
Abstract
Spontaneous brain activity, that is, activity in the absence of controlled stimulus input or an explicit active task, is topologically organized in multiple functional networks (FNs) maintaining a high degree of coherence. These “resting state networks” are constrained by the underlying anatomical connectivity between brain areas. They are also influenced by the history of task-related activation. The precise rules that link plastic changes and ongoing dynamics of resting-state functional connectivity (rs-FC) remain unclear. Using the framework of the open source neuroinformatics platform “The Virtual Brain,” we identify potential computational mechanisms that alter the dynamical landscape, leading to reconfigurations of FNs. Using a spiking neuron model, we first demonstrate that network activity in the absence of plasticity is characterized by irregular oscillations between low-amplitude asynchronous states and high-amplitude synchronous states. We then demonstrate the capability of spike-timing-dependent plasticity (STDP) combined with intrinsic alpha (8–12 Hz) oscillations to efficiently influence learning. Further, we show how alpha-state-dependent STDP alters the local area dynamics from an irregular to a highly periodic alpha-like state. This is an important finding, as the cortical input from the thalamus is at the rate of alpha. We demonstrate how resulting rhythmic cortical output in this frequency range acts as a neuronal tuner and, hence, leads to synchronization or de-synchronization between brain areas. Finally, we demonstrate that locally restricted structural connectivity changes influence local as well as global dynamics and lead to altered rs-FC.
Introduction
Structure and function
One of the major open questions in neuroscience is the relationship between structure and function. Several spatiotemporal features of resting-state functional connectivity (rs-FC) have been captured by existing modeling approaches (Deco et al., 2011, 2013b). A systematic framework for relating structural connectivity (SC) and functional connectivity (FC) has been recently provided by The Virtual Brain (TVB), a simulation platform of large-scale brain activity (Ritter et al., 2013; Sanz et al., 2013). There is a lack of mechanistic understanding of changes of network states due to learning/plasticity (Sigala et al., 2014). This article explores how to describe learning and plasticity-related changes of FC with biophysically plausible brain models. With learning and plasticity, we refer to training/exposure-related changes of SC at all spatial brain scales and the resulting functional consequences. Computational insights gained due to structural modifications will add to our understanding of the structure-function relationship provided by previous computational studies (Alstott et al., 2009; Cabral et al., 2012a, 2012b). A key ingredient of our model is the structural brain network defined by empirically derived long-range brain connectivity between regions, resulting in biologically plausible conduction delays. We use a subset of cortical regions represented as nodes in the large-scale brain network. The nodes are dynamical units built of highly interconnected excitatory and inhibitory neurons. The interaction between those regions in the network needs to be uncovered to understand the origin of correlated activity fluctuations in the brain during resting-state and task conditions. Here, we evaluate existing modeling approaches and advance them by incorporating state-dependent local plasticity mechanisms. Specifically, we explore how subcortical input to a cortical region shapes the local SC at a microscopic scale. Subsequently, we investigate the interaction with other cortical regions connected via the realistic anatomical connectivity matrix. Using computational and analytical methods, we show how input oscillations in the alpha (8–12 Hz) frequency range along with plasticity parameters (plasticity time constant, input oscillation frequency) critically influence transition from asynchronous to synchronous state in a local neuronal population. Local oscillations, in turn, reshape evolution of covariance on a longer time scale across a subset of anatomically connected cortical regions. We evaluate theoretical scenarios, how brain states in terms of slow power fluctuation of the local field potential's (LFP) alpha frequency band influence local plasticity. Restructuring of cortical node level synaptic weights due to spike-timing-dependent plasticity (STDP) results in learning phases of firing. In other words, the neurons learn to elicit output population spikes at specific phases of input background oscillations. We find that spatiotemporal low frequency fluctuations (<0.1 Hz) of the simulated signal—similar to those observed in blood oxygenation level-dependent (BOLD) functional magnetic resonance imaging (fMRI) signals during rest—are significantly shaped not only by given anatomical connectivity but also by ongoing synaptic plasticity within cortical regions. Combining brain states and plasticity provides a general theoretical framework for alterations of rs-FC, which reflects ongoing network dynamics and, hence, predicts the outcome of behavioral performance.
Plasticity shapes rs-FC
Typical large-scale interactions between brain areas are concerned with neural assemblies that are farther apart in the brain (>1 cm) with transmission delays in the order of 8–10 msec spanning poly-synapses (Varela et al., 2001). Resting-state networks (RSN) arise from coordinated distributed functional clusters. They give rise to coherent neural activity as a signature of large-scale brain operations in the absence of an explicit task or stimuli (Fox and Raichle, 2007; Smith et al., 2009). RSNs resemble both task-related functional networks (FNs) (Biswal et al., 1995; Fox and Raichle, 2007; Vincent et al., 2007) and anatomical networks, that is, regions directly connected via anatomical pathways (Honey et al., 2009). Recent studies have shown that cortical activation can restructure resting-state activity after task performance (Lewis et al., 2009; Riedl et al., 2011; Tambini et al., 2010; Urner et al., 2013; Zhang et al., 2012). Finally, evidence suggests that perceptual learning modifies the large-scale functional covariance between networks engaged in a task (Cole et al., 2012; Lewis et al., 2009). In line with this, only 30 min of repetitive sensory stimulation (RSS) (Freyer et al., 2012a)—a paradigm known to induce neural plasticity and counteracting sensory decline due to neurological trauma or aging (Kalisch et al., 2008, 2010)—leads to significant changes of the time-lagged coherence in resting-state alpha rhythm within sensorimotor cortical areas (Freyer et al., 2012a). While learning alters resting-state activity, this activity also influences the ability to learn. A recent electroencephalography (EEG) study on perceptual learning showed that high resting-state alpha power contralateral to the stimulated finger is predictive for a greater behavioral improvement after RSS at an inter-subject level (Freyer et al., 2013). These findings provide evidence that the resting brain is not a passive sensory-motor analyzer driven by sensory inputs. Rather, it actively generates predictions about forthcoming sensory stimuli by maintaining a structured memory of past activations that finds expression in the ongoing brain states. Here, we explore the effects of the interplay of RSS-induced plasticity and ongoing brain states in the alpha frequency range as a mechanism that could provide explanation for the corresponding change in FC between somatosensory and motor cortex connected via realistic SC.
Computational modeling
To understand the mechanisms underlying the complex interaction at the neuronal population level during rest and under task-based conditions, brain imaging as well as computational models play complementary roles. Brain imaging monitors cognitive states in the whole brain network at a macroscopic scale. For example, in fMRI, a cortical voxel describes the population activity of a few million neurons. Computational models provide potential underlying mechanisms of interaction between neuronal populations that give rise to the observed spatiotemporal dynamics of BOLD or EEG activity. This is realized by representing functional clusters of neurons by mean field or neural mass models. Mean field or neural mass models attempt to model the dynamics of large (theoretically infinite) populations of neurons. Numerous recent resting-state modeling studies (Cabral et al., 2011; Deco et al., 2013a, 2013c) successfully reconstructed empirically observed RSNs between clusters, patterns of positive and negative correlations as described in many experimental studies by looking at the interplay between local network dynamics and large-scale structure of the brain. In principle, global FC can be altered within milliseconds by perturbation, while the large-scale SC matrix remains fixed or under natural conditions changes only over longer time periods through plasticity processes. However, on a microscopic scale, synaptic dynamics and connectivity are altered within milliseconds (Caporale and Dan, 2008). Emergent FC depends on the dynamics of individual nodes, which are shaped by the local and global neuronal context. In the framework of TVB, this is accounted for by allowing for inputs from multiple connected regions to individual sub-cortical or cortical regions. Key signatures of emergent network dynamics are neuronal oscillations that can be detected by non-invasive brain imaging methods such as EEG.
Overview of the study
In the first section, we discuss ongoing activity at rest and underlying dynamics using two types of models: a spiking model (Deco and Jirsa, 2012), a mean field firing rate model derived from the spiking model (Deco and Jirsa, 2012; Deco et al., 2014). All are capable of generating a similar bifurcation structure. The dynamic mean field (DMF) model and the spiking model are capable of generating a similar bifurcation structure as shown qualitatively in Figure 2D. This is relevant, as empirical work shows multistable behavior in resting-state brain dynamics (Freyer et al. 2009).
In the second section, we introduce time-dependent input from the thalamus to the cortical neuron model equipped with STDP. In the local brain area—that is somatosensory cortex in our model—synaptic plasticity results in learning to spike at input phases of oscillations in the feed-forward network between the thalamus and somatosensory cortex. Moreover, when STDP is combined with an oscillation in the external input current, STDP acts robustly to detect and learn repeating patterns. It has been previously demonstrated in single neurons that theoretically ongoing oscillations may facilitate information decoding using a principle known as phase-of-firing coding (PoFC) (Masquelier et al., 2009). Here, we extend this finding by implementing STDP and PoFC in a neuronal population model with anatomically derived cortical connectivity between brain regions. There, STDP combined with rhythmic input synchronizes the spike output of the local spiking network model. The local node activity on a longer time scale yields periodic firing patterns in the alpha band range that exhibit local (within node) synchrony.
Finally, we carry out simulations by connecting a subset of distributed cortical areas in both right and left hemispheres with synaptic plasticity implemented in the somatosensory cortical area. We use a small realistic SC matrix customized for our simulations comprising the thalamus, somatosensory S1, and motor M1 areas of both hemispheres. We investigate the organization of FC on a time scale of 60 sec. These simulations provide a candidate framework to explore global coherence between areas under structural modifications.
Ongoing Oscillatory Activity and Dynamical Landscape at Rest
RSNs have been interpreted as the “ground state” of cognitive architecture (Deco et al., 2013c). In Table 1, we summarize network models that address the underlying neurodynamical mechanisms of RSN features as observed in fMRI, EEG, and magnetencephalography (MEG) (Cabral et al., 2011; Deco and Jirsa, 2012; Deco et al., 2013c; Freyer et al., 2011, 2012b; Ghosh et al., 2008a,b; Honey et al., 2007, 2009; Ritter et al., 2013). We list those mean field models that exhibit either fixed-point attractor or oscillatory or chaotic dynamics. Figure 1 summarizes the basic ingredients required to build up a realistic large-scale model of the cortex as done in TVB [for details, see Ritter and coworkers (2013)]. The main ingredients are a large-scale SC matrix determining connections between brain areas in terms of their distance and strength and a choice of cortical node models; for example, oscillatory (Cabral et al., 2011; Ghosh et al., 2008a), chaotic (Honey et al., 2009), and non-oscillatory (Deco and Jirsa et al., 2012). Ghosh et al. (2008a) proposed a stochastic mechanism to underlie the resting-state fluctuations via a careful optimization of the operating point close to criticality, that is, at the edge of dynamic instability. This mechanism has been adopted in many of the network models (Deco and Jirsa, 2012; Deco et al., 2013c).
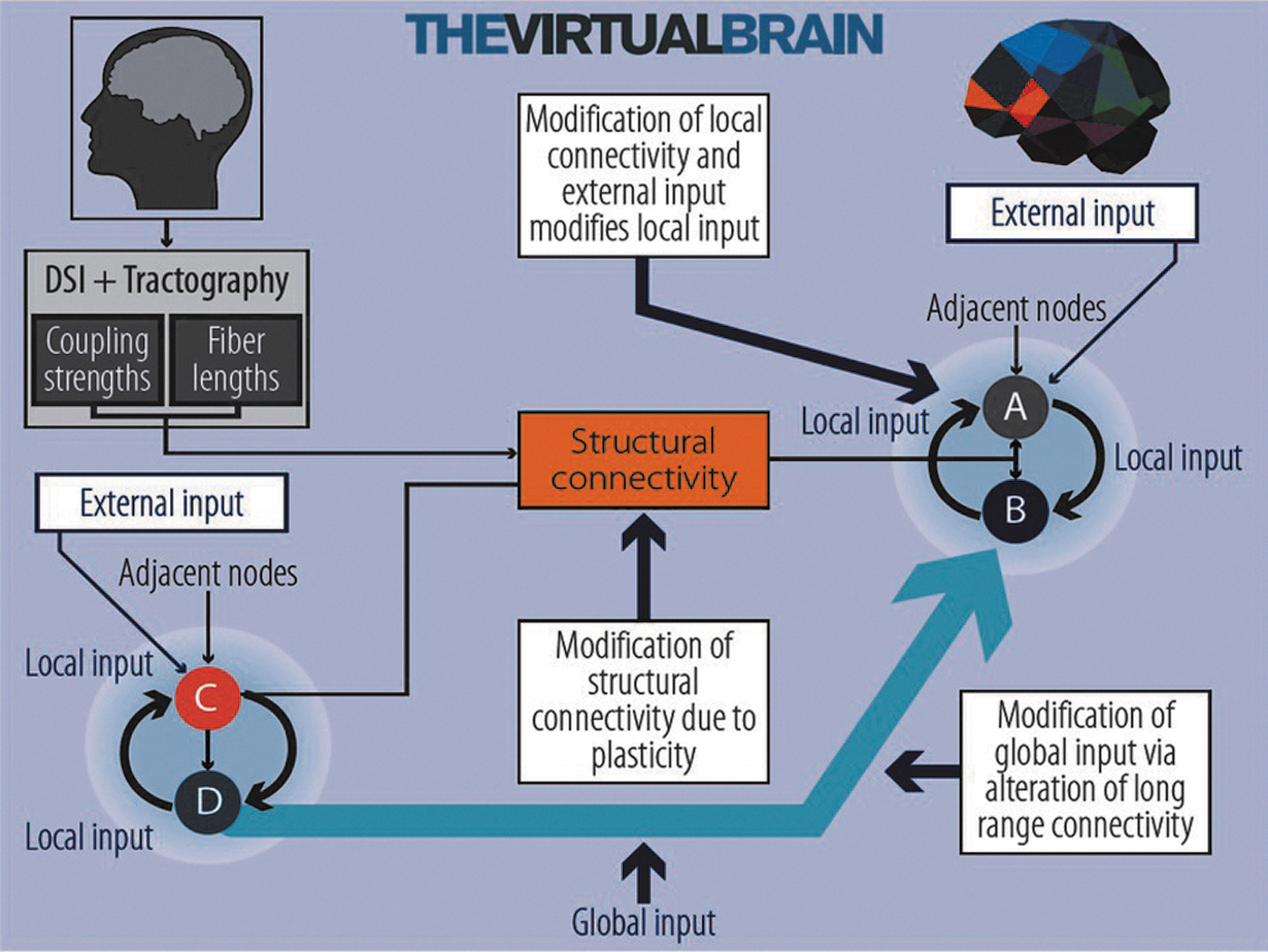
External input as well as local and global context shape ongoing brain dynamics. In the Virtual Brain (TVB) framework, we use informed structural connectivity (SC) (orange box) combined with local models described as nodes A, B, C, and D, respectively. TVB provides the choice of defining specific node dynamics using spiking attractor dynamics, firing rate dynamics, or oscillator models. The local models receive external time-varying input reflecting external stimuli, e.g. noisy input, external constant current input, local input from adjacent neural populations, and a global contextual input from distant populations. There are various ways to parametrically modulate the input received by an individual node; for example, by modifying SC, the local network connectivity (local scaling factor), long distance connectivity (global coupling scaling factor), and external input. Color images available online at
A Selection of Models Providing Important Insights into the Origin of Observed SpatioTemporal Pattern During Rest
Models capturing underlying neurodynamic mechanisms and features of empirical observations in neuroimaging research using fMRI, magnetencephalography and EEG. In all models, structural information is extracted from databases compiling different types of tracing studies; for example, in the macaque cortex, this information is provided by the CoCoMac neuroinformatics platform (Kotter, 2004) or by tractography using DTI data (Nooner et al.) and DSI (Hagmann et al., 2008).
BOLD, blood oxygenation level dependent; DSI, diffusion spectrum imaging; DTI, diffusion tensor imaging; EEG, electroencephalography; fMRI, functional magnetic resonance imaging; IF, integrate-and-fire; RSN, resting state networks.
Single brain area spiking model
We introduce a network of integrate-and-fire (IF) spiking neurons with excitatory (α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid [AMPA] receptors, N-methyl-D-aspartate [NMDA] receptors) and inhibitory populations with gamma-aminobutyric acid (GABA) receptor types. This spiking neuron model is similar to the one used in Brunel and Wang (2001) and Deco and Jirsa (2012). Each cortical region is modeled as a fully recurrently connected network comprising NE
excitatory neurons and NI
inhibitory neurons (Table 2). We describe the dynamical evolution of a single neuron's membrane potential V(t) that is driven by incoming total excitatory and inhibitory inputs of the same cortical area and the long-range excitatory input from directly connected brain areas. Hence, the time evolution of the membrane potential for an arbitrary neuron i obeys the following differential equation:
where gm
is the membrane leak conductance, Cm
is the membrane capacitance, and VL
is the resting potential; membrane time constant is defined as
Neurons, Connections, and Synaptic Parameters in Each Local Brain Area
Isyn is the synaptic current and comprises AMPA currents, NMDA currents, and GABA-A currents as defined next. The spike is transmitted to other neurons, and the membrane potential is instantaneously reset to Vreset and further maintained there until a refractory period is reached τref during which neurons are unable to produce further spikes.
Postsynaptic excitatory potentials are generated on the target neuron mediated via the conductance-based model specified by synaptic receptors, namely: glutamatergic AMPA external excitatory currents, AMPA, NMDA recurrent excitatory currents, and GABAergic recurrent inhibitory currents. The respective synaptic conduct gAMPA,ext
, gAMPA,rec
, gNMDA,rec
and gGABA,rec
, VE
, V1
are excitatory and inhibitory reversal potentials, respectively. Each one of the current excitatory feed-forward input currents, recurrent excitatory as well as inhibitory synaptic currents are described next:
Dimensionless parameters wij are the synaptic weights of recurrent all-to-all connectivity between excitatory and inhibitory neurons in each local brain area. By modifying these weights, one can either up- or down regulate the four local connectivity strength parameters (wEE , wEI , wIE , wII ). Unless otherwise specified, synaptic weights of the local recurrent connectivity are held at fixed values throughout our simulation of models. The recurrent self-excitation wEE =W +=1.4 and the remaining three synaptic weights are wEI =wIE =wII =1, respectively.
The fraction of open channels of neurons are given by the kinetics of the gating variables
NMDA channels are represented by a separate decay and rise-time kinetics. The sums over index p represent all the spikes emitted by presynaptic neuron j at times
In addition, all neurons in the network receive an AMPA-mediated external background current derived from a Poisson process of uncorrelated spikes with time-varying firing rate
where τn =30 msec, baseline firing rate=2.18 kHz (background). Local and global connectivity parameters are given in Table 2. Parameter values in Table 2 are modified from Brunel and Wang (2001)—where the network balance is shifted more toward an inhibitory regime—to incorporate local balance of excitatory and inhibitory synaptic currents as shown in Deco and Jirsa (2012), Albantakis and Deco (2011), and Deco and coworkers (2013c, 2014). Parameters for the single cortical area neuronal synapse model are summarized in Table 2. One of the key motivations for the choice of single area parameters as shown in Table 2 was to keep the uncoupled local area network to fire asynchronously at around 3–4 Hz, which closely resembles the biological range of spontaneous firing activity given in any isolated cortical region (Haider et al., 2006). Hence, this asynchronous spontaneous firing rate is maintained at about 3–4 Hz when isolated from the global network and in the absence of plasticity and brain state-dependent feed-forward drive. This type of local balance of excitation and inhibition in local cortical networks is sufficient to achieve oscillations in the desired frequency range, maximal synchrony and criticality in the resting-state dynamics (Poil et al., 2012). The balance (local excitation-inhibition ratio) impacts the spontaneous and evoked large-scale brain dynamics (Deco et al., 2014).
Single brain area represented by a DMF model
To model the RSN dynamics and prediction of empirical functional connectivity at the macroscopic level, Deco and coworkers (2013b, 2014) proposed a large-scale DMF model derived from the spiking model described earlier. As shown in Deco and associates (2014), the reduced DMF model consistently expresses the time evolution of the ensemble activity of the different excitatory and inhibitory neural populations building up the spiking network. In the DMF approach, each population's firing rate depends on the input currents into that population. The input currents, in turn, depend on the cortical pool population firing rates. The populations firing rate can be determined by a reduced system of coupled nonlinear differential equations that are driven by the respective input currents. The large-scale model connects these local sub-networks according to the SC matrix defined by the anatomical connections between those brain areas in the human, as obtained by diffusion spectrum imaging (DSI) and described earlier. The inter-area connections are established as long-range excitatory synaptic connections either exclusively between excitatory pools of different areas or both between excitatory pools as well as excitatory pools and inhibitory pools of different areas. Inter-areal connections are weighted by the strength specified in the SC matrix, denoting the density of fibers between those regions and by the state parameter global coupling strength G. When each cortical area is embedded in a large-scale network, their global brain dynamics are governed by the following sets of coupled stochastic nonlinear differential equations for the excitatory pool and the inhibitory pool, respectively:
In Eq. (6),
In order to obtain maximum excitatory firing rate, the global coupling strength is varied as a free parameter. The result is displayed in Figure 2D. Simulation of the global model with a range of low global coupling strength G=[0.0 – 0.5] shows spontaneous population activity with an excitatory mean firing rate of 3–4 Hz, which is what we have obtained for the global spiking model by varying the global coupling strength parameter. At even higher values of G=[1.5 – 4.0] both low and high population firing rates co-exist as steady-state solutions. For coupling above G>4.0, fixed point attractors destabilize and the system enters into a regime coinciding with a high mean excitatory population firing rate. This is shown for both the spiking model and the DMF model separately to visualize their dynamical transition properties with a range of control parameters. The critical values for phase transition are shown in Figure 2C and D as well as in Figure 8.
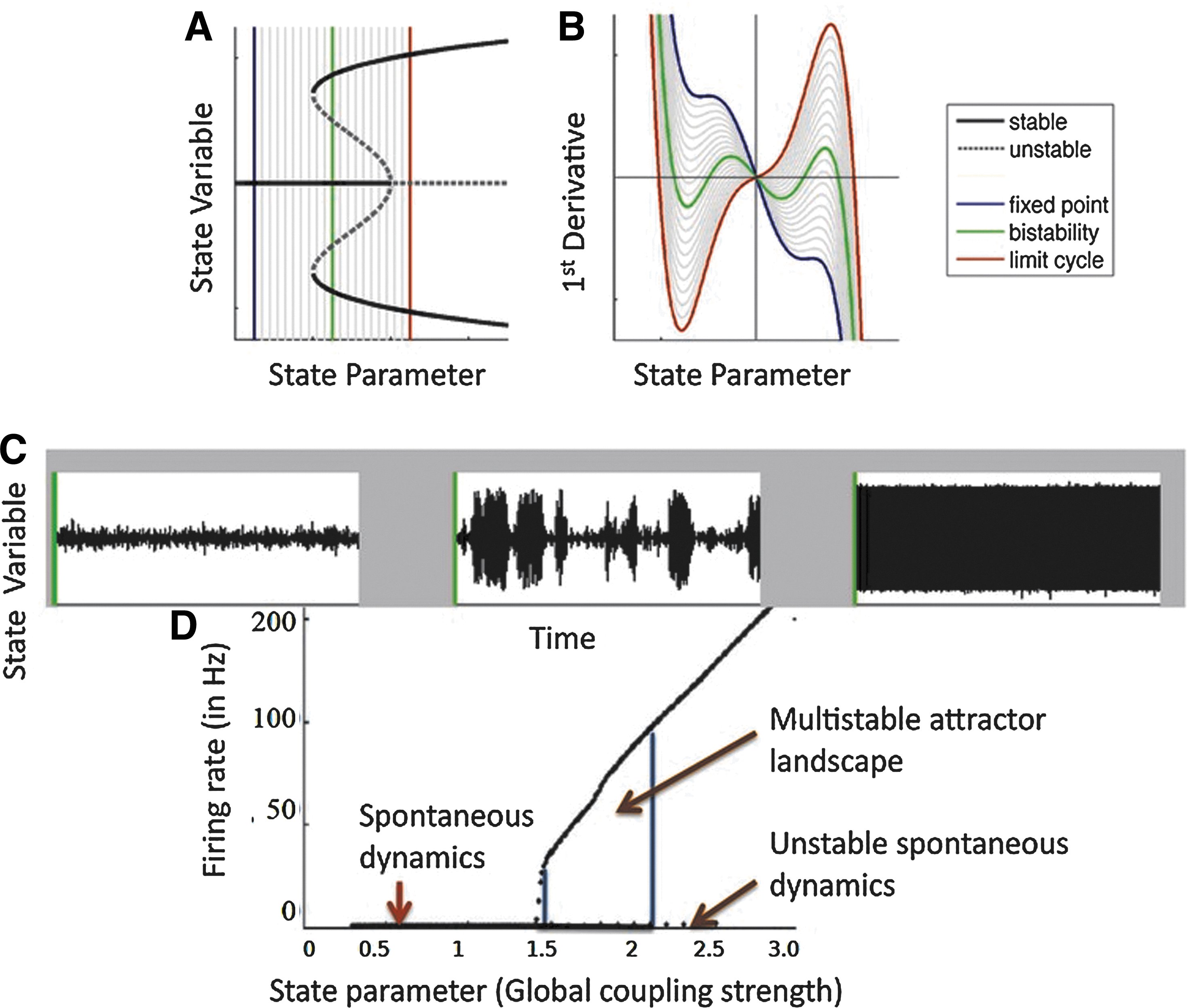
Resting-state dynamical landscape. Figure adapted and modified from Freyer and coworkers (2012b).
Numerical procedure
The local pool represented by a cortical spiking IF model when uncoupled from the global network is simulated using Python 2.7, scipy, and numpy. Resting-state spontaneous activity and the mean firing rate of each cortical module are obtained using a numerical integration procedure in the C code environment. STDP in local cortical areas is implemented using an exponential STDP class as implemented in Brian, a spiking network simulator, version 1.4. Time-frequency domain analysis is carried out on the LFP-like activity from cortical pools using the custom-made MATLAB code. BOLD signals are simulated using hemodynamic response function, and covariance between relevant brain areas is estimated using custom-made MATLAB scripts. DMF model with fully connected brain areas is simulated using custom-made MATLAB scripts that are available from the open source computational models database
Alteration of Dynamic Landscape Due to Locally Restricted Plasticity
In principle, it is possible to implement plasticity in both spiking and DMF models while taking advantage of a mechanism such as the balance of excitatory and inhibitory currents as recently proposed by Deco and coworkers (2014). This mechanism can be introduced in the DMF to clip the local area asynchronous firing rate to 3–4 Hz. Plasticity crucially depends on neurons with a similarly timed activity. Hence, in this study, we are more focused on the presynaptic-postsynaptic spike timing difference and the role of feed-forward input oscillations for the spiking of the output neurons at the precise phases of the input oscillations. Hence, amplification of network firing rates can be realized via appropriate modifications of spike timing in a mean field rate model setting. For the investigation of various effects of plasticity, we chose a spiking neuron model with a biophysically realistic attractor network. This attractor network as shown in Eqs. (1 –5) is a dynamical system with an intrinsic tendency to settle in stationary states—also called “attractors.” Attractors are typically characterized by stable patterns of firing (Fig. 2). Individual neurons are modeled as IF spiking neurons with biologically realistic excitatory (AMPA and NMDA) and inhibitory (GABA-A) synaptic receptor types. Realistic description of the synapses and nonlinear dependency of the synaptic current on the excitatory currents received from other neurons allow for including STDP. However, integrating the full spiking model is computationally expensive. This makes the exploration of the parameter space to find the parametric conditions matching the experimental findings a challenge. We know from empirical data that learning alters the rs-FC by restructuring connections employing Hebbian-like rules (Freyer et al., 2012a; Harmelech et al., 2013). We aim here to reproduce similar effects with a large-scale model implementing local STDP. The implementation of local plasticity at the node level is motivated from several relevant studies of microcircuit modeling (Morrison et al., 2007; Vogels et al., 2011). The implication for decoding of afferents in readout neuronal assemblies has been recently demonstrated, in studies that combine the PoFC and STDP mechanisms (Deco et al., 2011; Masquelier et al., 2009). It still remains an open question how these mechanisms may influence learning-related changes in distant brain areas. Recently, STDP was introduced in hierarchically modular realistic brain networks comprising leaky IF neurons (Rubinov et al., 2011) to investigate emergent criticality in spontaneous network dynamics. In this study, authors have found that STDP enabled a phase transition from random subcritical dynamics to ordered supercritical dynamics, hence broadening the parameter regime in which balanced criticality reigns. We here go beyond this work by (1) exploring the role of ongoing states for STDP-related local reorganization and (2) illuminating the spatial (global) effects that evolve from local plastic changes and resulting altered dynamic repertoire at the node level.
Implementing STDP and PoFC in a spike model
In this section, we test the hypothesis that background oscillations alter capability for plasticity in a connected network of cortical neurons. We build a spike model with STDP, where the cortical layer receives about 1000 thalamic afferent external inputs mediated via conductance-based excitatory AMPA synapses (Fig. 3). In the present spike model, node level dynamics is asynchronous with a firing rate of 3–4 Hz in the absence of an oscillatory input—as shown in Figure 4. In addition, cortical populations receive a periodic modulation of feed-forward thalamic input spikes generated at a rate of the alpha frequency (8–12 Hz). Put differently, afferent spike output from thalamus to cortex is modulated via a sinusoid as shown in Figure 3D. As a consequence, firing rates of the inputs change on a timescale highly relevant to STDP. Thalamic input spike trains are approximated as a continuous sinusoid. Randomness in the thalamic spike phases is introduced via a Poisson process. About 1000 thalamic spike trains drive a cortical node represented by IF neurons. Plasticity introduced between thalamic input layer and cortex induces a systematic change in the local microcircuit on a short time scale of milliseconds. This is carried out by systematic potentiation and depression of local cortico-thalamic excitatory synaptic weights wtc
and altering recurrent connectivity between excitatory neurons (Figs. 4 and 5). In the spiking model, the recurrent self-excitation within each excitatory population is given by the weight wEE
=W
+ and recurrent all-to-all connectivity between excitatory-inhibitory population is given by the weights wEI
=wIE
=wII
=1 and further described in detail in the spiking model section. We include plasticity in the local AMPA receptor-mediated excitatory-excitatory synapses between thalamus and cortical area S1 neurons with synaptic weight wtc
between thalamus and cortex. Recurrent self-excitation is used as a fixed parameter in the node model with and without plasticity (Deco and Jirsa, 2012). In this model, thalamocortical feed-forward synaptic weight changes in the excitatory synapses due to plasticity serve as a critical control parameter. We assume that local excitatory synapses between pyramidal cells have at least two distinct conductance states due to plasticity: a high conductance state resulting from long term potentiation (LTP) of AMPA receptors at the postsynaptic site as described by Eq. (4), and conversely a low conductance state due to long term depression (LTD) of AMPA receptors (Brunel, 2003). Transitions from a low conductance to a high conductance state can be mediated by Hebbian plasticity, that is, selective potentiation and depression of synapses (Brunel, 2003; Mikkelsen et al., 2013). In our work, LTP/LTD windowing functions are based on those proposed by Song et al. (2000). LTP/LTD can be elicited by activating NMDA-type glutamate receptors, typically by the coincident activity of pre- and postsynaptic neurons. The windowing functions describe the time window when consequences of the near-coincidence of pre- and postsynaptic neuronal activity take place. Figure 3 illustrates a model combining oscillatory input patterns with cortical spiking IF neurons equipped with local STDP. The local neural population comprising excitatory and inhibitory spiking neuron populations receives rhythmic spike inputs from multiple neurons—1000 input neurons are shown in Figure 3A. Figure 3C illustrates how Hebbian plasticity is realized through STDP and depends on the relative timing of pre-and postsynaptic spikes. In addition, here we employ a precise relationship between phase of the background oscillations and the spike phase of output neurons. This is schematically illustrated in Figure 3D. If the phase of the background oscillations, that is, phase of oscillatory LFP activity, and the spike phase, that is, phase of oscillatory postsynaptic spikes (phase delay A−
or advance A
+) matches, then the synaptic recurrent excitatory weights are updated via a linear additive rule as proposed by Song et al. (2000) shown in Figure 3D. STDP exploits this precise relationship between LFP to spike phase and has previously been successfully employed as a parsimonious learning mechanism to demonstrate stable phase locking in a feed-forward network model (Muller et al., 2011). The synaptic AMPA kinetics is governed by the following equations:
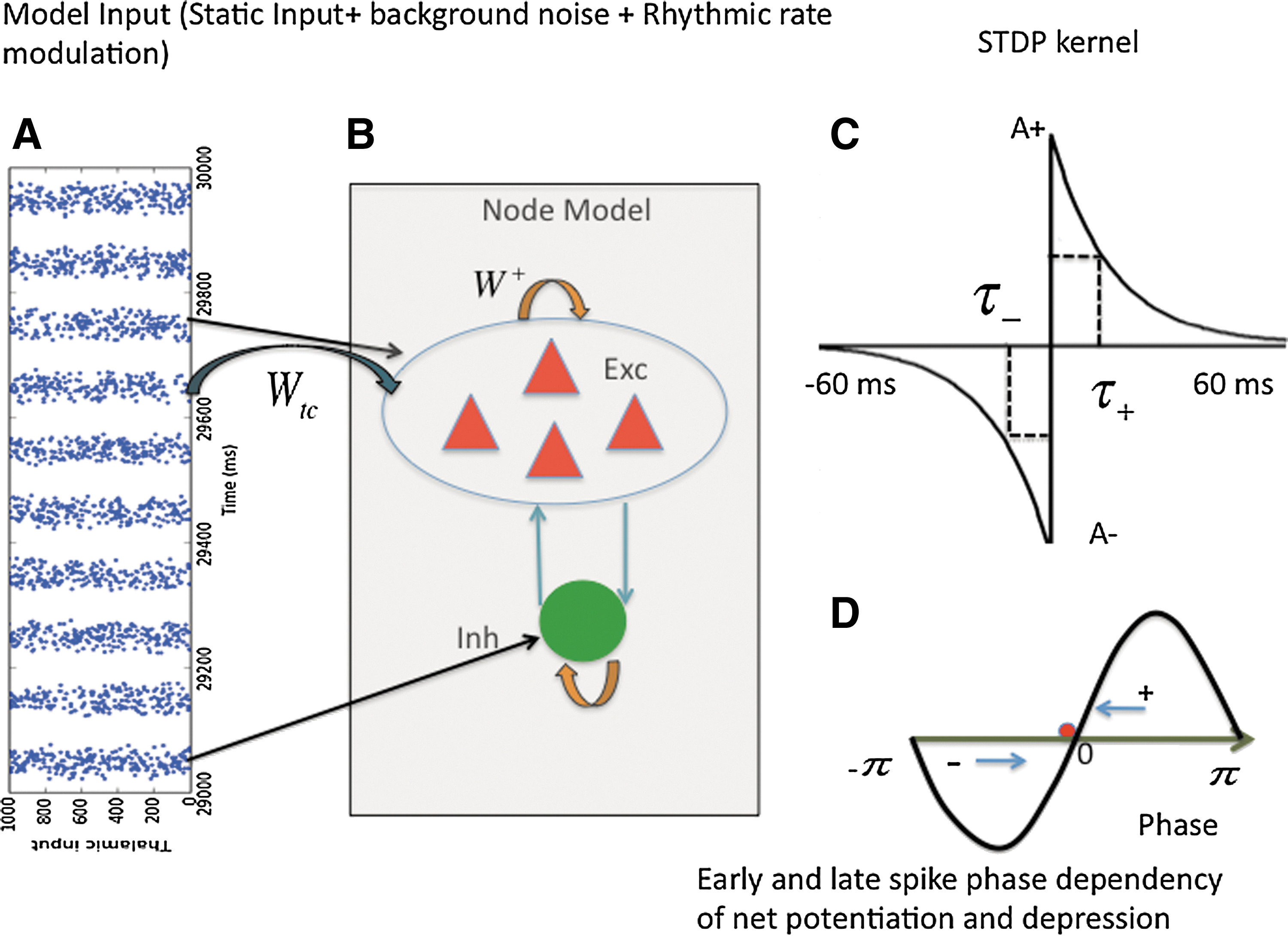
Interplay between rhythmic input and spike-timing dependent plasticity (STDP) modulates learning at the node level.
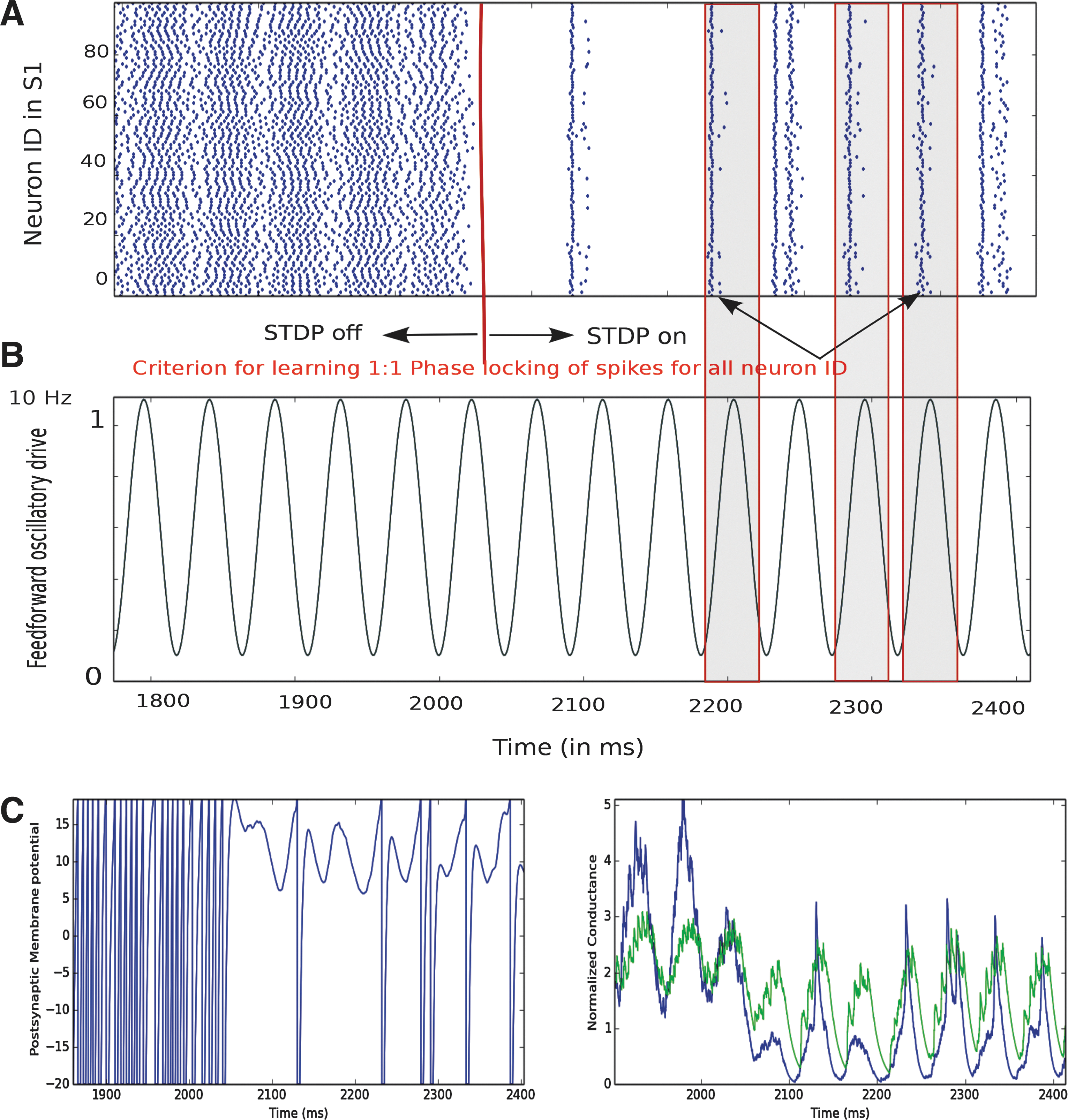
Learning with STDP in a local population S1 cortical integrate-and-fire (IF) neuron model with feed-forward oscillatory thalamic drive at alpha frequency around 10 Hz. A primary somatosensory cortical network neuron group is simulated for a total duration of 12 sec. STDP is turned on after about 2000 msec of simulation time (shown with a red marker). For clarity, 1800–2400 msec of cortical output activity are shown. Intrinsic network parameters are the same as shown in Table 2.
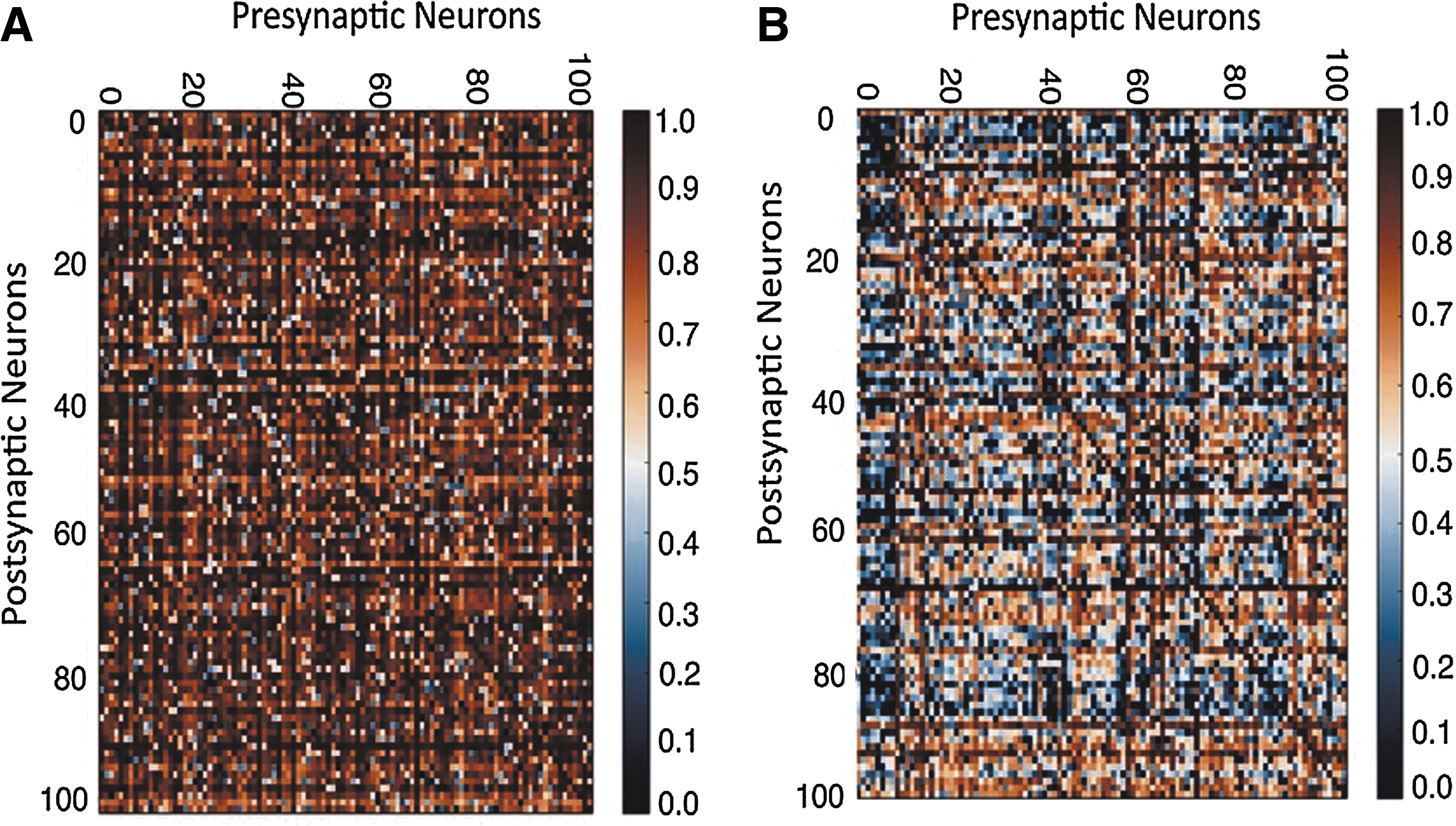
Alteration of thalamus-S1 synaptic weights (coupling strength) due to STDP, with and without brain state dependent input. Here, for clarity, only 100 excitatory presynaptic thalamic inputs are shown.
Input connections to the IF neuron are mediated by exponential synapses with excitatory-excitatory corticothalamic synaptic weights wtc
without delays. When a presynaptic spike occurs, the excitatory synaptic weight is incremented by an amount w. The STDP rule is implemented with all-to-all spike pairings (Song et al., 2000). The synaptic weights are restricted to a range between 0 and wmax
. For a given spike pairing with temporal difference x (spike time window), the synapse between the pre- and postsynaptic neurons is modified according to wtc
=w+f(x)w
max. Thus, the value of the STDP window is given by a piecewise exponential function f(x) for a given temporal difference x between pre- and postsynaptic spike times. Further, relative spike times determine the percentage of wmax
added to the feed-forward corticothalamic synaptic weights wtc
. In our simulation, when a presynaptic spike occurs in the input, the fraction of open AMPA receptor channels is updated as
where x is defined as the difference in pre- and postsynaptic spike times (tpost − tpre ), τ+ and τ− are the STDP time constants for potentiation and de-potentiation, A + is the amount of potentiation for an optimally potentiating pre-post pairing, and A − is the same for a de-potentiating post-pre pairing. The ratio of de-potentiation to potentiation (A −τ−/A +τ+) will be termed the STDP ratio. For the case of identical time constants considered here, we write (A −/A +). The value of the function f(x) determines the percent change for a synaptic weight on a given pairing, relative to wmax . Each of the input neurons in Figure 3 is modeled as an oscillating inhomogeneous Poisson process. In Figure 3A, we show 1000 periodically modulated Poisson process spike inputs. The number of inputs is chosen as a compromise between biological realism and computational feasibility. Inputs are fed to the IF neuron by exponential, current-based synapses with a 5-ms decay time constant. As shown in the scheme in Figure 3D, recurrent weights are potentiated and depressed depending on the phase of the background oscillation. By default, the local node model is in a balanced Ex/Inh regime for intrinsic network parameter combinations characterized by low-amplitude asynchronous firing activity. When input oscillations are in the alpha frequency range yielding the most prominent spontaneous oscillations of the human brain, the presynaptic firing rate changes on a time scale relevant for the STDP time window Δt=0 to 5 msec (Babadi and Abbott, 2013; Caporale and Dan, 2008). A defining feature of STDP is very high temporal asymmetry (Guetig et al., 2003). When a presynaptic spike precedes the postsynaptic spike in time, then the synapse between the two is potentiated (pre-post pairing); while in the converse scenario, the synapse between the two is depressed (post-pre pairing). The interplay between thalamic input in the presence of oscillations and STDP is illustrated with a simulation of a node model representing primary somatosensory cortical area S1 composed of 100 excitatory neurons and 25 inhibitory IF spiking neurons. The node model is simulated for a total duration of 12 sec. STDP is turned on after 2000 msec of simulation time. The network runs for another 10 sec. In Figure 4, for clarity, the simulation results for cortical output neurons are shown for the time period 1800 to 2400 msec. Intrinsic network parameters are the same as displayed in Table 2. In Figure 4A, a raster plot of population spikes from 100 cortical excitatory neurons is shown. Before 2000 msec, cortical output exhibits asynchronous spiking activity even in the presence of oscillations in the thalamic input. When STDP is turned on after 2000 msec, cortical neurons start exhibiting a precise phase of firing, that is, phase locking, followed by transient events of silence that corresponds to phasic inhibition. If the output spike occurs at the trough of the input oscillation cycle, that is, “early,” it leads to de-potentiation in the cortico-thalamic synaptic weights wtc in accordance with phase-dependent STDP. In contrast, when the spike occurs at the peak of input oscillation cycle, that is, “late,” this leads to net potentiation in the cortico-thalamic synaptic weights (Fig. 3). In Figure 4B, the simulated postsynaptic membrane potential is shown both before and after STDP. Before STDP, the membrane potential oscillates due to oscillatory feed-forward thalamic input. The membrane potential reaches the threshold of −20 mV once in every cycle. There is no phase preference. After STDP, that is, after 2000 msec, the postsynaptic membrane potential is modulated by the input phase. The membrane potential reaches the threshold only when there is potentiation due to phase matching of spikes at the peak of the input oscillation cycle. In Figure 4C, excitatory synaptic conductance (blue) and inhibitory synaptic conductance (green) are shown. The effect of phasic inhibition is strongly present in the temporal dynamics of both conductance types. Before STDP, excitatory conductance is typically higher than the inhibitory conductance due to the feed-forward excitatory drive, resulting in a number of spikes at arbitrary times as shown in the population raster plot between 1800 and 2000 msec. After STDP, excitatory conductance as well as inhibitory conductance exhibits strong phase modulation. Potentiation in this scenario leads to a homeostatic balance between excitatory-inhibitory conductance, leading to phase locking of spikes as shown in the raster plot. However, depression in corticothalamic synaptic weights leads to lowering of the excitatory drive received by the cortical neurons and thus resulting in a strong cortical, that is, recurrent or lateral inhibition dominating the excitatory conductance as shown in green (between 2000 and 2400 msec). As a consequence, silencing takes place in cortical output neurons at those time points as shown in the raster plot in Figure 4A. An increase in phase locking of spikes to the background input oscillatory phase (LFP-spike precise phase coupling) after STDP paves way for learning in the local node model that, in turn, plays a crucial role in influencing network dynamics in the globally connected nodes.
Next, we investigate how systematic potentiation and depression of local excitatory synaptic weights wtc may modify local connectivity between pre- and postsynaptic neuron pairs in a node model in the presence and absence of brain-state dependent input. To address this, we access local cortical SC by calculating synaptic weights in the presence and absence of rhythmic input and STDP (Fig. 5). Initially, presynaptic and postsynaptic neurons are connected in an all-to-all fashion. Hence, all synaptic weights are 1. Without rhythmicity in the input, that is, if STDP is random, weight changes between pre-post are also random and show up in the result (Fig. 5A). This is intuitively correct, as plasticity in this scenario is less prevalent and any change in weights between pairs, that is, potentiation or depression, due to the realization of STDP is rather random. In contrast, with a rhythmic background input, structure emerges in the layout of connectivity between pre- and postsynaptic neurons and local clustering is observed (Fig. 5B). The clustering of weights plays an important role in constraining local area node dynamics as shown in computational (Voges and Perrinet, 2012) and experimental work using two-photon calcium imaging in vivo and multiple whole-cell recordings in vitro in rodents' primary visual cortex (Ko et al., 2013). To obtain a systematic insight about local node model behavior under various conditions on a longer time scale, activity of the node network is simulated for 60 sec. Resulting summed membrane potential activity, that is, LFP and corresponding time frequency plots are displayed in Figure 6. The rhythmic input leads to an oscillation in the population-firing rate, as otherwise is not possible when relying only on the intrinsic network parameters used for the spiking node model. Integrated spontaneous activity shows intermittent low-frequency (3–8 Hz) locking (Fig. 6B). With a rhythmic input, LFP network activity exhibits network oscillations in the alpha frequency range (Fig. 6C). For a rhythmic input with STDP network, activity is shown in a time frequency plot in Figure 6D demonstrating continuous periodic oscillations and phase locking to the alpha frequency band.
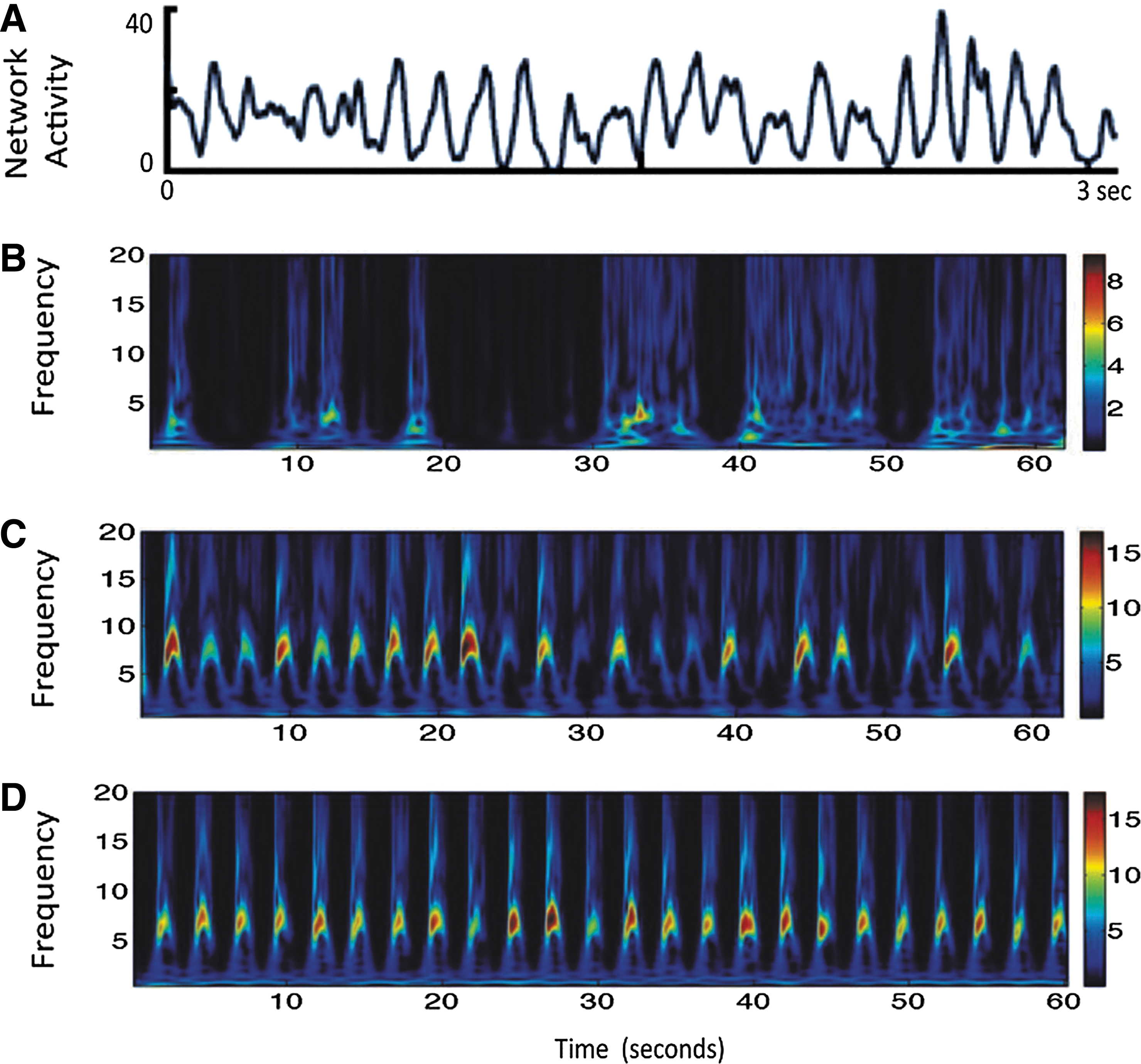
Spiking IF neuron model network activity is simulated for 60 sec. It exhibits band-restricted oscillations in the alpha frequency range.
Analytical approximation of synaptic weight change in time due to STDP in the cortical model
Rates of the presynaptic spike trains are approximated by a continuous sinusoid. Hence, feed-forward input to a cortical region is described as an inhomogeneous Poisson process:
where f is the angular frequency, and r is the peak firing rate. The spikes in the output are delivered as series of delta pulses of the following form:
where ψ is the phase offset of the output spikes. The phase distribution of the delta function reflects the assumption of phase locking in population spikes. We have verified numerically the behavior that is similar for any moderately peaked unimodal distributions.
In order to compute a correlation between input oscillations and output pulses, we perform the following integration:
where Δt is the difference in the spike times between tpre −tpost
. In other words, this reflects the STDP time window. The correlation integral is further simplified:
To obtain the change of weight w(t) over time, we make use of D(J) in Eq. (12) to write it as a function of input phase of oscillations:
Now, we can carry out an explicit integration in the subsequent steps,
We can further simplify Eq. (14) as follows:
Eq. (15) describes how—in the presence of plasticity—the synaptic weights w(t) evolve in time as a function of frequency of input oscillation f, input modulatory phase ψ, synaptic plasticity time constants τ
+, τ
−, or the ratio of depotentiation-potentiation
Alteration of FC and Dynamic Landscape Due to Locally Restricted Plasticity
In this section, we test the hypothesis that a change in local node spiking dynamics due to altered SC between thalamus and a single cortical node has an impact on adjacent as well as on distant node correlations. Here, we systematically explore how the local plasticity alters the global repertoire. How the local plastic change translates into global functional and structural restructuring is illustrated schematically in Figure 7. In a previous work, we have reported learning-related changes in empirical rs-FC as reflected by the difference in the imaginary component of coherence before and after learning (Freyer et al., 2012a). Imaginary coherence reflects the functional correlation between the areas from which EEG waveforms are recorded on the scalp, avoiding effects of volume conduction. To access the FC before and after plasticity, we simulate a global brain model consisting of six brain areas connected via long-range SC. Therefore, it will have an impact on global functional correlations across brain areas. As a first step, in the absence of plastic changes, we obtain a bifurcation diagram by varying the global scaling input parameter G to six brain regions using both spiking and the DMF model. A comparison of global dynamics between the spiking and mean field description is plotted in Figure 8A. Qualitatively, both models exhibit multistability in a range of global coupling parameter values. As shown in Figure 8B using the DMF model, a value of global scaling parameter G>4.0 network destabilizes and the selected six cortical regions exhibit high firing rates. For the color-coded representative points, we obtain population firing rates from the six selected brain regions. They display low asynchronous population firing rates around 3–4 Hz and high firing rates around 50–60 Hz for stronger scaling parameter values. In between, we obtain multi stability in the global model (Fig. 8C). Next, we introduced brain state-driven feed-forward thalamic input to a cortical node and modified the synaptic weights within that node in accordance with our synaptic plasticity rule. Using plasticity in the node model, first we estimate local coherence from resulting spike times and subsequently using local coherence to estimate a free energy function. The degree of synchronization of the spiking neurons in the local cortical area is measured by order parameter
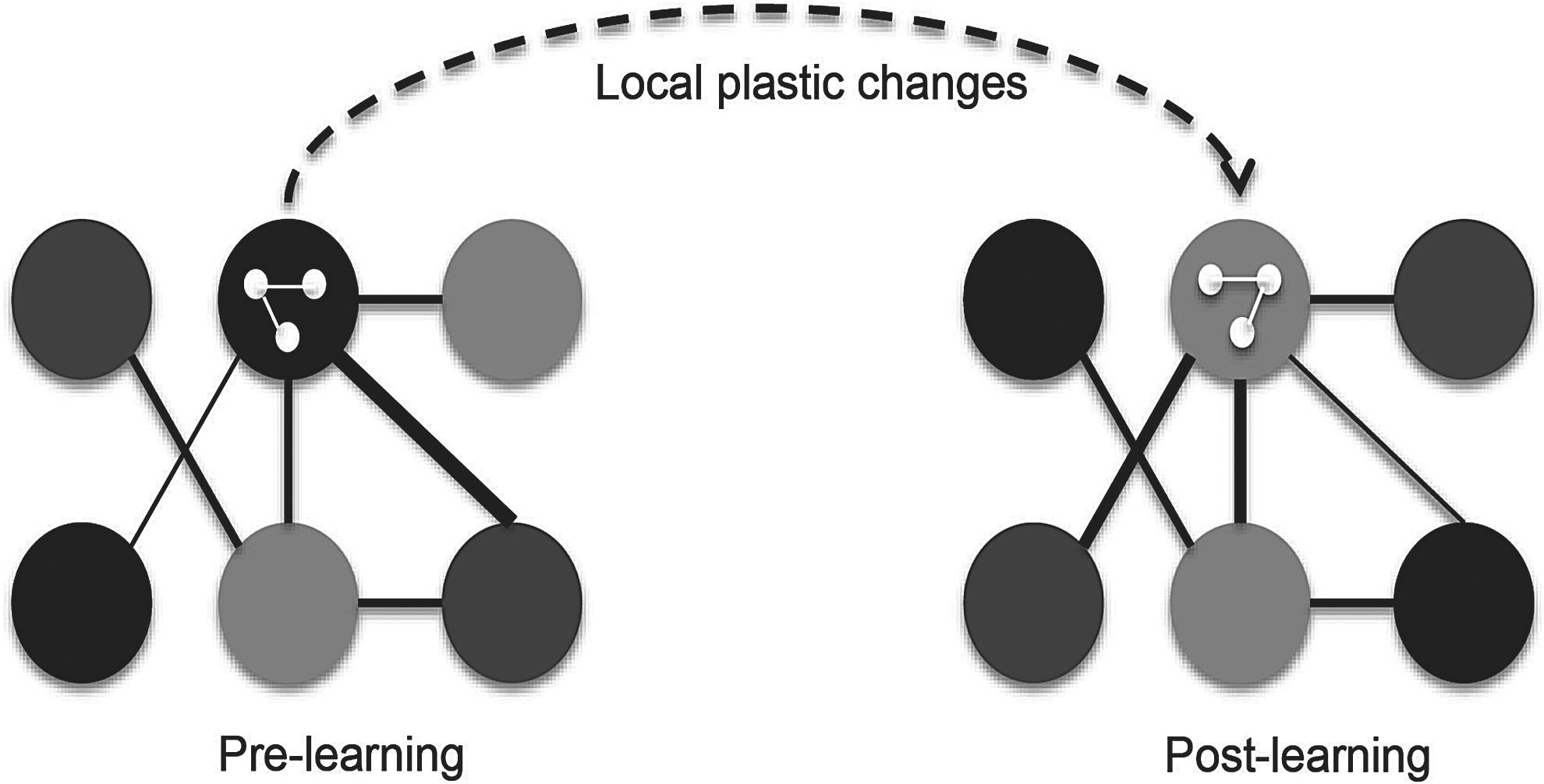
SC between six simulated cortical nodes and changes in global SC due to local plastic effects. Upper panel: A functional network is represented by six interconnected neural populations or regions represented by six large circles. Each node comprises the same number of excitatory and inhibitory neurons connected via recurrent connections within regions, represented by three small white pellets connected by white lines in the central-upper large pellet. SC is illustrated by black lines connecting the regions. Resting-state functional connectivity (rs-FC), that is, degree of correlation, is represented by the gray scales in the six regions, with similar scales indicating a high degree of coherence. After learning, as the dashed arrow indicates, local plasticity within a region changes. This is exemplified by the change of white local connections. Postlearning local changes are paralleled by global changes in FC, illustrated here by the reconfiguration of the connecting lines and the gray colors among the six regions.
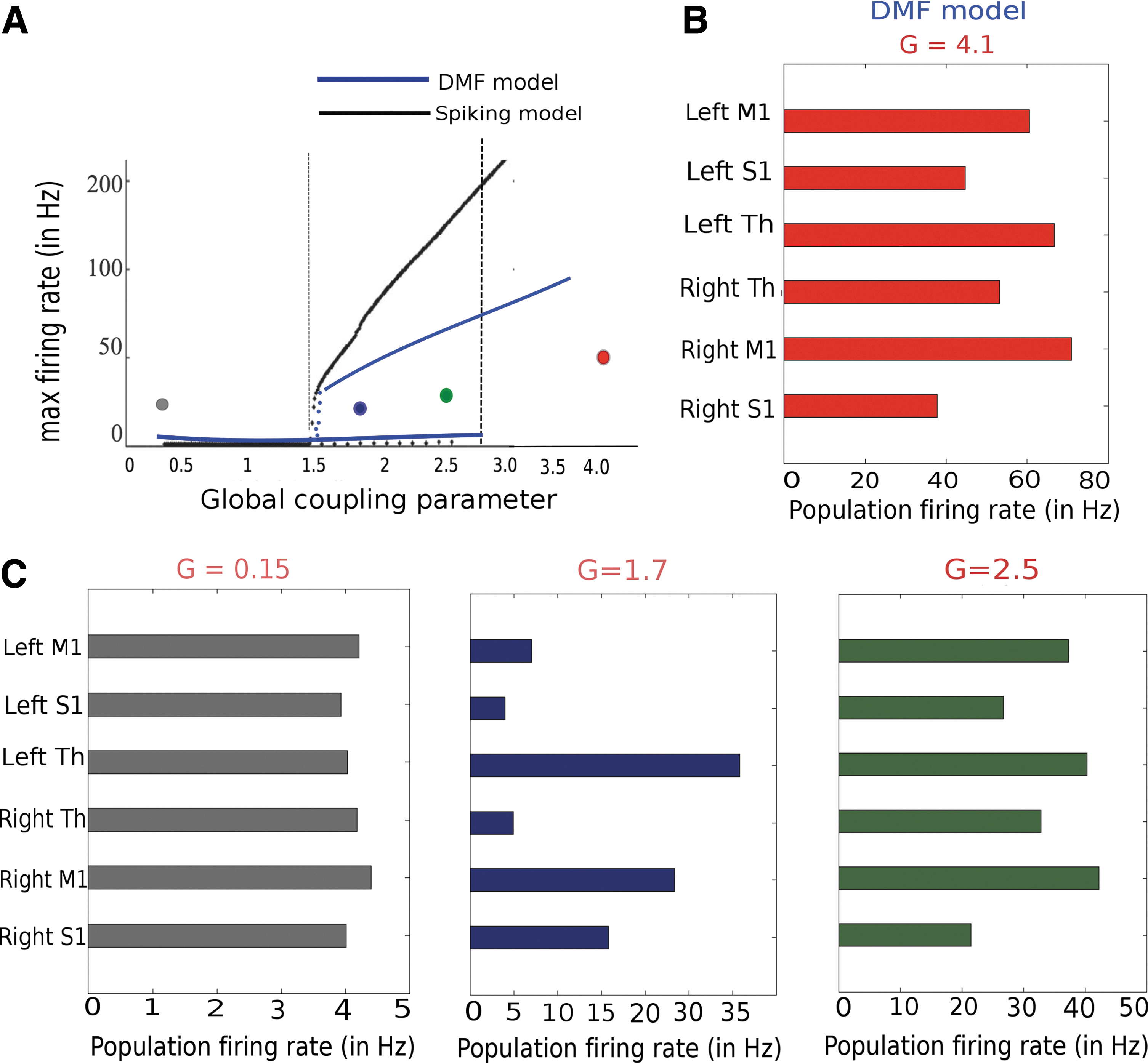
Bifurcation diagrams of the full spiking model and the reduced dynamic mean field (DMF) model.
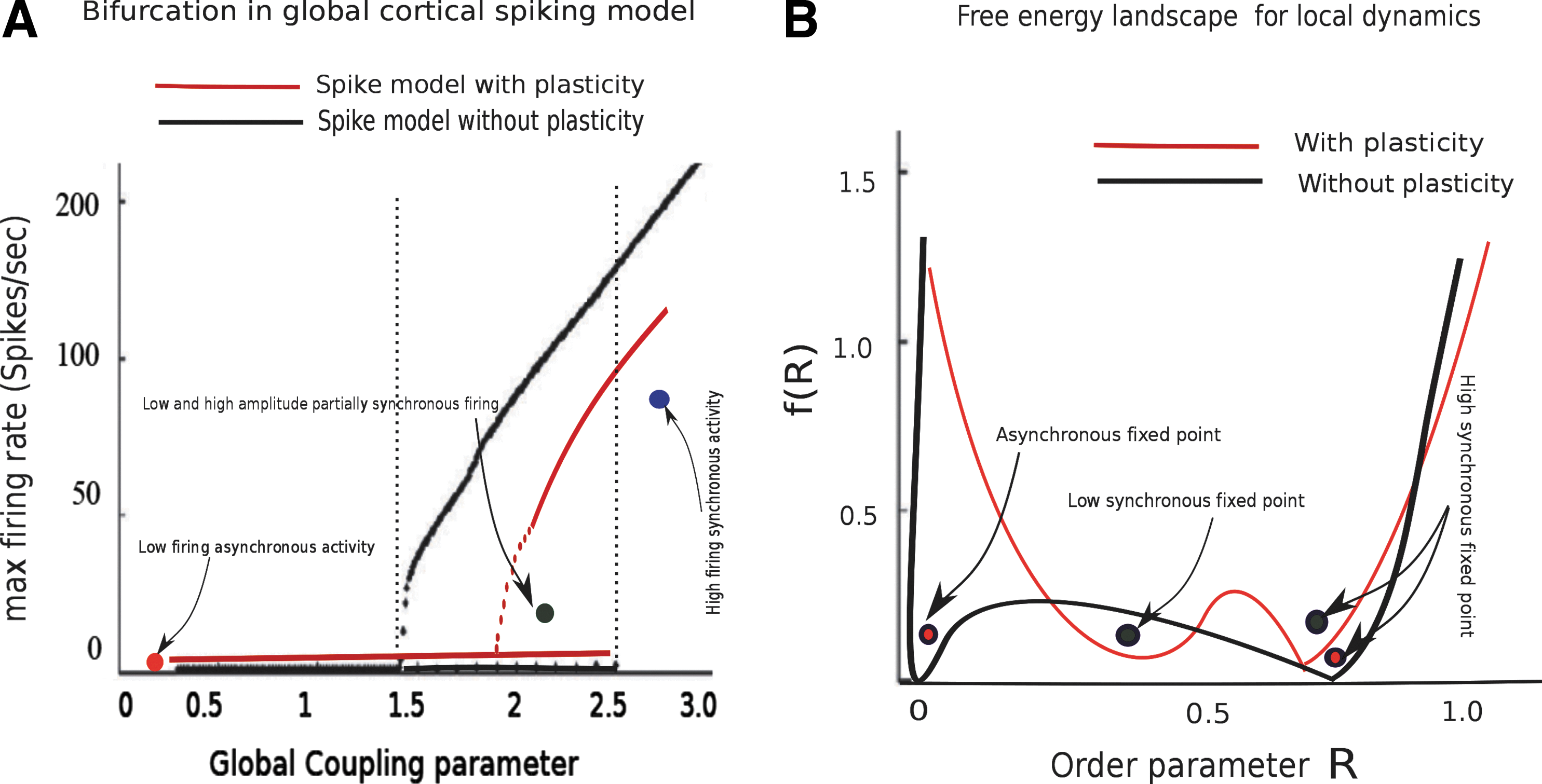
Bifurcation in the global cortical model and relationship with local area dynamics unfolded on a free energy landscape with STDP.
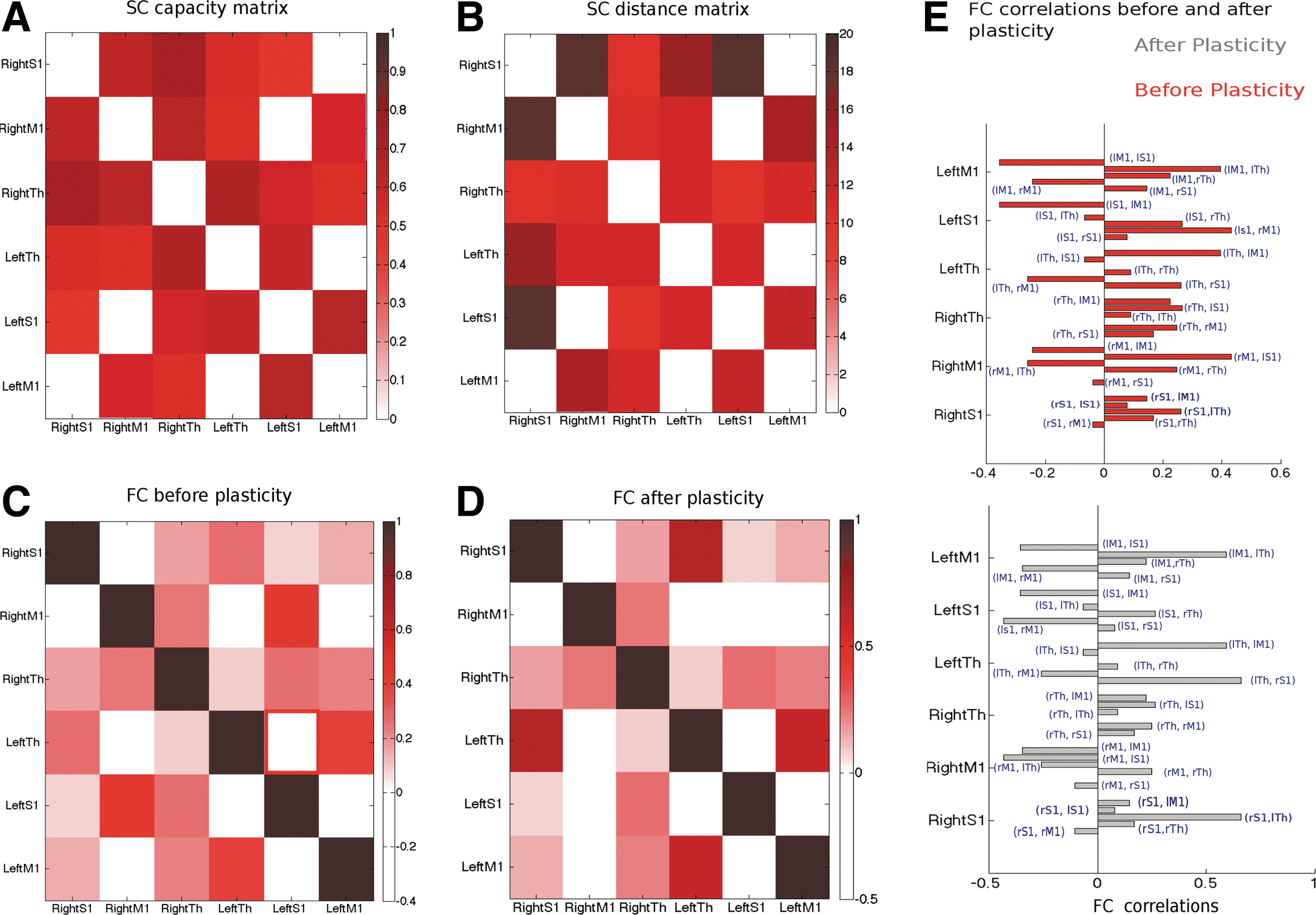
Realistic SC and simulated FC between salient brain regions.
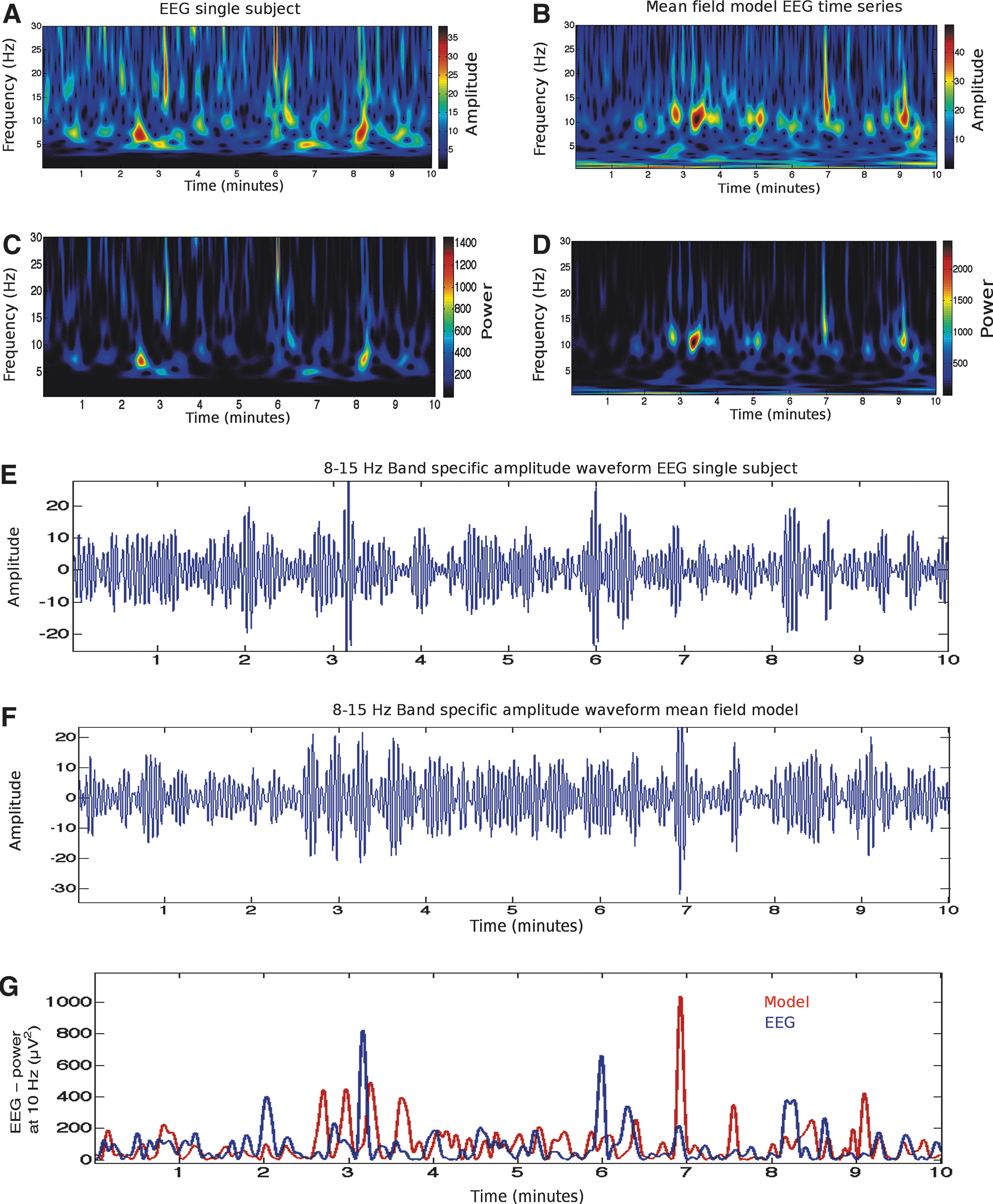
Time series analysis of experimental electroencephalography (EEG) data and the output of a mean field model.
Analytical approximation of activity covariance in the global cortical model
In order to estimate the network activity and statistics, we use statistical first- and second-order moments equations to approximate the deterministic gating equations in the DMF model as displayed in Eq. (6). We first express stochastic differential equations in terms of their first and second moments of the distribution of the gating variable:
where <.> expectation values over many realizations. In the vector field form, synaptic gating variable equations are expressed as
where for excitatory and inhibitory populations Eq. (18) can be expressed as (simply following as defined in Eq. (6) deterministic dynamics of SDE) follows:
Since we are dealing with a deterministic system, a Taylor expansion of the vector field of gating variable
With many such realizations, one can average out
Using the equation cited earlier, we can rewrite the equations for the mean of the gating variables as
where in Eq. (22), mean input current to cortical region a is thus given by
These mean input currents for cortical area a are given as follows:
In Eq. (23), wtc , w(t) are the corticothlamic synaptic weights and weight update due to synaptic plasticity.
In the equation cited earlier, widely used Bienstock-Cooper-Munro (BCM) rate-based plasticity rule replaces STDP rule. We compute a change of corticothalamic synaptic weight wtc
(t) directly as
In Eq. (24) θ−
is a sliding threshold that approximates piecewise the exponential function f(x).
Moreover, one can write the following for fluctuations about the mean gating variables:
If p is the covariance matrix between gating variables, it is written as a block diagonal matrix as given next:
where in Eq. (27)
The equation cited earlier expresses a continuity equation for the covariance matrix, which can be solved finally to obtain the Jacobian M of the deterministic system and the noise factor η
b
coming from other cortical areas. The derivatives are written out explicitly using Eq. (27):
In Eq. (27), I is an identity matrix of the dimension of the SC matrix. C is the SC between connected brain regions. Taken together, Eqs. (27) and (28) indicate the evolution of the covariance matrix and are clearly dependent on both global anatomical structure and local underlying dynamical inputs between cortical regions a, b. Since we have no other assumptions here, the result is very general. The stationary solution corresponds to a spontaneous asynchronous state as shown in Figure 11 for a specific regime of cortical stability as a function of global coupling parameter G. Moreover, these solutions of the vector field can be constructed analytically using Eigenvalue decompositions of the corresponding matrix equation of covariance as displayed in Eq. (27).
Discussion
In this work, we used a modeling approach to investigate the role of locally restricted plasticity in shaping the dynamic repertoire of the brain. A modification of synaptic weights between subcortical and cortical structure while the brain is learning or in a consolidation stage can have a dramatic impact on the resulting FNs organization. We demonstrated that incorporating plasticity in between thalamus (driver) and a local brain area of a full brain computational model enables the theoretical exploration of these mechanisms. We induced changes in the local microcircuit on a short millisecond time scale via synaptic scaling of excitatory and inhibitory conductance. This results from systematic potentiation and depression of feed-forward cortico thalamic excitatory synaptic weights wtc , which serve as a control parameter in our STDP model. The STDP description in the local cortical region is based on all-to-all spike pairing. When looking at local network activity, there are several possible sources of scaling synaptic conductance (excitatory as well as inhibitory). Hence, one should be careful with the potentiation of synaptic weights. We have applied a hard bound [0, w max] to clip the synaptic weights. Specifically, due to the phase locked stability as shown semi-analytically in the analytical approximation of synaptic weight change, overall stability of synaptic plasticity in this model is ensured. However, one should be careful of getting close to a parameter regime where the network firing rate increases monotonically without any bound, leading to an unstable spontaneous high firing rate (runaway excitation). The other important point is the spike rate adaptation in the recurrent spiking network. We have not tested this exhaustively in the local area spiking model.
One more critical assumption in this study is the low asynchronous firing activity in the local brain areas. This is indeed well supported by recent in vivo studies in the neo cortex (Haider et al., 2006; Sakata and Harris, 2009) as well applying diverse recording techniques (whole-cell patch clamp, sharp electrode recording, and two photon calcium imaging) probing laminar differences of cortical activity in a variety of sensory cortex, leading to the observation that spontaneous activity ranges between 1 and 5 Hz [for a review see Barth and Poulet (2012)]. While these observations are certainly valid, more careful comparisons are necessary to monitor the range of large-scale cortical activities from various sensory and other association cortices to further see concordance between different brain scales.
The other simplifying assumption that allows for analytical approximations is the stationary behavior of functional connectivity at rest throughout the cortex. Numerous resting-state modeling studies describe emergence of stationary resting FC as an interplay between anatomical SC and underlying cortical dynamics (Deco et al., 2011; Ghosh et al., 2008b). However, many recent studies have shown that the time-varying aspect of functional correlations plays a substantial role in the formation and dissolution of spatiotemporal patterns over the entire cortex (Allen et al., 2014; de Pasquale et al., 2012). In the context of synaptic plasticity/homeostasis, the time-varying nature of FC is even more crucial.
Nevertheless, in this study, we have exploited oscillatory features of neuronal population via STDP to explain modified FC under repetitive time-dependent stimulation. Moreover, with an analytical estimation of covariance matrix, we have made an attempt to demonstrate that among other important factors, local oscillations (local dynamics) also plays a crucial role in determining FC on slower time scales by modifying the underlying dynamical landscape.
More specifically, we have demonstrated that rhythmic input provides an oscillatory context and it is essential for neurons equipped with STDP in the output layer to detect and learn input spikes extending the concept of Masquelier and colleagues (2009), where a similar idea was implemented for a single downstream neuron equipped with STDP to facilitate learning (Masquelier et al., 2009). Second, as a result of brain-state dependent learning, local connectivity between excitatory neurons changes and clustering of local recurrent connectivity emerges. This finding is particularly interesting in the light of a recent study where Rubinov and coworkers (2011) have shown that spiking leaky IF neurons equipped with STDP with a hierarchically modular network exhibit self-organized critical dynamics (Rubinov et al., 2011). Moreover, a transition to supercritical dynamics, that is, random-order spike timing, is more evident in a regime where between-module connectivity follows a power law density distribution. Sparser local connectivity due to plastic influences and hierarchically modular SC may provide an ideal neurobiological determinant of constrained local as well as global dynamics.
Finally, we verify our hypothesis about how state-dependent input oscillations in the alpha frequency band combined with STDP facilitate learning phases of population spikes in the readout neuronal populations at a faster time scale. Based on local neuronal spike times, we analytically estimate the phases at which the system makes a transition from partially synchronous to highly synchronous fixed points (Fig. 9). Local and global population mean that firing rate is systematically modulated depending on whether the local input is received at the asynchronous or synchronous phase of firing. When the cortical region where synaptic plasticity is implemented is embedded in a larger network using realistic SC and conduction delays, the FC on a slower time scale in the order of multiple seconds is altered. Finally, we analytically approximate the prediction of FC and its dependence on local dynamics as well as given anatomical connectivity. As a possible extension of this work, it would interesting to look at a global model where plasticity/homeostatics is implemented in multiple cortical areas.
In summary, large-scale neural models of brain dynamics are promising tools to explore the non-trivial relationship between anatomical and functional brain connectivity. In particular, these models enable an investigation of the role of different factors in the network dynamics and their topological properties. Furthermore, investigating the impact of such factors in the SC-FC relationship helps understanding the mechanisms underlying healthy resting-state activity and its breakdown in disease or aging. We expect that similar results could be reproduced with virtually any model of macroscopic brain dynamics with brain-inspired SC. The DMF model discussed in our work in Eq. (6) simplifies a description of neural population at the node level and allows for analytic descriptions. One could, in principle, also explore plasticity-driven stochastic Wilson–Cowan units as shown in a recent study (Benayoun et al., 2010). It would be interesting to see how the correlated input shapes the dynamical landscape locally and globally in such models or any systematic difference than the one found in this study.
Positive and negative correlations often exhibit band limited power fluctuations (Deco and Corbetta, 2011; Fox and Raichle, 2007). The underlying mechanism of this observation may be rhythm-specific neural synchronizations. Recent computational studies further suggest that selective communication is achieved by coherence mechanisms between gain (signal to noise ratio, amplitude scaling) and firing rate modulations (Akam and Kullmann, 2012). For example, rhythmic synchrony decreases reaction time in attention and enhances speed of information transfer between different neuronal assemblies as reviewed in few recent studies (Battaglia et al., 2012; Deco et al., 2011). Hence, knowing the mechanisms in the brain that lead to altered FC is crucial to understand basic principles of brain function. A tool for their systematic exploration in a standardized and well-documented fashion is offered by TVB. In line with the synaptic homeostasis hypothesis (Tononi and Cirelli, 2014), the present simulation results suggest that an energetically, costly continuous change of synaptic weights yields the brain's full dynamic repertoire.
Footnotes
Acknowledgments
The authors acknowledge the support of the German Ministry of Education and Research (Bernstein Focus State Dependencies of Learning 01GQ0971) to P.R., the James S. McDonnell Foundation (Brain Network Recovery Group JSMF22002082) to M.B., A.R.M., V.J., G.D., and P.R., and the Max-Planck Society (Minerva Program) to P.R. G.D. was supported by ERC Advanced grant DYSTRUCTURE
Author Disclosure Statement
No competing financial interests exist.