
Abstract
Reading is an essential skill in modern society, but many people have deficits in the decoding and word recognition aspects of reading, a difficulty often referred to as dyslexia. The primary focus of neuroimaging studies to date in dyslexia has been on cortical regions; however, subcortical regions may also be important for explaining this disability. Here, we used diffusion tensor imaging to examine the association between thalamo-cortical connectivity and children's reading ability in 20 children with typically developed reading ability (age range 8–17/10–17 years old from two imaging centers) and 19 children with developmental dyslexia (DYS) (age range 9–17/9–16 years old). To measure thalamo-cortical connections, the structural images were segmented into cortical and subcortical anatomical regions that were used as target and seed regions in the probabilistic tractography analysis. Abnormal thalamic connectivity was found in the dyslexic group in the sensorimotor and lateral prefrontal cortices. These results suggest that the thalamus may play a key role in reading behavior by mediating the functions of task-specific cortical regions; such findings lay the foundation for future studies to investigate further neurobiological anomalies in the development of thalamo-cortical connectivity in DYS.
Introduction
Reading is an essential skill in modern society, but a large portion of population (5–17%) experiences difficulty in reading (Shaywitz, 1998), including deficits in the decoding and word recognition, or dyslexia. Dyslexia is a developmental reading disorder that is characterized by deficits in phonological processing which consequently impedes the development of adequate word recognition/decoding (Lyon, 1995). These deficits are specific, and are present despite adequate instruction and intelligence. While the behavioral characteristics of dyslexia are fairly well known, the neurobiological characteristics of this disorder are still under examination. With the advent of neuroimaging, especially functional magnetic resonance imaging (fMRI), over the last two decades in particular, studies have mapped the areas in the brain that are associated with good and poor reading. Converging findings reveal that proficient reading performance is associated with a coordinated left hemisphere network that involves temporo-parietal, occipitotemporal, inferior frontal (Eden et al., 2004; Pugh et al., 2000; Richlan et al., 2011; Shaywitz et al., 2004), and perisylvian regions, including premotor, pars opercularis, and triangularis (Broca's) areas (Heilman et al., 1996). Conversely, in those who demonstrate impaired performance (dyslexia), the homologous right hemisphere regions are recruited (Gilger et al., 2013; Pugh et al., 2000; Simos et al., 2000, 2007), with underactivation seen in particular in left fusiform gyrus (Richlan et al., 2011). There is also evidence for a cerebellar origin for developmental dyslexia (Buckner, 2013; Leiner et al., 1993; Pernet et al., 2009; Ramus et al., 2003; Stoodley and Stein, 2011).
In addition to differences in functional activation, the connectivity between these brain regions may also influence reading skill. Accurate and fluent reading requires a synthesis of information between the cortical processing regions via white matter tracts connecting these regions. For this reason, in order to understand more about dyslexia, structural neuroimaging studies have been employed to map white matter microstructure using diffusion tensor imaging (DTI). Consistent with findings from functional imaging studies, an initial study reported decreased fractional anisotropy (FA) of white matter associated with poor reading, and correlations between FA and reading scores in left temporo-parietal regions (Klingberg et al., 2000). These findings have also been replicated in children with a wide range of reading scores (Beaulieu et al., 2005; Deutsch et al., 2005; Niogi and McCandliss, 2006). Along with the left temporo-parietal regions, brain regions such as frontal regions (Carter et al., 2009; Lebel et al., 2013; Qiu et al., 2008; Rimrodt et al., 2010; Steinbrink et al., 2008), superior longitudinal fasciculus (Yeatman et al., 2011), and callosal tracts (Ben-Shachar et al., 2007; Dougherty et al., 2007; Frye et al., 2008) have also been reported to show white matter microstructural anomalies that are related to reading ability. In addition to the voxel-based morphometry analyses that focus on the localized white matter microstructural properties, diffusion tractography was employed to map the white matter fiber pathways which are important for reading, including arcuate fasciculus, corpus callosum, inferior longitudinal fasciculus, and inferior frontal occipital fasciculus (Catani et al., 2005; Odegard et al., 2009; Rauschecker et al., 2009; Saygin et al., 2013; Yeatman et al., 2012) (for a review, see Vandermosten et al., 2012; Wandell, 2011). Brain connectivity between distant regions, such as thalamus, angular gyrus, superior temporal cortex, and insula, has been shown to be related to reading ability in children (Davis et al., 2010). These findings suggest that the characteristics of white matter pathways between distant cortical regions are potentially an important aspect of the neurobiology of dyslexia.
While the primary focus of neuroimaging work to date in dyslexia has been on cortical regions, subcortical regions have been implicated. In particular, studies have reported individual variability in functional activities in the thalamus (Brunswick et al., 1999; Díaz et al., 2012; Fiebach et al., 2002; Hoeft et al., 2007; Maisog et al., 2008; Preston et al., 2010; Price and Friston, 1997; Price et al., 1994; Pugh et al., 2013; Turkeltaub et al., 2002; Wolf et al., 2010), as well as in structural properties (Brown et al., 2001; Galaburda and Eidelberg, 1982; Galaburda et al., 2006; Lebel et al., 2013) that are associated with differences in reading skills. The fact that there is thalamic involvement in dyslexia is not surprising, because the thalamus acts as an information processing way station for the brain, relaying signals contributing to the regulation of arousal (Portas et al., 1998) and cognition (Johnson and Ojemann, 2000; Karussis et al., 2000). It is, therefore, logical that the cortical regions implicated in dyslexia may be due to anomalous thalamo-cortical connectivity.
Nevertheless, despite the thalamus' central “relay station” role along with the abundant functional imaging studies showing thalamic anomalies in dyslexia, in vivo studies examining thalamo-cortical structural connectivity as related to reading ability are very limited, partially because it is technically difficult to capture thalamo-cortical connectivity, given the small size of the white matter tracts within the thalamus with regard to the typical MRI resolution in vivo (Metzger et al., 2013). However, studies have demonstrated the agreement between thalamic subdivisions identified by cytoarchitecture techniques and those identified by diffusion tractography-based connectivity measures (Behrens et al., 2003a; Johansen-Berg et al., 2005; Klein et al., 2010; Rushworth et al., 2006). In fact, the diffusion tractography-based thalamo-cortical connectivity analyses have recently been used to reveal structural alterations in neurological disorders, such as Alzheimer's disease (Zarei et al., 2010) and schizophrenia disease (Marenco et al., 2012), suggesting that an examination of thalamo-cortical connectivity in vivo is feasible.
Here, we compare the thalamo-cortical connectivity in children with typically developed (TD) reading ability and with dyslexia (DYS). Examining whether white matter connectivity anomalies are present from such a critical subcortical region of the brain, as well as where the anomalies may exist, may add knowledge to the subcortical-cortical relationships in developmental dyslexia. Given the lack of previous studies in dyslexia examining potential thalamic connectivity differences, our hypotheses were necessarily general; however, given the differences in functional activation in the thalamus between typically developing and dyslexic groups, we expected that we would find differences in thalamo-cortical connectivity between the TD and DYS groups. While specificity of these regions was difficult to pinpoint, in the one previous diffusion tractography-based connectivity study (Davis et al., 2010), we showed differences in thalamo-cortical connectivity between responders and nonresponders to reading intervention. Therefore, we expected thalamo-cortical differences between groups in the standard language-related regions near the perisylvian cortex, such as the occipito-temporal cortex (OTC) and the temporo-parietal cortex (TPC). The main focus of this study is the thalamus and cerebral cortex, and thus the cerebellum was not included in the analyses.
Materials and Methods
Participants
Participants of this study were a part of a larger study investigating the neurobiological basis of reading in children with a history of reading difficulty, Neurofibromatosis Type 1, and normal reading ability; it should be noted that due to the specificity of our hypotheses regarding reading ability, no participants with Neurofibromatosis Type 1 were selected to be in the current study. A total of 55 children participated in the current study. Before entering the study, parents of children were administered an informal screening measure over the phone to ensure that participants met the study's inclusion criteria: (1) native English speakers, (2) normal hearing and vision, (3) no history of major psychiatric illness, (4) no traumatic brain injury/epilepsy, and (5) no contraindication to MRI. Each parent gave written consent, while a separate written assent was obtained from each child at the start of the study, with procedures carried out in accordance with the university's Institutional Review Board. Eligible participants completed a battery of standardized tests to determine reading ability, that is, DYS or TD. The battery consisted of the following standardized measures of intellectual and academic achievement: Rapid Naming subtest from the Comprehensive Test of Phonological Processing (CTOPP) (Wagner et al., 1999); the Test of Silent Contextual Reading Fluency (TOSCRF-Form A) (Hammill et al., 2006); Phonological Decoding Efficiency (PDE) and Sight Word Efficiency (SWE) from the Test of Word Reading Efficiency (TOWRE) (Torgesen et al., 1997); Verbal Comprehension Index (VCI) and Perceptual Reasoning Index (PRI) subsets from Wechsler Intelligence Scale for Children, Fourth Edition (WISC-IV) (Wechsler, 2003); Spelling and Fundamental Literacy Index (FLI) subsets from the Word Identification and Spelling Test (WIST) (Wilson and Felton, 2004); and Word Attack (WA), Word Identification (Word ID), and Passage Comprehension subtests from the Woodcock Johnson-III (WJ-III) (Woodcock et al., 2001, 2003).
Participants met criteria for DYS if they had a standard score at or below the 25th percentile on the Basic Reading Standard Score (BR) on the WJ-III, which is the composite of the Word ID and WA measures. Participants met criteria to be TD by having a standard score at or above the 39th percentile on the BR. Five children were removed from the analysis due to motion artifacts in their imaging data; one was removed due to low VCI standard score (<75); three were removed due to low comprehension scores despite high word reading scores; and seven were removed, because their standard BR score fell in the range of 26th to 38th percentile. Of the eligible participants, 19 met criteria for DYS, and 20 met criteria for TD. All subjects are right handed. Behavioral profiles can be found in Table 1.
Demographic, Volumetric, and Neuropsychological Profile of Each Group (Mean±SEM)
BR, Basic Reading standard score (from WRMT-R/NU); CTOPP, Comprehensive Test of Phonological Processing; DYS, dyslexia; F, female; FLI, Fundamental Literacy Index; ICV, intracranial volume; LWID, Letter Word Identification standard score (from WRMT-R/NU); M, male; PComp, Passage Comprehension standard score (from WRMT-R/NU); PDE, Phonological Decoding Efficiency; PRI, Perceptual Reasoning Index; RaN, rapid naming; SEM, standard error in mean; Spell, spelling; SWE, Sight Word Efficiency; TD, typically developed; TOSCRF, Test of Silent Contextual Reading Fluency (Form A); TotWRE, Total Word Reading Efficiency; TOWRE, Test of Word Reading Efficiency; VCI, Verbal Comprehension Index; WA, Word Attack standard score (from WRMT-R/NU); WISC, Wechsler Intelligence Scale for Children; WIST, Word Identification and Spelling Test; WJ, Woodcock Johnson; ηp 2, partial eta squared as a measure of effect size.
Imaging data
MR data acquisition
Diffusion-weighted (DW) images and MPRAGE images were acquired on a Philips 3T MR scanner with an 8-channel head coil. The DW scan was acquired using the single shot echo planar imaging (EPI) sequence, TR/TE=6237/75 msec, flip angle=90°, SENSE factor=2.5 (AP), b=700 sec/mm2. The DTI data were acquired in 96×96 matrices with a field of view (FOV) of 212 (RL)×212 (AP) mm2, and zero filled to 256×256 matrices, yielding 0.83 mm isotropic in-plane resolution. Sixty axial slices were acquired with no gap, slice thickness=2.2 mm. Diffusion weighting was applied along 32 gradient directions that were evenly distributed on a unit sphere, and three non-DW images were acquired for averaging. Acquisition time was 3 min and 38 sec per scan. The MPRAGE images were acquired in coronal slices with no gap. Two hundred slices were collected in total. Each was acquired in a 256×200 matrix, with an FOV of 256 (FH)×200 (RL) mm2 and an isotropic voxel size of (1 mm)3. TR/TE=8.0/3.7 msec, flip angle=8°, SENSE factor=2 (AP), acquisition time was 7 min per scan.
It should be noted that data were acquired at both the Johns Hopkins School (JHU) of Medicine Kennedy Krieger Institute (KKI) and the Vanderbilt Institute for Imagining Sciences. To investigate any differences due to scanner location, all analyses were initially conducted with site (JHU-KKI and VUIIS) as a factor; it should be noted that unless indicated otherwise, results were not significant, and sites were removed from further analyses.
Image processing
The image analysis was performed in Freesurfer (
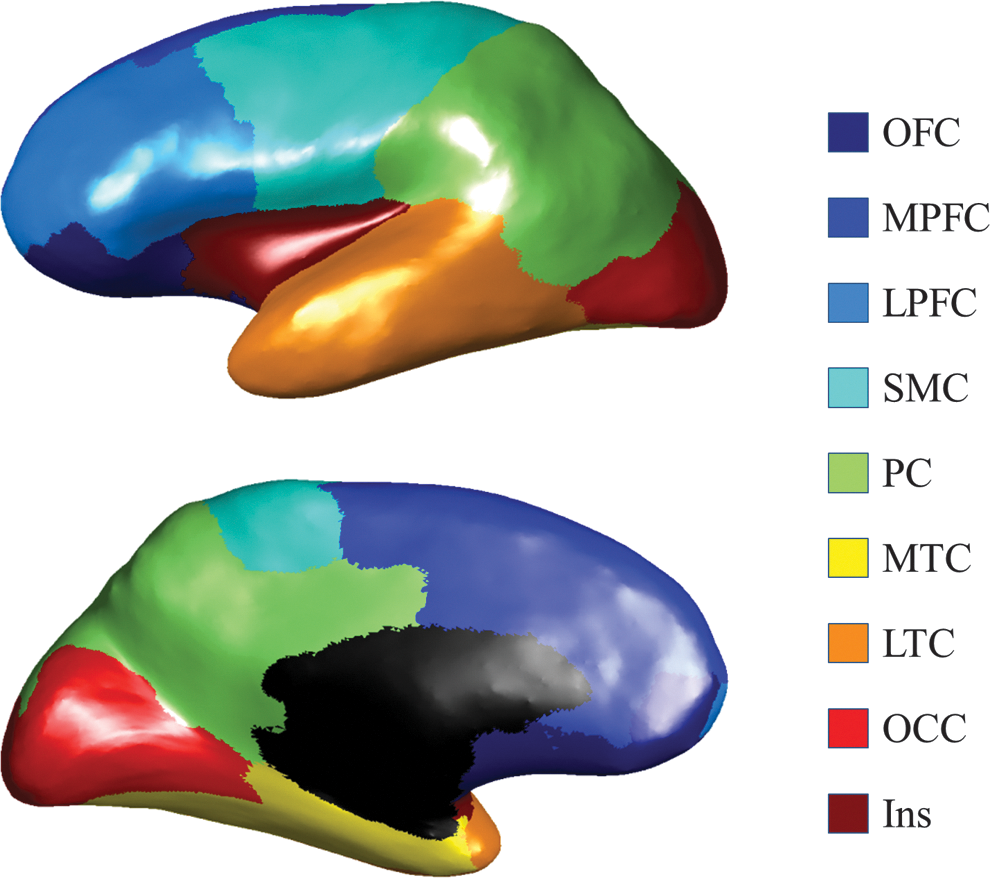
Cortical ROIs illustration. The subdivisions of the cortex used in the analysis are shown for the left hemisphere in a lateral (top) and medial (bottom) view. Bilateral ROIs were used in the analysis, whereas unilateral ROIs are shown here for visualization. The cortex was segmented into nine nonoverlapping regions with the original Freesurfer parcellations (Desikan et al., 2006): the LTC (transverse temporal cortex, superior temporal gyrus, banks of the superior temporal sulcus, inferior temporal gyrus, middle temporal gyrus, and temporal pole), the SMC (precentral gyrus, caudal middle frontal gyrus, postcentral gyrus, and paracentral lobule), the Ins, the MTC (entorhinal cortex, parahippocampal gyrus, and fusiform gyrus), the OCC (pericalcarine cortex, lingual gyrus, lateral occipital cortex, and cuneus cortex), the OFC (pars orbitalis, medial orbitofrontal cortex, and lateral orbitofrontal cortex), the LPFC (pars triangularis, frontal pole, rostral middle frontal gyrus, and pars opercularis), the PC (inferior parietal cortex, supramarginal gyrus, precuneus cortex, posterior cingulate cortex, isthmus cingulate, and superior parietal cortex), and the MPFC (caudal anterior cingulate, rostral anterior cingulate, and superior frontal gyrus). Ins, insula cortex; LPFC, lateral prefrontal cortex; LTC, lateral temporal cortex; MPFC, medial prefrontal cortex; MTC, medial temporal cortex; PC, parietal cortex; OCC, occipital cortex; OFC, orbitofrontal cortex; ROIs, regions of interest; SMC, sensorimotor cortex.
Probabilistic fiber tracking
The probabilistic fiber tracking was performed using the FDT tool in FSL (Behrens et al., 2003b). The probabilistic diffusion parameters were modeled with approximately two fiber compartments (Behrens et al., 2007), and probabilistic tractography was performed between the thalamus and the ipsi-lateral nine nonoverlapping cortical targets (heretofore referred to as ROIs, Fig. 1). From each thalamic voxel, 25,000 samples were drawn for probabilistic fiber tracking. Ventricles were avoided for fiber tracking. For each voxel in the thalamus, the number of samples reaching the nine cortical targets was counted, respectively, and the connection ratio to a specific cortical target was calculated as the ratio of the number of samples reaching this cortical target versus the total number of samples reaching any of the cortical targets.
Thalamo-cortical connectivity
For each cortical target, the thalamo-cortical connectivity was calculated as the average connection ratio in the ipsi-lateral thalamus. Thalamic voxels with zero connection densities were excluded from the connectivity calculation, in order to increase the specificity to thalamic regions from where a cortical connection exists.
Statistical analyses
The repeated-measure ANOVA (R-ANOVA) was conducted to compare the thalamo-cortical connectivity indices between the TD and DYS groups, where ROI (nine cortical targets), side (right and left) were within-subject factors, group (TD and DYS) was the between-subjects factor, and, because of group differences observed in verbal IQ, VCI was used as a covariate. Since the connection density was calculated as the ratio of number of fiber tracking streamlines reaching one ROI versus the total number reaching any of the ipsi-lateral ROIs, the relative size of the cortical target ROIs could potentially exert an impact on the calculated connectivity quantities. Therefore, as control analyses, the R-ANOVA analyses were also performed on the relative sizes normalized by the intracranial volume (%) (Buckner et al., 2004), as well as absolute sizes (in cm3), of the nine ROIs, in order to rule out the possibility that the group difference was driven by volumetric differences in target regions.
Results
Demographic data
Demographic variables were compared between groups using multivariate ANOVA, and a significant group difference was found (F[13,25]=8.69, p<0.001, ηp 2=0.819). As expected, children with DYS had significantly lower reading test scores than TD children (Table 1, all ps<0.001); they also had significantly lower Verbal IQs, as measured by the VCI on the WISC-IV (Wechsler, 2003). Results revealed no significant group differences in performance IQ, as measured by the PRI on the WISC-IV, or age, suggesting that the groups were similar with regard to both nonverbal IQ and age. Chi-square analyses also indicated no significant differences between the groups for gender distribution (χ 2=0.033, p=0.556). Due to the significant differences in VCI between groups, which is often seen in those with DYS (Kibby et al., 2009; Shaywitz et al., 2004), VCI was entered as a covariate for all subsequent analyses. No group difference was found in intracranial volume between groups.
Volumetric data
An R-ANOVA was performed to compare the absolute sizes of the 10 ROIs (9 cortical ROIs and the thalamus) between the TD and DYS groups, with ROI and side as within-subject factors, group as the between-subject factor, and age and VCI as covariates. A main effect of ROI was found (F[9,27]=11.541, p<0.001, ηp 2=0.248); no significant effect of side (left/right) or group (TD/DYS) or interactions was found. Follow-up analyses for the main effect of ROI indicated, as one would expect, that most of the 10 ROIs have significantly different absolute sizes from each other, with only two comparisons showing no significant differences: the medial prefrontal cortex (MPFC) versus the lateral prefrontal cortex (LPFC, p=0.878) and the insula versus the thalamus (p=1.000). All the rest of the pairwise analyses comparing absolute ROI sizes were ps<0.018 (corrected by Sidak). See Table 2 for the estimated marginal means of brain region volumes.
Repeated-Measure ANOVA Analysis of Absolute Sizes of Cortical Rois (cm3)
Note that the p-values reported in this and the next tables were corrected for multiple comparisons using Sidak.
The abbreviations in this and the next tables are consistent with Figure 1.
Thal, thalamus.
For relative ROI size, similarly, an R-ANOVA was performed to compare the relative ROI sizes between the TD and DYS groups, with ROI (nine cortical ROIs and the thalamus) and side (left and right) as within-subject factors, group as the between-subject factor, and age and VCI as covariate. Results indicated a main effect of ROI (F[9,27]=27.446, p<0.001, ηp 2=0.440) and age (F[1,35]=24.664, p<0.001, ηp 2=0.413); no significant effect of side (left/right) or group (TD/DYS) was found. A significant interaction of ROI by age was found (F[9,27]=9.312, p<0.001, ηp 2=0.210), and no other interactions was significant. Follow-up analyses for the main effects of ROI were as would be expected from the results of the absolute size analysis; all 10 ROIs showed significantly different relative sizes from each other, except for two comparisons: the MPFC versus the LPFC (p=0.796) and the insula versus the thalamus (p=1.000). All other pairwise analyses showed ps<0.023 (Table 3).
Group Comparison of Relative Volumetric Sizes of Target Regions of Interest a (%)
The relative size was calculated as the ratio of individual ROI size versus the intracranial volume.
ROI, regions of interest.
Thalamo-cortical connectivity data
An R-ANOVA was performed to compare the thalamo-cortical connectivity between groups, with the nine cortical ROIs as within-subject factors, group as the between-subject factor, and age and VCI as covariates. The R-ANOVA for thalamo-cortical connectivity values revealed a significant group×ROI interaction (F[8,28]=3.865, p=0.038, ηp 2=0.072); no significant effect of age or side was found; no other significant interaction was found. The overall between-subject effect of group (TD vs. DYS) was significant (F[1,35]=5.77, p<0.022, ηp 2=0.142). In the follow-up analyses, two regions were revealed to have significant thalamo-cortical connectivity differences between groups: the LPFC and the sensorimotor cortex (SMC). Both showed greater thalamic connectivity in the DYS group than in the TD group (see Table 4 for p-values). The R-ANOVA analysis showed that thalamic-SMC connectivity is significantly correlated with WJ-standard basic reading scores (Fig. 2), where a main effect of side was found (F[1,37]=7.409, p<0.01, ηp 2=0.167), with left thalamic-SMC connectivity higher than the right side (p<0.001). A significant interaction between side and behavioral score was found (F[1,37]=4.141, p<0.049, ηp 2=0.101), and both sides were negatively correlated with basic reading scores.
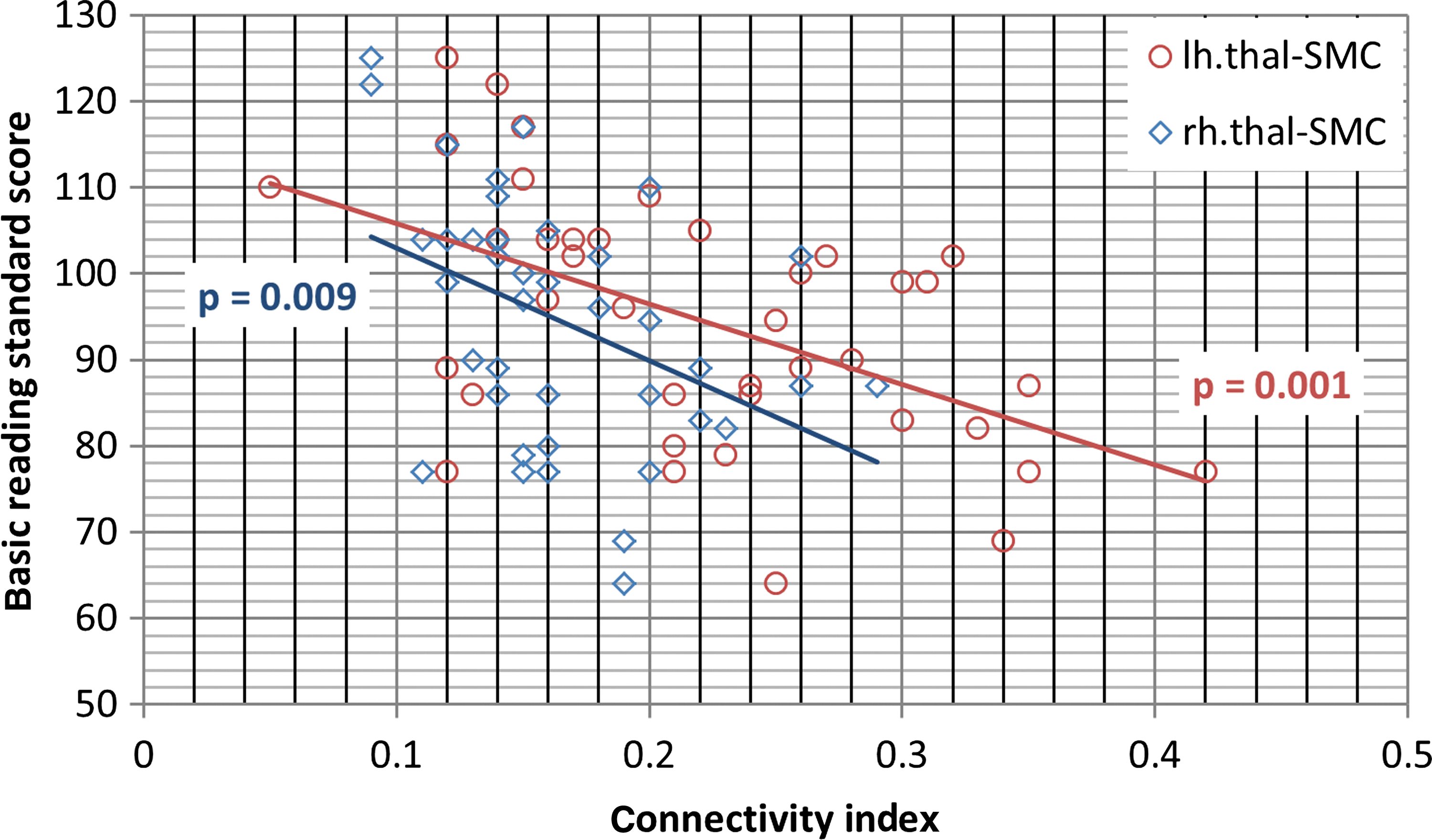
Illustration of correlation between thalamo-SMC connectivity and WJ-basic reading standard scores.
Group Comparison of Thalamo-Cortical Connectivity a (%)
Note that significant differences are reported for left and right hemispheres for informational purposes only; the overall group×ROI×side interaction was not significant (see text).
For each voxel in the thalamus, the number of samples reaching each target region was counted, and the connection ratio to a specific cortical region is calculated as the ratio of the number of samples reaching this cortical region versus the total number of samples reaching any of the cortical regions. For each ROI in the cortex, the thalamo-cortical connectivity was calculated as the average connection ratio in the ipsi-lateral thalamus.
The mean difference is significant at the 0.05 level, corrected for multiple comparisons using Sidak.
In Figure 3, an illustration of thalamic connection with SMC is shown. The probabilistic fiber tracking demonstrated that the streamlines originating from seed voxels in the thalamus followed the corona radiata, and then dispersed as they ascend and project to territories of the SMC. To locate these probabilistic fiber tracts within the thalamus, a tractography-based thalamic connectivity map was illustrated in Figure 4. The voxels in the thalamus were color coded, according to which cortical target they are most likely connected to (i.e., noted by connection ratio defined in Probabilistic Fiber Tracking section). In comparison to the multiarchitectonic division of the thalamus, it suggestes that the thalamic voxels primarily connected to the SMC were in the territories of the ventral lateral nucleus and the ventral posterior lateral nucleus.
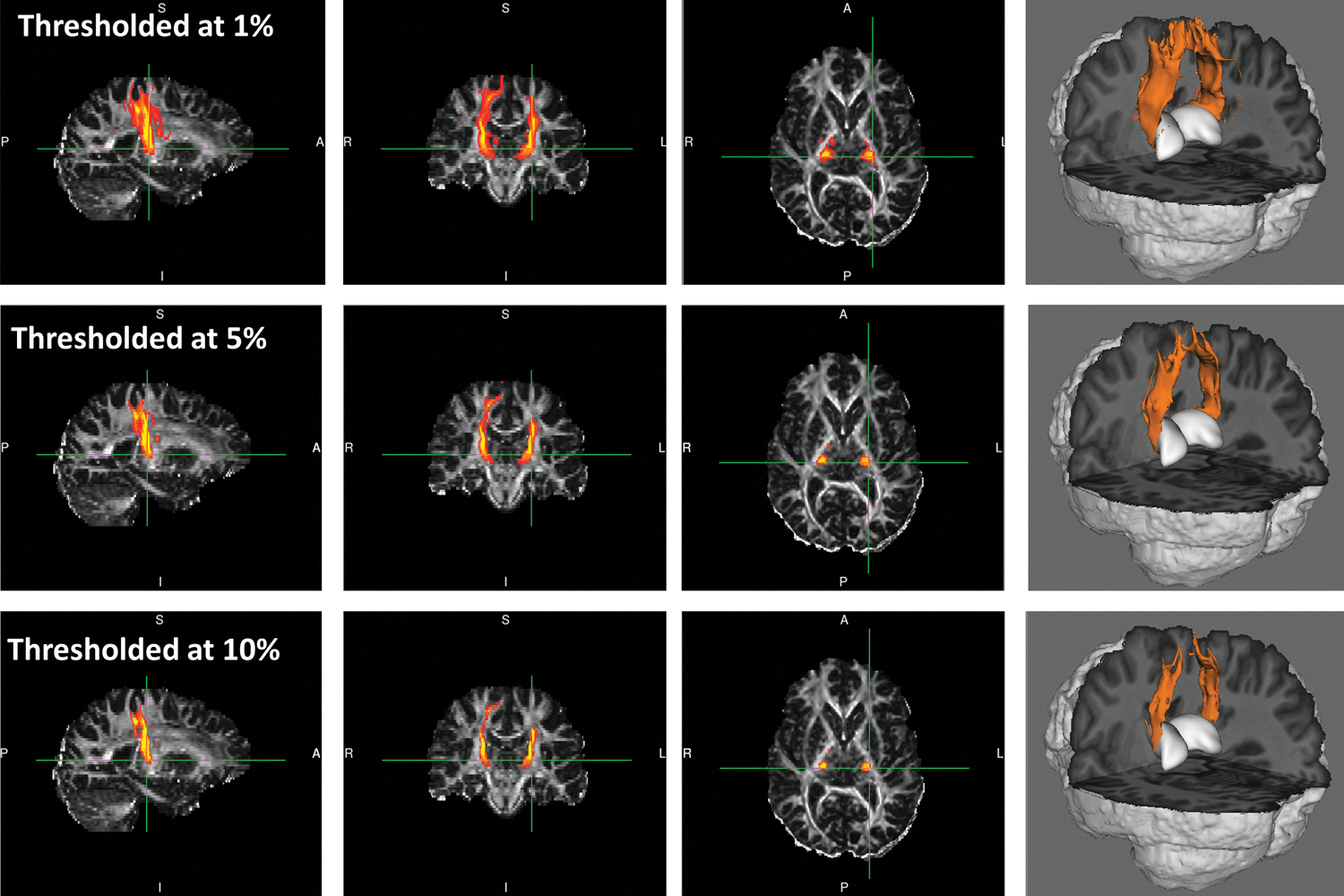
Illustration of the thalamo-SMC connection in one TD subject. Maps of the probability of connections at different thresholds are provided in three-orthogonal views and 3D view to illustrate the connectivity estimation. TD, typically developed.
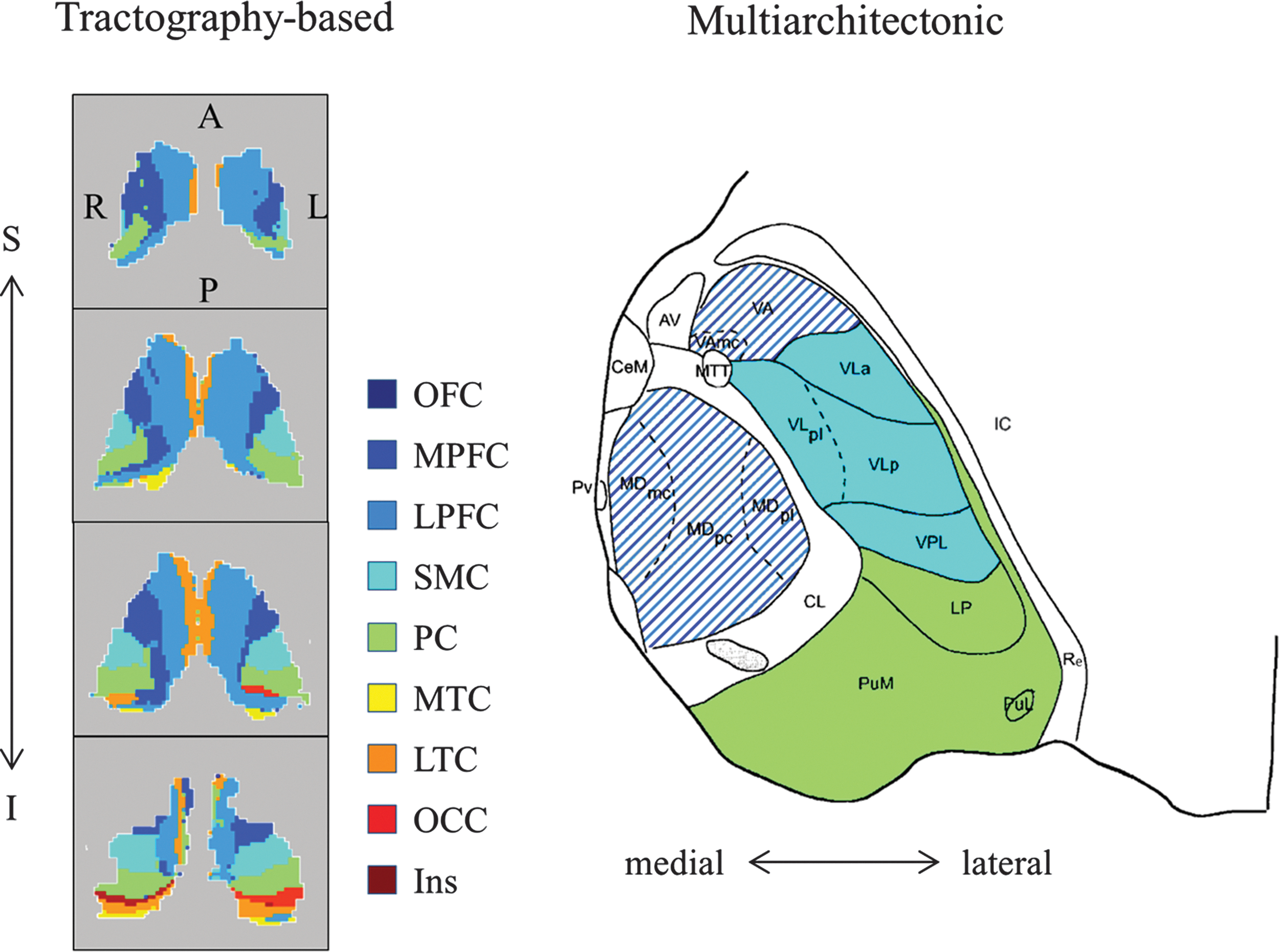
Tractography-based and cytoarchitecture-based thalamic subdivisions side by side. On the left is an illustration of thalamic segments based on probabilistic tractography. Each thalamic voxel was labeled with the color corresponding to the cortical target it is most likely to connect to. The VA and mediodorsal nuclei of the thalamus are connected with the broad frontal cortex and thus marked by interleaved purple and dark blue lines. From top to bottom, every two axial slices of the thalamus from superior to inferior were shown. Given that cross-subject registration of thalamus is a great challenge and is not the main focus of this article, only one TD subject was shown for exemplary purpose. In comparison to the multiarchitectonic division of thalamus on the right (Morel et al., 1997), the thalamic voxels primarily connected to the SMC were in the territories of the VL and VPL nuclei. A, anterior; AV, anteroventral nucleus; I, inferior; IC, internal capsule; L, left; MDmc, mediodorsal nucleus, magnocellular division; MDpc, mediodorsal nucleus, parvocellular division; MDpl, mediodorsal nucleus, paralamellar division; MTT, mammillothalamic tract; P, posterior; PuL, lateral pulvinar; PuM, medial pulvinar; Pv, paraventricular nuclei; R, right; Re, reticular thalamic nucleus; S, superior; VA, ventral anterior nucleus; VAmc, ventral anterior nucleus, magnocellular division; VL, ventral lateral; VLa, ventral lateral anterior nucleus; VLp, ventral lateral posterior nucleus; VPL, ventral posterior lateral.
Given that the sensorimotor territory includes multiple functionally distinct brain regions, explicitly, supplementary motor area (SMA) in the caudal middle frontal cortex, the primary motor cortex (PMC) in the precentral gyrus [excluding the paracentral lobule (PCL)], somatosensory cortex (SSC) in the postcentral gyrus (excluding the PCL), and foot and leg representation of the SMC (PCL), to investigate whether these subregions have distinct thalamic connectivity characteristics, another R-ANOVA analysis was performed to compare the thalamic connectivity between the TD and DYS groups, with the ROI (SMA, PMC, SSC, and PCL) as within-subject factor, group (TD vs. DYS) as between-subject factor, and age and VCI as covariates. No significant main effect of age, ROI, side, or interaction between them was found except for the overall between-subject effect of group (TD vs. DYS); however, the marginal Group by ROI interaction supported our SMC findings (F[3,33]=2.78, p=0.056, ηp 2=0.072), with the interaction primarily driven by PMC and SSC (p=0.005 and 0.107, respectively).
To further examine the white matter tracts corresponding to the SMC findings, the probabilistic tracts thresholded at 10% of maximum was used as a tract mask to calculate the volume, mean FA, and diffusivities of the tracts, with similar R-ANOVA analyses. Overall, the analyses revealed the significant age correlation in diffusivities and FA, while no significant effect was found in group or side. Specifically, the averaged radial diffusivity along the tract is negatively correlated with age (F[1,35]=6.908, p=0.013, ηp 2=0.165), while axial diffusivity is not (p>0.769). As a result, the mean FA along the tract is positively correlated with age (F[1,35]=7.827, p=0.008, ηp 2=0.183). No significant main effect was found for group or side. Tract volume is not different between groups, and not correlated with age, either with or without covariate of intracranial volume.
In summary, no significant difference was found for volumetric sizes of the thalamus or cortical target regions between the TD and DYS groups. The thalamic connectivity with the SMC and the LPFC was found to be greater in the DYS group than in the TD group.
Discussion
This study used diffusion tractography to examine the association between thalamo-cortical connectivity and children's reading ability. Given the thalamus' central role in neural function, we hypothesized that we would see thalamo-cortical connectivity differences in reading related regions between groups of good and poor readers. Results showed no differences in the thalamic connectivity with OTC or the TPC; however, we found group differences in the sensorimotor regions. Specifically, the DYS group showed greater thalamic connectivity than the TD group. An additional finding is that the DYS group had higher thalamo-LPFC connectivity.
The central findings of higher thalamic connectivity with the SMC suggests a thalamo-cortical role in reading that until now has not been a central focus in neuroimaging studies of reading difficulty, although a role of sensori-motor functioning in reading development has long been hypothesized. More specifically, learning to read involves establishing the connection between specific articulatory gestures with associated graphemes. Indeed, sensorimotor and SMA activations were found in overt speech production with silent phonotactic judgment, in support of a role of the SMC in phonetic encoding, articulation, and self-monitoring processes (Indefrey et al., 2001). Furthermore, in the process of establishing this phonological connections with orthographic forms, the ventral posterior medial nucleus of the thalamus is posited to play a key role in acquiring the tactile and proprioceptive feedback from the articulatory apparatus (Heilman et al., 1996). Regarding learning to read, the connections between the thalamus and SMC would be most critical in the early phases of reading acquisition, and would presumably be less relied on with increased reading proficiency. This could reflect a migration of core brain regions in the circuits to deal with new cognitive demands in response to learning and practicing, the result of which is increased automaticity. Actually, this mechanism of migration in brain regions intensively engaged in learning to read has already been observed in extrastriate regions in young readers when performing word generation tasks—a strong recruitment initially, which gradually diminishes with age (Brown et al., 2005; Schlaggar et al., 2002).
From this standpoint, the greater thalamo-SMC connectivity in the DYS group as compared with the TD group may reflect a prolonged multisensory engagement phase in developing reading skills for the children with developmental dyslexia. During development, pruning of gray matter is a central part of maturation. Consistently, it has been shown that the process of cortical brain maturation begins first in dorsal parietal cortices, particularly the primary sensorimotor areas, and then spreads rostrally over the frontal cortex and caudally and laterally over the parietal, occipital, and finally the temporal cortex (Gogtay et al., 2004). This finding, in combination with the greater connectivity of the SMC, may suggest a less mature subcortical-SMC system in children with developmental dyslexia. Indeed, an early motor delay is suggested to be associated with a delay in language development and in acquiring fluent reading skill (Viholainen et al., 2006). Although this finding was obtained from a younger population ranging from 2 to 5 years old, the delay in acquiring reading skills could cascade to the adolescent stage, due to the deficiency in an integrated neural system. Specifically, when a child first comes to read, his/her neural basis for linguistics, such as visual and motor processes, has already been well established; however, these systems are not integrated in a way that supports reading. In response to the demands of learning to read on extant circuits for mapping print to sounds, novel functional connections also form between circuits (Schlaggar and McCandliss, 2007), as seen in the case of multisensory integration studies of letter and letter sounds (van Atteveldt et al., 2004).
Similar to the SMC, a higher thalamic connectivity with the LPFC was also found in the DYS group. It may be that the DYS group over-relies on the thalamic connectivity with the LPFC as a compensatory mechanism, such as over-relying on working memory to mitigate weaknesses, as the prefrontal cortex is well accepted to play a critical role in working memory (Braver et al., 1997), and an important element of working memory is to assist executive functions, including cognitive control (Floresco et al., 1999; Miller and Cohen, 2001). Meanwhile, the prefrontal cortex has prominent afferent projections from the mediodorsal nucleus (MDN) of thalamus, which also receives return projections from the prefrontal cortex (Fuster, 2008); damages in the MDN often lead to disruptive executive functions that require working memory (Daum and Ackermann, 1994; Van der Werf et al., 2003; Zoppelt et al., 2003), indicating an indispensable role of the MDN in the working memory circuitry. Interestingly, the MDN has a divergent connection with many other cortices (Cappe et al., 2009; Klein et al., 2010). These connections with the broad cortical regions form an extensive network surrounding the thalamus, and may provide a convenient way for the supervisory information, perhaps integrated with inputs from other cortices in the thalamus (Cappe et al., 2009), to be sent out to the other cortices, and hence subserve the compensatory attentional regulation of relevant information.
No difference was found in the occipito-temporal and the temporo-parietal regions, which was not as expected. This may be due to the coarse parcellation of the cortical region (i.e., the ipsi-lateral cortex was segmented into nine nonoverlapping segments instead of finer segments), which reduces the specificity of the measurement. However, the stability of the connectivity indices across (repeated MR scans of healthy) subjects decreases as the size of the cortical target regions decreases (due to noise in the imaging data), which will increase the chance of false positives and difficulty to interpret the results as well. We found nine cortical segments is a reasonable balance between measurement stability and anatomical specificity for this particular dataset. Readers should reckon the results reported in this article as preliminary findings. Further investigation will benefit greatly from diffusion MR data with more diffusion weighting directions (i.e., ≥60), which may provide the essential signal-to-noise ratio to enable connectivity analyses on a finer scale in the occipito-temporal and the temporo-parietal regions.
The thalamo-cortical connectivity indices reported in this article is measured as the mean connection ratio with each cortical ROI. The difference was relative, that is, a higher connection ration could be due to more connections to SMC or less connections to the rest of the cortex. However, with MRI, it is difficult to measure the absolute axon quantities. A higher connectivity with SMC, for example, may indicate that the part of the thalamus which is connected with SMC has a larger fraction of connections with SMC with regard to the total connections projecting from (or to) this region. Previous studies have shown that the thalamus is widely connected with the cortex (Behrens et al., 2003a), one particular region inside the thalamus can project to multiple (cerebral) cortical regions (Cappe et al., 2009), and how the thalamus is divided in terms of which cerebral cortical region it is preferentially connected with is largely variable across individuals (Johansen-Berg et al., 2005). Given the integrative role of the thalamus in mediating cerebral cognitive behavior, this structural connectivity characteristic may reveal how effectively the thalamus is integrating, processing, and transfer information among distinct cortical regions during cognitive activities. Nevertheless, the difference reported reflects the intrinsic structural connectivity properties within the thalamus rather than the tracts in the cerebral white matter, as no differences were observed between groups along these tracts.
In summary, the current study examined the in vivo relationship between thalamo-cortical connectivity and children's reading ability. Our findings suggest that children with DYS have substantially greater thalamic connectivity with the SMC and the LPFC. These findings present one possible intriguing interpretation: that during normal reading development, there is an initial reliance on thalamic sensorimotor connectivity for developing phonemic representations and orthographic forms; however, thalamic connectivity for TD readers expands away from primary motor/sensorimotor regions to tertiary cortical regions, resulting in a tuned reading network with increased efficiency. The results suggest that the thalamus may play a key role in reading behavior by mediating the functions of task-specific cortical regions. Clearly, longitudinal experimental designs will be needed to provide more direct evidence for these postulations, but the current study adds knowledge to the field by providing neurobiological evidence of developmental dyslexia in the perspective of subcortical-cortical connectivity.
Conclusion
This study shows that diffusion tractography can be useful to identify evidence of the role of the thalamus in mediating reading behavior in children with TD reading ability and developmental dyslexia. Similar approaches might be useful in the study of subcortical structures in other neurodevelopmental or neurodegenerative disorders.
Footnotes
Acknowledgments
This research was supported in part by grants to Laurie Cutting from the National Institute of Neurological Disorders and Strokes through the Neurobiology and Treatment of Reading Disability in NF1 (7R01NS049096-05), from NIH through Cognitive and Neural Processes in Reading Comprehension (R01HD044073), and from NIH through Predicting Late-Emerging RD: Neurobiological and Cognitive Factors (R01HD067254), to Vanderbilt University Kennedy Center for Research on Human Development (P30 HD15052), and to Vanderbilt University from the National Center for Advancing Translational Sciences (2UL1 TR000445-06) and the National Center for Research Resources (UL1 RR024975-01). This work was conducted in part using the resources of the Advanced Computing Center for Research and Education at Vanderbilt University. The content is solely the responsibility of the authors and does not necessarily represent the official views of the NIH.
Author Disclosure Statement
The authors declare that no competing financial interests exist.