
Abstract
This analysis examines the neuronal foundation of drug-induced psychomimetic symptoms by relating the severity of these symptoms to changes in functional connectivity for a range of different psychoactive compounds with varying degrees of psychomimetic effects. The repeated measures design included 323 resting-state functional magnetic resonance imaging time series and measures of subjective effects in 36 healthy male volunteers. Four different pharmacological challenges with ethanol, morphine, Δ9-tetrahydrocannabinol, and ketamine (12 subjects per drug) were applied. A set of 10 “template” resting-state networks was used to determine individual connectivity maps. Linear regression was used for each individual subject to relate these connectivity maps to three clusters of drug-induced subjective psychomimetic effects (“perception,” “relaxation,” and “dysphoria”) as measured with visual analogue scales. Group analysis showed that the subjective effects of perception correlated significantly across drugs with the connectivity of the posterior cingulate cortex and precentral gyrus with the sensorimotor network (p < 0.005, corrected). No significant correlations were found for relaxation or dysphoria. The posterior cingulate cortex has a role in visuospatial evaluation and the precentral gyrus has been associated with auditory hallucinations. Both the posterior cingulate cortex and the precentral gyrus show changes in activation in patients with schizophrenia, which can be related to the severity of positive symptoms (i.e., hallucinations and delusions), and have previously been related to changes induced by psychoactive drugs. The similarity of functional connectivity changes for drug-induced psychomimetic effects and symptoms of psychosis provides further support for the use of pharmacological challenges with psychomimetic drugs as models for psychosis.
Introduction
Pharmacological challenges constitute a useful instrument in clinical pharmacology research. A pharmacological challenge consists of the administration of a drug with a known pharmacology followed by the (repeated) measurement of quantifiable effects. The relationship between the administered drug and the measured effects provides functional information about the targeted system.
Several drugs have been used to elicit psychomimetic symptoms in healthy volunteers and patients (reviewed in detail by Gouzoulis-Mayfrank et al., 1998). Psychomimetic symptoms are changes in subjective feelings or experiences that bear some resemblance to psychotic symptoms. For example, ketamine (e.g., Abel et al., 2003; Gouzoulis-Mayfrank et al., 2005; Krystal et al., 1994) and Δ9-tetrahydrocannabinol (THC; e.g., D'Souza et al., 2004; Kleinloog et al., 2012; Liem-Moolenaar et al., 2010) have been examined as models for psychosis. The basis for these models is the resemblance between the drug-elicited psychomimetic symptoms and the symptoms of psychosis and schizophrenia (in particular the so-called “positive” symptoms, such as hallucinations and delusions).
The visual analogue scale is sensitive to drug effects and often used to measure psychomimetic effects. It typically consists of a 100-mm-long line with two subjective states on the extremes (e.g., “drowsy” and “alert” or “not at all high” to “extremely high”). To provide an integrated measure of subjective effects, multiple scales are combined in a composite scale of a certain subjective state.
Functional magnetic resonance imaging (fMRI) of functional brain connectivity networks constitutes an important tool in understanding the physiology, pathophysiology, and pharmacology of the brain (Anticevic et al., 2013). Different groups have confirmed the existence of resting-state connectivity networks (also referred to as intrinsic connectivity networks) and their spatial consistency in resting conditions (e.g., Beckmann et al., 2005; Biswal et al., 1995, 2010; Damoiseaux et al., 2006; Smith et al., 2009; van de Ven et al., 2004). The spatial distribution of these networks is related to functional domains (Smith et al., 2009). Changes in connectivity can be detected following administration of drugs that act on the central nervous system (Cole et al., 2013; Khalili-Mahani et al., 2012; Klumpers et al., 2012; Niesters et al., 2012).
Driesen et al. (2013) have used fMRI of intrinsic connectivity networks to study the neuronal foundation of psychomimetic effects of ketamine. They found a positive relationship between psychomimetic effects, and connectivity in several regions implicated in sensory processing. In this study, we apply resting-state fMRI with four different pharmacological challenges to study common effects that are shared between functional brain connectivity and psychomimetic symptoms. Furthermore, the identified brain areas are compared to the regions associated with functional connectivity changes in psychosis, to investigate whether these drug-induced psychomimetic symptoms can serve as a model for psychosis.
Many studies on functional connectivity have focused only on the “default-mode network.” Important nodes of this network are located in the midline areas of the posterior cingulate cortex and within the medial prefrontal cortex (Holt et al., 2011; Raichle et al., 2001). The default-mode network is characterized by a decrease in the tonic resting-state activity during the execution of tasks and is attributed a large evolutionary significance (Raichle et al., 2001). Within this default-mode network, the posterior cingulate cortex serves an adaptive function and regulates incoming information from internal and external environments, whereas the medial prefrontal cortex plays a role in social cognition and emotional processing (Broyd et al., 2009).
Several studies have shown an altered connectivity of the default-mode network in patients with schizophrenia, related to positive symptoms (Bluhm et al., 2007; Garrity et al., 2007; van Lutterveld et al., 2014; Orliac et al., 2013; Tang et al., 2013; Whitfield-Gabrieli et al., 2009). Following the assumed relationship between drug-elicited psychomimetic symptoms and symptoms of psychosis and schizophrenia, we hypothesized that the connectivity within (structures associated with) the default-mode network would be related to the psychomimetic effects. Of the structures within the default-mode network, the posterior cingulate cortex in particular was expected to show such a relationship (Bluhm et al., 2007; Broyd et al., 2009; Holt et al., 2011; van Lutterveld et al., 2014).
Material and Methods
Data sets
The current analysis used data previously collected in three studies within the same centre. Khalili-Mahani et al. (2012) compared the effect of pseudo-steady state levels of ethanol (∼600 mg/L during 2.5 h) and morphine (∼80 nmol/L during 2.5 h) in 12 healthy male volunteers. Klumpers et al. (2012) studied the effect of three subsequent doses of THC (2, 6, and 6 mg with 90 min intervals) in 12 healthy volunteers. Niesters et al. (2012) compared the effect of an increasing concentration of S(+)-ketamine (20 mg/70 kg/h during 1 h followed by 40 mg/70 kg/h for another hour) in 12 healthy male volunteers. These dosing regimens were considered sufficient to induce clinically relevant psychomimetic effects. There were no significant differences in age or body–mass index.
All studies were placebo controlled and included repeated measurements (including baseline measurements) of resting-state fMRI, visual analogue scales for psychedelic effects (Bowdle et al., 1998), and visual analogue scales for alertness, calmness, and mood (Bond and Lader, 1974). From these visual analogue scales, three profile scores (“perception,” “relaxation” and “dysphoria”) were calculated to describe the subjective psychomimetic effects (Kleinloog et al., 2014). Perception consisted of the items “thoughts,” “high,” and “time”; relaxation consisted of the items “mentally slow,” “dreamy,” “drowsy,” and “muzzy”; and dysphoria consisted of the items “suspicious,” “meaning,” and “voices.” For each time point, the profile score was calculated as the mean of the items assigned to a certain profile (each item ranged from 0 to 100 mm).
Imaging protocol
All resting-state images from the three studies were collected on a 3T Philips Achieva scanner with the same sequence parameters (whole brain volume obtained with a gradient echo, echo planar imaging sequence with: repetition time—TR-2180 msec; echo time—TE-30 msec; 80° flip angle; 220 volumes; 3.44 mm isotropic voxels; and a matrix size of 64 × 64 × 38). A T1-weighted high-resolution structural image (TR 9.7 msec; TE 4.6 msec; 8° flip angle; 2-mm isotropic voxels, and a matrix size of 256 × 256 × 140) was acquired at the start of each session and used for registration purposes. Lighting was dimmed and subjects were instructed to lie still with their eyes open and to stay awake.
Software
All imaging data were preprocessed and analyzed in FSL version 5.0.4 (Oxford Centre for Functional Magnetic Resonance Imaging of the Brain [FMRIB] Software Library; Jenkinson et al., 2012;
Preprocessing
Imaging data were preprocessed as previously described by Filippini et al. (2009). For spatial smoothing, a Gaussian kernel of full-width at half maximum of 6 mm and a high-pass temporal filter of 150 sec (0.007 Hz) were used to reduce noise. Resting-state images were registered to the individual's T1-weighted high-resolution structural scan. This structural scan was registered to standard space (MNI-152; Montreal Neurological Institute, nonlinear sixth generation) using a 12 degree of freedom linear search followed by nonlinear registration with a warp resolution of 10 mm. Registration parameters were combined to put the resting-state fMRI scans in MNI standard space, interpolated at a resolution of 2 × 2 × 2 mm.
Dual regression
Dual regression (Beckmann et al., 2009; Cole et al., 2013; Filippini et al., 2009; Khalili-Mahani et al., 2012, 2013) was used with the 10 “template” connectivity networks described by Smith et al. (2009) to derive the spatial and temporal definition of each template network within each scan session. The 10 networks are “medial visual network,” “occipital visual network,” “lateral visual network,” “default-mode network,” “cerebellum network,” “sensorimotor network,” “auditory network,” “executive control network,” “right frontoparietal network,” and “left frontoparietal network.”
Dual regression consists of two consecutive linear regression models. In the first step, the 10 template networks (and 10 additional nuisance networks described by Smith et al., 2009 to control for artefacts) are used in a spatial regression model to extract the session-specific mean time course of each network. These 20 individualized template time courses are then used in a temporal regression model to describe the time course for each voxel as a linear, weighted combination of these template time courses. This results in three-dimensional maps (one per network) containing the regression coefficient (the relative contribution) of each voxel to that specific network.
No additional corrections for physiological functions (heart rate and respiration) were performed. Changes in physiological functions are directly related to the pharmacodynamics of the drug challenges and are often concentration dependent. Template-based dual regression analysis has previously been found to be robust to physiologic noise (Khalili-Mahani et al., 2013).
Subject-level analysis
Following dual regression, a voxel-wise comparison throughout the entire brain was performed for the 10 template networks per subject to determine the linear relationship between the relative change on the subjective measure (clusters perception, relaxation and dysphoria) and the change in connectivity with each network, using the FSL function “randomize.” Baseline measurements were included in this analysis.
Group-level analysis
The resulting estimates were analyzed per network and subjective effect. For the group-level analysis, a one sample group-mean test with Monte Carlo permutation testing of 10,000 random sign-flips of individual estimates was used (as implemented in randomize, see Nichols and Holmes, 2002). A corrected p-value smaller than 0.005 (corrected for 10 networks of interest), following threshold-free cluster enhancement (Smith and Nichols, 2009), was considered statistically significant.
Extracting information on structural location and function
The location of each identified cluster was described using the Harvard–Oxford cortical and subcortical structural atlases (Desikan et al., 2006).
Results
Data sets
Data from a total of 36 healthy male volunteers were included in the analysis. For each treatment (ethanol, morphine, THC and ketamine), data from 12 subjects were available for evaluation. The subjects who received ethanol and morphine were the same (both were included in the data set by Khalili-Mahani et al., 2012). Due to differences in study designs, the study days for ketamine treatment included five scan sessions, the study days for ethanol and morphine included seven scan sessions, and the study days for THC included eight scan sessions. Data from one scan session of one subject receiving ketamine were missing due to adverse events. The total number of scan sessions used for evaluation was therefore 323.
Subjective effects
An overview of the responses on the visual analogue scales is presented in Table 1. The items within the relaxation profile are bidirectional (two different subjective states at the extremes). Subjects were instructed that a normal resting condition would be somewhere halfway between the two extreme subjective states, in line with the original publication of the scales (Norris, 1971), which means the theoretical zero point for these scales is at 50 mm. In contrast, the items within the perception and dysphoria profiles are unidirectional (one subjective state, ranging from “not at all” to “extremely”), resulting in a theoretical zero point of 0 mm. Administration of ketamine resulted in the largest change in all profiles of subjective effects. The induced changes that were seen were smallest for ethanol, followed by morphine, and substantially larger for THC.
Overview of Maximum Subjective Effects as Measured Using Visual Analogue Scales (Mean ± SD)
Statistically significant difference from placebo (p < 0.05 in paired t-test).
Cluster of correlation
The connectivity between one cluster (MNI coordinates maximum (6, −28, 46); size 456 mm3; primarily posterior cingulate and precentral gyrus) and the sensorimotor network correlated significantly with the subjective effects of perception across all treatments (Fig. 1). As shown in the upper panel of Figure 1, the spatial location of the cluster remained constrained to the posterior cingulate gyrus and the precentral gyrus, also when a more liberal p-value of 0.05 was used as threshold as shown in the panel below.

Overview of significant cluster within MNI-152 standard space (blue area: sensorimotor connectivity network; red-yellow area describes cluster). MNI, Montreal Neurological Institute.
For the subjective effects of relaxation and dysphoria, no clusters with significant correlation were identified. Figure 2 presents an overview of the average correlation between the sensorimotor network connectivity and subjective effects within the significant cluster, divided by treatment. The figure also includes the nonsignificant relationships with relaxation and dysphoria, to illustrate the specificity of the association for perception.
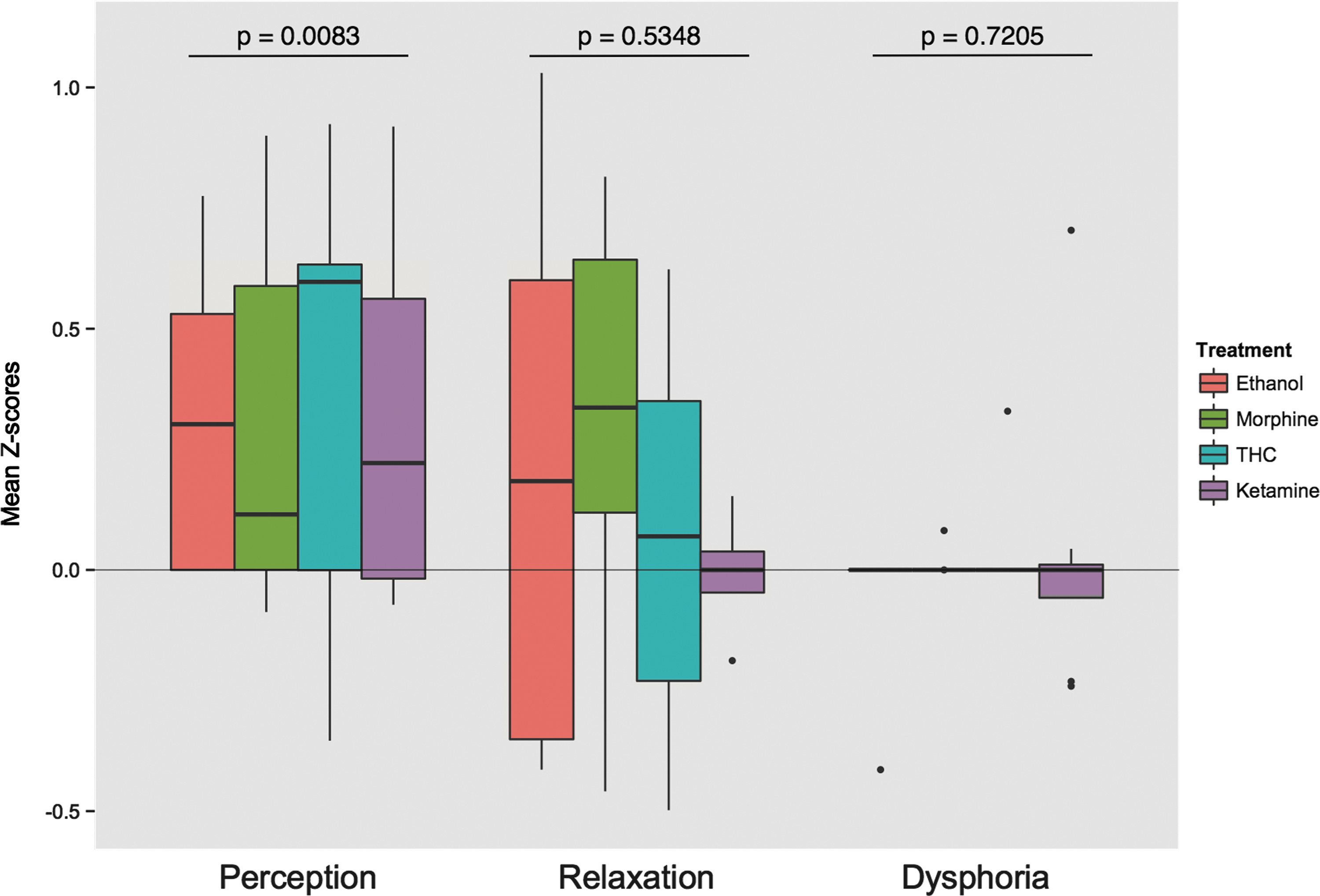
Average correlation of connectivity between the sensorimotor network and subjective effects within the identified cluster, divided by treatment. (Boxes represent the 25th and 75th percentile, the band inside the box, the median, and the whiskers represent the highest/lowest value within 1.5 times the interquartile range; dots represent outliers; p-values are based on a Wilcoxon signed-rank test).
Discussion
A positive correlation, for four different drugs, was found between changes in the functional connectivity and specific subjective psychomimetic effects. Across compounds, the connectivity between a cluster located in the posterior cingulate cortex and the precentral gyrus and sensorimotor network was associated with changes in perception.
We hypothesized that the subjective effects would be associated with changes in connectivity within the default-mode network. Although the posterior cingulate cortex has a role in the default-mode network (Garrity et al., 2007; van Lutterveld et al., 2014; Orliac et al., 2013; Tang et al., 2013; Whitfield-Gabrieli et al., 2009), the relationship between perception and the posterior cingulate cortex in the current analysis was seen within the sensorimotor network. This might be explained because the current analysis examined 10 different networks at the same time, including internetwork connectivity, whereas the previous studies specifically examined the default-mode network and did not look at other connectivity networks.
The cingulate cortex plays a role in the integration and processing of sensory, motor, cognitive, and emotional information (Bush et al., 2000). The identified cluster is located in the area of the cingulate cortex that is functionally described as the visuospatial evaluative region and is involved in the evaluation of the relevance of visual and auditory stimuli (Vogt et al., 1992).
The main functional role of the precentral gyrus is in motor function, but it has also been associated with auditory hallucinations (Diederen et al., 2010; Jardri et al., 2011). Intrinsic connectivity within the precentral gyrus was previously associated with psychomimetic effects of ketamine, which the authors hypothesized to be a result of abnormal sensory processing (Driesen et al., 2013). It is therefore not surprising that these areas are related to psychomimetic changes in perception, which are the result of disruptions in the evaluation of external stimuli. The current analysis demonstrates that these changes are shared by four different pharmacological challenges. Administration of ketamine was previously associated with an increased activation in the precentral gyrus (Daumann et al., 2010; Driesen et al., 2013).
There is also a relationship between the identified brain area and aspects of psychoactive drug use that were not included in the current analysis. Salomon et al. (2012) describe a relationship between life-time ecstasy exposure and functional connectivity in the precentral gyrus. The use of cocaine (Kübler et al., 2005) and administration of psilocybin (Gouzoulis-Mayfrank et al., 1999) have been associated with decreased activation of both the posterior cingulate cortex and the precentral gyrus during cognitive tasks.
The strength of the current study is the combination of drugs with different mechanisms, which results in an outcome that is unrelated to the direct mechanism of action and based on functional changes that are shared by these drugs. A weakness in this approach is that the absence of drug-induced changes in a specific area for one of the drugs would reduce the chance of identifying this change in this combined analysis of all drugs. Of the four drugs used in this analysis, ketamine had the strongest psychomimetic effects, but the relationships were also found for drugs with less pronounced effects (Fig. 2).
The possible influence of changes in physiologic parameters, such as heart rate and respiration, has not been included in the analysis model. Although correction for physiologic parameters is being performed more often, changes in these parameters are related to the pharmacodynamics of the pharmacological challenges and these should therefore not necessarily be seen as artifactual nuisance (Iacovella and Hasson, 2011). In fact, the incorrect use of nuisance regression, might have unintended negative effects (Hallquist et al., 2013). For example, heart rate has been shown to be one of the most sensitive biomarkers for THC effect and there is a strong correlation between an increase in heart rate and THC concentration (Zuurman et al., 2009). Therefore, correcting for heart rate could indirectly mean a correction for drug concentration.
Furthermore, we used template-based dual-regression analysis, which has previously been found to be robust to physiologic noise in resting-state connectivity following a pharmacological challenge (Khalili-Mahani et al., 2013). Other factors, such as system noise and scanner drift could influence the BOLD signal as well. In part, these nuisance factors should have been removed by the high-pass filtering and inclusion of nuisance networks, but some noise may remain. Including global signals in white matter and/or cerebrospinal fluid as a regressor could be an alternative strategy. This approach is believed to include both correction for physiologic parameters and other potential influential factors (Birn, 2012; Weissenbacher et al., 2009).
In a post hoc analysis, where white matter and cerebrospinal fluid signals were included in the dual regression (using FMRIB's tissue probability templates for white matter and cerebrospinal fluid with >90% probability), the identified association between the functional connectivity and subjective effects was blunted and no longer met criteria for statistical significance.
For the subjective effects of relaxation and dysphoria, no clusters were identified that had a significant relationship with changes in connectivity consistent for all four drugs. This can, in part, be explained by the different characteristics of the included drugs, which had various pharmacological activities and effect profiles. Any shared effects on the relaxation profile, which includes various aspects of sedation and anxiolysis, may result from different mechanisms of central nervous system depression.
This also seems to be the case for dysphoria, which describes a range of poorly specified negative emotional states, although it is still conceivable that drug-induced negative mood effects share common neuronal networks. Such changes may not have been detected in our study because the effect of dysphoria occurred in only a limited number of subjects (Kleinloog et al., 2014), which may have prevented the detection of significant clusters for the whole group. These hypotheses are consistent with the distributions presented in Figure 2.
The analytic approach used in the current analysis places a strong emphasis on the template connectivity networks described by Smith et al. (2009). The use of this standardized set of template networks improves the interpretation of the findings and makes the results more easily comparable with other studies. On the other hand, the use of the dual-regression approach allows for individual differences in exact spatial and temporal definitions of the connectivity networks. It would have been too rigid to directly apply the template networks to the data (without dual regression). Although the template connectivity networks are based on a model-free approach, the use of this standard set of networks introduces a framework for the intrinsic functional architecture of the brain.
The measurement of resting-state functional connectivity has been shown to be highly consistent over repeated measurements (Shehzad et al., 2009; Zuo et al., 2010) without intervention. It is therefore likely that changes seen following a pharmacological challenge are the result of the administered drug.
To accommodate the use of repeated measurements in the analysis, the different measures of connectivity within each subject (for a given treatment) were related to the subjective effects. This has the additional benefit that individual differences in “anchor” points on the visual analogue scale (the range between feeling “not at all high” and “extremely high” can be different for different subjects) are accounted for. In other words, the relative changes in connectivity are related to the relative changes in subjective effect. However, the use of linear regression to compare functional connectivity and subjective effects assumes a linear relationship, which is not necessarily the case.
As mentioned in the introduction, the psychomimetic symptoms that are elicited by the different challenge agents were hypothesized to be related to symptoms of psychosis and schizophrenia. The identified cluster is part of the areas within the cingulate cortex and the precentral gyrus where functional connectivity was previously associated with the positive symptoms of schizophrenia (Choi et al., 2005; Garrity et al., 2007; Silbersweig et al. 1995).
Olney et al. (1999) propose a glutamate receptor hypofunction model of schizophrenia, which attributes the occurrence of psychotic symptoms to specific neurons within the posterior cingulate. Hyperstimulation of these neurons, for example, by administration of a drug (i.e., ketamine), was associated with a limited and reversible psychomimetic reaction (Olney et al., 1999).
There are similarities between drug-induced psychomimetic symptoms and psychosis, but also differences (Gouzoulis-Mayfrank et al., 1998). Models using psychoactive drugs to mimic symptoms of psychosis do not model the full spectrum of psychosis and in particular do not model the complex syndrome of schizophrenia.
The four drugs (ethanol, morphine, THC, and ketamine) included in this analysis, all have a different mechanism of action. The psychoactive effects of these drugs are clearly not identical, although there are similarities among them. The rationale for grouping these specific drugs was partly pragmatic, as they had already been used in resting-state fMRI studies in our centre and the data of these studies were available. The measurement of psychomimetic symptoms with the visual analogue scales used in the current analysis is not validated to measure psychotic symptoms in psychiatric illness, and it is possible that drug-induced psychomimetic symptoms have a different neurophysiological origin than symptoms of psychosis. Nevertheless, the current findings are compatible with the notion that drug-induced psychomimetic effects and positive symptoms of psychosis may share an underlying relationship with functional connectivity in networks that involve the posterior cingulate and precentral gyrus.
Conclusions
The intrinsic functional architecture of the brain can be described using connectivity networks. Within these networks, a relationship was found between drug-induced changes in functional connectivity in the posterior cingulate cortex and the precentral gyrus and drug-induced changes in subjective psychomimetic effects following pharmacologic challenges with four psychoactive drugs (ethanol, morphine, THC, and ketamine). Previous studies in patients with schizophrenia have shown a relationship between the functional connectivity and positive symptoms of schizophrenia in the same area. Our findings provide further support for the use of pharmacological challenges with psychomimetic drugs as model for psychosis and for the use of resting-state fMRI as a method to study the effects of drugs and diseases in the brain.
Footnotes
Acknowledgment
This research is supported by grants provided by the Netherlands Organisation for Scientific Research (NWO).
Author Disclosure Statement
No competing financial interests exist.