
Abstract
Functional imaging studies have indicated hemispheric asymmetry of activation in bilateral supplementary motor area (SMA) during unimanual motor tasks. However, the hemispherically special roles of bilateral SMAs on primary motor cortex (M1) in the effective connectivity networks (ECN) during lateralized tasks remain unclear. Aiming to study the differential contribution of bilateral SMAs during the motor execution and motor imagery tasks, and the hemispherically asymmetric patterns of ECN among regions involved, the present study used dynamic causal modeling to analyze the functional magnetic resonance imaging data of the unimanual motor execution/imagery tasks in 12 right-handed subjects. Our results demonstrated that distributions of network parameters underlying motor execution and motor imagery were significantly different. The variation was mainly induced by task condition modulations of intrinsic coupling. Particularly, regardless of the performing hand, the task input modulations of intrinsic coupling from the contralateral SMA to contralateral M1 were positive during motor execution, while varied to be negative during motor imagery. The results suggested that the inhibitive modulation suppressed the overt movement during motor imagery. In addition, the left SMA also helped accomplishing left hand tasks through task input modulation of left SMA→right SMA connection, implying that hemispheric recruitment occurred when performing nondominant hand tasks. The results specified differential and altered contributions of bilateral SMAs to the ECN during unimanual motor execution and motor imagery, and highlighted the contributions induced by the task input of motor execution/imagery.
Introduction
Supplementary motor area (SMA) has been found to play a dominant role in motor planning, preparation, and control (Babiloni et al., 2003; Brinkman, 1981; Cunnington et al., 2002; Jenkins et al., 2000; Nair et al., 2003; Serrien et al., 2002; Tanji and Shima, 1994), while the primary motor cortex (M1) directly participates in movement execution (Haaland and Harrington, 1996; Nair et al., 2003; Porro et al., 1996). Thereby, these two areas have been consistently found to be significantly activated during both motor execution and motor imagery tasks (Gao et al., 2011; Gerardin et al., 2000; Guillot et al., 2008; Hanakawa et al., 2003; Jeannerod, 1994; Michelon et al., 2006; Solodkin et al., 2004; Szameitat et al., 2007). These functional results were largely in agreement with the anatomical findings in monkeys and humans, showing that the SMA was densely and reciprocally connected with the hand area of M1 (Arai et al., 2012; Johansen-Berg et al., 2004; Luppino et al., 1993; Muakkassa and Strick, 1979; Rouiller et al., 1994). In addition, the SMA has shown hemispherically asymmetric patterns of activation during both motor execution and motor imagery tasks (Babiloni et al., 2003; Haaland and Harrington, 1996; Jancke et al., 2000; Rogers et al., 2004), demonstrating a left dominant activation of SMA in right-handed subjects. The results implied the presence of asymmetry in the left/right SMA and M1 connectivity networks. Thereby, it is very important to clarify the information transfer and integration among the left SMA, right SMA, and M1 during motor-related tasks, to specify the roles of ipsilateral and contralateral SMA during the tasks.
Some studies have been done on the connectivity of SMA and M1 during motor execution in terms of effective connectivity, which refers explicitly to the influence that one neural system exerts over another (Friston, 2011). The influence of SMA on M1 has been consistently confirmed by many effective connectivity measurements such as Granger causality analysis (Chen et al., 2009; Gao et al., 2008, 2011; Sato et al., 2006), structural equation modeling (Solodkin et al., 2004), and dynamic causal modeling (DCM) (Grefkes et al., 2008). These results demonstrated a predominantly facilitatory function of the SMA in the top-down processes during motor execution (Arai et al., 2012; Gao et al., 2011; Grefkes et al., 2008), and implied that ipsilateral and contralateral SMAs played different roles on contralateral M1 (Gao et al., 2011).
As for motor imagery where there is no overt body movement, the connectivity network analyses have shown a similar SMA-M1 connectivity pattern (Gao et al., 2011; Solodkin et al., 2004). Importantly, using DCM, Kasess et al. (2008) showed that the SMA exerts suppressive influence over contralateral M1 during motor imagery, whereas Grefkes et al. (2008) demonstrated the facilitation of SMA on contralateral M1 during motor execution. This alteration of the influence that SMA exerts over contralateral M1 during motor execution and motor imagery conditions implied the different roles of SMA induced by different task conditions. However, the specifically different contributions of ipsilateral and contralateral SMAs to M1 during motor execution and motor imagery tasks, and the possible hemispherically asymmetric patterns of effective connectivity networks (ECN) among the left/right SMA and M1 during tasks remain unclear. Furthermore, motor execution and motor imagery task conditions may possibly modulate the intrinsic coupling of the network in different ways and contribute to the mechanism of ECN associated with different task conditions, which also need to be clarified.
In summary, earlier studies have demonstrated the dominant role of SMA on M1 during both motor execution and motor imagery; however, the differential contributions of the left and right SMAs in motor execution and imagery networks remain to be evaluated. In the present study, we hypothesized that (1) ipsilateral and contralateral SMAs played different roles on contralateral M1 in terms of effective connectivity; and (2) these roles were modulated by different task conditions of motor execution and motor imagery. To test our hypotheses, functional magnetic resonance imaging (fMRI) experiment of unimanual motor execution and motor imagery tasks was performed, and DCM was used to analyze the ECN among left SMA, right SMA, and contralateral M1. DCM provides an effective method to construct mechanistic models of brain function, which not only takes into account the direct influence of experimental inputs on regional activity but also considers interactions among neuronal populations and how these interactions are modulated by experimental perturbations (Stephan et al., 2010). Specifically, to clarify the different roles of ipsilateral and contralateral SMA on contralateral M1, we chose regions of interest (ROI) in left SMA and right SMA separately; more importantly, the task stimuli modulation of the intrinsic coupling and the influence on the activated regions were highlighted using DCM in the present study, which distinguished the present study from our previous studies (Chen et al., 2009; Gao et al., 2011). In addition, the alteration of the networks under different conditions of motor execution and motor imagery and the dominant/nondominant hand issue were further discussed.
Materials and Methods
Subjects
Twelve healthy subjects (5 females, age range 20–24 years) with no history of psychiatric or neurological illness participated in the study. Handedness was evaluated by the Edinburgh Handedness Inventory (EHI). All the subjects were right handed (mean EHI score was 91.06, with a standard deviation of 6.01). The study was approved by the local ethics committee and all subjects provided informed written consent.
Experimental paradigm
The experiment was performed on a 3.0-T Siemens Trio scanner (State Key Laboratory of Brain and Cognitive Science, Institute of Biophysics, Chinese Academy of Sciences, Beijing, China) using a gradient-recalled echo planar imaging sequence with an eight-channel head coil (TR=2000 ms, TE=30 ms, FOV=24 cm, matrix=64×64, voxel size=3.75×3.75×5 mm3, 30 transverse slices without slice gap, and flip angle=90°). The experiment had two runs: the first one for right hand tasks, and the second one for left hand tasks. Each run included 10 trials, and each trial lasted 30 sec, including 4 sec for sequence informing, 10 sec for motor imagery, 6 sec for motor execution, and 10 sec for resting in order. During each trial, a visual stimulus of four sequentially presented pictures first appeared for 4 sec by a projector, indicating a random order of finger tapping. Subjects then started to imagine tapping their fingers continuously for 10 sec in the order informed by the visual stimulus. Another cue for the next 6 sec was presented on the screen informing the participants to perform the finger tapping exactly as they had imagined. All subjects have been trained for about 1 h to perform the behavior experiments before scanning. At the end of the scanning session, the subjects were debriefed and asked about their performance of the tasks. All subjects reported performing both the motor execution and motor imagery tasks successfully. High-resolution T1-weighted anatomical images were also acquired in axial orientation using a 3D spoiled gradient-recalled sequence (TR=8.5 ms, TE=3.4 ms, flip angle=12°, matrix size=512×512×156, and voxel size=0.47×0.47×1 mm3).
Data preprocessing and signal extraction
Data preprocessing was performed using statistical parametric mapping (SPM) software (SPM8,
The present study was restricted to bilateral SMAs and contralateral M1 for our purpose. The ROIs were defined as spheres within the activated regions in conjunction analysis in contralateral M1 corresponding to the performing hand, and the bilateral SMAs, with the center at the nearest local maximum in relation to the group analysis. The radius of the sphere was selected to ensure that the spheres located in bilateral SMAs were not intersected, which was 4 mm in the present study. The mean time course of each ROI for each subject was then calculated and linearly detrended. To extract the signals for motor execution and motor imagery conditions, respectively, the mean time course was first adjusted for realignment parameters for head movements, and then deconvoluted by the classic hemodynamic response function in SPM. The maximum of the delay correlation coefficient between the deconvoluted time course and the experiment pattern was then calculated to determine the time of the hemodynamic delay, which was 6 sec. A boxcar vector was defined for each condition of interest, with the onset lagging 6 sec after the onset of the motor execution or motor imagery condition accounting for the hemodynamic delay. The boxcar vectors were considered as task inputs in DCM. Finally, these boxcar vectors were multiplied with the mean time series to extract the signals for each condition separately (Gao et al., 2011).
Dynamical causal modeling analysis
In the current study, the DCM method as implemented in SPM8 for effective connectivity analysis was performed among the selected ROIs during motor execution and motor imagery to evaluate our models. As a powerful approach to inferring effective connectivity from fMRI data, DCM has increasing applications. The validity of DCM has been evaluated by numerous studies (Bitan et al., 2005; Friston et al., 2003; Kasess et al., 2008; Penny et al., 2004b; Smith et al., 2006; Stephan et al., 2005; Szameitat et al., 2007). DCM uses a bilinear model where the changes in neuronal states over time can be evaluated by the following:
where Z represents the neuronal states and u is the input. A represents the intrinsic coupling among the states, B includes the parameters that allow the inputs to modulate the intrinsic coupling, and C is the influence of extrinsic input on the states (Friston et al., 2003). The parameters A, B, and C need to be estimated to infer the connectivity between the neural sources that give rise to these regional measurements (Friston et al., 2003; Stephan et al., 2007).
As an effective connectivity method, DCM is hypothesis driven and requires a priori model structure definition, for which the connectivity parameters are estimated (Kasess et al., 2008). Totally, five model classes were defined according to previous studies. The variations of the model classes are shown in Figure 1. Common to all the model classes, the task input (motor execution or motor imagery) was projected to all the three activated regions. Further support for this hypothesis came from the results of Kasess et al. (2008) demonstrating that, SMA and M1 as nodes of an interactive closed-loop control circuit subserving motor task execution should both be modulated by task stimuli (Brigadoi et al., 2012; Kasess et al., 2008). Since the activated regions were detected using a general linear model in which task input was the main regressor, the influences of the inputs C were assumed to be >0. The bidirectional connections between bilateral SMAs were based on studies demonstrating that the bilateral SMAs were reciprocally connected in the macaque brain (Grefkes et al., 2008; McGuire et al., 1991; Rouiller et al., 1994) and were intrinsically connected in the DCM network in the human brain during hand movements (Grefkes et al., 2008). Moreover, we assumed a forward influence from contralateral SMA to contralateral M1 during unilateral tasks since the connection was also identified in both the macaque brain (Rouiller et al., 1994) and human brain (Grefkes et al., 2008). The remaining intrinsic couplings among regions were varied for the different classes. Since modulation of an intrinsic connection can only occur if the connection exists, the modulating influences from the inputs on the coupling (i.e., B parameters) were varied accordingly. In model class 1, there were three possible modulating influences. Varying the three influences resulted in 23=8 possible models. Similarly, in model class 2, there were 24=16 possible models. Varying all five model classes in this way led to a total of 23+24+24+25+26=136 models to be estimated.
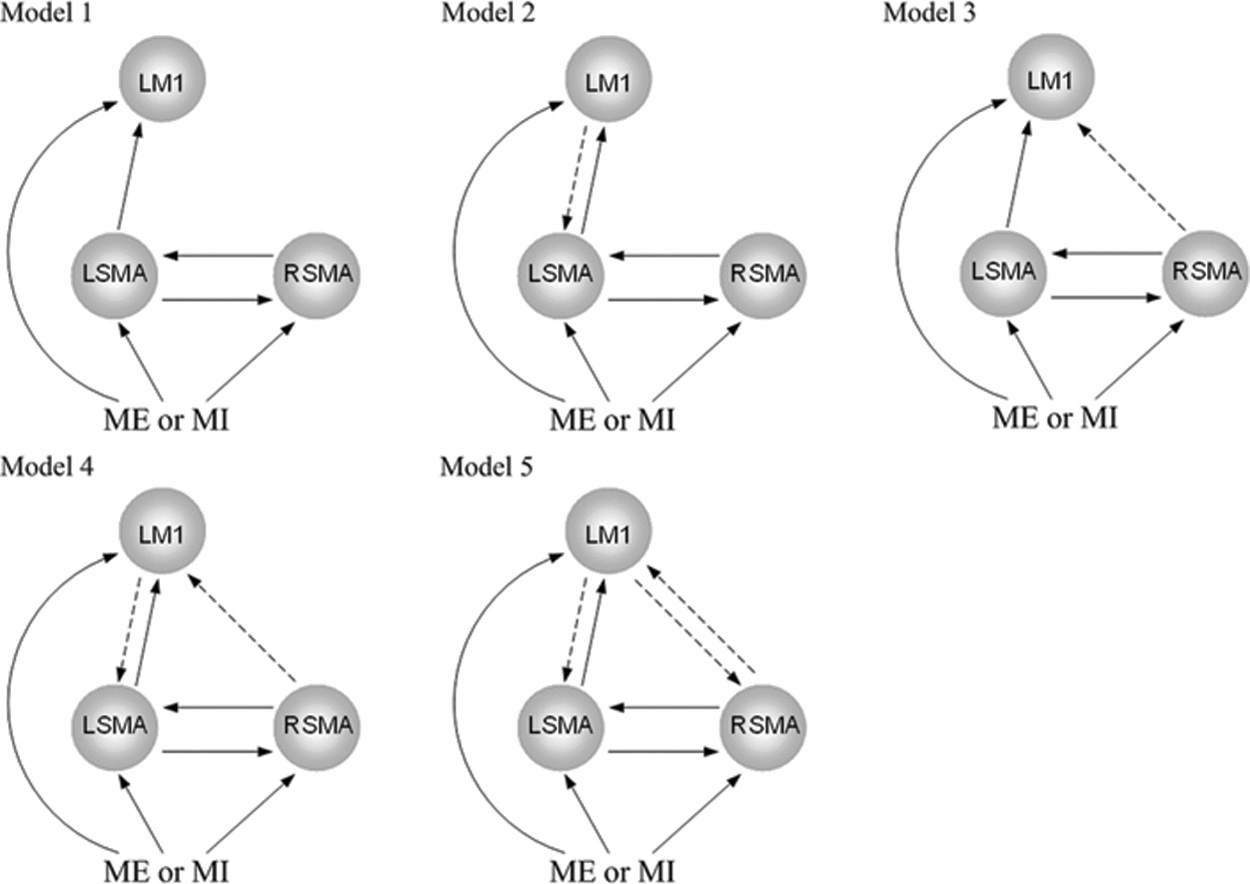
The five basic model classes. Connections that were constant throughout all models are shown as solid arrows. The modulating influences from the inputs on the coupling were varied accordingly. L, left; M1, primary motor cortex; ME, motor execution; MI, motor imagery; R, right; SMA, supplementary motor area.
The models were compared using the Bayesian model selection to identify the connectivity model showing the highest (posterior) evidence (Grefkes et al., 2008; Penny et al., 2004a). The signals were first averaged across subjects to yield the data for DCM to obtain the best model pattern (Kasess et al., 2008; Stephan et al., 2010); then, to extend the model to the general population from which the subjects were drawn, we further estimated the model parameters for each subject using the optimal model pattern and then performed a second-level inference using a one-sample t-test (Grefkes et al., 2008; Stephan et al., 2007, 2010). We performed the tests separately for each parameter of A, B, and C of the optimal model. Connections were considered to exist if they passed a statistically significant threshold of p<0.05 (Bonferroni corrected for multiple comparisons). The group DCM results were assessed as averages over the subjects' DCM parameters.
Results
Brain activation during tasks
The information of peak voxels and statistical t-value for conjunction analysis of motor execution and motor imagery are summarized in Table 1. Only selected regions in bilateral SMAs and M1 are reported for our purpose.
Local Maxima of Significantly Activated Regions During Motor Execution and Motor Imagery
Only selected regions in bilateral SMAs and M1 are reported for our purpose.
BA, Brodmann's area; L, left; M1, primary motor cortex; R, right; SMA, supplementary motor area.
DCM of right hand tasks
Figure 2 depicts the best model patterns suggested by Bayesian factors for motor execution and motor imagery, respectively. The bidirectional connectivity between bilateral SMAs and between left SMA and left M1 existed in both models; the task stimuli modulated the coupling from left SMA to left M1, from left SMA to right SMA, and from right SMA to left SMA. Figure 2a demonstrates the significant parameters of the effective network among the three regions during motor execution. The left SMA had a very strong positive effective connectivity on left M1, and left M1 had a positive feedback to left SMA; the bidirectional connections between the left SMA and right SMA were also positive. Task input had a strong negative modulation of the coupling from left SMA to right SMA (−0.42, p<0.01, Bonferroni corrected). The indirect suppressive influence of the task input on the right SMA resulted in the inhibition of left hand movement during right hand tasks.
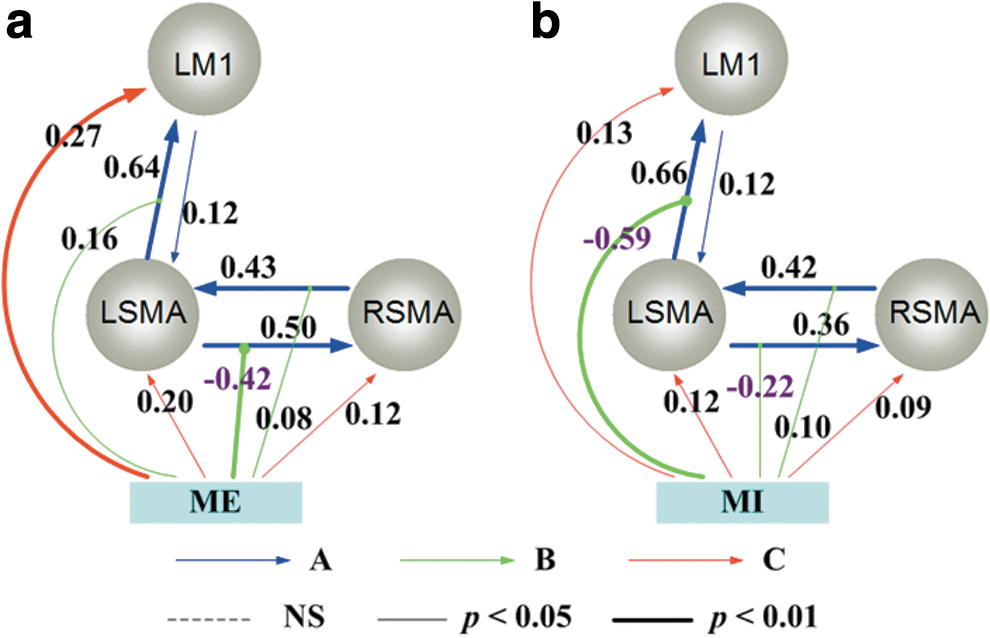
The effective connectivity networks (ECN) during right hand tasks.
Similar to the motor execution network, all effective connections among active regions were positive during motor imagery (Fig. 2b). However, since there was no overt body movement, the positive modulation of the coupling from the left SMA to left M1 turned to be negative (−0.59, p<0.01, Bonferroni corrected). The result indicated the inhibitive function of the task input modulation during motor imagery.
DCM of left hand tasks
Figure 3 depicts the best models of left hand tasks during motor execution and motor imagery, respectively, which was almost mirror reversed to the networks during right hand tasks (Fig. 2). During motor execution (Fig. 3a), all the intrinsic couplings were positive (the negative connection from left SMA to right M1 was not statistically significant). The task input had positive influences on the three regions. The effective network during left hand motor imagery is shown in Figure 3b. Similar to left hand motor execution, positive connections existed among all activated regions. The task input had negative modulation of the coupling from the right SMA to right M1 (−0.49, p<0.01, Bonferroni corrected) during motor imagery.
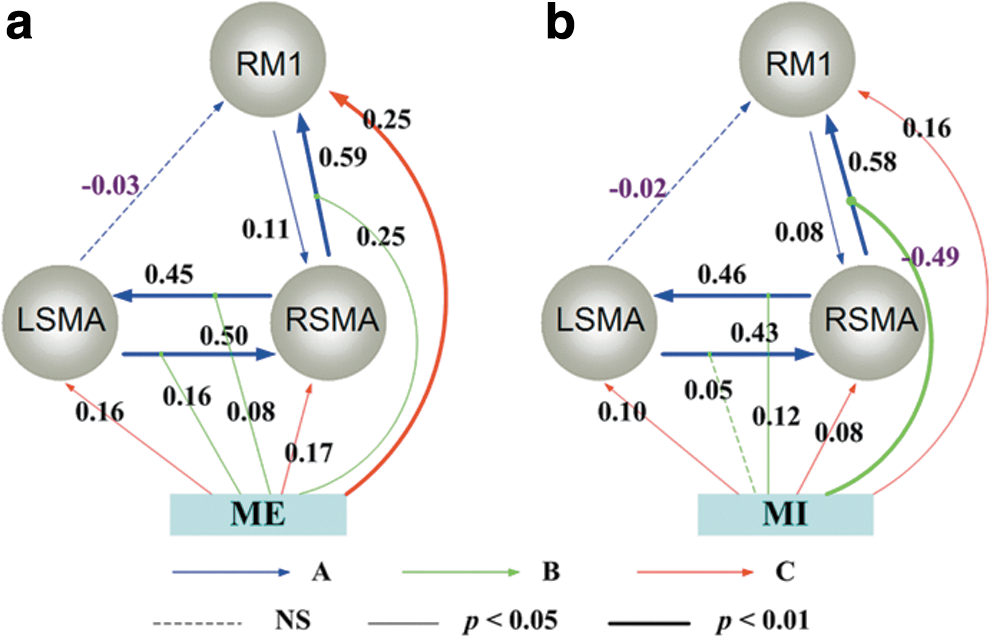
The ECN during left hand tasks.
Comparison of motor execution and motor imagery networks
To compare the dynamic causal networks during motor execution and motor imagery, the significant connectivity parameters of right hand motor execution, right hand motor imagery, left hand motor execution, and left hand motor imagery were entered in the repeated-measure analysis of variance. Since the activation in ipsilateral M1 was not significantly detected during the tasks by group analysis in our study, the parameters associated with M1 during right hand tasks and left hand tasks were compared mirror reversed. The result showed a significant difference of the four set of parameters (p<0.01, Bonferroni corrected). The following paired t-tests suggested a significant difference existed between motor execution and motor imagery conditions (p<0.01, Bonferroni corrected) rather than performing hand conditions. Furthermore, a Kolmogorov–Smirnov test was performed to compare the distributions of motor execution and motor imagery parameters. The results suggested that the two datasets were drawn from two different underlying distributions (p<0.01, Bonferroni corrected).
The paired t-test was further performed to compare each connection during motor execution and motor imagery, respectively, to detect where the difference came from. Table 2 depicted the connectivity parameters with significant differences during motor execution and motor imagery (p<0.01, Bonferroni corrected). None of the intrinsic connections showed significant differences between motor execution and motor imagery conditions nor did the influence of the inputs on regions. The results suggested that the significant difference between motor execution and motor imagery was mainly induced by the task input modulation of the intrinsic connections.
Significant Alterations of Task Stimuli Modulation/Influence During Motor Execution and Motor Imagery
ME, motor execution; MI, motor imagery.
Discussion
To improving our understanding of hemispheric asymmetry of movement, it is very important to understand the mechanism of inter- and intrahemispheric information integration, which strongly depends on task stimuli (Haaland and Harrington, 1996; Stephan et al., 2007). The challenge here is to test which candidate mechanism (model) of information transfer and integration represents the best balance between prediction accuracy and model complexity (Stephan et al., 2010). In the present study, DCM was used to model the ECN among bilateral SMAs and contralateral M1 during dominant and nondominant hand motor execution/imagery tasks in right-handed subjects. Our results specified different contributions of bilateral SMAs during unimanual motor execution/imagery tasks and demonstrated that the intrinsic couplings among bilateral SMAs and contralateral M1 were task independent. Moreover, the alteration of motor execution/imagery networks was mainly induced by the task inputs, emphasizing the input modulations of the intrinsic coupling.
We should note that the order of the task conditions was the motor imagery condition followed by the motor execution condition. The reason that the motor imagery task was followed by the motor execution task was to ensure that the participants concentrated on the motor imagery task and imagined the finger tapping correctly. In addition, the order ensured the motor imagery signal against contamination by the motor execution signal, for the motor imagery signal was weaker than the latter.
Methodological considerations
DCM is a powerful approach to explore the dynamic mechanisms in systems. The use of it is becoming increasingly widespread in many domains of cognitive neuroscience and neurophysiology (Stephan et al., 2010). However, there were some issues we should be aware of in its application. First, from a mathematical and computer science perspective, the number of possible DCM models was combinatorial explosion when increasing the number of regions and inputs (Lohmann et al., 2012; Stephan et al., 2010). Therefore, model space should be well defined by prior knowledge to reduce the number of the plausible alternatives. Second, we should note that the winning model is from the candidate set that we have defined and extremely dependent on the choice of the candidate set (Lohmann et al., 2012). Therefore, in practice, it is critical to ensure a clear definition and systematic exploration of model space (Stephan et al., 2010). To do this, it is helpful to make full use of previous empirical studies in humans or animals (Stephan et al., 2010).
Intrinsic coupling during motor execution and motor imagery
As Equation (1) showed, theoretically, the connectivity matrix A represented the first-order connectivity among the regions in the absence of input (Friston et al., 2003). This means that the intrinsic connectivity is always present during the experiment and, hence, represents the task-independent component of interregional coupling (Grefkes et al., 2008). Thereby, in the study of Grefkes et al. (2008), they separated the parameter estimation process into two steps by first estimating matrix A in the absence of task stimuli and then analyzing the task modulation of the intrinsic coupling. In our present study, version DCM8 was used in which inputs were not mean centered. Therefore, the connectivity matrix A represented the baseline connectivity in the absence of input. In addition, matrix A was not estimated separately in DCM8. Our statistical analysis demonstrated that the connectivity parameters (A, B, and C) of motor execution and motor imagery were from different distributions, although (p<0.01, Bonferroni corrected) the intrinsic couplings did not show differences between motor execution and motor imagery. Our results verified that intrinsic connectivity was not influenced or driven by task-related activity, and expanded the findings to motor imagery, implying that motor execution and motor imagery shared the same intrinsic coupling patterns among bilateral SMAs and contralateral M1.
Regardless of the performing hand, all the intrinsic couplings were significantly positive in both motor execution and motor imagery networks (p<0.05, Bonferroni corrected). The effective circuit between bilateral SMAs existed in all four networks. The anatomic studies had reported that SMAs were reciprocally connected and each projected to both contralateral and ipsilateral primary motor areas (McGuire et al., 1991; Muakkassa and Strick, 1979; Pandya and Vignolo, 1971; Rouiller et al., 1994), which anatomically supported our results on the functional interactions of bilateral SMAs. In functional studies, it had been convinced that bilateral SMAs were both activated during unimanual movement (Deecke, 1987), and the interaction of bilateral SMAs played a crucial role in both unimanual and bimanual coordination (Grefkes et al., 2008; Seitz et al., 2004; Stancak et al., 2003). These results implied the information transfer between bilateral SMAs during both unimanual and bimanual movements. Our result was well compatible with these earlier findings. Furthermore, our results demonstrated that this interconnection also existed during left/right hand motor imagery tasks, and this intrinsic circuit permanently existed regardless of the task conditions.
Furthermore, in the unimanual tasks, contralateral SMA had a very strong effective connectivity to contralateral M1 regardless of the performing hand or motor execution/imagery tasks. The feedback from contralateral M1 to contralateral SMA also existed throughout all four networks. Neuroimaging experiments with human subjects demonstrated the involvement of SMA in many unimanual tasks, consistently showing the special role of SMA for motor planning, preparation, and control (Cunnington et al., 2002; Deecke, 1987), in particular, for movement sequencing, internal pacing, and temporal organizing (Jenkins et al., 2000; Passingham, 1989). The forward and backward interactions among contralateral SMA and M1 found in our results suggested the function integration among the regions to accomplish the motor-related tasks. Moreover, the earlier study on single-cell recording data in monkeys showed that a large majority of SMA neurons exclusively respond to contralateral hand movements only (Grefkes et al., 2008; Kazennikov et al., 1999). Especially, our results specified this effective circuit in contralateral hemisphere, expended the results to the motor imagery condition, and showed similar mechanisms in motor execution and motor imagery. Our findings suggested that the contralateral SMA had a predominant influence on contralateral M1 during the unimanual tasks.
The task conditions influence/modulation
When the network parameters were compared during motor execution and motor imagery, our results indicated that parameters during motor execution and motor imagery were drawn from different underlying distributions, suggesting that different connectivity patterns were involved during motor execution and motor imagery. This variation was mainly induced by the task input modulation of the intrinsic connections.
As Table 2 showed, during both right and left hand motor imagery, the task input modulation of coupling from contralateral SMA to contralateral M1 changed from significantly positive to significantly negative comparing to motor execution. The results suggested that during motor imagery where there was no overt body movement, the task input had a suppressive role on contralateral M1 through a very strong negative modulation of the intrinsic coupling from contralateral SMA to contralateral M1 (−0.59 for right hand motor imagery and −0.49 for left hand motor imagery, p<0.01, Bonferroni corrected). This resulted in the suppressive role of contralateral SMA to contralateral M1. Some earlier studies had reported that the strong activity in M1 during motor execution was significantly suppressed during motor imagery (Lotze et al., 1999; Porro et al., 1996, 2000). Our results demonstrated that the decreased activity in M1 was induced by the suppressive function of task input through contralateral SMA. The result was concordant with the results of the Kasess et al. (2008) group, which demonstrated the suppressive influence of SMA on M1 during motor imagery by DCM. Moreover, our study specifically located this suppressive influence in contralateral SMA during motor imagery. Furthermore, this suppressive pattern during left hand motor imagery was totally mirror reversed to that during right hand motor imagery, suggesting similar inhibitory mechanisms in both dominant and nondominant hand motor imagery.
Dominant and nondominant hand tasks
From the point of view of network parameter distribution, our results demonstrated that the underlying distributions of parameters from right hand and left hand motor execution did not show significant differences. Neither did those from right hand and left hand motor imagery. Regardless of the performing hand, the facilitatory modulation of the coupling from contralateral SMA to contralateral M1 during motor execution consistently changed to be negative during motor imagery, indicating an inhibitory effect of contralateral SMA to contralateral M1 during motor imagery.
We further compared SMA roles during dominant and nondominant hand motor execution. During right hand tasks, task stimuli had a negative influence on the intrinsic coupling from left SMA to right SMA. This might appropriately help the right SMA to modulate the right M1 activity and thereby to suppress the left hand movement. However, during left hand tasks, the expected mirror reversed results were not found. In addition, the task input had a positive modulation of coupling from left SMA to right SMA in even in left hand tasks. The results implied that left SMA also played a facilitatory role in left hand tasks and expended the earlier results of the dominant role of left SMA during unimanual motor execution (Babiloni et al., 2003; Rogers et al., 2004) to motor imagery. This might due to the right-handed subjects in our study. In the right-handed subjects, left SMA plays a dominant role in unilateral movements of either hand over right SMA (Babiloni et al., 2003; Rogers et al., 2004). This hemispherically asymmetric modulation of connectivity has been predicted by the hemispheric recruitment theory of interhemispheric integration (Banich, 1998; Stephan et al., 2007). The theory stated that the interhemispheric connectivity could dynamically undergo adjustment depending on task demands to better meet task demands or learn a task (Banich, 1998). Specifically, if the neural resources in the hemisphere receiving a stimulus are insufficient for optimal processing, the benefits of the distributing processing load across both hemispheres should outweigh the costs of transcallosal information transfer (Stephan et al., 2007). In our results, left SMA and right SMA were tightly intrinsically connected, while left SMA was further recruited by left hand tasks. The results suggested that left SMA helped accomplishing the nondominant hand tasks, and verified that left SMA is dominant over right SMA in the ECN regardless of the performing hand or motor execution/imagery conditions in right-handed subjects.
Conclusions
We studied the ECN among overlapped activated regions in bilateral SMAs and contralateral M1 during motor execution and motor imagery using DCM. Motor execution and motor imagery did share the intrinsic coupling network, which was independent to the task stimuli. However, the network parameter distribution during motor execution and motor imagery tasks was significantly different suggesting that variation of the connectivity patterns between motor execution and motor imagery was mainly induced by the input modulation of the intrinsic connections. This variation was mainly presented in the input modulation of coupling from contralateral SMA to contralateral M1, where the significantly positive parameters during motor execution changed to be significantly negative during motor imagery regardless of the performing hand. This suppressed the overt movement during motor imagery. Moreover, the left SMA further helped accomplishing left hand tasks, implying that hemispheric recruitment occurred when performing nondominant hand tasks. Our results highlighted that the differential contribution of bilateral SMAs to the ECN during unimanual motor execution and motor imagery was induced by task conditions by means of task input modulation of intrinsic coupling between regions. Still some problems remained in the present study. First, the activation in ipsilateral M1 was not significant in group analysis; thereby, the differential roles of bilateral SMAs on ipsilateral M1 were not able to be studied. Second, the present results suggested that handedness may also be an issue that contributes to the ECN. The experiment on both left-handed and right-handed subjects may be designed and studied to identify the effective networks of motor execution and motor imagery associated with handedness in further studies.
Footnotes
Acknowledgments
This work was supported by grants from the Natural Science Foundation of China (91132721 and 61125304); the Fundamental Research Funds for the Central Universities of China (ZYGX2012J108); the Postdoctoral Science Foundation of China (20100481378); and the Special Postdoctoral Science Foundation of China (2012T50772). The funders had no role in the study design, data collection and analysis, decision to publish, or preparation of the article.
Author Disclosure Statement
The authors have no commercial associations that might create a conflict of interest in connection with this article.