
Abstract
The human brain has been described as a complex network, which integrates information with high efficiency. However, the relationships between the efficiency of human brain functional networks and handedness and brain size remain unclear. Twenty-one left-handed and 32 right-handed healthy subjects underwent a resting-state functional magnetic resonance imaging scan. The whole brain functional networks were constructed by thresholding Pearson correlation matrices of 90 cortical and subcortical regions. Graph theory-based methods were employed to further analyze their topological properties. As expected, all participants demonstrated small-world topology, suggesting a highly efficient topological structure. Furthermore, we found that smaller brains showed higher local efficiency, whereas larger brains showed higher global efficiency, reflecting a suitable efficiency balance between local specialization and global integration of brain functional activity. Compared with right-handers, significant alterations in nodal efficiency were revealed in left-handers, involving the anterior and median cingulate gyrus, middle temporal gyrus, angular gyrus, and amygdala. Our findings indicated that the functional network organization in the human brain was associated with handedness and brain size.
Introduction
Handedness has been considered as not only a natural and noninvasive marker of functional asymmetries, but also a hallmark of human evolution. Behavioral studies have associated handedness with differences in language lateralization. Specifically, right-handers have shown left hemisphere dominance in language (Price, 2012), whereas left-handers are primarily characterized by rightward asymmetry (Knecht et al., 2000) or the bilateral speech representations (Rasmussen and Milner, 1977). For example, left-handers showed a less left lateralized morphometry of planum temporale than right-handers (Steinmetz, 1996) and anatomical leftward asymmetry of central sulcus decreased significantly from consistent right- to mixed- to consistent left-handers (Amunts et al., 2000). The difference in lateralization of face- and body-related visual areas between left-handers and right-handers also has been observed (Badzakova-Trajkov et al., 2010; Willems et al., 2010). In right-handers, much larger activation was located in the right fusiform face area and extrastriate body area, however, in left-handers, activation in these areas was similar in the two hemispheres (Willems et al., 2010). In addition, non-right-handers (left-handers and mixed-handers) were reported to show a higher ratio in developmental disorders occurrence, such as autism (Dane and Balci, 2007), schizophrenia (Dragovic and Hammond, 2005), and epilepsy (Sveller et al., 2006). Left-handers, however, are often excluded from research cohorts to reduce variance in the data, which limit the findings to generalize to the whole population. For a more detailed review of variation and research necessity of left-handers see reference (Willems et al., 2014).
Brain size is an important factor that shapes neural systems and accounts for the behavioral and cognitive variability between individuals. It is often considered as a determining factor in cognitive abilities (Reader and Laland, 2002), behavioral flexibility (Sol et al., 2005), and intelligence (Gould, 1974). That is, higher cognitive abilities were possibly correlated with larger brain size (Reader and Laland, 2002). However, increasing brain size may impose constraints on distributed and integrated networks through longer information transfer delays. The advances in neuroimaging techniques and analysis methods have allowed us to better understand the information transformation. For example, the human brain has been described as a complex network characterized by small-world properties, which not only support the local neighborhoods and distributed information processing, but also maximize the efficiency of information transmission at a relatively low wiring cost (Bullmore and Sporns, 2009). Brain size effect and efficient network architecture of human brain raise the question of a possible relationship between how efficiently the spatially distributed brain regions are organized with varied brain sizes.
Multiple lines of evidence suggest that the functional efficiency may correlate with the brain size and handedness. For example, smaller brains have shown higher local efficiency in females, but not in males using diffusion tensor imaging (DTI) techniques (Yan et al., 2011) and female brains are on an average 10% smaller when compared with male brains (Peters et al., 1998). These collective findings reinforce the expectation that brain size has substantial effects on the efficiency of functional connectivity. In addition, brain size was reported to correlate with handedness, that is, left-handers had a larger brain size as compared to right-handers (Josse et al., 2006), which, however, contradicted with one study on a large sample of 112 subjects (Herve et al., 2006). Furthermore, some effects of brain volumes would have been less significant if handedness had not been taken into account (Josse et al., 2006). Quantifying the variability of functional efficiency and brain size with a different handedness cohort is, therefore, necessary.
In this study, we explored whether different organizational efficiency (e.g., global efficiency, local efficiency, nodal efficiency) of functional brain network vary with brain size and handedness. The resting-state functional magnetic resonance imaging (fMRI) was employed to construct the binarized functional network for each participant and the graph theoretical method was used to calculate topological parameters to investigate their associations with brain size and handedness. We found that the brain functional efficiency was varied with brain size both in terms of local efficiency (C and E loc) and in global efficiency (Lp, λ, and E glo) and nodes located in language- and emotion-related regions were changed in left-handers. These results tentatively suggest that the variation of handedness and brain size provide new insights into the understanding of the brain functioning.
Materials and Methods
Participants
Thirty-two right-handers (15 females; mean age±SD: 25.87±1.89 years; years of education: 18.44±1.63 years) and 21 left-handers (nine females; mean age±SD: 24.00±2.59 years; years of education: 16.95±2.56 years) were included in this study. These participants were partially from our previous study (Li et al., 2014). All participants showed absence of psychiatric and neurological illness, which were confirmed using the Structured Clinical Interview for DSM-IV, Non-patient Edition. They were specifically recruited and came primarily from the student population at Tianjin Medical University. Handedness was assessed according to the criterion of Chinese revised version of the Edinburgh Handedness Inventory (Oldfield, 1971) (Supplementary Table S1; Supplementary Data are available online at
Image acquisition and data preprocessing
Experiments were performed on a Signa HDx 3.0 Tesla MR scanner (General Electric, Milwaukee, WI) in Tianjin Medical University General Hospital. Sagittal three-dimensional (3D) T1-weighted images were acquired by a brain volume (BRAVO) sequence (repetition time [TR]=8.1 msec; echo time [TE]=3.1 msec; flip angle [FA]=13°; field of view [FOV]=256×256 mm; in-plane matrix=256×256; slice thickness=1 mm, no gap; 176 slices). Resting-state images were collected by an echo planar imaging sequence (TR=2000 msec; TE=30 msec; FA=90°; FOV=220×220 mm; in-plane matrix=64×64; slices: 38; thickness=3 mm, gap=1 mm). The subjects were instructed simply to rest with their eyes closed, not to think of anything in particular, stay awake, and minimize head movement. For each subject the resting-state scanning lasted 6 min 20 sec with dummy scanning 10 volumes, thus collecting 180 volumes.
Preprocessing of the T1-weighted images first involved manual anterior–posterior commissure realignment. Second, all images were segmented into grey matter (GM), white matter (WM), cerebrospinal fluid (CSF) using the VBM8 toolbox (Ashburner and Friston, 2000). Brain size for each participant was then obtained from the total volumes of GM, WM, and CSF.
All fMRI preprocessing steps were performed with the SPM8 (
Functional connectivity networks construction
To determine the nodes of brain functional connectivity networks, we employed the automated anatomical labeling (AAL) atlas (Tzourio-Mazoyer et al., 2002) to divide the whole cerebral cortex into 90 anatomical regions of interest (ROIs).
For each subject, the representative time series of each ROI was obtained by averaging the fMRI time series across all voxels within that region. By calculating the Pearson correlation coefficients between the processed time series of every pair of ROIs, a temporal correlation matrix (90×90) was obtained for each subject. Considering the ambiguous biological interpretations of negative connections (Fox et al., 2009), we only focused on positive correlations. A Fisher's r-to-z transformation was then applied to the correlation matrices, and then each correlation matrix was thresholded into a binary matrix with a sparsity value. Sparsity value was defined as the ratio of total number of edges in a network to the maximum possible number of edges, ensuring that networks from two comparison groups had the same number of edges. Although there is currently no formal consensus regarding threshold selection, we defined a range of sparsity values from 0.10 to 0.46 at the interval of 0.01, following previous studies (Liao et al., 2013; Zhang et al., 2011a). Specifically, (1) the minimum sparsity was selected to assure that the mean degree of each thresholded network was larger than 2log N, where N is the number of ROIs (2) the maximum sparsity was selected to ensure that the small-worldness of each thresholded network was larger than 1.1.
Network analysis
Graph theoretical analysis was carried out using the Brain Connectivity Toolbox (
Statistical analyses
To determine the relationships between the topological properties (Lp, C, E glo, E loc, λ, γ, and E nodal) and handedness and brain size, a multiple regression analysis was performed, with gender, age, and years of education, as covariates. The network parameters were dependent variables, and the predictor variables were handedness, brain size, and their interaction. A value of p<0.05 was considered significant for all analyses.
Validation analysis
Possible concerns regarding the generalizability of this study were considered. First, although the head motion was matched between the left-handers and right-handers (p=0.4621, t=0.7411), the effect of head motion was evaluated with the scrubbing (data with FD>0.2 mm was discarded) procedure on the preprocessed images to ensure that the results were minimally affected by motion-related artifacts (Power et al., 2012), as increasing studies have reported that head motion could have an influence on the functional connectivity (Van Dijk et al., 2012; Zeng et al., 2014). Second, we recalculated the brain size using the total volumes of GM and WM considering the relative increase of CSF volume in atrophic brains, which might lead to spurious results. Finally, high-resolution parcellation network with 1024 ROIs (512 ROIs for each hemisphere) were constructed considering the variability introduced by fitting different brain sizes to the AAL atlas (Zhang et al., 2011b). Specifically, each node composing the low-resolution AAL template was subdivided into a set of micronodes of approximately the same size across both hemispheres.
Results
Small-world functional networks
Global and local efficiency of brain functional networks were compared with the same parameters estimated in random networks over the sparsity range. As expected, efficiency as a function of cost in all networks was monotonically increased; the random graph had higher global efficiency and lower local efficiency than the handedness graph (Supplementary Fig. S1), providing evidence for the small-worldness of functional networks in right- and left-handed subjects.
Handedness effect on brain size
We didn't find any statistically significant difference in the total brain size (p=0.860) or other tissue volumes (GM: p=0.386, WM: p=0.976, CSF: p=0.504) between left-handers and right-handers, which was consistent with one prior study on a large sample of 112 subjects (Herve et al., 2006).
Relationships between global parameters and handedness/brain size
We didn't find any main effect or interaction on the normalized clustering coefficient γ. However, we found the brain size effect on other topological parameters.
C and E loc
The multiple regression analysis revealed significant negative correlations, between C and brain size (t =−3.14, p=0.003), and between E loc and brain size (t=−3.28, p=0.001) (Fig. 1A).

The relationships between fitted network properties and brain size.
Lp, λ, and E glo
The multiple regression analysis also showed significant negative correlations between Lp and brain size (t=−2.51, p=0.015), between λ and brain size (t=−2.57, p=0.013), and positive correlation between E glo and brain size (t=2.87, p=0.007) (Fig. 1B).
Reproducibility of the results
Further analysis of the scrubbed data (about 6.1% images were removed across all subjects) revealed that the majority of the results remained largely reproducible (Supplementary Fig. S2). Repeat analysis using the GM and WM volumes as brain size has demonstrated unchanged results (Supplementary Fig. S3). Not all significant, but similar trend (smaller brains had higher local efficiency, whereas larger brains had higher global efficiency) was also predicted with the high-resolution AAL atlas (Supplementary Fig. S4).
Relationship between nodal efficiency and handedness
Compared with right-handers, the left-handers revealed significant changed nodal efficiency in the left middle temporal gyrus (MTG.L), left anterior cingulate gyri (ACC.L), left median cingulate gyri (MCC.L), right angular gyrus (ANG.R), and right amygdala (AMYG.R). Significant negative correlations between the brain size and nodal efficiency were discovered in the right superior occipital gyrus (SOG.R), right interior tempora gyrus, left parahippocampal gyrus, and left thalamus(THA.L). No significant positive correlation was found. Compared with right-handers, the left-handers showed increased correlations between nodal efficiency and brain size in the MTG.L, ACC.L, and MCC.L, and decreased correlations in the ANG.R, AMYG.R, and right temporal pole: superior temporal gyrus (Fig. 2; Table 1).
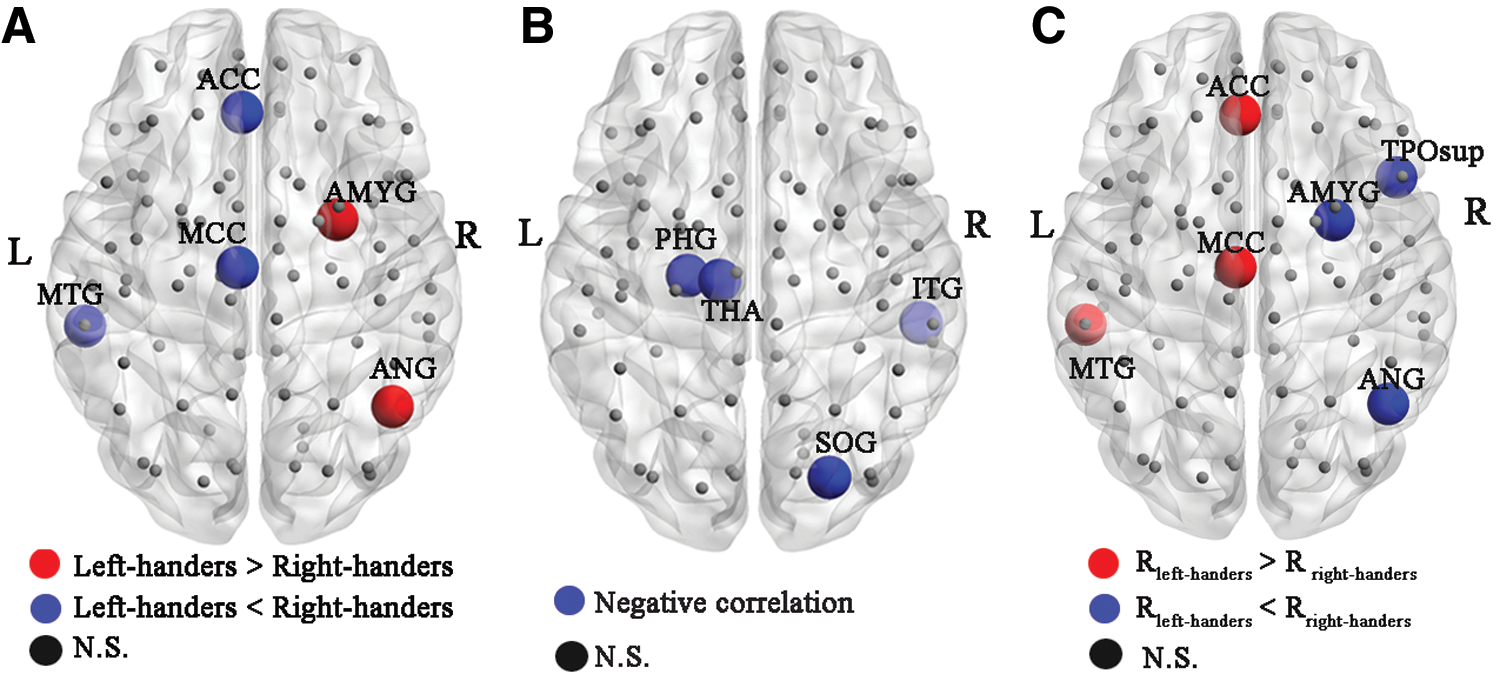
Handedness and brain size effects on nodal efficiency of functional networks.
Handedness and Brain Size Effects on Regional Nodal Efficiency of Functional Networks
R left-handers indicates the relationship between nodal efficiency and brain size in the left-handed group; R right-handers presents the relationship between nodal efficiency and brain size in the right-handed group.
L, left; R, right; MTG.L, left middle temporal gyrus; ACC.L, left anterior cingulate gyri; MCC.L, left median cingulate gyri; ANG.R, right angular gyrus; AMYG.R, right amygdale; SOG.R, right superior occipital gyrus; ITG.R, right interior temporal gyrus; PHG.L, left parahippocampal gyrus; THA.L, left thalamus; TPOsup.R, right temporal pole: superior temporal gyrus.
Discussion
Despite different topological metrics of the brain networks, we observed compatible results for the correlations between local efficiency (C, E loc) and brain size, and correlations between global efficiency (Lp, λ, E glo) and brain size. As for the correlations between nodal efficiency of functional networks and handedness/brain size, we found changes in the left-handers, including the ACC, MCC, MTG, ANG, and AMYG.
Brain size effect on local efficiency of cortical functional networks
Brain networks with higher clustering coefficient and higher local efficiency are robust in local information processing even if some neurons are inefficient or suffer attack (Zhao et al., 2008). In this study, the brain size was shown to be significantly and negatively correlated with C and E loc (Fig. 1A), suggesting that smaller human brain networks had more local functional interconnections, and thus would be more efficient for local information transmission. Cerebral GM contains neuronal cell bodies, dendrites, and short protrusions that are crucial for regional information processing. Consistently, several studies (Im et al., 2008; Leonard et al., 2008) have indicated that people with smaller brains have relatively higher proportions of GM. Therefore, a high percentage of GM corresponds to a large proportion of tissue available for local information processing, which provides further evidence for our finding of higher local network efficiency in smaller brains. Similarly, a DTI study (Yan et al., 2011) has reported that brain size is significantly and negatively correlated with the local clustering coefficient of structural cortical network.
Brain size effect on global efficiency of cortical functional networks
Brain networks with shorter characteristic path length and higher global efficiency are significant in minimizing noise, shortening signal delay, and increasing synchrony (Kaiser and Hilgetag, 2004). In this study, we demonstrated that the brain size was significantly and negatively correlated with Lp and λ, and positively correlated with E glo (Fig. 1B), implying that larger human brains would be more efficient in global information transfer. Im and colleagues (2008) found that WM, rather than GM, would be favored in larger brains, which showed increased number of neurons and glial cells, to support cortical connectivity (Larsen et al., 2006). Additionally, global efficiency has been shown to be affected by the number of long-range connections (Latora and Marchiori, 2001). The disruption/loss of long-range communication among parts of the brain would lead to the decrease of global efficiency of human brain network (Wang et al., 2009). In general, larger brains may need more brain regions concurrently so as to achieve comparable input information, resulting in more extensive connections between distant cortical areas. Consequently, we speculate that greater global efficiency and shorter path length of connections are required to ensure the concurrent packets of information exchange (Latora and Marchiori, 2001), since information is delayed by the increasing time constraints of transcallosal conduction when brain size increased.
Together, higher global efficiency in larger brains, and higher local efficiency in smaller brains might suggest a perfect balance between local necessities and wide-scope interactions as brain size increased. Simultaneously, it further proved that brain networks might have been evolved to maximize the cost efficiency of parallel information processing (Sporns and Zwi, 2004), corresponding to an optimal small-world network organization.
The validation analysis suggests a reproducibility of our findings. However, the repeated high-resolution analysis predicted the same results with a not-very-significant statistical level, which may be caused by the limited samples or other unknown factors. Larger dataset is needed to confirm this exploratory study in the future.
Handedness and brain size effects on nodal efficiency of cortical functional networks
Previous studies have reported that in right-handers, the left hemisphere is dominant for language, but in left-handers, there is a more variable lateralization of language (Haberling et al., 2011). It is clear that the language-relevant cortices are mainly focused on the Broca's area, Wernicke's area, as well as parts of the MTG and ANG. Therefore, the decreased role in the MTG and increased role in the ANG in the left-handers may support the slight rightward speech laterality in left-handers. In addition, the ACC has been found to be strongly associated with emotions and mood regulation (Mayberg et al., 2002). Besides, amygdala hyperactivity has been widely reported to perturb the effective circuit that underlies the overprocessing of negative emotion, which will make someone more sensitive to negative mood (Frick et al., 2012). Therefore, the increased nodal efficiency in the AMYG and decreased efficiency in the ACC suggested irregular emotion processing and regulation in left-handers, which was consistent with that left-handers were more anxious when they responded in a new situation (Wright and Hardie, 2012). Furthermore, the findings provided implications for the understanding of higher ratios of non-right-handers in variant mental diseases (Dane and Balci, 2007; Dragovic and Hammond, 2005), which may be associated with the irregular emotion regulation of left-handers. Further analyses are imperative to investigate whether atypical handedness was associated with emotion dysregulation and illness risk.
Brain size also had significant effects on the nodal efficiency of functional networks. In primates, the THA afferents to the primary visual cortex (Godement et al., 1979) and object recognition takes place in a pathway known as the ventral visual object recognition stream (Ungerleider et al., 1998). Recently, Rilling (2006) indicated that the primary visual cortex was much smaller than the allometric predictions of human brain size. As such, Yan and colleagues (2011) reported that smaller brains had higher nodal centrality in the occipital lobe. Accordingly, our findings about the visual processing (the SOG, THA) were coincided with previous studies. We also found significant interactions between handedness and brain size on nodal efficiency, which had a large overlap with those showing handedness-related differences in nodal efficiency.
A few limitations should be noted. First, our study was limited by a relatively small sample size, larger imaging datasets will be necessary to confirm our preliminary findings. Second, we constructed the functional network excluding the cerebellum; on the other hand, the brain size included the cerebellum. Therefore, the results should not be overextended without careful examinations. Third, more justifiable evidences such as the neuronal activity are needed to better investigate the relationships between the information processing efficiency and brain size and handedness, most of the detailed information are already lost when just considering the BOLD signal acquired by fMRI. Furthermore, using the resting-state fMRI, we were unable to completely eliminate the effects of physiological noises such as cardiac and respiratory cycle with a relatively low sampling rate (TR=2 sec). A more rigorous approach should be applied to remove such noises. Finally, 1 mm slice gap is not optimal for fMRI studies, although it will minimize the interference effects between slices, the missing data in reconstruction may exclude brain regions that are of interest and increase the statistical significance errors (Vaden et al., 2012).
Conclusions
In summary, we found that larger brains possess increased global efficiency and smaller brains possess increased local efficiency, suggesting a perfect organizational principle and regulatory mechanism of human brain. Furthermore, irregular nodal efficiency in different brain regions was revealed in the left-handers. Our results provided novel insights into the understanding of relationships between functional network organization and brain size and handedness. Future studies may clarify whether the altered topological organization in handedness cohort is correlated with specific brain disorders.
Footnotes
Acknowledgments
This work was supported by grants from 973 Project (2012CB517901), the Natural Science Foundation of China (61035006), the Specialized Research Fund for the Doctoral Program of Higher Education of China (20120185110028), the Scholarship Award for Excellent Doctoral Student granted by the Ministry of Education (A03003023901010) and the Fundamental Research Funds for the Central Universities (ZYGX2013Z004).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.