., 2010). No studies so far, however, have examined whether rsFC can predict learning performance when subjects are not explicitly informed of task goals and are learning probabilistic relationships that they cannot verbalize.
Here, we investigate whether individual differences in IPSL can be predicted by the rsFC of a task-relevant region, the caudate. We measured rsFC of a bilateral caudate seed either before learning or well outside the timeframe when consolidation might be expected to influence intrinsic connectivity patterns. We then correlated rsFC with SL performance, as well as with average response speed on the TLT. Our primary interest was in the relationship between rsFC and sequence-specific learning, but we examined average response speed to test whether the striato-cortical networks supporting sequence specific learning are distinct from those supporting other aspects of task performance. We examined whole-brain caudate connectivity but were particularly interested in correlations between the caudate and MTL, given evidence that these regions interact during IPSL tasks in healthy populations. We hypothesized that greater positive correlations in the spontaneous activity of the caudate and MTL at rest (indicating stronger intrinsic communication between these regions) would predict better SL, but not response speed.
Materials and Methods
Subjects
Twenty-two Georgetown University undergraduates (14 female) aged 18 to 22 years (M±SD=20.43±0.98) participated for payment. They were recruited from a pool of 50 subjects in an unrelated experiment in which resting-state scans were acquired. As a result of this recruitment procedure, behavioral testing occurred on a varying number of days (17–367 days; M±SD=96±102 days) after the scan. In addition, two subjects who had completed behavioral testing 179 and 542 days before the scan were retained, because the long delay made it highly unlikely that learning-related consolidation influenced their resting-state connectivity. Correlational analysis confirmed that the time between the scan and behavioral testing was unrelated to the behavioral measures. Furthermore, our results did not change when these two subjects were removed.
Procedures were approved by Georgetown University's Institutional Review Board. Exclusion criteria included (1) self-reported use of psychotropic medication (e.g., stimulants, anti-anxiety/depression); (2) self-reported history of neurological injury or disease, seizure disorder, psychiatric diagnosis; and (3) contraindications for MRI—for example, metal implants in the body, or pregnancy.
Behavioral testing
Subjects completed the TLT (Howard et al., 2008) on a separate test day either after (N=20; M±SD=96±102; ranging from=17–367 days) or preceding (N=2; M±SD=360±256 days) the scanning session. A schematic of the TLT is shown in Figure 1. Subjects viewed a horizontal row of four open circles centered on a computer screen. In each trial, a three-event sequence of circles (a “triplet”) was filled in sequentially red, red, and then green. Subjects were instructed to observe the first two red “cues” and to indicate the location of the green “target” by pressing a spatially corresponding button as quickly as possible with their right hand. Cues were displayed one after the other for 120 ms each (150 ms inter-stimulus interval) and were followed by the target, which remained in view until participants made a correct response. The next trial began after a 650 ms delay. Repetitions (e.g., 111) and trills (e.g., 121) were not presented, because they have strong pre-existing response tendencies (Boyer et al., 2005; Cleeremans and McClelland, 1991; Howard et al., 2008). Without being known to subjects, the TLT contained a probabilistic regularity such that for a randomly chosen set of 16 triplets the target was likely to follow the cues (High Probability triplets, p=0.90) whereas for the remaining 32 triplets, the target was unlikely (Low Probability triplets, p=0.10) (Simon et al., 2011a).
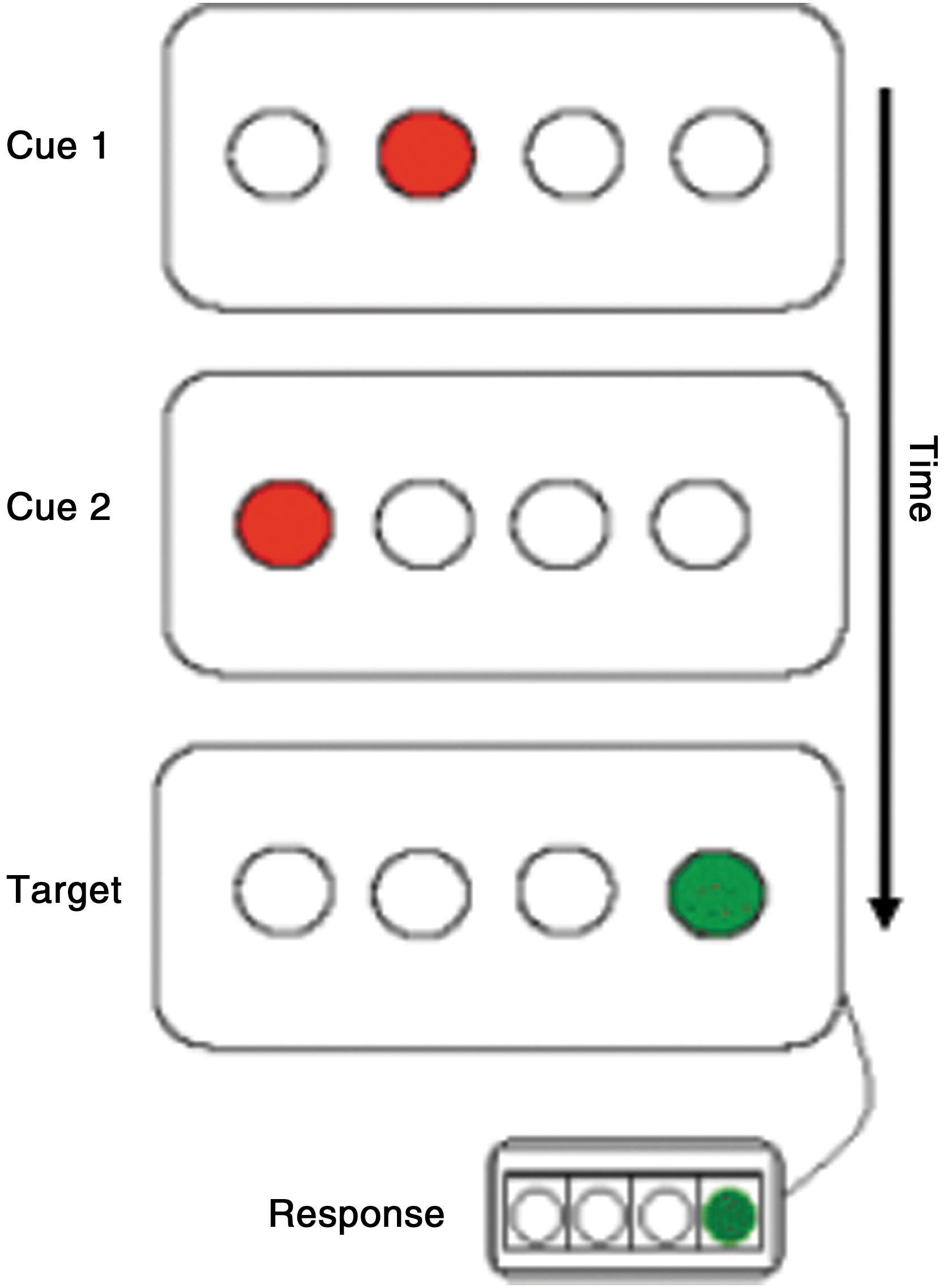
Schematic of the Triplets Learning Task. Each trial, or “triplet,” consists of the sequential presentation of two red cues and a green target. Subjects are instructed to view the first two red cues of each triplet and to respond only to the location of the green target using a corresponding response button.
Subjects completed 750 trials in ∼30 min. Short breaks occurred after every 50-trial block, during which the subject's mean RT and accuracy were displayed along with instructions to “focus more on speed,” “focus more on accuracy,” or “speed and accuracy are about right.” The instructions were based on the subject's mean accuracy for the preceding block of trials and were intended to drive all subjects to 92% accuracy.
Calculating SL scores
As in previous studies using the TLT, we used SL scores, which measure learning unbiased by individual or group differences in overall RT (Howard et al., 2008; Simon et al., 2011a). To determine these scores, median RTs were calculated for all correct responses for each triplet for each subject (overall accuracy in the task was high, ∼93%, so we omitted a few trials). These median RTs were then correlated with the actual triplet frequencies for that subject. Subjects who show greater sequence-specific learning display greater negative correlations between RT and triplet frequency, as responses are faster to triplets that occur with greater frequency and, thus, have more predictable targets. For ease of interpretation, the correlations were multiplied by −1 so that higher SL scores reflect greater SL.
fMRI data acquisition
Subjects were scanned for 5:04 min during the resting state, in which they were told to relax with eyes closed but to stay awake and to not think of anything in particular. All subjects verbally confirmed that they had remained awake for the duration of the scan. Imaging was performed on a Siemens Trio 3T scanner (Erlangen, Germany). For the resting scan, 152 whole-brain images were acquired using a gradient echo pulse sequence (37 slices, TR=2000 ms, TE=30 ms, 192×192 mm FOV, 90 degree flip angle, and voxel dimensions 3 mm isotropic). The first 4 images of this run were discarded to allow for signal stabilization. This scan was followed by acquisition of a high-resolution T1-weighted structural scan (MPRAGE) lasting 4:18 min (TR/TE=2300/2.94 ms, TI=900 ms, 90 degree flip angle, 1 slab, 160 sagittal slices with a 1.0 mm thickness, FOV=256×256 mm, and matrix=256×256, resulting in an effective resolution of 1.03 mm isotropic voxels).
Image preprocessing
Using SPM8 (Wellcome Department of Cognitive Neurology, London, United Kingdom) implemented in MATLAB (Version 7.10; Mathworks, Inc., Sherborn, MA), images were corrected for translational and rotational motion by realigning to the first image of the resting-state scan. All subjects demonstrated less than 1.0 mm of translational motion in any one direction (max translation=0.94 mm) and less than 0.5° of rotation around any one axis (max rotation=0.35°). Since micro-movements have been shown to lead to spurious correlations in previous functional connectivity studies (Power et al., 2012), framewise displacement (M±SD=18±0.08 mm) was included as a regressor of no interest in all group-level analyses reported next, a procedure recommended to reduce motion artifact while preserving the power of the study's design and reducing the likelihood of Type II error (Satterthwaite et al., 2012; Van-Dijk et al., 2012). Images were slice-time corrected, normalized to an EPI template, and smoothed using a Gaussian kernel with full width at half maximum of 8 mm. For normalization, all functional images for each subject were realigned to the first image using 2nd-degree B-spline interpolation. This image was then used as the source image, which was transformed to a standard SPM-EPI template in Montreal Neurological Institute (MNI) atlas space available in SPM8. The parameters used to transform the source image to the template were then applied to all functional images. The quality of the registration process was assessed by visual inspection.
Finally, a band-pass filter of 0.01–0.1 Hz was applied to the data to remove non-neuronal temporal trends and to restrict signal variation to the frequency range established in the literature for fluctuations in resting-state data (Biswal et al., 1995).
Functional connectivity calculation
Seed region-of-interest creation
A bilateral dorsal caudate (DC) seed region of interest (ROI) based on coordinates from Di Martino et al. (2008) was created using Marsbar (Brett et al., 2002) as two spheres of radius 6 mm centered around the coordinates [13 15 9] and [−13 15 9]. The seed location is depicted in Figure 2. Following Di Martino and colleagues, we chose our seed coordinates to be consistent with findings from a meta-analysis of fMRI and positron-emission tomography human neuroimaging studies (Postuma and Dagher, 2006), which demonstrated that there are functionally distinct anatomical regions within the human striatum. Specifically, dorsal regions of the caudate (defined as those where z>7 mm) were found to be more coactivated with cortical regions involved in cognitive processes, while ventral regions are coactivated with cortical regions underlying limbic and motivational functions. In addition, this seed is consistent with a previous event-related fMRI study using the TLT, which reported greater task-dependent activation in dorsal regions of caudate body in response to sequences occurring with high versus low probability (Simon et al., 2012).

Location of the bilateral dorsal caudate (DC) seed. The seed was created as two spheres centered around coordinates [13 15 9] and [−13 15 9].
Nuisance signal identification
To identify the effects of motion and physiological noise (i.e., respiration, heart rate), time series approximating these signals were calculated for the resting-state scan. Physiological noise was approximated by obtaining signal time series from white matter and cerebrospinal fluid segmentations of the MPRAGE image (Van-Dijk et al., 2010). Motion was obtained from the six realignment parameter time courses from the motion correction preprocessing step. Global signal regression was not performed as a preprocessing step, because studies have shown that this procedure can artificially induce negative correlations in functional connectivity data (Murphy et al., 2009).
Voxelwise rsFC calculation
For each subject, partial correlations were conducted between the bilateral DC seed time series and the time series of every voxel in the brain, while partialling out the motion and physiological noise time series. Each subject's individual connectivity maps were visually inspected in order to confirm that they were not corrupted by motion artifact. The resulting r-values were converted to normally distributed Z-scores using Fisher's transformation to allow further statistical analysis of correlation strengths. This produced a brain map of intrinsic connectivity strength with the DC during rest for each subject.
Overall connectivity
We identified overall patterns of connectivity with the DC by entering subjects' individual connectivity maps into a voxelwise one-sample t-test using SPM8. All group-level results were restricted to gray matter using an explicit gray matter mask from the SPM toolbox. Monte-Carlo-based correction for multiple comparisons (Ward, 2000) was carried out using tools implemented in Analysis of Functional Neuroimages (AFNI) software (Cox, 1996). Briefly, we estimated the smoothness of the preprocessed data using the 3dFWHMx utility, and then used the 3DClustSim tool to conduct 10,000 Monte-Carlo simulations of random noise activations with that same smoothness, within the same whole brain volume as the primary analyses. These simulations revealed that clusters exceeding both a voxelwise alpha level of p<0.005 and a size of 100 voxels occurred less than 5% of the time, corresponding to a corrected p<0.05 cluster-level significance threshold.
Connectivity correlations with SL scores and response speed
The single-subject whole-brain maps of DC connectivity strength were entered as the dependent variable into two regressions testing for correlations with subjects' SL scores and average response speed, respectively, in a voxelwise fashion. Average response speed was calculated by determining each subject's median RT for all correct responses in each 50-trial block, and then averaging across blocks to obtain a single mean RT value for each subject. Results were corrected for multiple comparisons at p<0.05 using the Monte-Carlo correction threshold described earlier.
Results
Sequence learning performance
Subjects' SL scores indicated that they had become sensitive to the probabilistic regularity in the task: An independent sample t-test confirmed that overall SL scores (M±SD=0.19±0.08) were significantly greater than zero, t(21)=10.81, p<0.0001. As predicted, subjects also responded significantly faster on average to high probability (M±SD=326±32 ms) than to low probability (M±SD=343±35 ms) triplets, t(21)=7.8, p<0.0001.
Importantly, individual SL scores did not correlate with subjects' overall RT or accuracy, indicating that our learning measure was not biased by individual differences in speed or accuracy. Further, overall mean accuracy (M±SD=0.93±0.03) indicated that the end-of-block feedback was successful at driving subjects to respond with ∼92% accuracy.
DC connectivity networks
Mean resting-state connectivity networks of the DC observed across all subjects are shown in Figure 3 and Table 1. Connectivity patterns were similar to those observed by Di Martino et al. (2008) using the same seed.
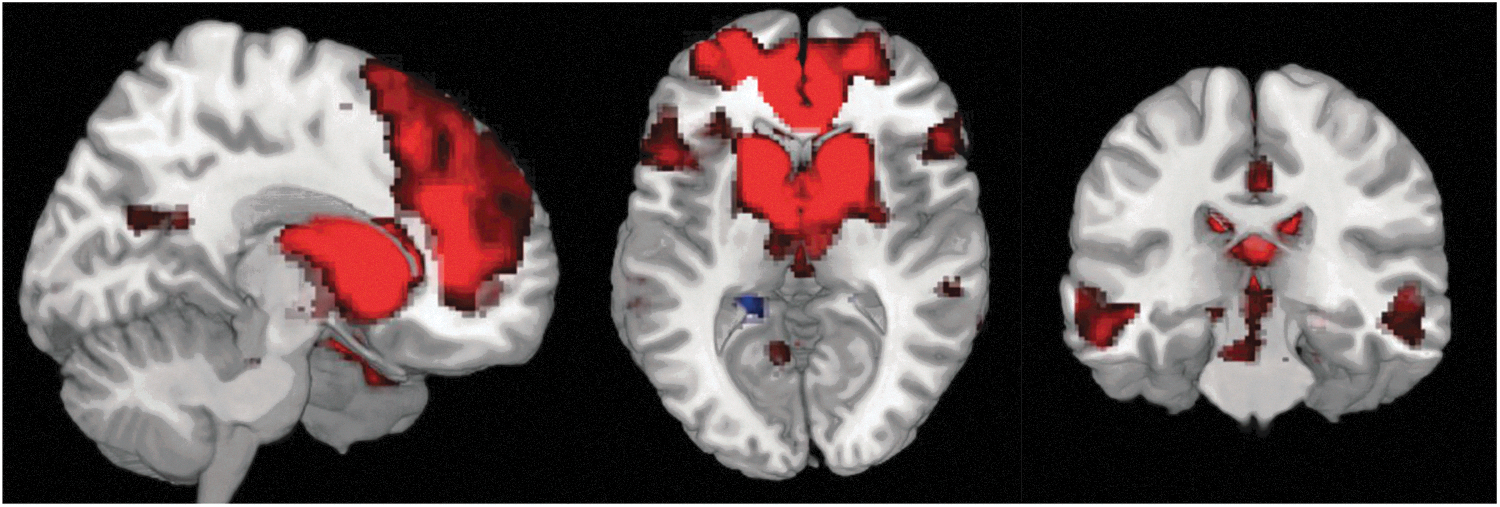
Resting state functional connectivity (rsFC) network associated with a bilateral DC seed across all subjects. Red represents positive connectivity, and blue represents negative connectivity. Results are corrected for multiple comparisons at p<0.05.
Whole-Brain Connectivity of the Dorsal Caudate Seed
MNI, Montreal Neurological Institute.
Correlations between DC connectivity and SL scores
Consistent with our hypothesis, voxelwise regression using individuals' SL scores as the predictor resulted in a positive correlation with a cluster in the right parahippocampal gyrus extending into the right hippocampus (k=117 voxels; peak MNI coordinates [38 −30 −14]; r=0.74, p<0.0001), such that subjects who had greater connectivity between the DC and this region tended to show greater learning (Fig. 4). Notably, this was the only positive correlation between whole-brain intrinsic connectivity of the DC and SL scores to survive correction. This correlation occurred despite the fact that the average value of intrinsic connectivity between the DC and this region was close to zero at the group level (M±SD=−0.043±0.19).
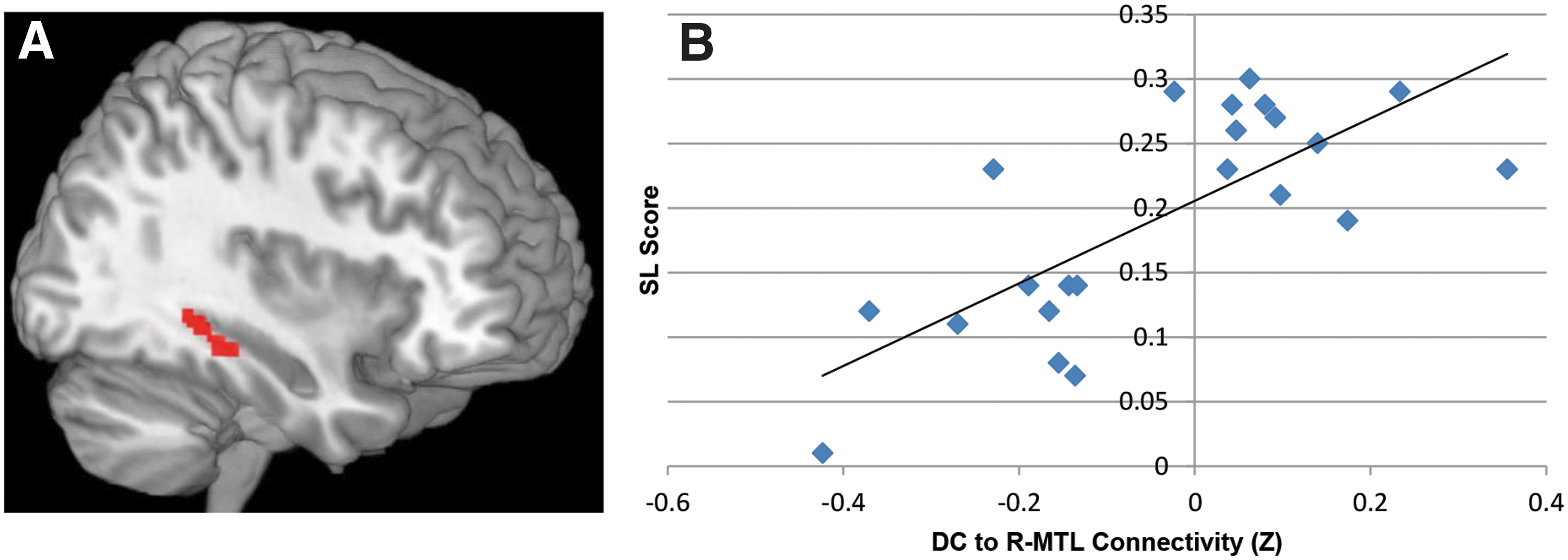
Positive correlation between rsFC of the DC seed and sequence learning scores.
Follow-up analysis revealed that the positive correlation between DC connectivity and SL scores was specifically driven by the right DC. We separately calculated the whole-brain connectivity maps of the left and right DC, and entered these as the dependent variables into two regressions testing for correlations with SL scores in a voxelwise fashion. As in the analysis using the bilateral DC connectivity maps, the regression using the right DC connectivity maps revealed a positive correlation with a similarly located cluster in the right parahippocampal gyrus (k=139 voxels; [38 −32 −14]; r=0.76, p<0.0001). This cluster was not present in the regression using the left DC connectivity maps.
In addition, there were three regions whose connectivity to the DC negatively correlated with SL scores, such that subjects with greater connectivity between the DC and these regions tended to show less learning: the left postcentral gyrus (BA1; k=210 voxels; [−46 −16 28]; r=−0.78, p<0.0001), the right precentral gyrus (BA8; k=108 voxels; [46 8 44]; r=−0.70, p<0.0001), and the right medial superior frontal gyrus (BA8; k=118 voxels; [12 28 46]; r=−0.71, p<0.0001; Figure 5).
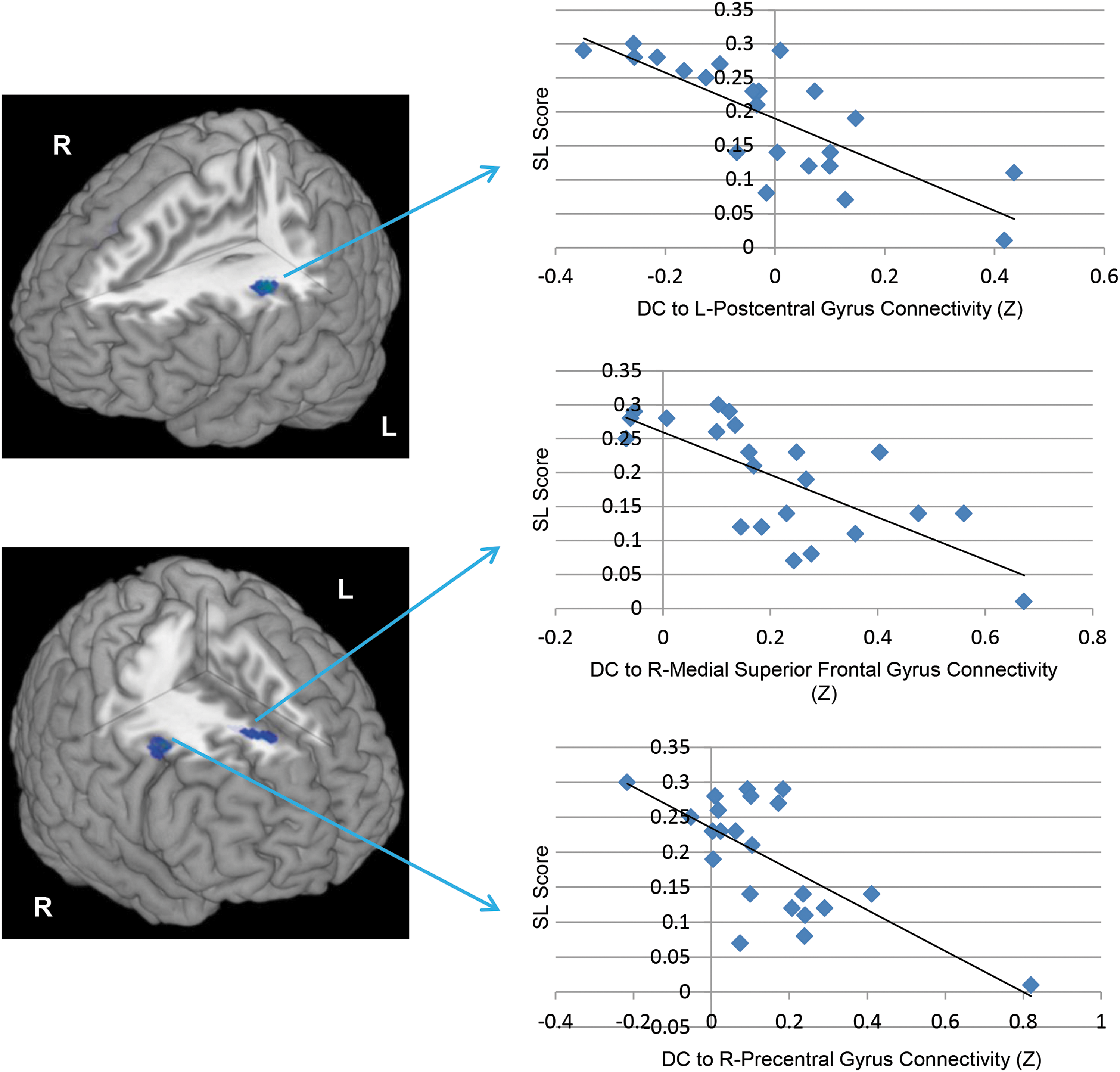
Negative correlations between resting functional connectivity of the DC seed and SL scores. As depicted in the left column of the figure, negative correlations between SL scores and DC resting connectivity were observed in clusters in the left postcentral gyrus (peak Z=4.38), right medial superior frontal gyrus (peak Z=3.81), and right precentral gyrus (peak Z=4.10). On the right are scatter plots depicting the z-transformed correlation coefficients between the DC and these clusters (x-axis) and SL scores (y-axis).
Correlations between DC connectivity and response speed
Average response speed correlated with connectivity of the DC to a cluster located in the right cuneus (BA 18; k=125 voxels, [16 −88 12]; r=−0.66, p=0.001) and left medial superior frontal gyrus (BA 10/32; k=140 voxels, [−14 44 22]; r=−0.67, p=0.001), such that subjects with stronger connectivity between the DC and these regions tended to have faster average response times. No regions correlated negatively with response speed. Further, the correlation between DC-MTL connectivity and average response speed (r=−0.15, p=0.50) was reliably smaller than that observed with SL scores (z=3.4, p=0.0007). In sum, DC-MTL connectivity predicted SL but not overall speed in the task.
Discussion
This study investigated whether DC connectivity at rest is related to individual differences in the implicit learning of sequential, probabilistic regularities in a sample of healthy adults. Supporting our hypothesis, subjects with greater positive rsFC between the DC and a cluster in the right MTL had superior learning. Importantly, individual differences in overall task performance cannot account for this relationship, as SL scores were not correlated with subjects' overall speed or accuracy, and DC-MTL connectivity was not related to subjects' overall response speed in the task. Faster overall response speed related, instead, to stronger connectivity of the DC with cuneus and medial frontal gyrus, regions associated with visual processing (Vanni et al., 2001) and response selection under uncertainty (Critchley et al., 2001), respectively. Thus, the strength of a DC-MTL network in the resting brain predicts how well one learns sequential probabilistic regularities, but does not predict overall task speed.
To our knowledge, this is the first study which shows that IPSL performance is correlated with connectivity of a task-relevant region (the DC) at rest. Since all scans were collected either before learning or well outside the timeframe when consolidation might be expected to occur, our finding suggests that rsFC is an intrinsic factor predictive of implicit SL aptitude. The relationships we observed between SL performance and rsFC of the DC with the MTL and frontal motor regions may, therefore, reflect the integrity and functional segregation of a task-relevant neural network.
Our results are consistent with those of previous studies using task-dependent fMRI, which find that implicit probabilistic types of learning depend on an interaction between the caudate and MTL. Specifically, MTL contributions to learning may predominate early in training, while the caudate becomes particularly relevant to performance later (Albouy et al., 2008; Poldrack et al., 2001; Rieckmann et al., 2010; Schendan et al., 2003; Simon et al., 2012). Further, implicit motor sequencing tasks show that greater SL is associated with a pattern of increasing striatal and decreasing MTL BOLD signal with practice, whereas individuals who do not show this pattern demonstrate less learning (Albouy et al., 2008; Rieckmann et al., 2010). These studies support the idea that a precisely coordinated interaction (i.e., a shift from hippocampus-dominant to caudate-dominant processing) between these regions is related to learning ability. As with these earlier task-dependent fMRI studies, our results also support the importance of the interaction between the DC and MTL by showing that the magnitude of IPSL is related to the strength of their connectivity at rest. A stronger positive intrinsic functional association between these regions at rest may enable more efficient interregional activation/inhibition during the course of learning (i.e., promote “readiness to learn”), thereby facilitating the observed interaction between the DC and MTL during SL. Future studies could examine this interpretation by combining task dependence with rsFC.
An important caveat is that it is still unclear exactly how the caudate and MTL interact during IPSL; the task-dependent activation of these regions during implicit tasks has been interpreted as both competitive (e.g., Poldrack and Packard, 2003) and cooperative (e.g., Schendan et al., 2003). Therefore, the positive relationship we report between DC-MTL rsFC and ISPL does not allow us to make any conclusions about whether the MTL is necessary for IPSL, as it is plausible that a more positive resting connection between the DC and MTL reflects the potential for more efficient suppression of the MTL during IPSL tasks. Interpretation of the MTL's role in IPSL is further complicated by the fact that there is some behavioral evidence of preserved SL in patients with MTL damage (Nissen and Bullemer, 1987; Nissen et al., 1989; Reber and Squire, 1994), suggesting that this region may not be necessary for IPSL. However, such clinical groups sometimes show less learning than healthy controls when the to-be-learned sequences are more complex (the sort studied here) and/or when training is extended (Curran, 1997; Nemeth et al., 2013; Vandenberghe et al., 2006), implying that MTL involvement may be necessary under certain conditions. Despite the mixed evidence from IPSL studies in clinical groups, the bulk of the (albeit correlational) evidence in healthy samples, such as the one tested here, have shown that both caudate and MTL are activated during IPSL, although frequently on different timescales. This could, therefore, indicate that these regions have distinct contributions to this type of learning (Gheysen et al., 2011).
It is notable that rsFC between DC and MTL predicts IPSL performance, even though there are a few direct anatomical connections between these regions. This is not surprising in light of evidence indicating that a direct anatomical connection is not necessary for functional connectivity in the resting state (Honey et al., 2009). For SL, our earlier work using DTI tractography suggests that this DC-MTL functional relationship may be mediated via connections with the dorsolateral prefrontal cortex (DLPFC) (Bennett et al., 2011). Early in training, SL scores were positively correlated with the microstructural integrity of the tract between the right MTL and DLPFC, while later in training, learning scores positively correlated with the integrity of left caudate to DLPFC tract. Moreover, individual differences in the integrity of the left caudate-DLPFC tract mediated the age differences in late-training learning between younger and older adults, such that accounting for that variability eliminated age differences in learning. Thus, it is possible that the observed connectivity between caudate and MTL was mediated by the DLPFC, and further, that variability in caudate-DLPFC and MTL-DLPFC anatomical connections could underlie learning differences. Future studies could combine fMRI functional connectivity and DTI to examine this possibility.
Although we did not predict any negative relationships, connectivity between DC and several motor planning regions was negatively correlated with SL. Higher SL scores were predicted by less positive, more negative connectivity between the DC and clusters in the left postcentral gyrus (BA1), right precentral gyrus (BA8), and right medial superior frontal gyrus (BA8). These negative correlations are interesting in light of anatomical (e.g., Lehéricy et al., 2004) and functional neuroimaging evidence (e.g., Lewis et al., 2003) in humans, suggesting that distinct striatal subregions underlie motor and cognitive processes. Specifically, the caudate may be primarily involved in networks supporting cognitive functions, receiving input from and projecting to the DLPFC, while the putamen may be primarily involved in motor functions, receiving input from and projecting to frontal motor areas [for reviews see (Alexander and DeLong, 1986; Seger, 2008)]. Thus, the superior learning we observed in subjects with less resting-state connectivity between the DC and these regions might indicate that SL is maximized when the supporting motor and cognitive circuits remain weakly associated at rest. Importantly, the negative correlations we detected do not imply that frontal motor regions are unimportant for task performance, but rather that they might be involved in distinct striato-cortical loops from those supporting sequence-specific learning. Such functional segregation of motor and cognitive networks at rest might facilitate more efficient adaptation to the limited motor demands of the TLT and a greater commitment of DC and MTL activity to sequence-specific learning. Another possibility is that these negative correlations occur because the TLT does not involve motor sequencing, a key difference between this task and more traditional SL tasks, such as the serial reaction time task. Replication using other tasks would, therefore, be informative.
Alternative interpretations
One important alternative interpretation of the present results relates to the implicit nature of the TLT and whether it, or any implicit task for that matter, is truly “process pure.” That is, there could be occasional influences of explicit memory. Several published studies have used sensitive recognition tasks to show that learning in the TLT is implicit (Howard et al., 2008; Simon et al., 2011a, 2011b, 2012), but our only measure of awareness in the present study came from a postexperiment interview (Simon et al., 2011a). Although no one accurately described the regularities from the TLT or reported using explicit strategies, this type of awareness test is arguably less sensitive than others used in prior studies. Therefore, one alternative interpretation for the present results is that subjects who displayed more effective cooperative interaction between explicit and implicit memory systems revealed enhanced learning. We cannot rule out this possibility here. However, if MTL activation during IPSL reflects influences from explicit memory processes, then MTL activation would be expected to increase with training, as people are most likely to become aware at later stages of the task. The fMRI studies of IPSL cited here, however, show that MTL activation decreases over the course of training. It is, therefore, unlikely that the implicit-explicit distinction can entirely account for MTL activation during IPSL.
Limitations and future directions
The present results should be interpreted in light of some limitations. First, the length of our resting scan was relatively short; longer scans may be more sensitive to individual differences (Anderson et al., 2011; Birn et al., 2013). However, reliable estimates of functional connectivity can be obtained in as little as 4 min (Van-Dijk et al., 2010; p. 15), suggesting that our scan length is sufficient. Second, our sample size is relatively small, although comparable (or larger) than that of other published studies using similar tasks (e.g., Bennett et al., 2011; Rieckmann et al., 2010; Schendan et al., 2003; Simon et al., 2012). Since the chances of Type I error increase in small samples, however, replication with a larger sample is desirable, and future studies focusing on individual differences should consider acquiring longer scans.
Third, our method of recruitment did not permit a tightly controlled interval between the scan and behavioral testing. Nevertheless, there was no evidence that the time interval separating the scan from behavioral testing influenced learning, and our pattern of results remained when the two subjects scanned after behavioral testing were removed from the analyses. Thus, our results do not reflect consolidation processes resulting from the learning task itself. More studies of rsFC are needed, however, in order to better understand the stability (and experience-induced plasticity) of resting networks over time.
Conclusion
Our results are the first to show a relationship between IPSL and rsFC of the DC and task-relevant regions and suggest that the predictive value of resting networks is not exclusive to explicit, goal-directed cognition. As predicted, subjects who had greater positive connectivity between the DC and MTL at rest revealed greater SL, adding to existing fMRI and DTI evidence that the MTL supports IPSL in conjunction with the caudate. In addition, our unexpected finding that subjects with more negative connectivity between the DC and frontal motor regions at rest revealed less learning suggests that there may be functional segregation within neural networks not only during tasks, but also at rest. Future research should extend these findings by examining additional characteristics associated with resting-state connectivity (e.g., age, genotype, and lifestyle) and by combining resting-state fMRI with task-dependent fMRI and DTI techniques. Such research would help bolster evidence that resting-state connectivity is an intrinsic factor contributing to individual differences in a variety of cognitive processes, now including IPSL.
Footnotes
Acknowledgments
The authors thank Stephanie Bean, Halley Feldman, and Megan Norr for assistance with subject recruitment and data collection. This work was supported by the National Institutes of Health (grant numbers MH86709 to C.J.V., MH088066 to E.M.G., and R01AG036863 to D.V.H and J.H.H., Jr.).
Author Disclosure Statement
None of the authors report any competing financial interests.