
Abstract
Patients may perceive paradoxical heat sensation during spinal anesthesia. This could be due to deafferentation-related functional changes at cortical, subcortical, or spinal levels. In the current study, the effect of spinal deafferentation on sensory (pain) sensitivity was studied and linked to whole-brain functional connectivity as assessed by resting-state functional magnetic resonance imaging (RS-fMRI) imaging. Deafferentation was induced by sham or spinal anesthesia (15 mg bupivacaine injected at L3–4) in 12 male volunteers. RS-fMRI brain connectivity was determined in relation to eight predefined and seven thalamic resting-state networks (RSNs) and measured before, and 1 and 2 h after spinal/sham injection. To measure the effect of deafferentation on pain sensitivity, responses to heat pain were measured at 15-min intervals on nondeafferented skin and correlated to RS-fMRI connectivity data. Spinal anesthesia altered functional brain connectivity within brain regions involved in the sensory discriminative (i.e., pain intensity related) and affective dimensions of pain perception in relation to somatosensory and thalamic RSNs. A significant enhancement of pain sensitivity on nondeafferented skin was observed after spinal anesthesia compared to sham (area-under-the-curve [mean (SEM)]: 190.4 [33.8] versus 13.7 [7.2]; p<0.001), which significantly correlated to functional connectivity changes observed within the thalamus in relation to the thalamo-prefrontal network, and in the anterior cingulate cortex and insula in relation to the thalamo-parietal network. Enhanced pain sensitivity from spinal deafferentation correlated with functional connectivity changes within brain regions involved in affective and sensory pain processing and areas involved in descending control of pain.
Introduction
Deafferentation is the disruption of afferent and efferent signals between the central and peripheral nervous system (Pawela et al., 2010). Several experimental human and animal studies show that when peripheral sensory and motor input is removed (e.g., by application of ischemia or local anesthetic-induced nerve blocks, cutaneous anesthesia, or peripheral nerve damage) detectable functional changes in the cortex occur (Björkman et al., 2005; Kaas, 1999; Pawela et al., 2010; Waberski et al., 2007; Weiss et al., 2004; Wernhahn et al., 2002). Also subcortical areas, including the thalamus, show changes upon deafferentation (Faggin et al., 1997; Krupa et al., 1999; Nicolelis et al., 1993; Pawela et al., 2010). These changes are best described as reorganization of neuronal interactions due to a rebalancing of excitatory and inhibitory factors that mediate adaptation and neuronal plasticity (Kaas, 1999). For example, the loss of sensory and motor input from the hand by peripheral nerve blockade is associated with supraspinal excitatory changes possibly mediated by disinhibition of unmasked (interhemispheric) cortical neuronal connections, and explains the enhanced functionality of the contralateral hand (Waberski et al., 2007; Weiss et al., 2004). These cortical changes coincide often with perceptual changes.
One form of deafferentation that is performed yearly in millions of patients worldwide is spinal anesthesia where the sensory information from the lower part of the body is temporary removed to allow surgical intervention without the perception of pain. It is well known that spinal anesthesia may coincide with sensory distortions. For example, some patients report body image distortions (such as swelling of the legs, illusionary limb position, and changes of the length of the limbs) during regional (including spinal) anesthesia (Gandevia and Phegan, 1999; Paqueron et al., 2003, 2004; Silva et al., 2011). Additionally, the affected limbs are often perceived as warm upon the administration of the local anesthetic while some patients perceive paradoxical heat sensation above the level of the anesthetic block (i.e., a cold stimulus is perceived as warm) during the assessment of the spread of the spinal anesthetic (Kanai et al., 2010; Paqueron et al., 2003). These observations are typically made during the initial rapid rise of the anesthetic level and are suggestive of changes in central sensory modulation, possibly related to the deafferentation from the spinal block. There is further observation that epidural anesthesia (another form of deafferentation) can lead to occurrence of painful sensations in the deafferented area in an otherwise healthy individual (Mihic and Pinckert, 1981). Existing evidence presented above suggests that deafferentiation from spinal anesthesia would lead to a change in functional organization of cortical and subcortical networks involved in sensory motor perception and pain. Possibly, the altered sensory perceptions during spinal anesthesia and functional changes in cortical and subcortical areas of the brain are causally related. Some evidence to that suggestion comes from data in patients where hyperexcitability of thalamic neurons coincides with neuropathic deafferentation pain (Rinaldi et al., 1991).
A well-known paradigm to evaluate the efficacy of the endogenous pain modulatory system is “conditioned pain modulation (CPM)” (Dahan et al., 2012; Ossipov et al., 2010). The CPM paradigm assumes that adding afferent nociceptive input at a remote area of the body inhibits the intensity of primary focal pain stimulus (“pain inhibits pain”) through activation of supra spinal centers including the anterior cingulate cortex (ACC), the insula, and the prefrontal cortex. Therefore, it would be plausible that blockade of afferent input would have the reverse effect on pain perception. This means that if afferent input becomes “disconnected,” then pain perception would become more intense. However, there are no human studies assessing the effect of pain perception on areas remote from deafferentation sites (such as pain perception on the arm during spinal anesthesia). We aim to use this model of acute deafferentation by spinal anesthesia in healthy participants to further understand the mechanisms involved in endogenous modulation of pain.
Our placebo (sham-spinal anesthesia), crossover, randomized study investigates (1) whether pain perception above the level of the anesthetic is altered by spinal deafferentation, and (2) whether we can detect an coinciding change in resting-state functional connectivity of cortical and thalamic networks in healthy humans. The thalamus was chosen as region of interest as it is an important pain modulatory center that receives input from multiple ascending pain pathways, and projects to various (sensory and affective) pain modulatory regions of the cortex and limbic system (Ab Aziz and Ahmad, 2006; Apkarian et al., 2005). In this study, we obtained repeated resting-state functional magnetic resonance images (RS-fMRI) in two sessions (spinal and sham spinal peripheral anesthesia). It has been shown that this technique can be reliably used to evaluate alterations in intrinsic brain connectivity following pharmacological interventions in humans (Cole et al., 2013; Khalili-Mahani et al., 2012; Klumpers et al., 2012; Niesters et al., 2012), and deafferentation in rats (Pawela et al., 2010).
Materials and Methods
Subjects
Twelve right-handed, healthy, male volunteers (age: 23.7 [3.4] year [mean (SD)]; body mass index: 21.3 [2.4] kg/m2) were enrolled in the study after approval by the local Institutional Review Board of the Leiden University Medical Center in Leiden, The Netherlands. All participants gave oral and written informed consent. The study was performed according to GCP guidelines and the ethical principles for medical research involving human subjects of the international Association of the Study of Pain (
Study design
The study was performed using a randomized crossover design. Upon arrival, an intravenous line was placed in the right hand or arm to allow fast administration of emergency medication when necessary. Next, baseline anatomical MRI (T1-weighted) and baseline RS-fMRI scans were obtained followed by baseline heat pain measurements. After baseline measurements were complete, subjects received an intrathecal injection with a local anesthetic on one occasion and a sham procedure on the other as described below (time of injection is t=0). Responses to heat pain and the height of the sensory block (measured by the response to a cold 4 cm2 surface applied to the skin in the left and right mid-axillary line) were measured at 15-min intervals. Additional RS-fMRI scans were obtained 1 and 2 h after the spinal injection or the sham procedure. At the end of the study, subjects were monitored until fully recovered from the spinal anesthetic, as defined by return of motor functions and diuresis, and then allowed to go home.
Intrathecal injection and sham procedure
The intrathecal injection was performed at the interspace between vertebrae L3 and L4 with 3 mL bupivacaine 5 mg/mL (AstraZeneca, Zoetermeer, The Netherlands) after the skin was locally infiltrated with 1–2 mL lidocaine 10 mg/mL (AstraZeneca). For the spinal puncture a 27 gauge Whitacre needle (Vygon, Valkenswaard, The Netherlands) was used to minimize the risk of postspinal headache. The sham procedure was performed by insertion of a spinal needle at the interspace between vertebrae L3 and L4 through the skin, after the skin was locally infiltrated with 1–2 mL lidocaine 10 mg/mL. The dura mater was not punctured and no bupivacaine was injected. An independent anesthesiologist, who was not involved in conducting or analyzing other measurements made during the study, performed the injections. The instructions to the subject were similar on both occasions so that the subject and the investigators did not know which treatment was given at the moment of injection.
Pain assessment
Heat pain was induced on the lower part of the left (i.e., nondominant) arm with a 3×3 cm thermal probe of the Pathway Neurosensory Analyzer (Medoc Ltd., Ramat Yishai, Israel). Baseline temperature was set at 32°C. During heat pain tests the temperature of the probe gradually increased (1.5°C/sec) toward a preset destination temperature that was held constant for 30 sec and then rapidly (6°C/sec) returned to baseline temperature. To quantify pain intensity of the heat pain stimulus, subjects rated the perceived pain stimulus using a computer-connected slider on an electrical potentiometer that ranged from 0 mm (no pain) to 100 mm (worst pain imaginable). This allowed for continuous electrical monitoring of the visual analogue scale during the noxious stimulation. The target temperature of the heat stimulus was determined at the start of each study day and was intended to evoke an electronic visual analogue scale (eVAS) of 40 mm. To evaluate pain responses after the intrathecal injection or sham procedures, pain tests were applied between imaging sessions at fixed time points: t=15, 30, 45, 90, 105, and 150 min.
RS-fMRI acquisition
A 3-Tesla Achieva Scanner (Philips Medical System, Best, The Netherlands) was used to acquire functional data at fixed time points (baseline, t=60 and t=120 min). The neuroimaging protocol included a high-resolution T1-weighted scan (repetition/echo time=9.7/4.6 msec, flip angle=8 degrees, 1 mm isotropic, 4 min) and three RS-fMRI series (each 220 T2*-weighted whole-brain volumes, obtained with a gradient echo planar with repetition/echo time=2180/30 msec, flip angle 80 degrees, 3.44 mm isotropic, duration 8 min; subjects were instructed to keep their eyes open and relax). A high-resolution T2*-weighted scan (∼30 sec) was acquired at the end of each repeated RS-fMRI to facilitate registering the functional data to the anatomical image.
RS-fMRI analysis
The following prestatistics processing was applied using FSL software on all individual RS-fMRI scans: motion correction; registration to standard space by applying six rigid-body transformations between RS-fMRI and high-resolution T2*, and high resolution T1, followed by an affine registration to the MNI152 template with 2 mm resampling (Smith et al., 2004); brain extraction; spatial smoothing using a 5-mm full width at half-maximum Gaussian kernel; grand-mean intensity normalization; and high-pass temporal filtering (Gaussian-weighted least-squares straight line fitting, with a 0.01 Hz cut-off).
Functional connectivity was assessed using two different approaches. First, to evaluate the general effects of deafferentation from spinal anesthesia on functional brain connectivity, we used a model-free analysis of eight predefined networks of interest (NOIs) as previously described by Khalili-Mahani et al. (2012). These canonical networks represent 80% of the total brain volume and are described based on their general function as the medial and lateral visual network, the auditory and somatosensory network, the sensorimotor network, the default mode network, the executive salience network, the visual-spatial network, and the working memory network (Beckmann et al., 2005). As we have previously shown for morphine, alcohol, δ(9)-tetrahydrocannabinol and ketamine, RS-fMRI data analysis using these networks reveals localized and drug-specific changes in functional brain connectivity (Cole et al., 2013; Khalili-Mahani et al., 2012; Klumpers et al., 2012; Niesters et al., 2012).
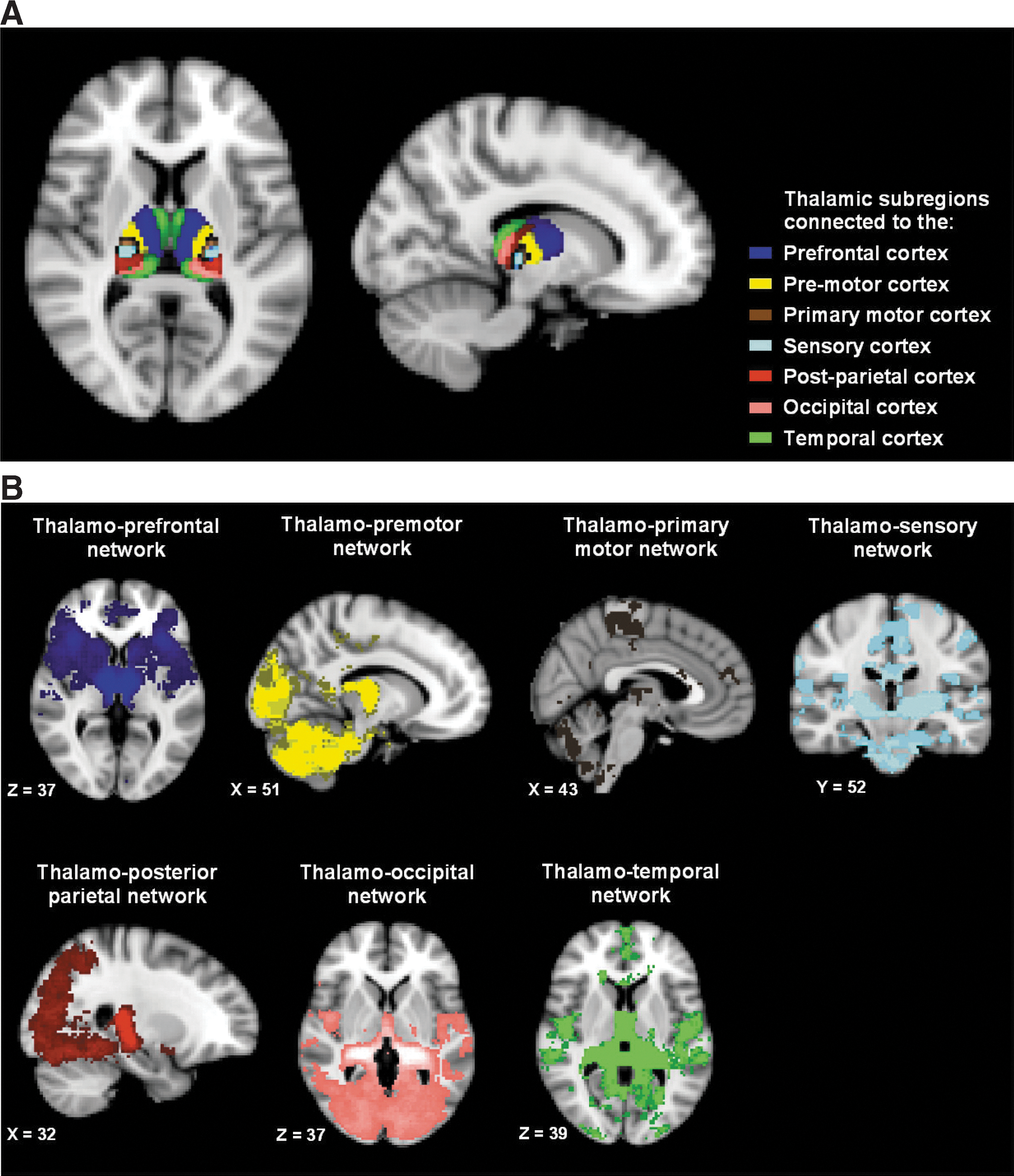
The second functional connectivity analysis focused on functional connectivity in relation to the thalamus. The thalamus was chosen as it receives projections from multiple ascending pain pathways, is involved in processing nociceptive information, and projects the information to various parts of the limbic and cortical structures involved in sensory discriminative and the affective dimensions of pain perception (Ab Aziz and Ahmad, 2006). We used seven thalamic subregions according to the Oxford thalamic connectivity atlas (Johansen-Berg et al., 2005). Our choice of this atlas is motivated by our principle to develop objective, easily reproducible, and standardized procedures for replication studies. The important advantage of this atlas is that it is constructed based on probabilistic diffusion tractography that describes the probability of corticothalamic white matter fibers connection between thalamic subregions and cortical segments (prefrontal cortex, temporal cortex, premotor cortex, primary motor cortex, sensory cortex, posterior-parietal cortex, and the occipital cortex). We refer to the resulting functional networks as the thalamo-prefrontal, thalamo-premotor, thalamo-primary motor, thalamo-sensory, thalamo-parietal, thalamo-occipital network, and thalamo-temporal network to indicate the reference region.
In both analyses, we used a dual regression analysis to define resting-state networks (RSNs) in relation to reference regions (the eight canonical NOIs or the seven subthalamic segments) (Beckmann et al., 2009). Briefly, dual regression involves multiple-regression of RS-fMRI time-series against several NOIs or thalamic subregions to estimate a representative vector of BOLD fluctuations within each reference region, and next regressing the RS-fMRI time-series against these time vectors to identify spatial representations of RSNs, that is, brain areas with similar fluctuations patterns as the reference regions. Nuisance variables corresponding to fluctuations in the deep white matter (measured from the center of the corpus callosum) and cerebrospinal fluid (measured from the centre of lateral ventricles) were included in the dual regression analysis to account for nonspecific and physiological variations (Birn, 2012). This resulted in statistical maps of z-scores, where each voxel of the brain represents the functional connectivity between that voxel and each of the NOIs or the thalamic subregions. These statistical maps were later used for voxel-wise inference testing of the spinal anesthesia on each network.
Data, power, and statistical analyses
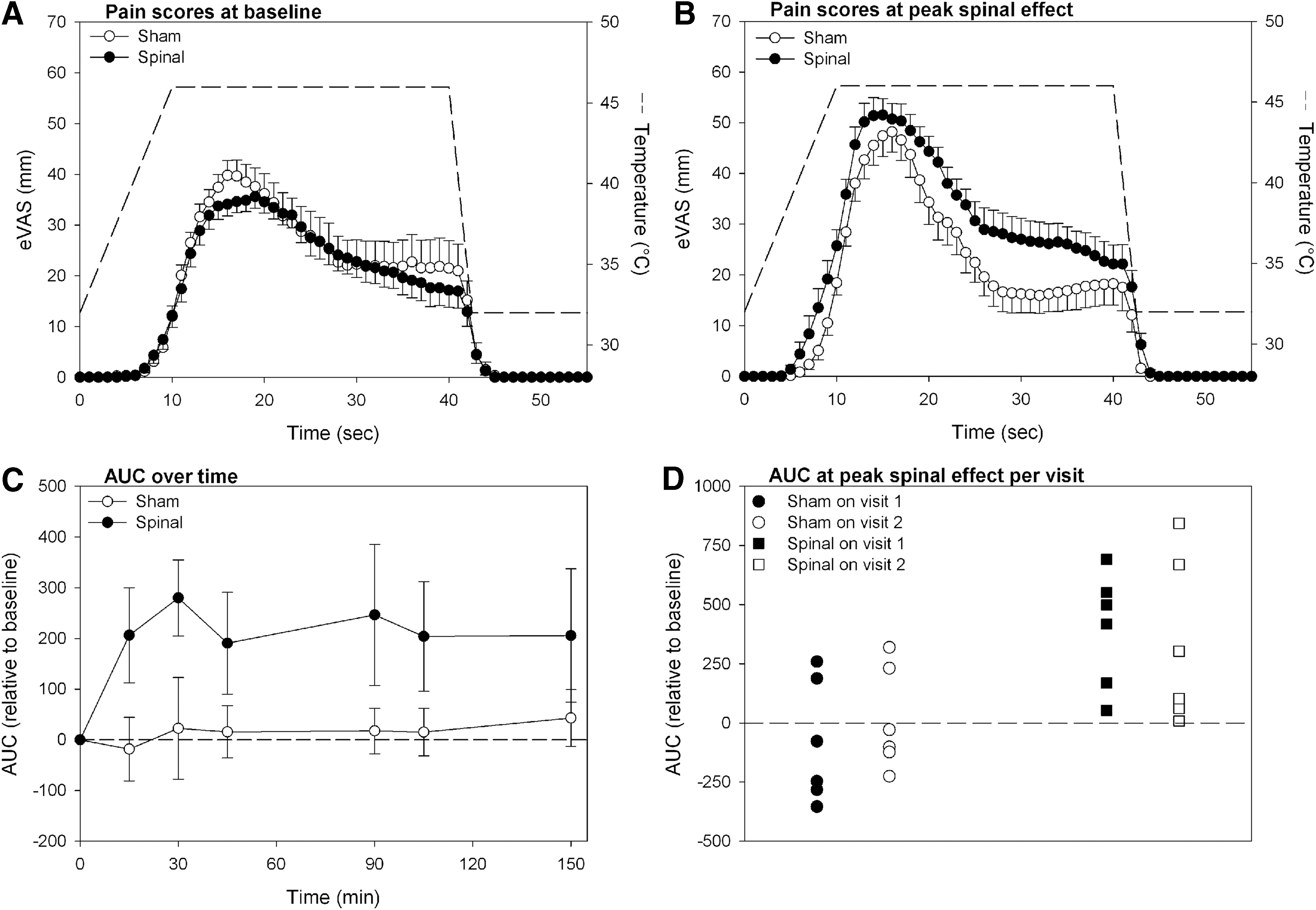
To quantify pain intensity, the area-under-the-curve (AUC) of each eVAS response was calculated and presented relative to the baseline measurement (Niesters et al., 2011). The study was powered to detect a 50% treatment difference in the eVAS AUC at peak spinal level (estimated SD 35%, α=0.05, 1—β=0.9).
The effect of spinal anesthesia on pain perception was tested by a repeated measures analysis of variance with post-hoc Bonferroni correction on the AUC values relative to baseline. The statistical analysis was performed in SigmaPlot version 12.0 (Systat Software, Inc., Chicago, IL) and p-values<0.05 were considered significant. Data are presented as mean (SEM) unless otherwise stated.
To determine the effect of deafferentation from spinal anesthesia on resting-state functional connectivity a mixed-effects analysis was applied with subject as random and time and drug as fixed within-subject variables. Voxel-wise statistical analysis on the z-score connectivity maps was performed using a permutation-based statistical inference with 5000 permutations. Statistical significance was set at p-value<0.05 after family wise error cluster-based correction (with cluster forming voxel-wise thresholds set at p<0.01) (Forman et al., 1995). To further control for spurious effects, we report clusters that included a minimum of 10 adjacent voxels, 80 mm3. We also performed a stepwise regression (without and with pain score as regressor in the model) to examine brain regions whose connectivity was modulated by the subjective perception of pain. In all stages of MRI analyses the FMRIB Software Library was used (FSL 4.1; Oxford, United Kingdom) (Smith et al., 2004).
Results
Spinal anesthesia
All subjects completed the study without the occurrence of major side effects. Peak sensory blockade was achieved after 45 min with 17.5 (1.0) blocked segments corresponding to a sensory block level from dermatomes S5 to Th5. This sensory block persisted throughout the whole study period. The mean time of spinal anesthesia to full recovery of diuresis and motor function was 369 (11) min. No sensory blockade was observed after the sham procedure in any of the subjects. The spinal anesthetic and sham procedure did not result in significant cardiorespiratory changes. Blood pressure remained within 5% of control values. Due to the absence of spinal block following the sham procedure, blinding of the study was rapidly lost to both investigators and volunteers.
Pain responses
The mean eVAS responses prior to treatment and at peak spinal anesthetic level and sham experiments are given in Figure 1A and B. Spinal anesthesia significantly increased pain sensitivity on the skin of the lower forearm. Mean AUC values at baseline were 844.7 (63.2) mm·s on the study day with spinal injection and 898.6 (122.6) mm·s on the day of the sham procedure (p=0.644). Mean AUC values at peak spinal level were 1165.0 (71.0) mm·s after spinal injection and 877.1 (105.8) mm·s after the sham procedure (p=0.005). Mean AUC values over time are shown in Figure 1C showing that the hyperalgesic responses lasted for at least 3 h (end of the study). There was no effect of study order on the pain AUC values (Fig. 1D).
Effect of spinal anesthesia on predefined general RSNs
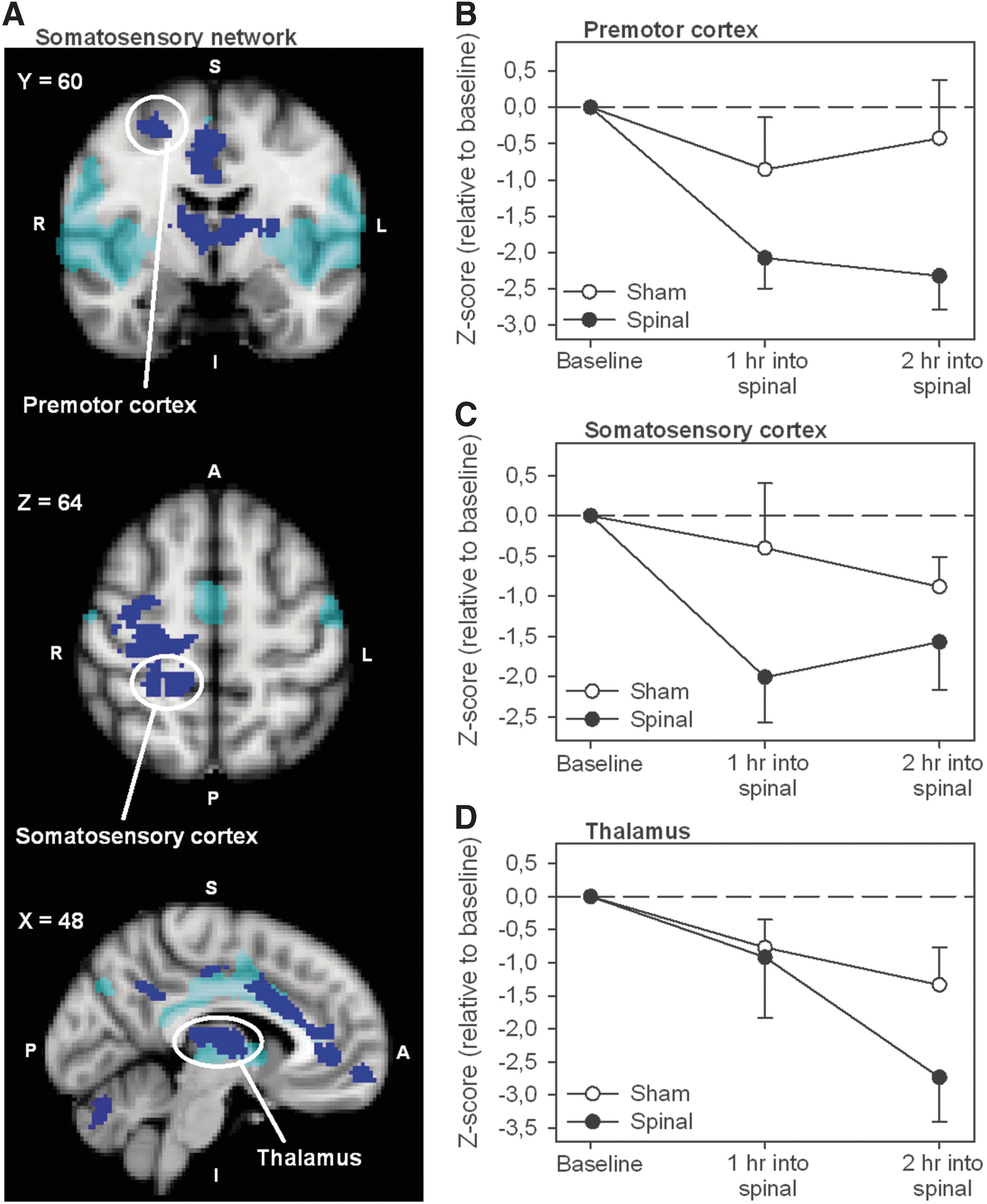
Spinal anesthesia induced significant changes in functional connectivity in relation to three of the eight canonical NOIs: the medial visual network (increase), the somatosensory network (decrease) and the default mode network (increase). Regions that demonstrate functional connectivity changes in relation to these three networks are given in Table 1 and include among others the thalamus, primary somatosensory cortex, primary motor cortex, premotor cortex, ACC, caudate nucleus, and the cerebellum. Figure 2A demonstrates the statistical connectivity map (cluster corrected; p<0.05) of the brain areas with a decrease in functional connectivity in relation to the somatosensory network. The effects of spinal anesthesia on functional connectivity over time for the premotor cortex, primary somatosensory cortex, and thalamus are shown in Figure 2B–D. Details regarding cluster size, z-score, and location of the areas that show functional connectivity changes are provided in Table 1. Adding treatment order (sham first vs. spinal first) as a covariate to the statistical model did not affect the number, extent, and location of these regions.
Effect of Spinal Anesthesia on Functional Connectivity in Relation to the Predefined General Resting-State Networks (Cluster p-Value <0.05)
Voxel dimension is 2×2×2 mm (voxel volume 0.008 mL); regions can be located within or outside the resting-state network.
B, bilateral; L, left; R, right.
Thalamic RSNs
Cortico-thalamic connectivity maps are shown in Figure 3A. To evaluate whether anatomically distinguishable networks were produced by the dual regression analyses of the thalamic subregions, average functional connectivity maps were obtained for each thalamic RSN. This was done by first thresholding and binarizing each functional connectivity map at a z-score>4.0 and next computing a probability map (with probabilities of connectivity>50%). Figure 3B represents the average functional connectivity probability maps of the RS-fMRI data acquired at baseline for all seven thalamic subregions. With one exception, all thalamic subregions were functionally connected to cortical areas as expected according to the anatomical atlas. The exception was one thalamic subregion that instead of predominantly connecting to the premotor cortex (as expected from the atlas) demonstrated functional connectivity to the occipital cortex and cerebellum (the yellow areas in Fig. 3B).
Effect of spinal anesthesia on thalamic RSNs
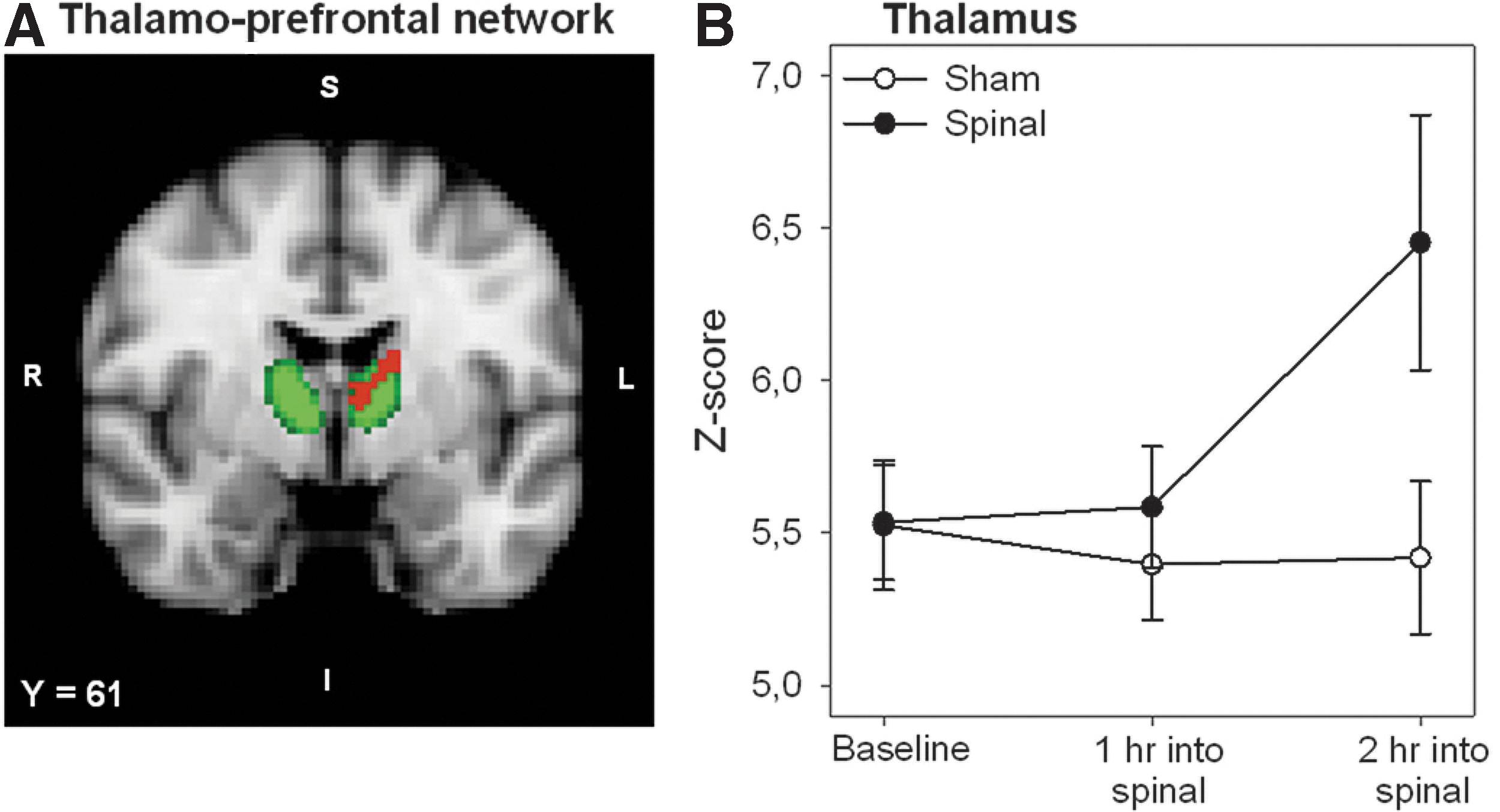
Spinal anesthesia induced a significant increase in functional connectivity in relation to three of the seven thalamic networks: the thalamo-prefrontal, the thalamo-parietal, and the thalamo-temporal network. These networks are involved in the sensory discriminative (i.e., pain intensity) and affective components of pain (Ab Aziz and Ahmad, 2006). Regions that show connectivity changes in relation to the three networks are given in Table 2 and include (partly) similar regions observed in the general RSN analysis: thalamus, primary somatosensory cortex, primary motor cortex, and ACC. Additional affected regions are the insula, precuneus cortex, the frontal lobe, and the posterior cingulate gyrus. The effect of deafferentation from spinal anesthesia in relation to the thalamo-prefrontal network is presented in the statistical connectivity map of Figure 4A, which shows a significant increase on thalamic connectivity (in red) overlapping the subregion of the thalamus that functionally and anatomically connects to the prefrontal cortex (in green). The main effect of treatment over time in this thalamic region is shown in Figure 4B. The significant effect from deafferentation from spinal anesthesia in relation to the thalamo-posterior parietal network is presented in the statistical map of Figure 5A. The main effect of treatment over time for the ACC, the posterior cingulate gyrus, and the insula is given in Figure 5B–D with significant treatment changes during the complete course of measurement. Details regarding cluster size, z-score, and location of the affected areas are provided in Table 2. Study order did not affect the location and extent of these clusters.
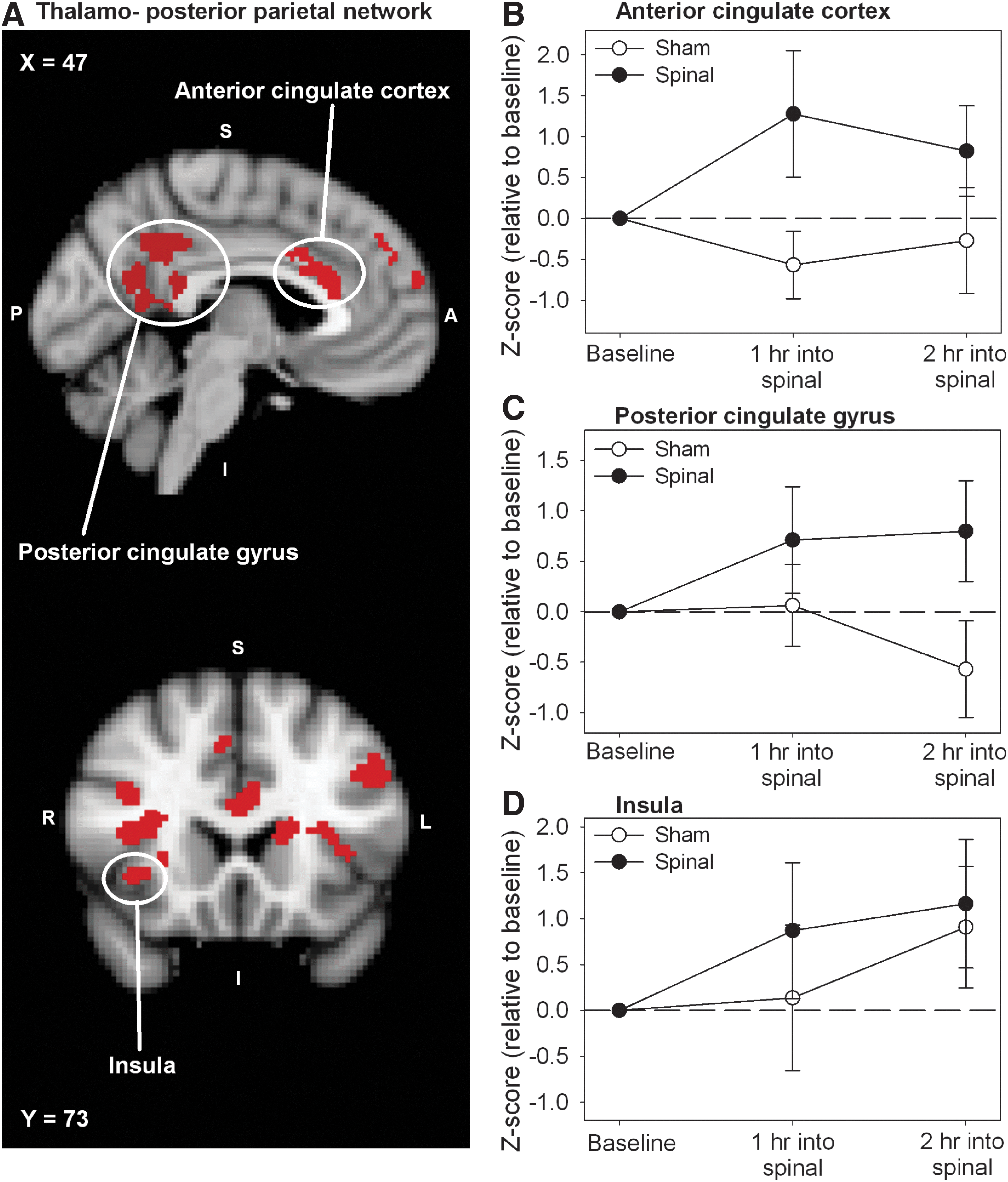
Effect of Spinal Anesthesia on Functional Connectivity in Relation to the Thalamic Resting-State Networks (Cluster p-Value <0.05)
Voxel dimension is 2×2×2 mm (voxel volume 0.008 mL); regions can be located within or outside the resting-state network.
Regions that disappear when pain is added as covariate to the statistical model.
Correlations between pain and functional resting-state connectivity
Table 2 lists brain areas whose connectivity was altered by including the absolute AUC pain scores as a covariate in the permutation testing (i.e., connectivity changes increased with greater pain scores). Illustrative examples of significant correlations observed between functional connectivity changes and pain responses are given in Figure 6 for the thalamus (in relation to the thalamo-prefrontal network, i.e., intra-thalamic), ACC, and insula (both in relation to the thalamo-parietal network).
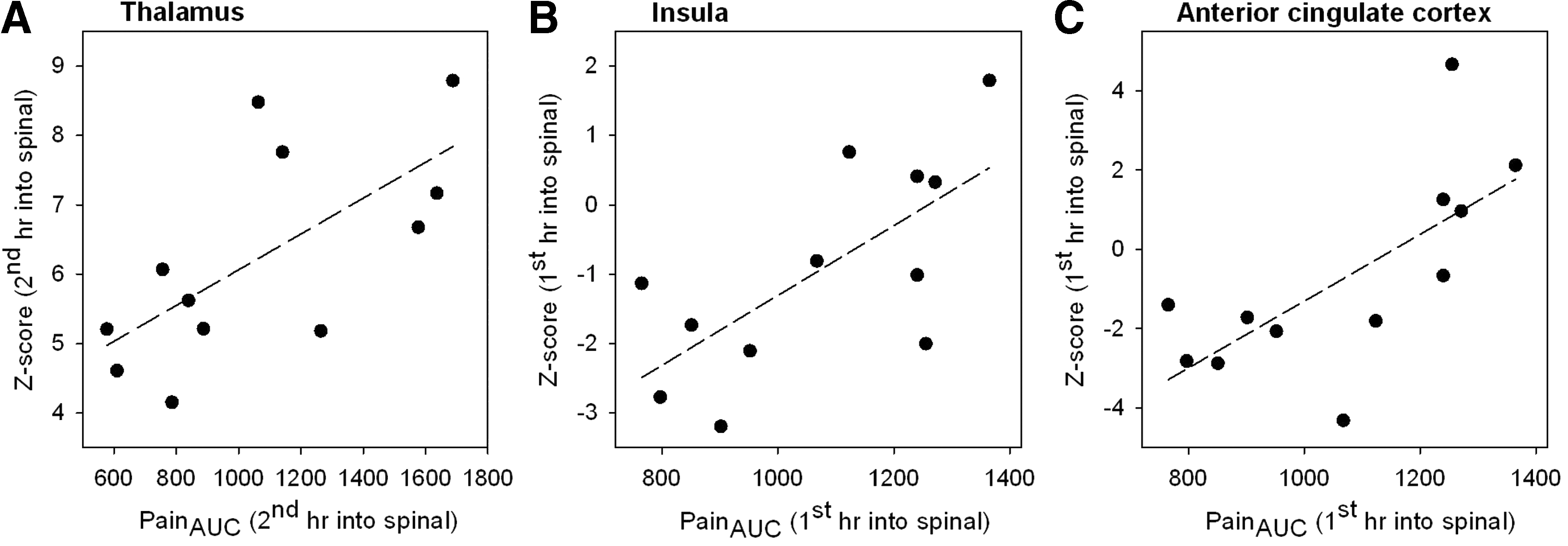
Illustrative scatterplots of the pain response area-under-the-curve scores in relation to the absolute connectivity z-scores for the
Discussion
Our hypothesis that spinal deafferentation would enhance pain sensitivity was confirmed by our finding that nociceptive stimuli applied to dermatomes above the level of spinal deafferentation were perceived as hyperalgesic. This observation is suggestive of transient central cortical and subcortical changes in neuronal organization. Indeed, we observed spinal deafferentation-induced connectivity changes in brain networks involved in the sensory discriminative dimension (e.g., thalamus, insula, and somatosensory cortex) and in the affective dimension (e.g., brainstem, thalamus, insula and ACC) of pain perception from two independent analyses of canonical NOIs and thalamic networks (Atlas et al., 2014; Brooks and Tracey, 2005). Furthermore, the increased pain sensitivity at nondeafferentated skin areas was correlated to thalamo-cortical connectivity changes within the thalamus, ACC, and insula. Our findings are in agreement with earlier animal and human studies showing that deafferentation is associated with changes in neuronal organization in the cortex and subcortical areas. These changes are associated with warm and referred sensations, perceptual illusions, neuropathic pain, and enhanced sensorimotor function of nondeafferented areas (Björkman et al., 2004, 2005; Kanai et al., 2010; Lundborg, 2000; Mihic and Pinckert, 1981; Navarro et al., 2007; Paqueron et al., 2003; Rinaldi et al., 1991; Silva et al., 2011; Waberski et al., 2007; Weiss et al., 2004; Werhahn et al., 2002).
Effect of deafferentation on canonical resting-state networks
In the current study, the RS-fMRI technique was successfully used to evaluate deafferentation-induced changes in brain connectivity in awake humans. The changes in RSN connectivity induced by the symmetric spinal deafferentation included areas involved in the sensory discriminative components of pain perception (sensory cortex, (pre)motor cortex, brainstem, thalamus) and the affective dimension of pain (insula, caudate nucleus, frontal pole, ACC, thalamus, brainstem and cerebellum), in relation to the medial visual network (increase in RSN connectivity), the somatosensory network (decrease) and the default mode network (increase) (see Table 1).
In two previous studies in an anesthetized rat model the effects of traumatic peripheral nerve- or spinal cord injury (SCI)-related deafferentation were studied using RS-fMRI (Pawela et al., 2010; Seminowicz et al., 2012). Both studies show changes in connectivity between the thalamus and cortical and subcortical areas of the brain (e.g., the primary somatosensory cortex). The authors argue that these changes are related to the loss of inhibitory influences within these brain neuronal networks. There is general agreement in the literature that deafferentation causes a rebalancing of excitatory and inhibitory neuronal activity toward disinhibition exposing formerly masked excitatory connections (Björkman et al., 2005; Faggin et al., 1997; Kaas, 1999; Nicolelis et al., 1993; Pawela et al., 2006; Seminowicz et al., 2012; Waberski et al., 2007; Weiss et al., 2007; Wernhahn et al., 2002). Krupa et al. (1999) further show that also feedback from cortex to thalamus plays an important role in plastic changes due to deafferentation [see also Kaas (1999)]. These deafferentation-related changes may be due to alterations in neuronal activity, such as due to reduced GABAergic inhibitory activity and/or enhanced glutamatergic excitatory activity, or due to changes in microcirculation, where reduced afferent input changes the neurovascular coupling (Jacobs and Donoghue, 1991; Koyama et al., 1993; Pawela et al., 2010). Synaptic sprouting and development of structural changes between brain areas take more time to develop and seem to play a role in chronic deafferentation (in SCI, peripheral nerve injury or amputation) (Lundborg, 2000; Navarro et al., 2007; Pawela et al., 2010). Given the fact that we are unable to determine from the RS-fMRI analyses whether changes in connectivity coincide with increases or decreases in neuronal activity, attribution of the observed changes in RS-fMRI connectivity during spinal anesthesia to a shift from inhibitory toward excitatory nociceptive pathways is currently at best speculative.
We observed changes in connectivity relative to medial visual, somatosensory, and default mode networks. The reason for the selective association of spinal deafferentation with connectivity changes relative to these specific canonical networks cannot be deduced from our study. Possibly compared to the other networks, these networks are most sensitive to loss of peripheral afferent input. Irrespective of the mechanism, we argue that the observed changes may cause specific behaviors associated with neuraxial blockade. For example, epidural anesthesia is associated with block-height dependent sedation and reduced brainstem auditory evoked potentials (Doufas et al., 2004). Further, several studies show that neuraxial blockade coincides with sedation and consequently reduced (volatile and intravenous) anesthetic requirements (Hodgson et al., 1999; Lu et al., 2010). These effects may be related to connectivity changes relative to the default mode and medial visual networks (Boveroux et al., 2010; Heine et al., 2012). Particularly the default mode network seems important in altered states of consciousness (anesthesia, coma, vegetative state, epileptic loss of consciousness and somnambulism) (Boly et al., 2008; Boveroux et al., 2010). We did not measure the arousal state in our study. Due to this limitation we cannot conclude whether in our population a change in arousal state occurred. Changes observed relative to the somatosensory network may be associated with nociceptive sensations (warm sensation/paradoxal heat sensation, and as observed here: hyperalgesia) and illusions of abnormal bodily position and recognition (Paqueron et al., 2003; Silva et al., 2011).
Effect of deafferentation on pain responses and thalamic resting-state networks
An important observation of this study is that pain sensitivity increased during spinal deafferentation. Similar observations were made in rats following experimental SCI where allodynia is perceived at dermatomes above the transection level in a majority of animals (Endo et al., 2008; Gerke et al., 2003). Gerke et al. (2003) further showed increased spontaneous firing of thalamic neurons in rats following SCI. Several other studies show spatio-temporal changes and neuronal hyperactivity in the thalamus upon deafferentation (either in experimental animal models or in patients with deafferentation pain), with augmented connections between the primary somatosensory cortex and the thalamus (Jung and Shin, 2002; Seminowicz et al., 2012). Consistent with these findings, we observed changes in functional connectivity within the thalamus in our general RSN analysis (Fig. 2D). The more specific analysis of the thalamic subnetworks revealed significant increases in connectivity between the thalamus and regions of the brain involved in sensory and affective pain processing and perception (Figs. 4 and 5; Table 2). This indicates the importance of neuronal activity changes in the thalamus upon deafferentation. Importantly, the enhanced pain sensitivity was also correlated with the thalamic RSN connectivity (Table 2). Positive correlations were observed between pain scores and intrathalamic and thalamo-cortical (involving the ACC and insula) functional connectivity (Fig. 6A–C), suggestive of a causal role for these networks in enhancement of pain sensitivity during acute deafferentation.
Interestingly, several brain areas that we identified in the hyperalgesic responses to deafferentation (e.g., thalamus, insula, and ACC) are involved in descending modulation of pain, where activation of these supraspinal brain areas causes either facilitation or inhibition of afferent nociceptive input at the level of the spinal cord dorsal horn (Brooks and Tracey, 2005; Ossipov et al., 2010). This suggests that spinal anesthesia-induced deafferentation causes the shift of the endogenous pain system toward pain facilitation. Our findings therefore support the CPM paradigm as we now observed that blockade of afferent inputs [i.e., the reverse of the CPM paradigm (Dahan et al., 2012; Ossipov et al., 2010)] enhances pain sensitivity. Of interest is that You et al. (2012) identified the medio-dorsal subregion of the thalamus of the rat being involved in pain facilitation as part of the endogenous pain modulatory system. This region corresponds to the human thalamic subregion anatomically connected to the prefrontal cortex (cf. Fig. 4). Several studies on chronic (deafferentation) pain syndromes have also observed altered functionality in these same brain regions. For example, Apkarian et al. (2004) showed that chronic low back pain was associated with abnormalities (i.e., loss) of the thalamus (and prefrontal) gray matter density. SCI in primates leads to a functional reduction of the GABAergic inhibitory circuitry of the thalamus, and in humans, abnormal thalamic bursting patterns and abnormal activity patterns in the ACC were observed following SCI (Dostrovsky, 2000; Gustin et al., 2010; Ralston, 2005; Saab, 2012). Knowledge on the mechanism of both afferent and efferent signaling pathways is important for our understanding of the (ab)normal perception of pain and may lead to new insights for the treatment of pathological pain syndromes. Speculating that the enhanced pain sensitivity we observed in dermatomes above the deafferentation level is associated with excitatory changes in thalamo-cortical connectivity, a therapy focused on inhibition of these excitatory networks may be indicated. For example, pain relief may occur by reconstituting GABAergic inhibitory activity, or inhibition of glutamatergic excitatory activity. Indeed, recent studies indicate that the N-methyl-D-aspartate receptor antagonist ketamine induces long-term relief of neuropathic pain by improving descending pain inhibition, possibly via a central inhibitory effect on excitatory pathways (Niesters et al., 2012, 2013; Sigtermans et al., 2009).
Blinding
The inability of blinding the anesthetic treatment in both subjects and investigators in our study is inevitable with the procedure and paradigm in question. Anticipation is a critical aspect of subjective pain perception and it is plausible that awareness of subjects of the nature of the effect of the spinal injection could have affected the study outcome. We controlled for possible experimental order effects and deblinding by including the order effect in our statistical model. In our small sample, we did not observe any order effect on the subjective scoring of pain intensity (Fig. 1D), nor did we find an effect on the RS-fMRI results. This, however, is not generalizable and some effect due to differences in the attention to the thermal pain in spinal versus sham sessions cannot be excluded (Miron et al., 1989; Wiech et al., 2008). Possibly, such an interoceptive effect became visible in the insula signal at 2-h into the sham spinal (Fig. 5D) (Craig, 2009).
The insula
In this study we focused on the thalamus in relation to other brain areas to explain the observations of hyperalgesia following spinal analgesia. We are aware that other important pain areas of the brain were involved in the effect of spinal deafferentation on pain sensitivity, such as the insula and ACC. The insula is involved in the sensory and affective dimensions of pain perception and in the processing and modulation of interoceptive sensations (Brooks and Tracey, 2005; Craig, 2009; Rainville, 2002). Although not part of our initial protocol, we performed a secondary analysis on the effect of spinal deafferentation on the functional connectivity in relation to the insula using a similar approach as presented for the thalamus network on the complete left and right insula (as a seed region). We observed that deafferentation changed connectivities between the insula and several brain areas including the ACC, frontal cortex and hippocampus (increased connectivity), cerebellum, occipital cortex, and brainstem (decreased connectivity) (Fig. 7 and Table 3). Interestingly, connectivity changes did increase when subjects had greater pain scores although the effect size was not as large as observed for the thalamus networks (data not shown). These data indicate that apart from an effect on pain intensity, deafferentation changes the pain affect and possibly also interoceptive sensations via changes in functional connectivities in the mentioned insula networks. Since the insula is topographically organized, further studies are needed to assess the deafferentation effect on networks relative to specific insula subregions.
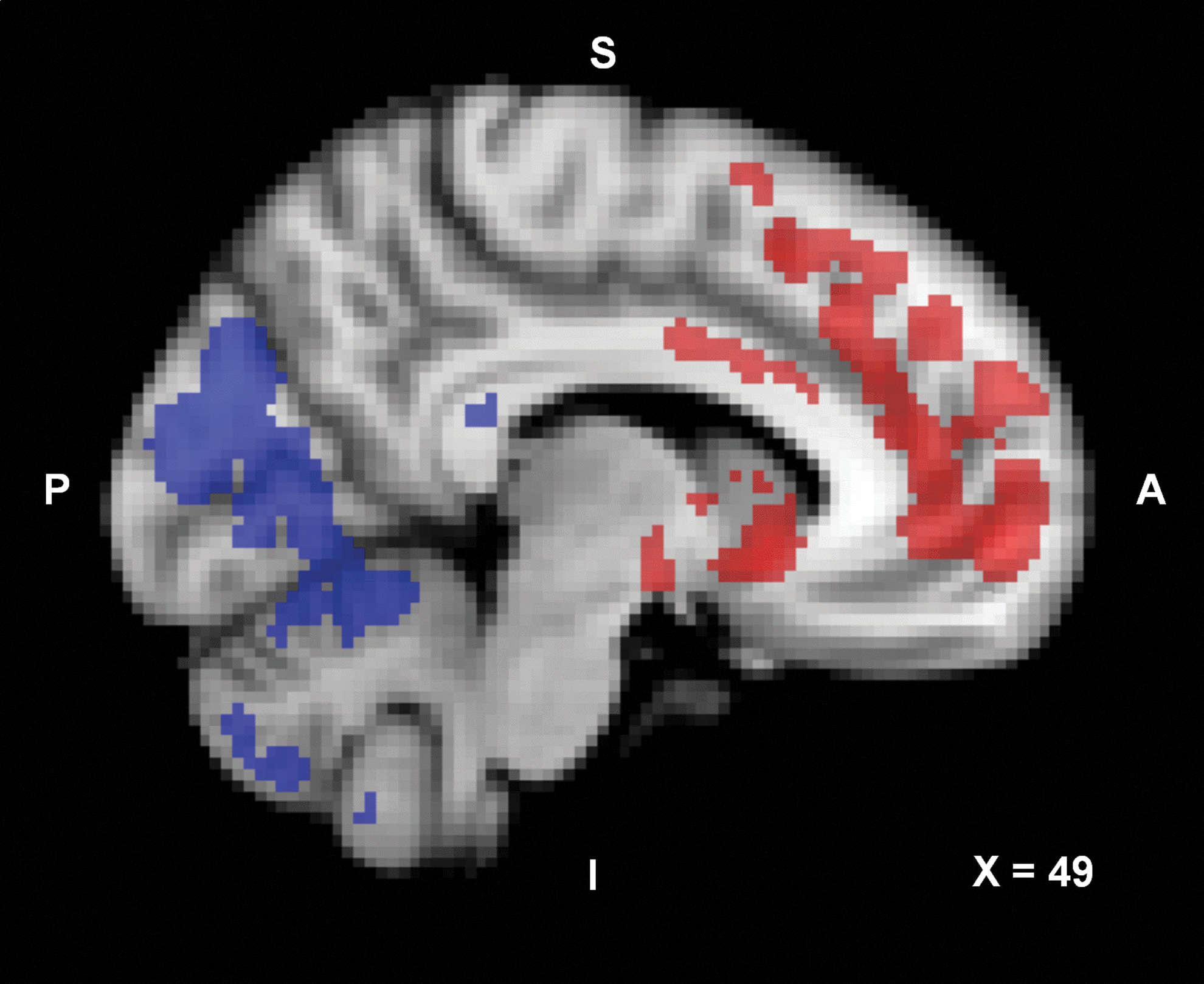
Statistical connectivity map (p<0.05; cluster corrected) of the increase (red) and decrease (blue) in resting-state network connectivity induced by spinal anesthesia in relation to the insula. Color images available online at
Effect of Spinal Anesthesia on Functional Connectivity in Relation to the Insula (Cluster p-Value <0.05)
Voxel dimension is 2×2×2 mm (voxel volume 0.008 mL);
Conclusions
Deafferentation from spinal anesthesia is associated with connectivity changes in the brain involving both cortical and subcortical areas. Furthermore, spinal anesthesia enhanced pain sensitivity that was correlated to enhanced connectivity patterns of the thalamus, ACC, and insula, areas associated with endogenous modulation of pain and the sensory and affective dimensions of pain perception.
Footnotes
Author Disclosure statement
No competing financial interests exist.