
Abstract
It is known that the default mode network (DMN) may be modulated by a cognitive task and by performance level. Changes in the DMN have been examined by investigating resting-state activation levels, but there have been very few studies examining the modulation of effective connectivity of the DMN during a task in healthy older subjects. In this study, the authors examined how effective connectivity changed in the DMN between rest and during a memory task. The authors also investigated whether there was any relationship between effective connectivity modulation in the DMN and memory performance, to establish whether variations in cognitive performance are related to neural network effective connectivity, either at rest or during task performance. Twenty-eight healthy older participants underwent a resting-state functional magnetic resonance imaging scan and an emotional face–name encoding task. Effective connectivity analyses were performed on the DMN to examine the effective connectivity modulation in these two different conditions. During the resting state, there was strong self-influence in the regions of the DMN, while the main regions with statistically significant cross-regional effective connectivity were the posterior cingulate cortex (PCC) and the hippocampus (HP). During the memory task, the self-influence effective connectivites remained statistically significant across the DMN, and there were statistically significant effective connectivites from the PCC, HP, amygdala (AM), and parahippocampal region to other DMN regions. The authors found that effective connectivities from PCC, HP, and AM (in both resting state and during task) were linearly correlated to memory performance. The results suggest that superior memory ability in this older cohort was associated with effective connectity both at rest and during the memory task of three DMN regions, which are also known to be important for memory fuction.
Introduction
Effective connectivity is defined as the influence of one brain region on another distant brain region (Friston et al., 1997, 2013) and has been studied extensively using functional magnetic resonance imaging (fMRI) in recent years. In particular, the focus has been on investigating the strength and changes in effective connectivity during performance of a task (Fairhall and Ishai, 2007; Pessoa and Adolphs, 2011; Vuilleumier et al., 2004). Currently, there is intense interest in quantifying effective connectivity during both resting-state and task-related activation, as well as the association, if any, between these. For example, in a pilot study that investigated the effects of therapy in stroke patients with resting-state fMRI measured pre- and post-therapy, it was found that the effective connectivity within motor networks increased in patients who showed a behavioral improvement (James et al., 2009). There is also evidence that connectivity at rest can predict task-related connectivity (Hamilton et al., 2011). In this study of resting-state connectivity in major depression patients, it was found that aberrant effective connectivity led to increased excitatory activity among limbic structures and increased inhibitory activity in the medial prefrontal cortex in patients compared to healthy subjects (Hamilton et al., 2011). In the current study, the authors were interested in investigating how accurate these predictions were for elderly subjects, and if they could quantify how brain regions are dynamically integrated in response to different tasks.
The default mode network (DMN) consists of a number of medial and lateral brain regions, including the posterior cingulate cortex (PCC), medial PFC, temporal cortex, and hippocampus, and is most active when a person is resting or mind-wandering but deactivates in response to goal-directed tasks (Raichle et al., 2001). The DMN appears to play an important role in cognitive function, particularly in later life. In a study of amnestic mild cognitive impairment (aMCI) (Dunn et al., 2014), which confers an increased risk of developing Alzheimer's disease (AD) (Stephan et al., 2010), aMCI participants showed reduced connectivity between two key DMN regions—the hippocampus and PCC—compared to individuals with nonamnestic MCI, and this disconnect in the DMN was related to impaired episodic memory performance. A combined fMRI-PET study found reduced deactivation of medial and posterior regions of the DMN during a memory encoding task in a cohort of cognitively intact, but high amyloid burden individuals (Sperling et al., 2009). Another recent study used dynamic causal modeling (DCM) to investigate the modulation in effective connectivity within the DMN during the performance of a gender judgment task (Li et al., 2012). It was found that during the task there was increased effective connectivity between regions as measured with DCM compared to the resting-state scan. At the same time, there was decreased self-inhibition connnectivities in the DMN during the task compared to resting state.
Effective connectivity therefore allows the investigation of the interactions among brain regions that comprise a network. For example, one could investigate interactions among the regions of the DMN under resting state and with an exogenous input (Li et al., 2012). This type of analysis can not only be used to investigate how the interactions are altered by an input into the network but also allows one to build mechanistic-based insights of how the connectivity in the network is altered. One issue that has not been investigated is how effective connectivity changes among the regions of the DMN between resting state and during a task-driven period. There has been considerable work examining these issues from the perspective of network activation and deactivation (among many see for example (Greicius et al., 2003; Greicius and Menon, 2004; Laird et al., 2013; Mennes et al., 2010), but from the perspective of effective connectivity, the work until now has been more limited.
There is a long history of studying connectivity loss in aging and AD subjects. Generally, it has been found that there is decreased functional and structural connectivity in these subjects (Clerx et al., 2012; Damoiseaux et al., 2009, 2012; Rose et al., 2000). However, only a few studies have been carried out to investigate effective connectivity (causality) in aging and AD. Causality analysis methods are very useful to investigate long-distance network integration in the human brain. However, due to the nature of the blood oxygenation level-dependent (BOLD) response, effective connectivity results from this method are modulated by different input stimuli. Therefore, it is necessary to conduct experiments to study effective connectivity changes under different conditions, that is, using different stimuli as brain system inputs. The purpose of this study was to achieve this goal. The authors studied dynamic brain network modulation effects using different stimuli and investigated the neuronal dynamic properties in aging subjects. The biological implication is that older adults and possible AD, there is decreased neuronal dynamics, which may contribute to reduced functional and structural connectivity. The loss of neuronal population dynamics could happen before function and structural changes are detected using neuroimaging; consequently, this could be a new biomarker for early AD diagnoses or for the detection of age-related changes in neural network dynamics. This study goes one step further to understanding the dynamics properties of functional brain characteristics in the aged brain.
Thus, in the current study, the authors were interested in investigating the modulations in effective connectivity of the DMN during a memory task. To investigate effective connectivity changes in response to different stimuli, the authors included two fMRI scans in the experimental design—a resting-state fMRI scan and an emotional face–name encoding task. The test group in this study comprised older healthy participants and the authors chose a task (emotional memory) that is relatively less vulnerable to age-related decline, so that the investigation of effective connectivity was not confounded by age-related memory loss. There have been many studies reporting relative preservation of emotional processing in healthy aging (Grady et al., 2001; Kehoe et al., 2013; Kensinger et al., 2002; Kensinger, 2008; Mather, 2012; Rosenbaum et al., 2010). Generally, it has been found that there is decreased functional and structural connectivity in older subjects (Clerx et al., 2012; Damoiseaux et al., 2009, 2012; Rose et al., 2000), however, the dynamic nature of networks has been less well studied in aging. This study goes one step further to understanding the dynamics properties of functional brain characteristics in the aged brain. This may allow for the investigating of whether task performance such as memory may be predicted from the effective connectivity in a network (Hampstead et al., 2011).
Based on these two functional scans, the authors set out to examine the effective connectivity in the DMN under resting-state and memory task conditions. The authors wished to address two main questions related to brain network modulation during the resting state and memory function. First, what are the causal relationships between different regions of the DMN during the resting-state task and in response to a face–name encoding task in elderly healthy subjects? And second, is there a significant correlation between network/regional causality and performance on the face–name memory task? Addressing these questions may help us to understand brain regional causality in the DMN, to understand the relationship in the DMN between resting-state effective connectivity and effective connectivity during a task, and finally to assess whether differences in effective connectivity during these experimental conditions can predict behavioral performance on a cognitive task.
Materials and Methods
Subjects
Twenty eight elderly, healthy right-handed subjects (14 males; age=69.8±6.4 years; age range=57–77 years) participated in the study. The study was performed with the informed consent of the subjects, had full ethical approval from the St. James and the Adelaide and Meath Hospitals (Dublin, Ireland), incorporating the National Children's Hospital Research Ethics Committee and followed the tenets of the Declaration of Helsinki. The participants underwent a brief neuropsychological examination to rule out cognitive impairment (The Consortium to Establish a Registry of Alzheimer's Diease [CERAD] battery) (Morris et al., 1988).
MRI data collection
A Philips Intera Achieva 3.0 T MR system (Best, The Netherlands) was used to acquire the MRI data. A T1-weighted sequence was used to collect a high-resolution, three-dimensional anatomical image (isotropic voxel at 0.9 mm) for structural localization and visualization. The structural scan was followed by the resting-state scan. During the resting-state scan, the BOLD signal changes were measured using a T2*-weighted echo-planar imaging sequence with TR=2000 msec and TE=30 msec. Each volume of data covered the entire brain with 37 slices, and the slices were acquired in an interleaved sequence from the inferior to superior direction, with voxel dimensions of 3×3×3.55 mm and the image size was 80×80 voxels. In total, 210 volumes were acquired during the resting fMRI. During the resting fMRI scan, the subjects were instructed to remain relaxed with eyes open and to look at a fixation cross on a screen, without thinking of anything in particular. This scan was always acquired before the memory task so that the resting-state activation was not influenced by the task.
Experimental tasks: face–name encoding task
The participants performed a face–name encoding task following the resting-state scan. During this task, the participants viewed a series of 27 emotional faces (Erwin et al., 1992) with a name presented underneath each one. This task was an implicit memory task, in that the participants later completed two surprise memory tasks to test their retention of both the faces and the face–name pairs, however, at the time of encoding, they were not explicitly asked to remember the face–name pairs. Rather, the participants were instructed to judge whether the names matched or suited the faces. It was explained that this was a subjective decision, with no right or wrong answer. The participants responded yes or no by pressing a button on a MR-compatible response pad held in their right or left hand, respectively, using the index finger of either hand. Each face–name combination was presented for 4 sec and was shown twice during the run. The faces were positive, negative, or neutral in valence and there were equal numbers of valence types as well as gender. The presentation of the face–name pairs was grouped according to the emotional valence of the faces. In each instance, a group of either two, three, or four faces of one valence type was presented randomly using an event-related paradigm, subsequently, there was a delay during which a white crosshair was presented (control condition). The duration of the white cross was varied according to the duration of the face stimulus. For instance, if a single face was presented for 4 sec the subsequent white cross was also shown for 4 sec and then the next block of faces began. (See Fig. 1a for a schema of the encoding task). The stimuli were delivered using Presentation v.16.1 (Neurobehavioral Systems, Albany, CA).
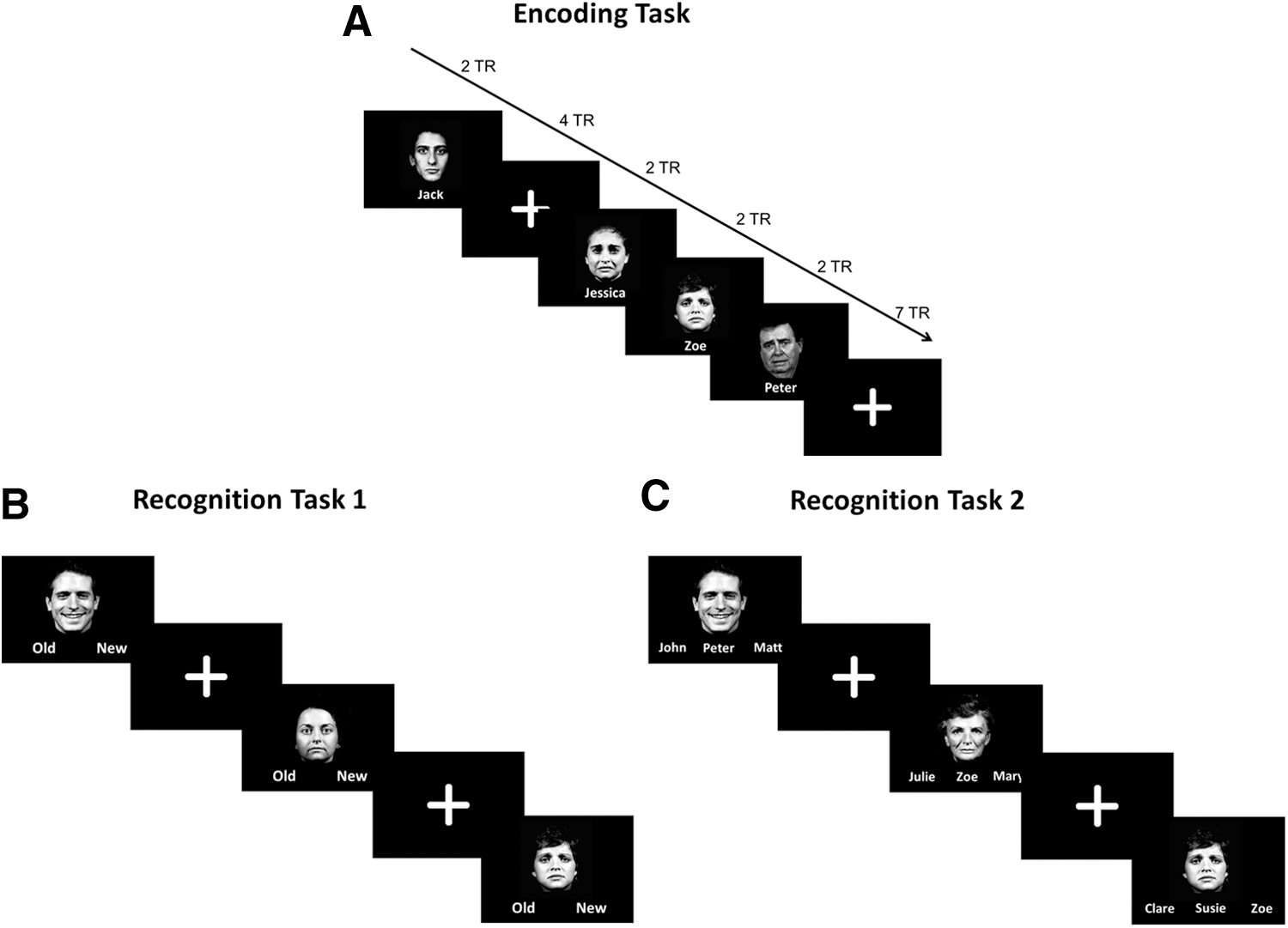
Schema of the task design.
Experimental tasks: recognition task 1
Approximately 15 min following the MRI scan, the participants performed two short computer-based recognition tasks. The first memory task was designed to test the participants' recognition memory for the faces presented during the encoding phase. During this task, the faces presented in the scanner were shown along with nine new faces matched for gender and valence (foils). The faces were presented one at a time in the center of a black background, with the words Old and New printed underneath (See Fig. 1b for a schema of recognition task 1). The participants were asked to select Old if they recognized the face as one which they saw before in the scanner, and New if they did not recognise the face. They made their selection by pressing a button on the left or right side of a keyboard to correspond to the position of the word on the screen. Each face was presented for 4 sec and followed by white crosshairs for 2 sec. The order of the faces was randomized relative to their order during the encoding phase. Before the task began, the participants completed a short practice run of five trials to familiarise them with the task.
Experimental tasks: recognition task 2
The second memory task was designed to test the participants' memory of the face–name pairs from the encoding phase. The emotional faces were presented one at a time on a black background with three names underneath (see Fig. 1c for a schema of recognition task 2). One of the names was the correct name; one name was a name that had been paired with a different face (distractor; incorrect name), while the third name was a new name (foil; incorrect name). The participants responded by pressing a button on the left, middle, or right side of a keyboard to correspond with the relative position of the name on the screen. The stimuli were presented for 5 sec and followed by an intertrial interval of 5 sec. This longer trial length was to facilitate performance of this task as it was quite challenging. Before the task began, the participants completed a short practice run of five trials.
Linear system identification method for studying effective connectivity
A linear system identification method (LSIM), which is a special case of the nonlinear system identification method (NSIM) (Li et al., 2010, 2011), was employed to study causality between regions of the DMN. The authors used the LSIM method because they wanted to take a conservative approach to the analysis. This method is based on the assumption that the authors only know the brain system inputs and outputs, that is, the experimental design is the system input and the system output is the fMRI (BOLD) signal. Therefore, the authors can describe the brain system using only system inputs and system outputs. The authors adopted a linear parameter approach to map the relationship between brain system inputs and outputs. This input–output relationship consists of three parts, that is, first, the linear basis obtained from the fMRI response of the connected regions, second, the experimental design or the system input, and finally, the drift terms, that is, for a two-connection network with first order of AR, the authors have (Li et al., 2010):
Once the system parameters/coefficients a 1, b 1, a 2, and b 2 in equations (1) and (2) have been identified using a least squares method, the authors can test the causality relationship between different regions using a statistical inference method. In the study, the authors used the zero order of polynomials, that is, constant shift terms a 0 and b 0 in equations (1) and (2) to model the mean to exclude its effect for the connectivity analysis. After the model was defined and the parameters calculated, the authors applied a T-test statistical method for causality inference between different connected regions, that is, tested the significance of the parameters for y 1(t − 1) and y 2(t − 1) in equations (1) and (2) for the causality analysis.
For the resting fMRI, the authors assume that the brain activity measured with the BOLD signal can be regarded as a special case of the previous model where no specific task or brain system external input is involved. Therefore, LSIM can be applied without including the system input to study the effective connectivity, that is, u(t)=0 in equations (1) and (2). When using this method, in the first step, the authors define the region of interest (ROI), that is, the structure of the network and the regional time series y 1(t), y 2(t) in equations (1) and (2) for the effective connectivity study. This can be achieved by applying a singular value decomposition to extract the representative time series for each region (Li et al., 2013). Then, LSIM can be applied based on the representative, regional, fMRI response time series for the effective connectivity study.
ROIs for effective connectivity study
The authors were interested in studying the DMN, which they defined as consisting of six ROIs as shown in Figure 2. The ROIs were located in the PCC, medial superior frontal gyrus, hippocampus, parahippocampal gyrus, amygdala, and superior parietal lobule (Raichle et al., 2001). The ROIs were based on the automated anatomical labeling template (Tzourio-Mazoyer et al., 2002), and regional fMRI time series were extracted from these masks. The authors were interested in how information flows (causality influence) in the medial temporal regions, which include the hippocampus, parahippocampal gyrus, and amygdala. Furthermore, the authors assumed that the information flow between these regions was bidirectional, resulting in the network structure as shown in Figure 2.
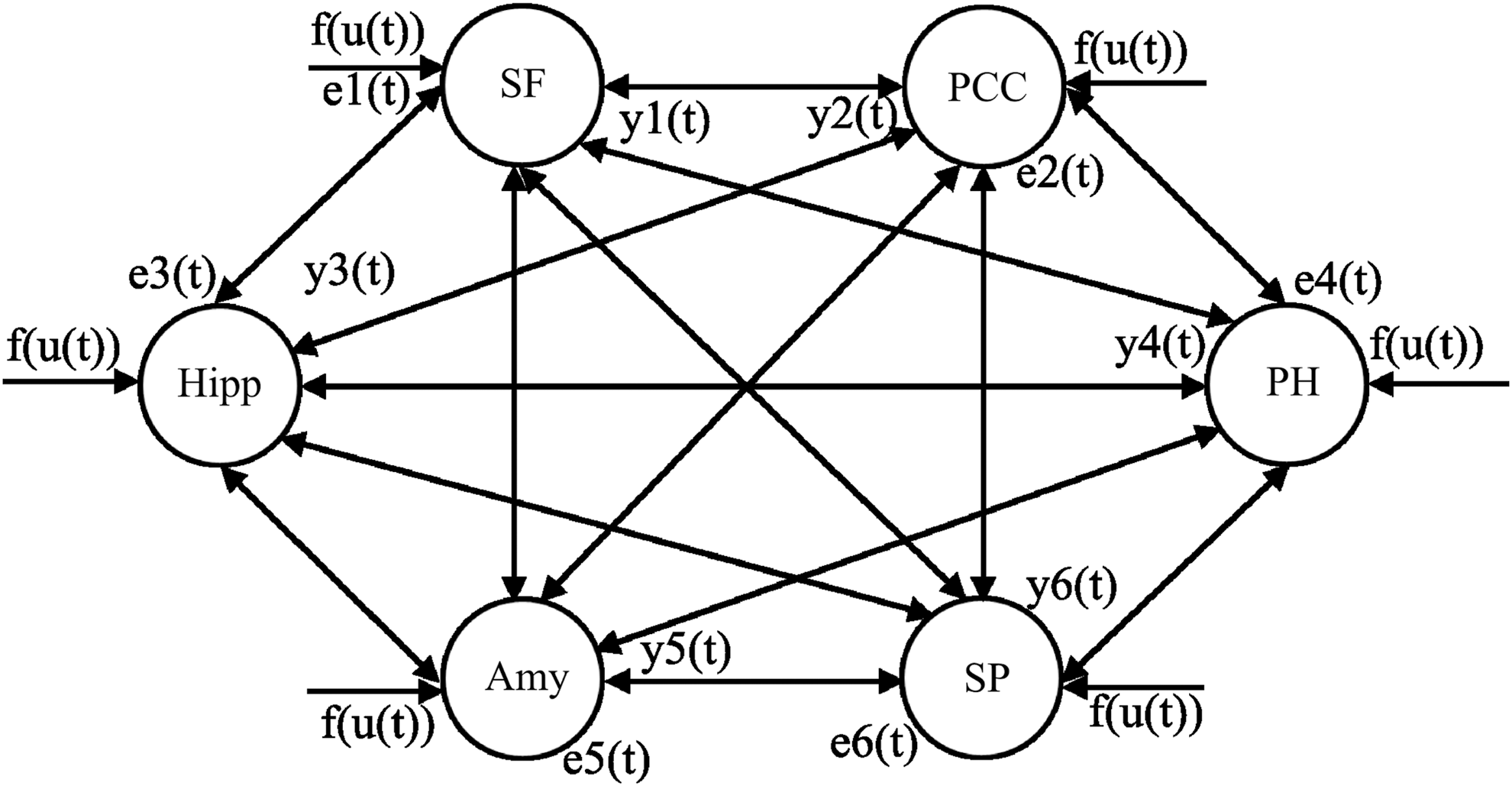
The DMN brain network. Amy, amygdala; DMN, default mode network; Hipp, hippocampus; PCC, posterior cingulate cortex; PH, parahippocampal gyrus; SF, medial superior frontal gyrus; SP, superior parietal lobule; f (u(t))=u(t) is the system input.
Results
Effective connectivity in the DMN during rest and task
In the resting state (Fig. 3A), there was a strong self-AR influence in the PCC (t-value=6.46), SF (t-value=4.58), HP (t-value=3.49), AM (t-value=5.65), and SP (t-value=10.30). Across the various regions of the DMN, the regions that had the largest number of statistically significant effective connectivity to other regions were the PCC (to all regions except SP), PH (negative effective connectivity to HP and AM), and AM (to HP and PH), while the SF had a negative effectivity connectivity to the PH. The PCC and HP had the strongest influence on the other regions of the DMN.

Modulation effect of the DMN. Matrix of t-values showing the effective connectivity in the DMN during
During performance of the task, the effective connectivity of the DMN was examined (Fig. 3B) and it was found that the self-AR effective connectivity was statistically significant in the same regions as during rest. The cross-regional effectivity connectivity had positive effective connectivities from the PCC to the HP (t-value=2.80), PH (t-value=3.81), and AM (t-value=2.36); the HP had positive effective connectivity to the SP (t-value=2.12) and AM (t-value=2.37); and the PH had negative effective connectivities to the HP (t-value=−2.57), AM (t-value=−3.54), and SP (t-value=−2.42).
Behavioral results
Memory performance results: recognition task 1
On recognition task 1 after the MRI scan, the participants correctly recognized 67.33%±15.59% of the faces from the scanning session, while they forgot 31.75%±11.60% of the faces. Nine foils (faces not seen in the encoding phase) were also presented and, of these, the participants on an average incorrectly classified 39.20%±11.62% as ones they had seen before, but correctly classified 61.31%±11.60% as new faces. This relatively high rate of false positives accords with many other previous aging studies, which have reported similar rates of false memory in older adults (e.g., Bender et al., 2010; Duarte et al., 2010; McCabe et al., 2009; Remy et al., 2008). Indeed, in a review of aging studies, McCabe et al. (2009) concluded that age-related increases in false hits are as reliably reported as age-related decreases in correct hits. Furthermore, the false memory effect may be particularly strong for positive emotional stimuli (e.g., Fernandes et al., 2008; Piguet et al., 2008; Werheid et al., 2010), which may induce a false sense of familiarity. Thus, to control for the confounding effect of false memory in the current study, performance on task 1 was quantified as the number of correctly recognized faces minus the number of new faces incorrectly rated as remembered (false positives) (Algarabel et al., 2012), that is, (% Old Correct) – (% New Incorrect). The average adjusted memory score of the participants was 28.12%±12.93% and this was used for the correlations with the fMRI results.
Memory performance results: recognition task 2
On recognition task 2, the participants on average selected the correct name for faces they had seen in the scanner on 55.69%±12.24% of the trials. On an average, they chose the incorrect name on 42.86%±12.11% of trials and there were 1.32%±5.72% missed trials (i.e., no response).
Association between effective connectivity and memory performance
The authors calculated the linear correlation coefficient between the participants' behavioral performance on the recognition memory tasks (postscanning) and the effective connectivity of the DMN during the resting state and memory task. The authors found three statistically significant linear correlations between memory performance on the face recognition task and the DMN effective connectivity during the task (r 26>0.37, p<0.05) (Table 1). For completeness, the authors also show the corresponding linear correlation between performance and effective connectivity during the resting state in the same regions.
The Regions of the Default Mode Network Where the Effective Connectivity During the Activation Task Was Statistically Significantly Linearly Correlated (p<0.05) with Performance in the Forced-Choice Face Recognition Task
In addition, the corresponding linear correlation coefficients between the regions during rest state and memory performance are shown.
CC, correlation coefficients; PCC, posterior cingulate cortex; PH, parahippocampal gyrus; SF, superior frontal gyrus; SP, superior parietal lobule.
For the face–name recognition task, the authors found five statistically significant linear correlations between the face–name recognition performance and effective connectivity in the DMN during the performance of the task (r 26>0.37, p<0.05) (Table 2). Again, the authors also show the corresponding linear correlations between effective connectivity and memory performance during the resting state.
The Regions of the Default Mode Network Where the Effective Connectivity During the Memory Task Was Statistically Significantly (p<0.05) Linearly Correlated with Performance in the Face–Name Recognition Task
In addition, the corresponding linear correlation coefficient between effective connectivity during rest and behavioral performance is shown.
AM, amygdala; HP, hippocampus.
The authors also found a statistically significant linear correlation between effective connectivity during the resting state and performance on recognition task 1 from the SF to the PCC (r=−0.3752); the corresponding correlation during the task was r=−0.3453. In addition, the authors found a significant correlation between recognition task 2 and resting-state effective connectivity, as shown in Table 3.
The Default Mode Network Regions Where the Effective Connectivity During the Resting State Was Statistically Significantly (p<0.05) Linearly Correlated with Performance in the Face–Name Recognition Task
In addition, the corresponding linear correlation coefficient between effective connectivity during task activation and memory performance is shown.
Discussion
In the current study, the authors investigated effective connectivity in the DMN under two different conditions (system inputs) in healthy older adults—at rest and during a memory encoding task. In this way, the authors examined the DMN and its modulation due to changes between the resting state and a cognitive task. One of the main findings of the study was the strong self-influence in the DMN regions both during the resting state and during the memory task. In addition, the cross-regional effective connectivities were strongest (number of statistically significant effective connectivities) with the PCC and hippocampus both during rest and the memory task. The strong self-influence across the network may indicate that one of the mechanisms driving the DMN is internal within each region of the DMN and may not depend on one central region or subgroup of the DMN. In addition, the cross-regional effective connectivites were centered on the PCC and hippocampus, two regions that have previously been defined as hubs of the brain (Thomas Yeo et al., 2011; van den Heuvel and Sporns, 2011). These regions have been implicated in memory function and in age-related cognitive decline as well as in AD.
Of note is that the effective connectivites between regions of the DMN had linear associations with performance in the emotional memory task—in particular the hippocampus, amygdale, and PCC—regions that have been shown to be involved in successful encoding. During the resting state, the effective connectivites from the amygdala to PCC, SF, and HP were linearly correlated with the face–name recognition performance level. Given the emotional content of the faces, it is not surprising that the amygdala is implicated in the performance level, but it was unexpected to see a negative correlation between the amygdala and the other regions. The effective connectivites in the DMN during the task were strongly associated with the hippocampus and amygdala (for face–name recognition) and PCC, SP, and SF for the forced-choice face recognition task. The strong involvement of the hippocampus and amygdala is consistent with the literature showing hippocampus activation in associate memory tasks (Celone et al., 2006; Killgore et al., 2000; Montaldi et al., 1998), and the authors extent these findings in terms of effective connectivity within the DMN.
Effective connectivity refers explicitly to the influence that one neural system exerts over another, either at a synaptic or population level. Effective connectivity is dynamic (activity dependent) and depends on a model of interactions or coupling (Friston, 2011b). Effective connectivity discounts other influences, therefore, this study has shown the influence that regions of the DMN have on each other and can be solely attributed to regions of the DMN. The study shows how these influences differ between the resting state and a cognitive task in healthy older subjects. The DMN is often considered a resting-state network, but this study clearly shows statistically significant effective connectivity of multiple regions in the DMN during a cognitive task. This study lends further support to the idea that the DMN plays an important role in memory function.
This method provides a new approach to evaluate the dynamic characteristics of brain networks, not only the networks that are recruited during a task but also the changes that occur in the DMN. The methods could be applied to investigate differences in the resting state between populations, for example, offering a new way of quantifying connectivity changes in neural networks associated with particular disease states. It could potentially be developed as a novel neuroimaging biomarker for disorders that are known to involve marked changes in the DMN, such as AD (Greicius et al., 2004; Mevel et al., 2011).
The DMN has been studied extensively using functional connectivity and independent component analysis (Greicius et al., 2004). These two approaches allow one to define which regions are linearly correlated in time, but do not quantify the influence of one region on the other, for example, by using effective connectivity measures. In the resting-state, fMRI effective connectivity has received relatively little attention. In the current study, the authors calculated the effective connectivity among regions of a network, where they examined all possible connections among the regions, to find the effective connectivity measures that were statistically significant. This approach provides estimates of information flow among regions of a network and may help us in understanding the nature of the resting state and the influence it may have on cognition.
Behavioral data correlation with effective connectivity
The authors found four significant linear correlations between performance on the memory tasks and the effective connectivity obtained from the DMN when the participants viewed the emotional face–name stimuli, while they only found one significant correlation using the effective connectivity obtained during the resting-state fMRI. This suggests that the effective connectivity in the DMN during the activation task is associated with memory performance of the healthy elderly participants. Similarly, a recent study (Sala-Llonch et al., 2012) found that memory performance can be predicted from resting-state connectivity in the period before performance of a working memory task.
Finally, the authors employed a newly developed analysis method, that is, LSIM (Li et al., 2010, 2011) for studying the effective connectivity. This method overcomes the limitations of conventional model-based methods that do not take the system input into account for the study of effective connectivity. Although the NSIM method can be used for studying effective connectivity, to be more conservative for the analysis, the authors only included linear terms and the first order of AR in the model for their fMRI effective connectivity studies. There is still an ongoing debate around using different methods, such as the Granger causality model (GCM), the dynamic causality model, and the moving average model for effective connectivity studies (Friston, 2011a). The method the authors applied, a simple form of the LSIM method to study effective connectivity, is most similar to GCM.
In this study, the authors have shown that the effective connectivity in the DMN is associated with cognitive function in helathy subjects. This study adds to the growing literature on the behavioral and cognitive aspects of the DMN.
Footnotes
Acknowledgments
This study was supported under the Centre of Excellence in Intelligent Systems (CoEIS) project, funded by InvestNI and the Integrated Development Fund, through ILEX. EGK was supported by the European Regional Development Fund through the Ireland Wales Programme 2007–13, and ALWB by the European Regional Development Fund through the Ireland Wales Programme 2007–13, and the Science Foundation Ireland (SFI) Stokes Programme.
Author Disclosure Statement
No competing financial interests exist.