
Abstract
This study reports dynamic functional network connectivity (dFNC) analysis on time courses of putative empathy networks—cognitive, emotional, and motor—and the default mode network (DMN) identified from independent components (ICs) derived by the group independent component analysis (ICA) method. The functional magnetic resonance imaging (fMRI) data were collected from 15 subjects watching movies of three genres, an animation (S1), Indian Hindi (S2), and a Hollywood English (S3) movie. The hypothesis of the study is that empathic engagement in a movie narrative would modulate the activation with the DMN. The clippings were individually rated for emotional expressions, context, and empathy self-response by the fMRI subjects post scanning and by 40 participants in an independent survey who rated at four time intervals in each clipping. The analysis illustrates the following: (a) the ICA method separated ICs with areas reported for empathy response and anterior/posterior DMNs. An IC indicating insula region activation reported to be crucial for the emotional empathy network was separated for S2 and S3 movies only, but not for S1, (b) the dFNC between DMN and ICs corresponding to cognitive empathy network showed higher positive periodical fluctuating correlations for all three movies, while ICs with areas crucial to motor or emotional empathy display lower positive or negative correlation values with no distinct periodicity. A possible explanation for the lower values and anticorrelation between the DMN and emotional empathy networks could possibly be inhibition due to internal self-reflections, attributed to DMN, while processing and preparing a response to external emotional content. The positive higher correlation values for cognitive empathy networks may reflect a functional overlap with DMN for enhanced internal self-reflections, inferring beliefs and intentions about the ‘other’, all triggered by the external stimuli. The findings are useful in the study of deviations in functional synergies of large complex networks associated with empathy responses and DMN in clinical applications like autism and schizophrenia.
Introduction
Temporal coherence between widely distributed active regions of the brain provides insights into anatomical and functional connectivity (Friston, 1994). Observation of the correlations over time in response to moving images like movies allows for investigation of the dynamic functional connectivity at any time instance. A movie is an interesting experience as it integrates the complexity of natural scenes of a sequence of events wrapped into a narrative in a compressed time, permitting the viewer to evolve multiple thought processes. Narratives in movies are designed to manipulate the viewer's feelings, even if temporary, to take others' (in this report, the ‘other’ implies the character or role played by the actor) perspective. The sequence of events depicting temporal developments that not only engage viewers but also promise an end is defined as narrative empathy (Keen, 2006). The viewer's reactions to the events in a movie as in real-life situations are subjective and a function of the ability to infer others' emotional state from the context or from facial expressions. The relationship that the viewer builds with the actor(s) could transcend from emotional (“I feel what you feel”) to cognitive (“I understand your feelings”) modes of empathy (Decety and Jackson, 2004; review: Walter, 2012). Investigation into the neural activation of a single region or multiple regions like the visual attention network using a movie paradigm has been extensively studied (Bartels and Zeki, 2005; Betti et al., 2013; Hasson et al., 2004; Levesque et al., 2003; Mantini et al., 2013). Responses to emotional contagion as presented in short clips from movies have also been studied, and networks specific to a multitude of emotions been identified (Goldin et al., 2005; Nummenmaa et al., 2012). The self-other feelings and engagement can vacillate in a narrative and not restricted to an impulsive reaction to a particular emotional scene or expression depicted by the actor(s), but a slowly evolving empathy response and hence investigation of dynamic functional connectivity will give insights into synergies between empathy modes. The mechanisms involved in empathy processes are reported to be top-down—from perspective or cognitive processing and also bottom-up—inputs from sensory systems, translating to distinct emotional and cognitive processes (Adams, 2001; Decety and Jackson, 2004; Preston and de Waal, 2002). Hence, it can be safely assumed that an emotional narrative, either in a real-life event or as presented in movies, by its complex content can trigger empathy comprising emotional responses, self-reflections, Theory of Mind (ToM), and reasoning (Decety and Lamm, 2006; Frith and Frith, 2003; Premack and Woodruff, 1978; Schnell et al., 2011).
Studies investigating neural correlates of empathy (meta-analysis: Fan et al., 2011; Sabatinelli et al., 2011) have looked at responses to pain inflicted on self versus other (Decety and Lamm, 2006; Lamm et al., 2007, 2011; Lamm and Decety, 2008; Morrison et al., 2004; Singer and Lamm, 2009; Singer et al., 2004; review: Lindquist et al., 2012) or of empathy toward someone of a different ethnicity (Lamm et al., 2010). A visual stimulus with actors depicting pain or an emotional state, although a temporary affiliation lasting only through the narrative, has shown to involve both the cognitive and emotional empathy modes (Blair, 2005; Decety and Jackson, 2004; Preston and de Waal, 2002). Inclusion of motor responses, as in imitation of feelings (of the other), is labeled as motor empathy. The areas associated with each of the modes reported (meta-analysis: Fan et al., 2011; Sabatinelli et al., 2011; lesion studies: Hynes et al., 2006; Schulte-Ruther et al., 2007; Shamay-Tsoory et al., 2009) are summarized into motor empathy: superior temporal cortex (BA 22), inferior parietal lobule (BA 39, 40), inferior frontal cortex (BA 44, 45), the somatosensory (BA 3), and the premotor (BA 6, 4) of the mirror neuron system (Gazzola et al., 2007; Rizzolatti and Craighero, 2004; Rizzolatti et al., 2001, 2005). Emotional empathy areas include the following: amgydala, insula, ventrolateral frontal cortex (BA 47), superior temporal cortex (BA 22), and inferior prefrontal gyrus (BA 44), while cognitive empathy covers the temporoparietal regions (BA 39, 40), temporal pole (BA 38), paracingulate cortex (BA 32), and mainly the dorsal prefrontal cortex (BA 9, 10, 46). The ToM, also phrased as cognitive ToM, includes the dorsal and ventral prefrontal cortex as main areas (Shamay-Tsoory et al., 2010) where the dorsal pathway is said to mediate cognitive empathy. Mimicking emotions of the other, considered to be an empathic response, has been found to have neural correlates covering the limbic lobe, ventrolateral prefrontal, superior temporal gyrus, and parietal region (Blair, 2005; Carr et al., 2003). The network of the insula (BA 13) and ventral anterior cingulate cortex (BA 24) as a crucial empathy circuit has been reviewed by Engen and Singer (2013).
The default mode network (DMN) covering the middle temporal lobe, posterior/anterior cingulate cortex, tempoparietal junction, and medial prefrontal cortex was originally reported in resting-state experiments and was shown to become deactivated during a goal-oriented task (Buckner et al., 2008; Mazoyer et al., 2001; Raichle et al., 2001). Later studies suggest coactivation of the DMN for cognitive tasks like projecting into the minds of others or ToM, self-reflections, for empathy response, inferring belief and intentions, autobiographical and prospective memory, or while engaged in mind-wandering in the resting-state condition (Buckner and Carroll, 2007; Buckner et al., 2008; Laird et al., 2011; Sestieri et al., 2011). The presence of the DMN for less challenging cognitive tasks (Greicius and Menon, 2004) and dynamic suppression or deactivation in these regions for focused performance in high-demand tasks (Kelly et al., 2008; Mayer et al., 2010; Mazoyer et al., 2001; Tomasi et al., 2006; Weissman et al., 2006) provides the basis for this study where we investigate the dynamic functional network connectivity (dFNC) of the identified DMN to that of the empathy networks.
Different analytical methods have been applied to study the intrinsic functional architecture that supports the interaction between two or more active regions or between networks formed of areas isolated from functional magnetic resonance imaging (fMRI) data. Rubinov and Sporns (2010) applied the graph theory to characterize functional networks where the fMRI time series of the functionally relevant area is a node and the edge is the time-series correlation between two such nodes. Considering only preselected areas might fail to reveal larger networks that are engaged in most cognitive tasks, the Bayes Net Model rectifies this by estimating on all the nodes simultaneously. A review of the various methods and advantages of each is presented by Smith et al. (2011). The ICA method is a blind source separation technique which provides statistically independent component (IC) networks from fMRI data (Biswal and Ulmer, 1999; Calhoun et al., 2001), and has been particularly applied to experiments that are non-epoch based experimental designs. The group ICA method was applied on fMRI data to putative empathy networks and were isolated for movie stimulus (Vemuri and Surmapudi, 2015). This method separates the mixed source signal from each voxel into spatially independent source signals, and the regions with temporally coherent source signals are the ICs (Calhoun et al., 2012; McKeown et al., 1998).
An estimate of the pairwise cross-correlation on the time course data of the ICs provides functional network connectivity (FNC) (Jafri et al., 2008). The question is, if the ICA method unmixes components on the principle of temporal independence, what would correlation analysis between time courses of two components suggest? There is a possibility that there exists weak temporal dependencies between components and hence analysis of the correlation between component time courses can be used to evaluate group differences (Jafri et al., 2008). Second, there is a functional overlap of the areas in more than one component sharing temporal coherence with other active regions and it is of interest to understand the role of these areas in different networks. A third aspect is the identification of networks that modulate the connectivity between two networks (Buchel and Friston, 2000). Hence, dynamic functional connectivity can identify communication between networks and investigating the fluctuations in correlations gives insights into the networks engaged in complex processing required for dynamic multimodal stimuli, such as movies.
Dynamic connectivity by calculating the cross-correlations, has been applied to study spontaneous fluctuations during the resting state (Allen et al., 2014; Arbabshirani et al., 2013; Fransson, 2005) using a sliding-window method for comparing healthy controls and schizophrenic patients (Jafri et al., 2008; Yuan et al., 2012) and has also been extended to cognitive task (Assaf et al., 2009; Coste et al., 2011; Fox et al., 2006; Sakoğlu et al., 2010). Working with patients diagnosed with the somatoform pain disorder and healthy controls, Otti and associates (2013) looked at the resting-state FNC in pain-related networks like the DMN, cingular-insular, and the sensorimotor and reported significant FNCs between these networks, but no group differences. The FNC from the time courses of DMN and the ventral and dorsal attention networks (Corbetta and Shulman, 2002) isolated by the ICA method for resting state was found to show modulation during task performance (Calhoun et al., 2008; Smith et al., 2009). Correlation comparison between active regions with known anatomical connections and unconnected active areas showed higher values for the anatomical networks while viewing natural scenes (Bartels and Zeki, 2005) demonstrating the relevance of free-viewing long duration stimuli to understand the relationship of the functional and anatomical networks. These studies offer evidence that (a) complex stimuli like movies with emotional contagion can possibly reveal many networks with spatially segregated active areas which can be identified into distinguishable DMN and stimulus-related responses like empathy networks and (b) exploring the cross-correlations from the time courses of the identified ICs provides insights into time-varying functional connectivity indicative of stimulus response.
In this study, we identify putative empathy networks from fMRI data analyzed by the group ICA method of three diverse movies, rated for emotional contagion and empathy eliciting context—a popular animation movie (S1: “Up”) clipping, an Indian Hindi movie (S2: “Taare Zameen par”), and a Hollywood movie (S3: “Green Mile”) the method for selection of ICs is reported by Vemuri and Surmapudi (2015). The story line for each is provided in the Appendix. We use the dFNC analysis methodology on time courses of the ICs estimated by the group ICA method. The analysis is expected to provide insights into the time-varying functional connectivity between the networks in response to the narrative. The hypothesis of the study is that empathic engagement in a movie narrative would perhaps modulate the activation with the DMN.
Methodology
Subjects
fMRI data were collected from 15 healthy multilingual college-going adults (age range of 20–25 years, females: 5 and males: 10). The data from one of the subjects were rejected due to excessive motion artifacts. Three movie clippings of ∼4 min each were used as stimuli. These clippings were additionally rated independently by 40 participants for emotion depiction, context, and self-emotion response at four time intervals. In a post scanning interview and survey the fMRI subjects rated S3 as the highest for all parameters, followed by S2 and S1. It should be noted that the fMRI subjects and majority of the independent survey participants were familiar with the Indian movie (S2). The order of the presentation was not counterbalanced across subjects but the familiar Hindi movie (S2) was second in the order, preceded and followed by unfamiliar movies. Resting-state data were also recorded for 5 min before the presentation of the stimulus but is not considered for analysis in this study. The Ethics Committee of the International Institute of Information Technology, Hyderabad had approved the study and all the required informed consents were obtained from the participants of the study.
fMRI image acquisition
fMRI data were collected from a 3T Philips Achieva scanner, using gradient echo, echo-planar images, with a repetition rate (TR) of 2 sec, echo time (TE)=35 msec, flip angle=90°, acquisition matrix=64×64, slice thickness of 5 mm at a gap of 1 mm resulting in 30 transverse slices at an acquisition voxel density of 3.59×3.59×5 mm. The three-dimensional T1-weighted structural image was acquired using a fast field echo technique and a Turbo Field Echo sequence, run for a duration of 4 min 46 sec with a TR of 8.39 msec, TE=3.7 msec giving 150 slices at a flip angle of 8°, field of view (FOV)=250×230 mm, and voxel volume: 0.98×0.98×1.0 mm. Due to variation in the length of the movie clippings, the number of images collected for each stimulus was different. While 130 scans each were recorded for S1 and S2, 250 scans were acquired for S3.
Preprocessing and data analysis
The preprocessing steps of realignment, coregistration, normalization, and smoothing were done using SPM8 (
Selection of ICs
The ICs were selected using three parallel methods, the first being visual inspection for artifacts, second process looks at correlation with white matter (WM), gray matter (GM), cerebral spinal fluid (CSF), and DMN template available in the GIFT templates. The third method was to acquire labels of all the areas of activation for each IC using the Talairach Daemon (3) with a 5-mm cube region of interest, and ICs with voxel clusters showing maximum white or CSF region activation were rejected. The ICs were spatially correlated with the four defined templates and selected based on highest correlation (GM and DMN) and lowest correlation criterion (WM and CSF), an approach that has been reported to be consistent (Garrity et al., 2007; Harrison et al., 2008). The templates used in this study are provided in the GIFT toolbox. In addition, the ICs were subjected to multiple regression analysis with respect to the self-reported rating on empathy collected at four time intervals for each movie clipping, with the rating as the regressor. For two ICs (IC13, IC15) of S1, the p-value was <0.05, while for all other ICs in the three stimuli, the regression coefficients were not significant. Consequently, one IC indicating posterior DMN and four ICs with areas reported for empathy modes (Vemuri and Surampudi, 2015) were selected, with conclusive identification from the significant activation in areas attributed to the three networks by examining all the 28 ICs isolated by the ICA method.
dFNC analysis
The time courses of the selected ICs were correlated pairwise to arrive at the dFNC. The approach (Allen et al., 2014) involves subtraction of remnant noise by linear detrending and replacing any outliers with the best estimate from a third-order spline fit from the smoother part of the time course. The next step is filtering using a high-frequency cutoff at 0.15 Hz, as previous work (Sun et al., 2004) has shown that functional association between the low-frequency component of the fMRI data is task related, while high frequency (>0.2 Hz) is usually attributed to noise from artifacts and hence shows no functional associations. A covariance matrix is generated by normalizing the variance of the time courses. The dynamic covariance was calculated by a Gaussian sliding window procedure with a window width of 30 TRs or 60 sec, as 64 sec was suggested to be optimal by Sakoğlu et al. (2010). The Gaussian window alpha value was 3 TRs (6 sec), and a sliding step size of 2 TRs or 4 sec was applied. The number of windows is the window size subtracted from the number of time points (TRs) which translates to 100 windows in our case. For this study we consider FNC time points only for the pairs of ICs of interest to test the hypothesis. The dFNC tool is provided in the GIFT toolbox and the methodology is reported by Allen et al. (2014). The correlation coefficients were Fisher z-transformed, and the average at each time point across all the subjects was calculated. Figure 1 is a schematic of the complete process followed in this study.
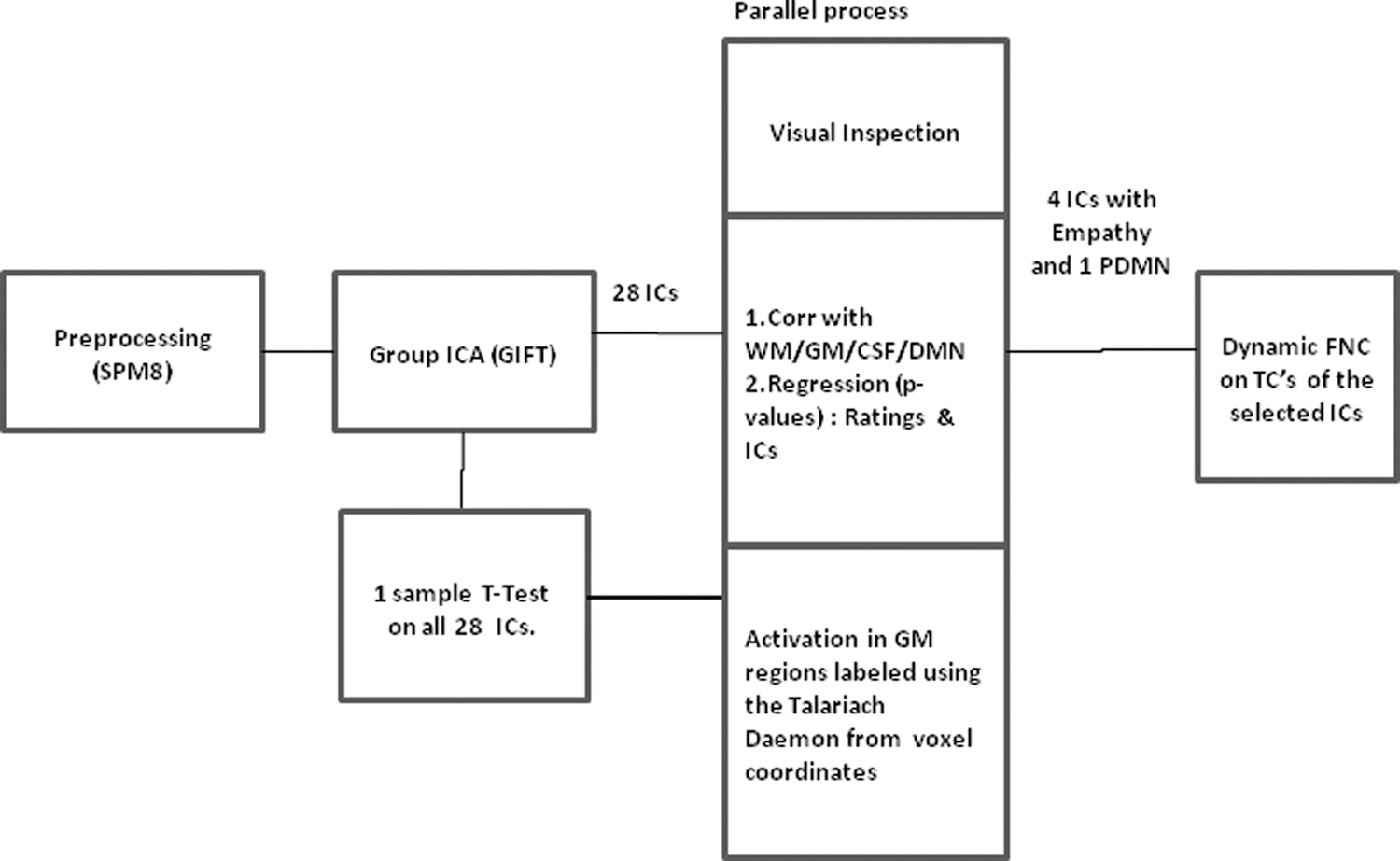
A block diagram representation of the analysis steps. GM, gray matter; CSF, cerebral spinal fluid; TCs, time courses; WM, white matter.
Results
We present dFNC analysis applied on the time courses of the IC corresponding to DMN and putative empathy networks and examine the time-varying pairwise cross-correlation cumulative of subjects' response to the events depicted in the movie narrative. The empathy response to specific events depicted in the movies collected from the fMRI subjects postscanning and through an independent survey rating on emotional self-response at four intervals of each clipping supports the following: (a) the ratings show that the movie clippings evoked emotional response and (b) the response varied across the narrative. The variation in the ratings suggests that attributing the activation seen in related areas in the spatial maps as empathy modes is reasonable. The correlation with DMN is of particular interest as it is found to be deactivated and shows very low signal changes for attentional and cognitive tasks (Mazoyer et al., 2001). For S1 (animation movie), one IC with areas covering the posterior DMN and four ICs with networks attributable to cognitive empathy were identified. Of the similar number of ICs selected for S2 (Indian Hindi movie) and S3 (Hollywood movie), it was possible to classify an IC with activation in the bilateral posterior cingulate implying a posterior DMN, an IC as an emotional empathy network, one IC with areas attributed to nearly all the empathy modes, and two ICs with a probable cognitive empathy network. An additional observation was that the components separated for S3 had significantly higher number of active areas covering the prefrontal, premotor, temporal, thalamus, and visual cortex regions in the identified ICs.
Animation movie (S1)
Table 1 lists the ICs selected for analysis from the animation movie data and Figure 2a presents the spatial maps along with the Fisher z-transformed average correlation (Fig. 2b) over the stimulus presentation duration, while Figure 2c is the survey rating plot. The fMRI participants' comparative rating on emotional response to S1 was low, while the scene-wise independent survey shows a sudden raise in the self-emotional response to the last scenes (Fig. 2c). Inspection of the ICs does not reveal activation in the crucial emotion processing areas like insula and temporal lobe (BA 22). The ICs (IC13, IC15, IC19, IC25) with significant activation in the frontal cortex (BA 9, 10, 46), parietal (BA 40), and cingulate gyrus (BA 32, 24) suggest a cognitive empathy network, and the presence of prepost central gyrus in IC13 suggests a motor empathy. The IC26 with posterior and anterior cingulate showed highest correlation to the defined reference DMN template (provided with the ICA toolbox) and hence can be considered as a posterior DMN. The average cross-correlation values for the DMN and IC25 are higher and positive with a periodic oscillation, while plots of the other three ICs (IC13, IC15, IC19) reveal lower variance and exhibit positive and negative correlations over the total scan period. The confidence interval at 90% estimated for each of the pairs (Fig. A1 in Appendix) show that for IC13 and IC15, the correlation is being equally spaced between the lower and upper interval bounds, while for IC19 and IC25, the variability seems much larger.
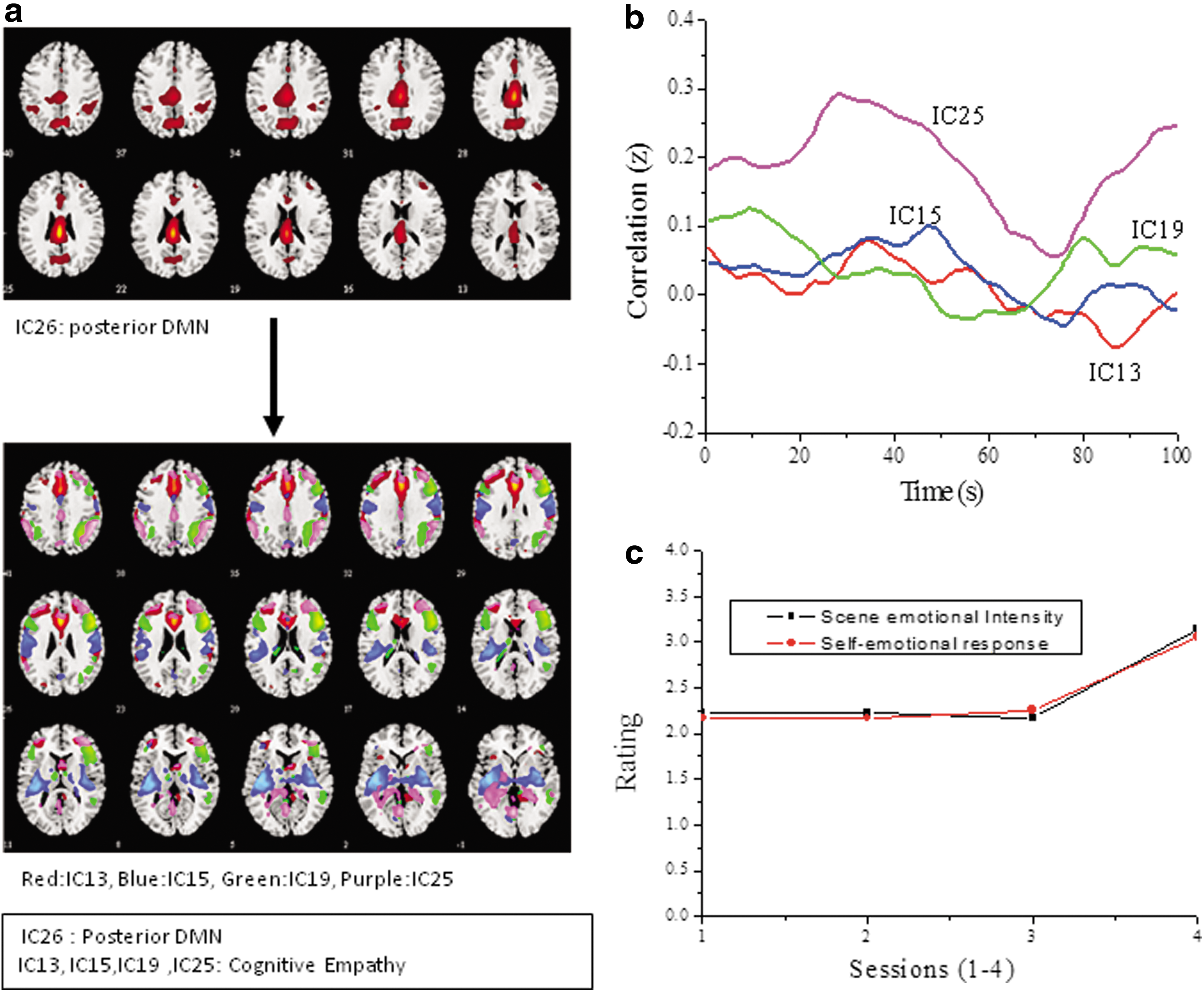
Independent components showing putative empathy networks and the DMN for the animation movie (S1).
The ICs Indicating DMN and Empathy Networks for the Animation Movie (S1)
T-value at p<0.001, FDR corrected <0.05, and voxel threshold: 10.
The MNI coordinates were transformed into Talairach space and Brodmann area labels extracted from the Talairach Daemon. The last column is the suggested putative empathy mode for each IC.
AC, anterior cingulated; CE, cognitive empathy; CG, cingulate gyrus; DMN, default mode network; ICs, independent components; IFG, inferior frontal gyrus; IPL, inferior parietal lobe; ME, motor empathy; MFG, medial frontal gyrus; MNI, Montreal Neurological Institute; PC, posterior cingulated; SFG, superior frontal gyrus; SPL, superior parietal lobe.
Indian Hindi movie (S2)
The IC identified as DMN (IC26) included medial frontal gyrus (BA 9, 46) in addition to the ventral posterior and ventral anterior cingulate gyrus (BA 23, 24) and dorsal anterior cingulate gyrus (BA 32) listed in Table 2. Figure 3 presents the spatial maps of the selected ICs (Fig. 3a), the corresponding FNC between DMN and empathy networks (Fig. 3b), and the rating plot from the survey (Fig. 3c). IC13, with activation extending to the frontal gyrus, precentral gyrus, insula, cingulate gyrus, and parietal lobe, suggests functional overlap of areas attributed to all the three empathy modes. The presence of significant activation in the insula (BA 13) in IC15 along with temporal lobe activity indicates an emotional empathy network and existence of mostly frontal lobe activation in IC19 and IC25 suggests cognitive empathy. The Hindi movie was rated second for emotional response by the fMRI participants and in the scene-wise rating by the independent participants the change is marginal with two scenes scoring slightly higher (Fig. 3c). The rating scores indicate that all the four scene windows were equally emotional with minor decreases for the first and last window. The FNC plots (Fig. 3b) show periodic fluctuations for three ICs (IC13, IC19, and IC25) with both positive and negative correlation values for IC19. The emotional empathy network (IC15) shows an initial negative and then an almost constant positive correlation or low FNC variability during most of the stimulus time duration with a sudden increase toward the end. The FNC dynamics for IC13, with activation in the frontal cortex and with a smaller cluster of activation in the insula that is reported for emotional empathy also shows lower fluctuations. In the Appendix (S2 in Fig. A1), the confidence interval at 90% for the averaged cross-correlation for each IC pair is presented. As can be seen, the confidence limits are tighter for IC13 and IC15 compared to IC19 and IC25.
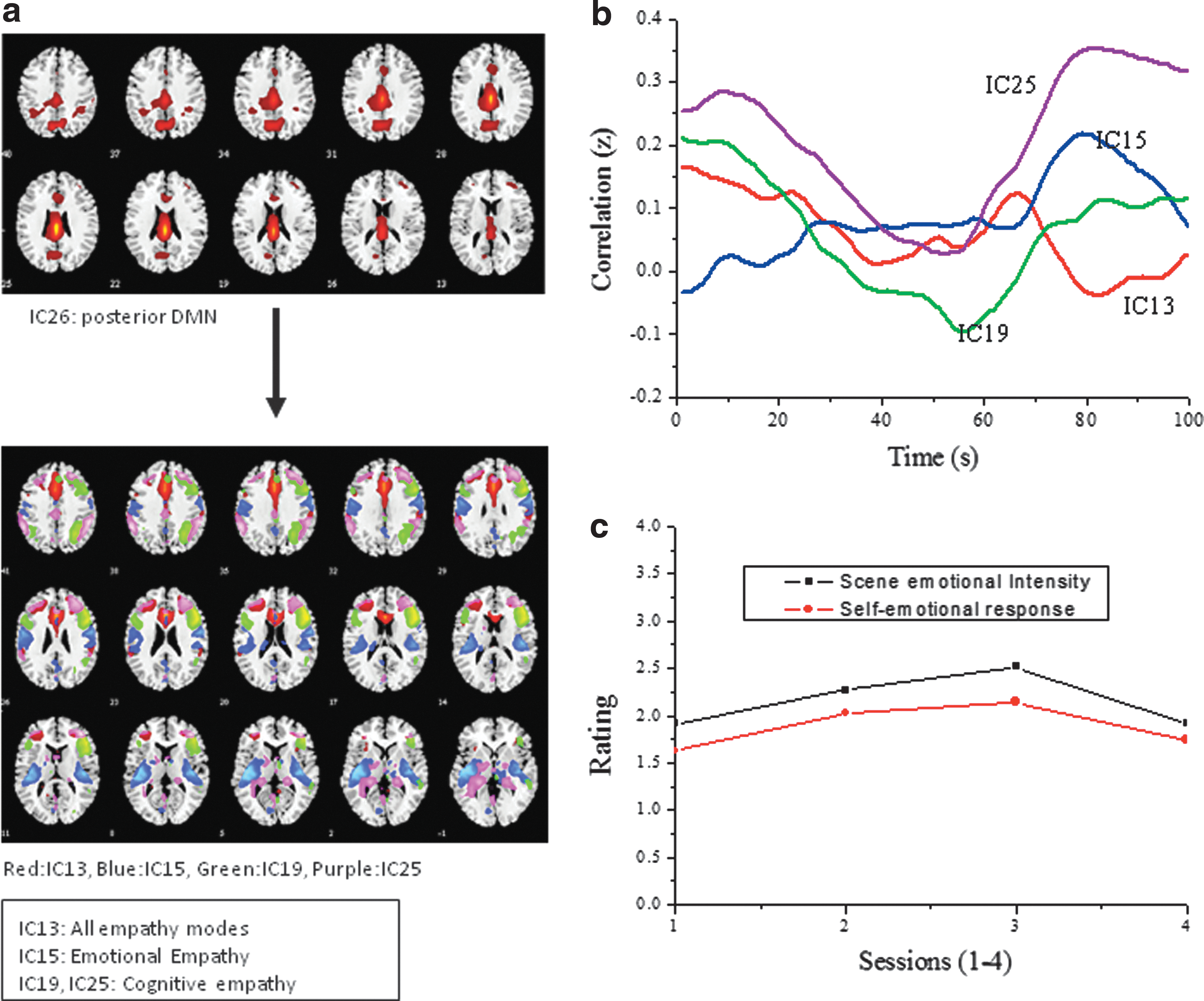
Independent components and correlation coefficients for the Indian Hindi movie (S2).
The Five ICs Representing the DMN and Putative Empathy Networks for the Hindi Movie (S2)
T-value at p<0.001 and voxel threshold: 10.
The final column is the suggested putative empathy mode labeled for each IC.
AG, angular gyrus; FG, fusiform gyrus; MTG, middle temporal gyrus; STG, superior temporal gyrus; TTG, transverse temporal gyrus.
Hollywood movie (S3)
The ICs for the Hollywood movie stimulus data reveal activation in a higher number of functionally relevant areas. Figure 4a presents selected five ICs with four of the ICs showing activation in areas reported for empathy networks (Table 3) in addition to an IC with posterior DMN. The FNC dynamics are calculated by taking the average of the Fisher z-transformed cross-correlation value for all subjects for the posterior DMN and the four ICs as shown in Figure 4b. The ICs (IC13, IC19, IC25) with significant activation in the frontal cortex areas (BA 9, 10, 46) along with bilateral inferior parietal lobule (BA 40 in IC13, IC25) and medial temporal lobe (BA 21) suggest cognitive empathy. Insula (BA 13) activation in IC15 suggests an emotional empathy network, while IC26 with activation in the posterior cingulate gyrus and precuneus indicates posterior DMN confirmed by conducting multiple regression analysis using a reference DMN template. The FNC dynamics plot (Fig. 4b) shows nearly uniform in-phase periodic fluctuations for all the four ICs with DMN while IC13 and IC15 show negative correlations too. Although the three ICs have frontal cortex activation and are identified as cognitive empathy networks the variation in the correlation with DMN for IC19 is lower compared to IC13 and IC25. The scene-wise emotion rating indicates a linear increase (Fig. 4c) in conjunction with the narrative and the scenes. The confidence interval at 90% is high and uniform for all the four IC pairs (S3 in Fig. A1).
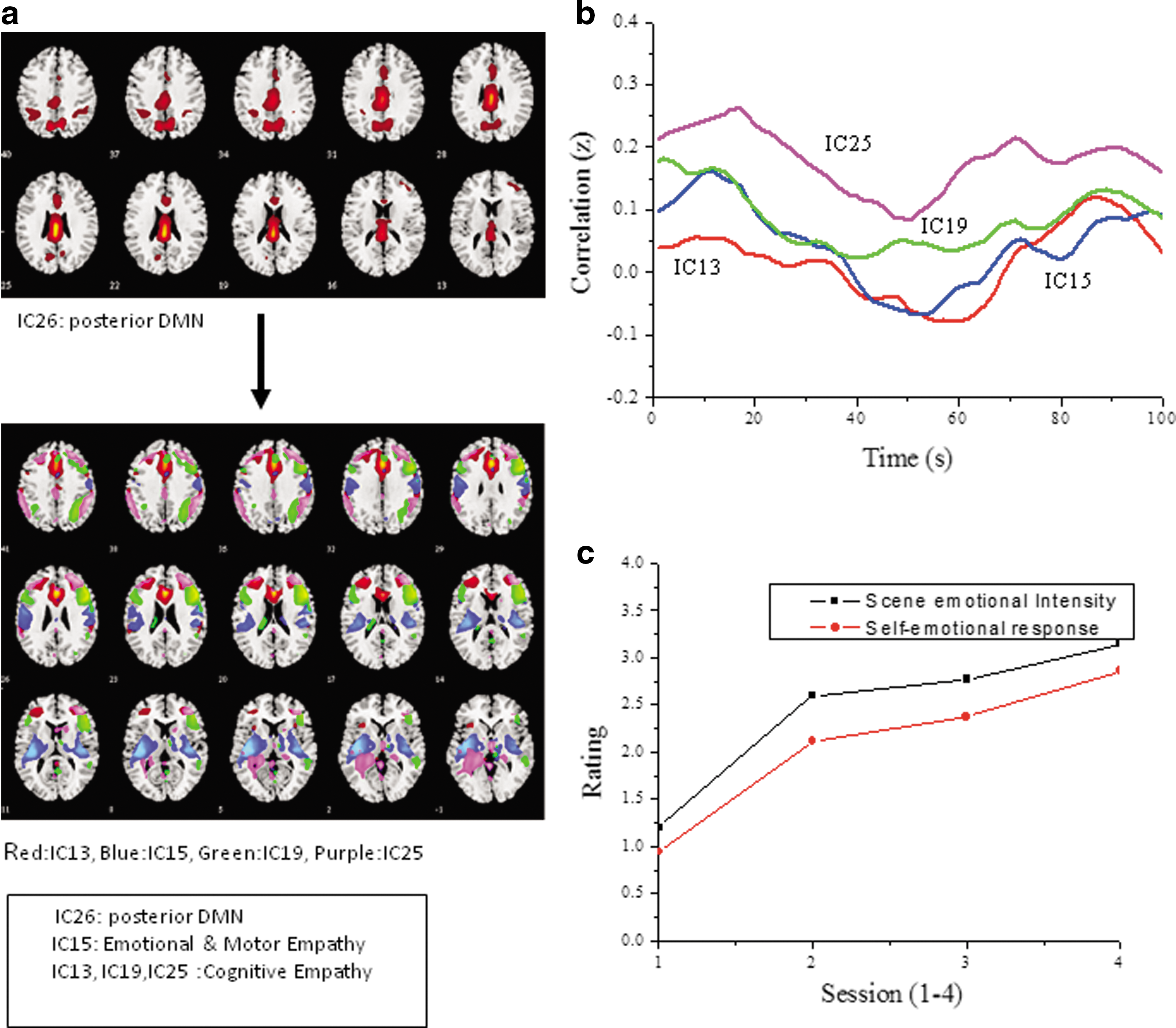
Independent components and correlation coefficients for the Hollywood movie (S3).
The Five ICs Selected for the Hollywood Movie (S3) with Posterior DMN and Putative Empathy Networks
T-value at p<0.001, with FDR corrected at <0.05, and voxel threshold: 10.
MDN, medial dorsal nucleus.
Discussion
The fMRI data were collected from 15 young adults (20–25 years of age) while they watched clippings selected from a diverse set of movies. The movie clippings were prerated by an independent set of participants and by the fMRI participants postscanning for emotional context. The rating scores provide reasonable ground to assume a varying empathy response to events, as processing of the movie narrative is mediated by various processes such as cognitive appraisal of the actor's situation and the emotional state induced in the viewer. The data-driven group ICA method was applied as the experiment was a free-viewing paradigm aimed at identifying functional networks that are active over the entire stimulus period rather than as a response for a specific event or at a certain time point. Three parallel processes were applied to identify stimulus-dependent ICs, with maximum weightage accorded to the filtering on CSF, WM, GM, and the areas of activation in each IC. The ICs from the group ICA analysis were examined and identified as cognitive, motor, and emotional empathy networks in addition to DMN, based on activation in the areas attributed to the these networks (meta-analysis: Fan et al., 2011; Sabatinelli et al., 2011; lesion study: Blair, 2005; Carr et al., 2003; Craig, 2009; Engen and Singer, 2013; Gazzola et al., 2007; Hynes et al., 2006; Rizzolatti, 2005; Rizzolatti et al., 2001; Schulte-Ruther et al., 2007; Shamay-Tsoory et al., 2009). The exploratory study identifies empathy networks based on reports in published research. As a result, the current study does not allow us to make major interpretations, but supported by the rating scores it allows for tentative inferences forming the basis for further research to consolidate the current findings.
The basic premise of our study is that multimodal complex stimuli like movies can evoke a multitude of time-varying event-specific responses which translate to simultaneous (de)activation in spatially separated regions of the brain and also across networks. The presence of simultaneous activation in many areas, which cannot be categorized into definite functional networks and conjecturing the role each area plays in different network configurations make interpretation of data from complex stimuli challenging. The dynamic correlation analysis between the DMN separated from the free-viewing experiment and emotional responses to the stimuli is aimed at inferring the role of the DMN in the nonresting-state condition. Understanding the change from the equilibrium state (Raichle et al., 2001) of this network for empathy response will help in analysis of clinical conditions like autism and schizophrenia, conditions that show deficits in emotional response. Considering that the ICA method was applied and motion or noise signals were not regressed out as usually done when using the general linear method (Desjardins et al., 2001) the effect of global signal regression leading to anticorrelations (Murphy et al., 2009) can be ruled out.
Functional connectivity—S1 (animation movie)
The dFNC of the animation movie is interesting and is in contrast to that of the live-action movie data as the ratings indicate constant emotional response for three-fourth the movie clipping with an increase for the last scene and the fMRI data do not reveal any activation in the areas attributed to emotional empathy, while cognitive and motor networks were comparable (Fig. 2). The dFNC plots indicate lower correlation for IC13, IC15, and IC19 with the DMN, where IC13 suggests a motor and cognitive empathy network and the latter two possibly indicating cognitive empathy. The low correlation values suggest possible disengagement with DMN, especially for the network (IC13) with motor empathy areas. The higher correlation and fluctuations for IC25, also identified as cognitive empathy, could be due to the presence of parahippocampal gyrus, which is reported as a primary hub of the DMN (Ward et al., 2014) and this area also did not show deactivation in a working memory task in contrast to other DMN areas (Koshino et al., 2014). The IC25 correlation with DMN (IC26) should also be interpreted cautiously from the 90% confidence interval estimates, which show instability at the upper interval (S1 in Appendix Fig. A1).
Functional connectivity for S2 (Indian Hindi movie)
The FNC with IC15, an IC with substantial activation in the insula (BA 13), hints at an emotional empathy network being relatively stable and has a very low correlation with an increase in correlation toward the end of the stimulus period. The rating on the emotional response from the survey indicates smaller changes between scenes and a decrease in rating at the end. The lower and relatively stable FNC of IC13 noticed for IC15 with areas that are identified to be from all the three empathy modes indicates a lower engagement between the networks, possibly due to a greater focus for emotion processes and inhibits mind-wandering for which the DMN is associated. The components (IC19, IC25) representing cognitive empathy show recurrent FNC fluctuations with DMN, with IC25 having higher correlation values possibly due to the presence of activation in the parahippocampal gyrus. The confidence interval at 90% indicates a relatively uniform lower confidence for the upper interval for all the four ICs.
Functional connectivity for S3 (Hollywood English movie)
The fMRI recording was for 460 sec and for group analysis the time points of all stimuli have to be equal (260 sec); hence, time points of scenes with no relevant action were cut—for example, in S3, there is a 90-sec scene showing Duncan being lead by the guards from his cell to the execution room. The cut scenes were checked for emotional content while also taking care of minimal disruption in the narrative. The FNC showed uniform periodic fluctuations in all the four selected ICs with IC25 and IC19 with a probable cognitive empathy network having significant activation in the dorsal medial prefrontal cortex showing higher correlation values, which could mean functional overlap with DMN as this network has also been implied for self-referential processing. The IC15 with activation in insula shows negative and lower correlation values for a short period of time, which is consistent with the findings for S2 and decidedly indicating that an emotional empathy response could lead to disengagement of DMN. The correlation plot of IC13 with areas covering cognitive empathy shows a similar trend, which could be due to the presence of significant activation in the anterior cingulate (BA 24) reported also for emotional empathy (Engen and Singer, 2013). The plots for S3 show nonrandom distinct periodic oscillation for all the ICs, except IC19, which shows lower modulations. The rating for emotional response increases over the movie narrative in line with the events supporting the presence of significant activations in the areas attributed for the emotional empathy network. The confidence interval estimate at 90% shows stability of correlations observed for all the four ICs, which is in contrast to the findings for the other two stimuli.
For all the stimuli the positive correlation of FNC with probable cognitive empathy networks is consistent with reported findings of DMNs involved in the ToM (review: Buckner et al., 2008; Spreng et al., 2010) process, a precursor for cognitive empathy (Mars et al., 2012) and for social cognition (Buckner and Carroll, 2007; Buckner et al., 2008; Laird et al., 2011; Mars et al., 2012). The negative and lower correlations noticed for ICs with emotional and motor empathy network indicate possible suppression of DMN as emotion processing requires focused attention observed in prior studies on attention and DMN (review: Buckner et al., 2008; Fox et al., 2005, 2006; Fransson, 2006). The stimulus-dependent response relationships between the DMN and motor empathy networks suggest a more complex affiliation than the posited suggestion of predominantly negative correlation for goal-directed tasks (Rizzolatti, 2005; Rizzolatti et al., 2001; Weissman et al., 2006) or during self-related cognition (Uddin et al., 2006), especially when extended to natural scene stimuli. The relationship between task-negative networks or DMN and the task-positive activation in a task paradigm comprising the dorsal parietal and lateral prefrontal cortex (Raichle et al., 2001; Shulman et al., 1997; Spreng et al., 2010; Uddin et al., 2009) is either one of tension between the two or a dual-process model (Evans, 2003; Jack et al., 2013) with independent and simultaneous activation in response to an external stimulus. In a study that compares working memory network to DMN it was also reported that during the task preparation state both the networks were activated, while the execution stage saw a deactivation (Koshino et al., 2014). This observation when applied to empathy can hint at a positive correlation during the cognitive processing of the events in the stimulus followed by negative correlation for scenes with a higher emotional contagion. The positive correlation values observed between DMN and the ICs with cognitive empathy and the lower correlation with nonperiodic fluctuations could possibly be due to emotional and motor empathy being more demanding. This inference needs to be verified in the future by more focused experiments.
Although intrastimulus comparison was the focus of the current study considering the diversity of the movies used it was interesting to examine the dFNC of each selected IC in a combined plot. Figure 5 depicts grouped data, combined according to the empathy network identified. The ICs (S1 and S2: IC13, S3: IC15) with activation in the precentral gyrus region (BA 3, BA 6), with either a cognitive or motor empathy network, show lower fluctuations for all the stimuli (Fig. 5a). The dFNC plot for the emotional empathy network (Fig. 5b) shows a lower fluctuation for S2 (IC13 and IC15), whereas for S3, we see an almost periodic fluctuation. The empathy response ratings for S3 were higher and the correlation plot for the empathy network (Fig. 5b) shows negative or inhibition for a time period. The plots for the cognitive empathy (Fig. 5c) network of IC25 with significant activation in the parietal lobe (BA 40) and hippocampus areas disclose comparable high periodic variation, while IC19 with common activations in the medial frontal cortex extending to BA 6 for S2 and S3 showed synchronous fluctuations. The correlation plots and the neural correlates identified for the ICs tentatively suggest that emotional empathy and motor cortex activation result in inhibition or even anticorrelation values, while cognitive empathy is mostly positive and higher.
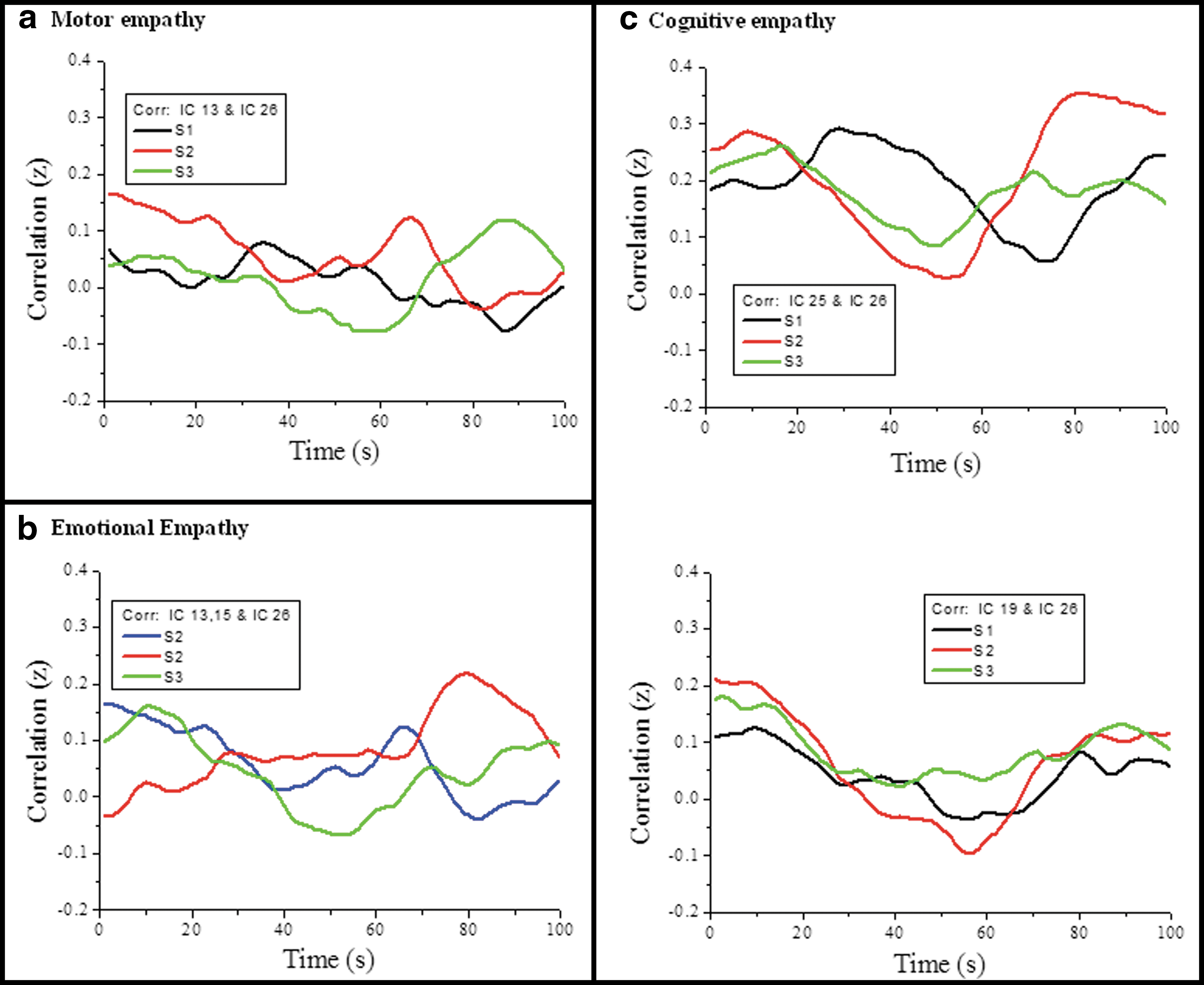
A combined plot of the cross-correlations for the three stimuli,
From the combined plot of Figure 5, we can infer a possible dual process, where distinct cognitive modes are applied for social information processing like inferring the state of mind of others and also of objects, as can be possible when watching complex stimulus-like movies. The motor and emotional empathy network correlation plots suggest a possible tension between the two networks, leading to an inhibition of the DMN.
Taken together, these results suggest a possibility of attention being more focused for emotional events reducing mind-wandering or self-reflections, which underlie cognitive empathy. As causality cannot be established by the current analysis, anticorrelation may also imply a control of emotional empathy by indulgence in perspective taking by distinguishing the self (viewer) and other (actor) or vice versa. This suggests that FNC analysis is an insightful tool to understand how regions or networks evolve and engage in real-life natural scene processing and may be helpful in investigating empathy deficiency in clinical conditions like schizophrenia and autism.
Limitations
In terms of limitations, we would like to mention that the stimuli were rated for emotional context of the narrative and the depiction of emotions by actors, but the participants did not tag specific events during the scanning process as it would intervene in the immersion in the narrative. This prohibits us from making conclusions on causes for change in state and the nature of engagement at different time points while viewing of the movie clipping. However, the rating provided by independent participants show the possible trends. Second, the regression analysis with the independent ratings taken at four intervals exhibited a significant p-value for some ICs; however, regression analysis by itself did not allow us to identify all the task-relevant ICs. A focused experimental design is required to make a direct inference between the empathy networks and the self-reported empathy response. Third, by not using a model-based functional connectivity analysis like dynamic causal modeling, our results do not allow for inferences on causality between the networks.
Conclusion
This exploratory study was aimed at understanding the dynamic connectivity changes between DMN and putative empathy networks isolated from a free-viewing experiment to decode the correlation between the networks during the processing of dynamic and complex stimuli. The identification of empathy networks is based on the areas reported to be active in many empathy response studies, and the confirmation that our stimulus does evoke empathy is evident from the variable scores provided by participants. The study using long natural viewing paradigm does not allow strong interpretations on the causal role of the areas and networks to the stimulus; however, we can safely assume that moving images with narratives are engaging for short periods of time if not all the while and this short attention span could be captured and analyzed with robust methods. Second, activation in certain areas reported for empathy for a particular experimental paradigm-like observation of someone being in pain might not necessarily be applicable for all emotion-evoking stimuli and this is the effort of all ongoing research on multiple roles assumed by a single area or a network by experimenting with different stimulus types. Our exploratory study is a step in this direction as natural viewing with no intermediate tasks permits us to look at complex constructs like empathy as it is not a sudden reflex response, but that which requires context assimilation and strategizing an appropriate response. Considering that the movies were diverse the scrutiny of the data was within the movie than intermovie, although the group ICA allows for comparison of global trends. Further experiments comparing short and long empathic narratives are being planned that will highlight differences in empathy networks and the dynamic correlations between time series of specific areas and networks, respectively.
Footnotes
Acknowledgments
Funding acknowledgment: partially funded by the National Programme on Perception Engineering Phase I, Department of Information technology (DIT), Government of India. The authors thank the National Brain Research Center (NBRC), Manesar, Gurgoan, India, for providing access to the fMRI equipment to conduct experiments and also to all the participants. The authors also thank the members of the SPM and ICA forums for timely advice and guidance.
Author Disclosure Statement
No competing financial interests exist.