
Abstract
Graph analysis of electroencephalography (EEG) has previously revealed developmental increases in connectivity between distant brain areas and a decrease in randomness and increased integration in the brain network with concurrent increased modularity. Comparisons of graph parameters across age groups, however, may be confounded with network degree distributions. In this study, we analyzed graph parameters from minimum spanning tree (MST) graphs and compared their developmental trajectories to those of graph parameters based on full graphs published previously. MST graphs are constructed by selecting only the strongest available connections avoiding loops, resulting in a backbone graph that is thought to reflect the major qualitative properties of the network, while allowing a better comparison across age groups by avoiding the degree of distribution confound. EEG was recorded in a large (n = 1500) population-based sample aged 5–71 years. Connectivity was assessed using phase lag index to reduce effects of volume conduction. Connectivity in the MST graph increased significantly from childhood to adolescence, continuing to grow nonsignificantly into adulthood, and decreasing significantly about 57 years of age. Leaf number, degree, degree correlation, and maximum centrality from the MST graph indicated a pattern of increased integration and decreased randomness from childhood into early adulthood. The observed development in network topology suggested that maturation at the neuronal level is aimed to increase connectivity as well as increase integration of the brain network. We confirm that brain network connectivity shows quantitative changes across the life span and additionally demonstrate parallel qualitative changes in the connectivity pattern.
Introduction
The brain is a complex network of highly connected brain areas under constant pressure for optimal performance. Describing the brain network using graph theoretical parameters has proven useful providing biomarkers for disease (Heuvel et al., 2010; Menon, 2011; Stam et al., 2009; Tijms et al., 2013; Zhao et al., 2012). In addition, it provides a theoretical underpinning for what might constitute optimal performance in an optimal network organization (Bullmore and Sporns, 2009; de Haan et al., 2009; Sporns, 2014; Stam, 2014). Ontological development may show similar pressure for increased optimal organization. Anatomically, the human brain shows developmental changes on the whole-brain scale (Casey et al., 2000; Courchesne et al., 2000; Giedd et al., 1999; Lenroot and Giedd, 2006; Paus et al., 2001; Westlye et al., 2010), on the intermediate scale of distinct brain areas (Gogtay et al., 2004; Lenroot and Giedd, 2006; Paus, 2005; Shaw et al., 2006), and also on the neuronal microscale (Huttenlocher, 1979; Huttenlocher and Dabholkar, 1997; Huttenlocher and de Courten, 1987).
These anatomical changes are accompanied by changes on a functional level, as measured using functional magnetic resonance imaging (fMRI), MEG, and electroencephalography (EEG) (M/EEG). The resting-state networks seem largely in place by the age of two (Fransson et al., 2010), but also show clear development by increasing (long-distance) connectivity as evidenced both from fMRI (Fair et al., 2008, 2009; Power et al., 2010) and M/EEG studies (Courchesne et al., 2000; Hanlon et al., 1999; Smit et al., 2012). Comparing young to older adults, modularity decreases for longer connectivity distances and across networks (Meunier et al., 2009), and in aging, connectivity decreases in strength (Smit et al., 2012).
Functional methods of determining connectivity use either direct (EEG, MEG) or indirect (fMRI blood-oxygen-level dependent [BOLD]) measures of correlated neuronal activity to derive coupling strength between brain areas. The high temporal resolution of M/EEG may be particularly useful for estimating short duration networks that appear and disappear on second scale (“fragile binding”). On a larger temporal scale, fMRI can also detect connectivity, as illustrated most clearly by the resting-state networks (Damoiseaux et al., 2006). Recent work has shown that both the fMRI- and M/EEG-based resting-state activity share a common ground (Britz et al., 2010; Mantini et al., 2007; Musso et al., 2010). Our previous investigations showed that connectivity showed substantial change over time closely following anatomical developmental curves of white matter (Boersma et al., 2010; Smit et al., 2010, 2012). Moreover, connectivity correlated with white matter volume (WMV) (Smit et al., 2012). When connectivity matrices from EEG were converted to graphs and analyzed following Watts and Strogatz (Watts and Strogatz, 1998), global network efficiency showed similar correlations with white matter and protracted development from childhood into young adulthood (Smit et al., 2012).
M/EEG recordings are subject to volume conduction effects that blur the recorded signals at the scalp or sensor level. Volume conduction is particularly problematic for determining functional connectivity between signals for algorithms like Coherence and synchronization likelihood (SL) (Nunez and Srinivasan, 2006; Nunez et al., 1997). For this reason, Stam and coworkers (2007) proposed the phase lag index (PLI), which was designed to reduce the effect of volume conduction by ignoring zero and π phase differences between pairs of signals. If the distribution of phase differences is symmetric around zero, this may be evidence for spurious connectivity due to volume conduction. Deviances from a symmetric distribution must be due to dependency between sources (direct or indirect). Flat distributions show no evidence for connectivity, spurious or not. Our first aim is to establish whether average connectivity, as well as the graphs derived from connectivity matrices, still show the strong developmental effects that we have reported earlier (Boersma et al., 2010, 2013; Smit et al., 2010, 2012).
A second limitation of previous studies may be that the comparison of networks across the different age groups is problematic, as networks have different average connectivity and degree (Tewarie et al., 2015; van Wijk et al., 2010). Although the use of graph parameters compared to those of randomized graphs is often thought to remove much of these comparability problems, this may only partly be the case (van Wijk et al., 2010). The use of the minimum spanning tree (MST) graph might provide additional information over the use of thresholded or weighted graphs (Boersma et al., 2013; Stam, 2014; Stam et al., 2014). MST graphs are connected graphs constructed from weighted, undirected connectivity matrices in such a way that they are fully connected and do not form loops. The Kruskal algorithm for finding the MST iteratively selects the edge with lowest weights (where weight is defined as “distance,” the inverse of “connection strength”) and adds the connection to the spanning tree only if no loops are formed. The algorithm stops when all vertices are connected (Kruskal, 1956).
Tewarie and coworkers (2015) showed using simulations that—even though tree graphs are biologically implausible—MST graph properties still capture topological features of the underlying full network, such as small-worldness and randomness of the graph. Kim and colleagues (2004) showed that spanning trees (like the MST) capture a large proportion of the betweenness centrality (BC) (e.g., ∼50% of BC was captured in a coauthorship network with only 16% of the nodes), supporting the idea that they capture most of the information flow in the network. The tree thus forms a “backbone” structure of the original graph. In this article, it is our aim to capture properties of this backbone structure, while keeping comparability across different age groups.
Graph parameters derived from MST graphs include the number of leaf nodes (nodes with degree 1; leaf number [LN]), diameter (DI), the tendency for preferential attachment (degree correlation [DC]), and measures that reflect the importance of the maximally central node (maximum degree Kmax, maximum eigencentrality [ECmax], and BCmax). Table 1 gives an overview of the measures. It has been argued that optimal network function is a tradeoff between a small diameter (i.e., the MST network reveals that the underlying brain network is compact), but not dependent on a single hub node (which has a very small diameter, but may be less resilient (Albert et al., 2000; Stam, 2014; Stam et al., 2014).
Minimum Spanning Tree Graph Parameters and Their Description
MST, minimum spanning tree.
Increased integration of the network may manifest itself as a move from low to high LN in MST graphs. How MST graphs relate to optimization of the brain is a matter of ongoing investigation (Lee et al., 2006; Serrano et al., 2009; Stam, 2014; Stam et al., 2014). However, it has been shown that MST parameters are altered in Parkinson's disease, epilepsy, Alzheimer's disease, and near brain tumors (Çiftçi, 2011; Dubbelink et al., 2014; Lee et al., 2006; Tewarie et al., 2014; van Dellen et al., 2014); for an overview, see Stam (2014).
In sum, we investigated whether the observed increase in network order remains after connectivity has been established with a measure less sensitive to volume conduction compared to the SL measure used previously (Smit et al., 2012; Stam and van Dijk, 2002). Next, we investigated whether the MST graphs from PLI connectivity networks provide a similar picture of increased integration with maturation. By reanalyzing the same dataset as previously (Smit et al., 2012), and by using a different connectivity measure and different graph type, we can make a comparison without the burden of sample fluctuation. Finally, while our previous analyses focused on a narrow age range in childhood (5 and 7 year olds) (Boersma et al., 2013), in this study, we provide data on subjects aged 5–71 from multiple large longitudinal EEG datasets covering adolescence and adulthood. Using these data we will establish whether the previously reported increase in MST order in childhood continues to adolescence and is maintained during adulthood.
Methods
Subjects and procedure
Data were collected as part of a study into the genetics of brain development and cognition. A total number of 1675 individuals (twins and additional siblings) accepted an invitation for extensive EEG measurement. For the present analyses, EEG data recorded during 3–4 min of eyes-closed rest were available from six measurement waves with ages centered approximately around 5, 7, 16, 18, 25, and 50 years. Table 2 shows the number of subjects broken down by wave and level of genetic overlap (twin zygosity/siblings). Note that the twin/family relatedness is not a part of the current, non-genetic analyses, but high degree of genetic overlap between subjects will result in a reduction of effective degrees of freedom. Part of the measurements consisted of longitudinal measurements at two ages (5–7 and 16–18 years). In addition, some of the subjects aged 16–18 were invited back for measurements at age 25. In total, this study incorporated 2453 EEG recordings. After data cleaning, 2206 recordings were available. The structure of the final subject set after data cleaning used in the present study was 362, 377, 425, 386, 359, and 297 respectively for the six measurement waves, which included 328 longitudinal observations between 5 and 7, 385 between 16 and 18, 103 between 16 and 25, 100 between 18 and 25.
Subject Count and Sample Overlap Between Measurement Waves
Numbers are valid subjects after EEG data cleaning. EEG data were collected in six separate waves, four with narrow age range (5–18), and two with wider age ranges (25–50). Waves 5 & 7 have complete overlap. Waves 16 & 18 have complete overlap. One hundred three subjects in adolescent waves also appear in wave 25. All others are independent observations.
DZ, dizogotic; MZ, monozygotic; EEG, electroencephalography.
Ethical permission was obtained by the “subcommissie voor de ethiek van het mensgebonden onderzoek” of the Academisch Ziekenhuis VU (currently METc of the VU University Medical Centre). All subjects (and parents/guardians for subjects under 18) were informed about the nature of the research. All subjects or parents/guardians were invited by letter to participate, and agreement to participate was obtained in writing. All subjects were treated in accordance with the Declaration of Helsinki.
EEG acquisition
The childhood and adolescent EEG were recorded with tin electrodes in an ElectroCap connected to a Nihon Kohden PV-441A polygraph with time constant 5 sec (corresponding to a 0.03 Hz high-pass filter) and low pass of 35 Hz, digitized at 250 Hz using an in-house built 12-bit A/D converter board, and stored for offline analysis. Leads were Fp1, Fp2, F7, F3, F4, F8, C3, C4, T5, P3, P4, T6, O1, O2, and bipolar horizontal and vertical electrooculogram (EOG) derivations. Electrode impedances were kept below 5 kΩ. Following the recommendation by Pivik and associates (1993), tin earlobe electrodes (A1, A2) were fed to separate high-impedance amplifiers, after which the electrically linked output signals served as reference to the EEG signals. Sine waves of 100 μV were used for calibration of the amplification/AD conversion before measurement of each subject.
Young adult and middle-aged EEG was recorded with Ag/AgCl electrodes mounted in an ElectroCap and registered using an AD amplifier developed by Twente Medical Systems (Enschede, The Netherlands) for 657 subjects and NeuroScan SynAmps 5083 amplifier for 103 subjects. Standard 10-20 positions were F7, F3, F1, Fz, F2, F4, F8, T7, C3, Cz, C4, T8, P7, P3, Pz, P4, P8, O1, and O2. For subjects measured with NeuroScan, Fp1, Fp2, and Oz were also recorded. The vertical EOG was recorded bipolarly between two Ag/AgCl electrodes, affixed one cm below the right eye and one cm above the eyebrow of the right eye. The horizontal EOG was recorded bipolarly between two Ag/AgCl electrodes affixed one cm left from the left eye and one cm right from the right eye. An Ag/AgCl electrode placed on the forehead was used as a ground electrode. Impedances of all EEG electrodes were kept below 5 kΩ, and impedances of the EOG electrodes were kept below 10 kΩ. The EEG was amplified, digitized at 250 Hz, and stored for offline processing.
EEG preprocessing
We selected 12 EEG signals (F7, F3, F4, F8, C3, C4, T5, P3, P4, T6, O1, and O2 and both EOG channels) for further analysis as the set with the most complete match between the different measurement waves/cohorts.
All signals were broadband filtered from 1 to 35 Hz with a zero-phase FIR filter with 6 dB roll-off. Next, we visually inspected the traces and removed bad signals based on absence of signals, excessive noise, extensive clipping, or muscle artifact. Note that for the network analysis, a full set of EEG signals was required, and therefore, any rejected EEG channel resulted in the loss of that subject. Next, we used the extended independent components analysis (ICA) decomposition implemented in EEGLAB (Delorme and Makeig, 2004) to remove artifacts, including eye movements, and blinks. After exclusion of components reflecting artifacts, the EEG signals were filtered into the alpha (6.0–13.0 Hz) frequency band. The peak alpha frequency developed from 8.1 Hz at age 5 to 9.9 Hz at age 18, after which a slow decline to 9.4 Hz was observed at around 50 years. The lower edge of the alpha filter was set such that alpha oscillation of all subjects was included from ∼2.0 Hz below the lowest peak frequency to ∼3.0 Hz above the highest average peak frequency. The cleaned filtered data were cut into sixteen 8-sec epochs.
Connectivity
Connectivity was calculated using the PLI. For a detailed description, we refer the reader to Stam and colleagues (2007). In short, the PLI inspects the distribution of phase differences between pairs of signals
where s(t) is the signal over time t, and H is the Hilbert transform, i is
for Δ φ modulated within the range −π and π.
To compare the results from PLI to a measure that does not take into account the effects of volume conduction, we calculated SL on the same data using the specifications as reported previously (Smit et al., 2012).
Graph analysis
MST graphs were created with the Kruskal algorithm (Kruskal, 1956) applied to the PLI connectivity matrices. Next, we derived parameters described in Table 1 from these graphs using a variety of MATLAB algorithms, including standard MATLAB code, the MIT graph toolbox (
Statistics
The effect of age was determined in several ways. First, we created developmental plots (scatterplots) from connectivity and each of the MST graph parameters on age. Next, local nonlinear-weighted regression trends were fitted (loess, on 65% window size second-order polynomials). 95% confidence intervals around the loess fit were obtained using a bootstrap with 10,000 repeats using percentiles. Since some observations are nested within family, the bootstrap was based on the independent unit (family) rather than individual. Note that the bootstrap was not used to establish significance. To test significance of developmental trends, we estimated different fixed-effect models. First, linear, quadratic, and cubic trends were fitted to the dataset, which we tested for significance sequentially. Because of the complex structure of data, including repeated measures and family dependencies, which even extended across the different age groups (siblings of twins might fall into a different age category than the proband twins), we used generalized estimating equations (GEE) to obtain p values. GEE is a random-effects model that corrects significance of fixed effects under nonindependence of observations, viz., under residual correlation within a known cluster of observations (in the current case, clusters are all observations within a family, including repeated measures). GEE with the “exchangeable” option estimates a single correlation between residuals within the cluster. Since all off-diagonal elements in the residual correlation matrix are estimated to be the same, this naturally handles uneven cluster sizes (including missing data and families of different size). Even though the residual matrix is arguably more complex than the single correlation (e.g., repeated measures and monozygotic twin correlations are expected to be higher than other within-family correlations), the robust standard errors (SEs) are not affected by this misspecification with regard to controlling for type I errors (Minică et al., 2014).
Second, we defined nine age groups using the following boundaries specified in years. The youngest four age groups were specified to match the measurement waves with relatively narrow age ranges (childhood and adolescence): 4.9–6.0, 6.0–7.4, 13.0–16.6, and 16.6–20.0. The adult waves had a larger age range and were split centered around decades 20.0–25.0, 25.0–35.0, 35.0–45.0, 45.0–57.5, and 57.5 and older. The youngest adult age group was chosen so as to not overlap with the late adolescent/young adult wave. The oldest age group's lower boundary was increased to 57.5 to reduce the large age range to 13.5 years. These were tested in an omnibus test using GEE with age group as the factor controlling for sex. To investigate post hoc-specific age group deviations, we also tested equality of means of age groups in a pair-wise manner with false discovery rate (FDR) correction for the n(n − 1)/2 comparisons tested (36 at n = 9) (Benjamini and Hochberg, 1995). In the post hoc comparisons, we further multiplied the p values by a factor of two to accommodate for multiple testing across the measures. Two was chosen rather than eight, since the dimensionality of the data showed a clear two-dimensional structure (see Results section).
Results
SL and PLI are different
SL and PLI were compared both on a lead pair by lead pair basis and on average connectivity. When comparing the 66 lead-pair connectivity scores, SL and PLI correlations ranged from 0.111 for childhood age groups to 0.34 for adolescent age groups. Averaged across the whole scalp, the correlation between SL and PLI ranged from 0.30 to 0.65. This suggests that PLI and SL show overall different patterns of connectivity. The average connectivity pattern for SL and PLI is shown in Figure 1. Compared to PLI, SL showed stronger connectivity for nearby electrodes grouped into three clusters (frontal, central, and posterior). This may partly reflect volume conduction effects. SL also shows evidence of stronger connectivity across homologues than PLI. These are less likely to reflect volume conduction effects (Nunez et al., 1997) due to their relatively large interelectrode distance. Since PLI connectivity for homologues is relatively low, we can conclude that lateralized alpha sources mainly oscillate in phase. PLI connectivity showed a more evenly distributed pattern of connectivity than SL, but a parietal hub was visible, while the frontal cluster disappeared. In sum, average PLI connectivity taps partly into the same sources of individual variation as SL, however, the connectivity pattern differs for PLI and SL.
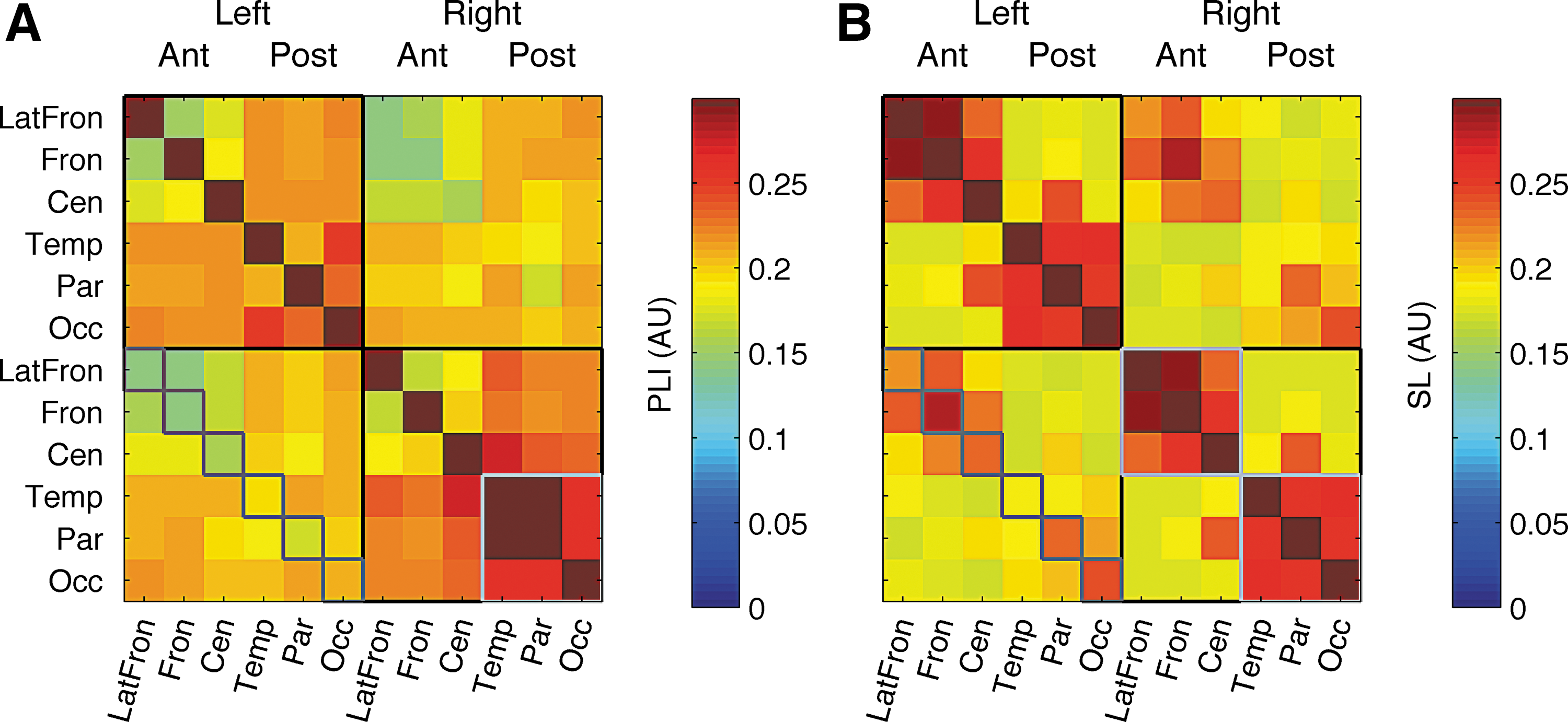
Connectivity matrices for PLI
Increased order with increased LN
Figure 2 shows the dependency of MST graph parameters on LN. Each point in the scatterplot represents the MST graph values of a single individual. Note that most values fall close to the polynomial regression. A second-order polynomial fit was significant in all cases. The centrality measures and Kmax showed a positive relationship with an upward curve as expected from random graph simulations (Boersma et al., 2013). Tree hierarchy (TH) also showed a positive dependence on LN, but with a downward curve, thus setting a limit to the effect of LN on TH. DI decreased with increasing LN in a linear manner, which may be expected since maximum and minimum values for DI may be derived analytically from LN (Stam, 2014). DI will lie between
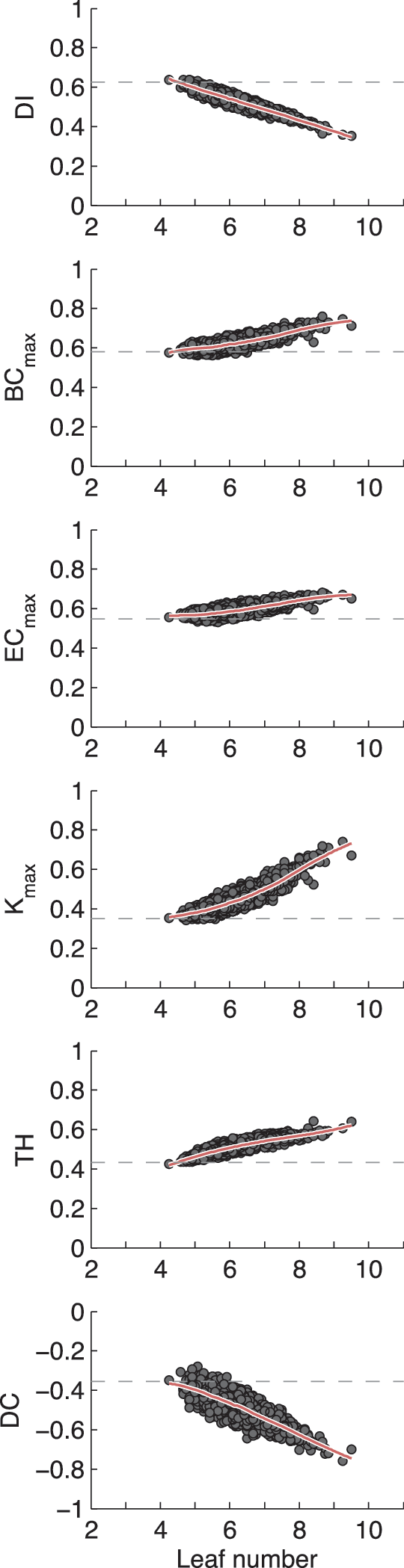
MST graph parameters covary with LN. From left to right on the x-axis, increased LN indicates increased hierarchical order and integration in the network. LN ranges from 2 (a linear configuration) to 11 (a star-like configuration) for a 12-vertex network, expressed in this study as a proportion from 0 to 1. Each plot contains an average MST graph parameter for each individual, plotted against average LN (averaged across multiple EEG epochs). The red line is a loess fit (50% width, second order). The dashed line indicates the average value obtained in 1000 graphs (n = 12) based on random signals. BC, betweenness centrality; DC, degree correlation; DI, diameter; EC, Eigenvector centrality; EEG, electroencephalography; LN, leaf number; MST, minimum spanning tree; TH, tree hierarchy. Color images available online at
PLI connectivity shows an inverted-U development
Figure 3 (top row) shows the results of average connectivity within the MST graph developing over age. Connectivity showed a pattern of development similar to those reported previously based on a different measure of connectivity (Smit et al., 2012). Top-left graph shows the development with loess fit (50% window size, second-order fit). The top-middle graph shows the same loess fit with bootstrap 95% confidence interval and reveals changes from childhood to early adulthood in average PLI, a decrease from 16 to 25. The omnibus test GEE model predicting average connectivity with age group as factor controlling for sex was highly significant, Wald χ 2(8) = 243.3, p = 4.5 · 10−48. Post hoc comparisons with FDR adjustment revealed that significant increases were found from childhood to adolescence, but also between age groups 5 and 7. MST connectivity declined in the 57.5+ age group and reached significance in comparison to the adolescent and ∼40 age groups.
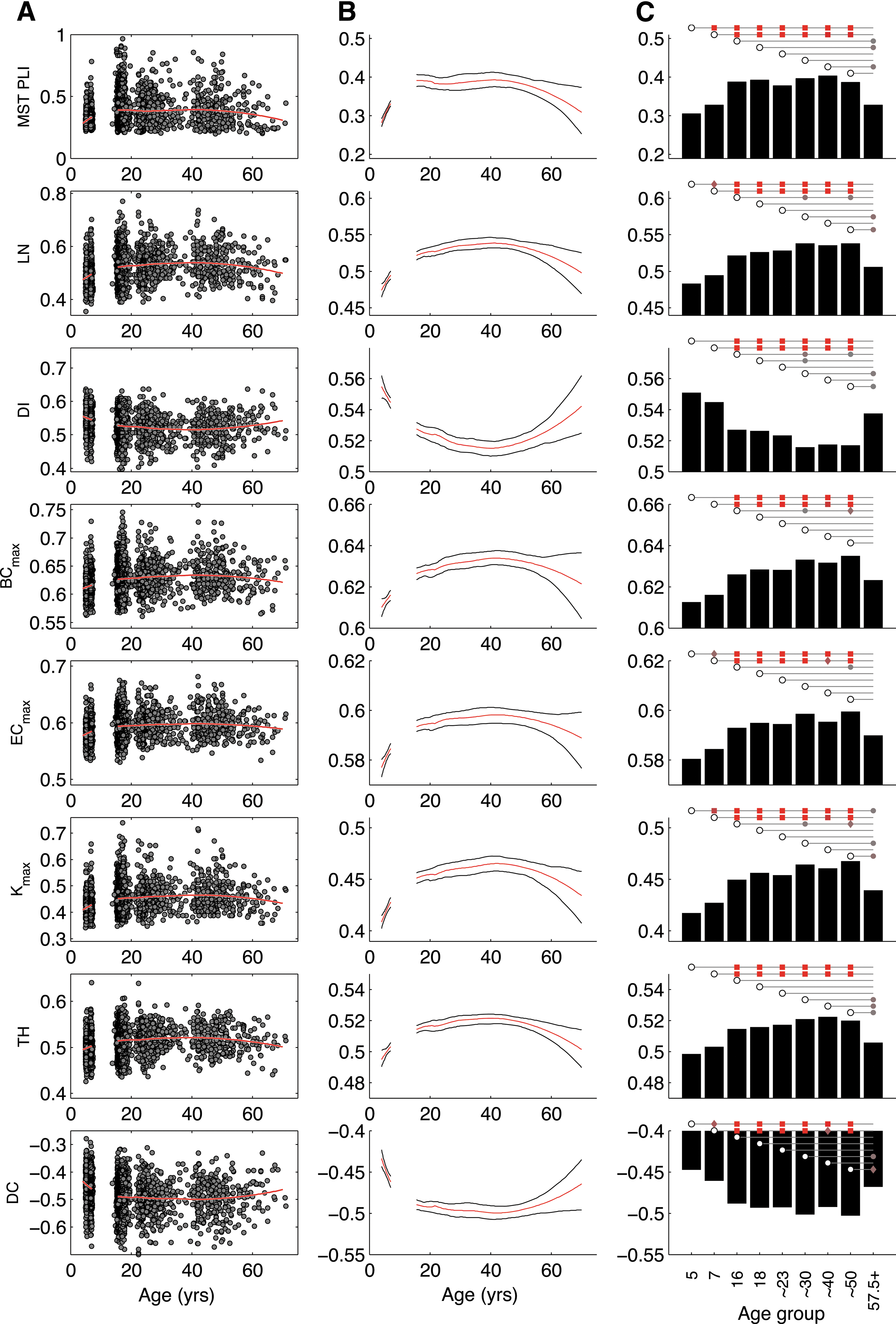
Age development plots for average connectivity strength (PLI) within the MST graph and MST network parameters of EEG alpha oscillations (6.0–13.0 Hz).
Connectivity patterns change with age
Although average PLI connectivity may change across different age groups, localized developmental differences may still occur. We assessed the connectivity between all possible pairs of signals and calculated change across the 9 age groups in a stepwise manner (i.e., 8 change values) expressed as rate per annum. Three-dimensional headplots were constructed using BrainNet Viewer (Xia et al., 2013) with approximate locations of the electrodes (Fig. 4 left column). The thickness of the edges was rescaled to average increase per annum making them comparable across headplots. Red colors indicate increase, blue decrease, and green indicates no change.
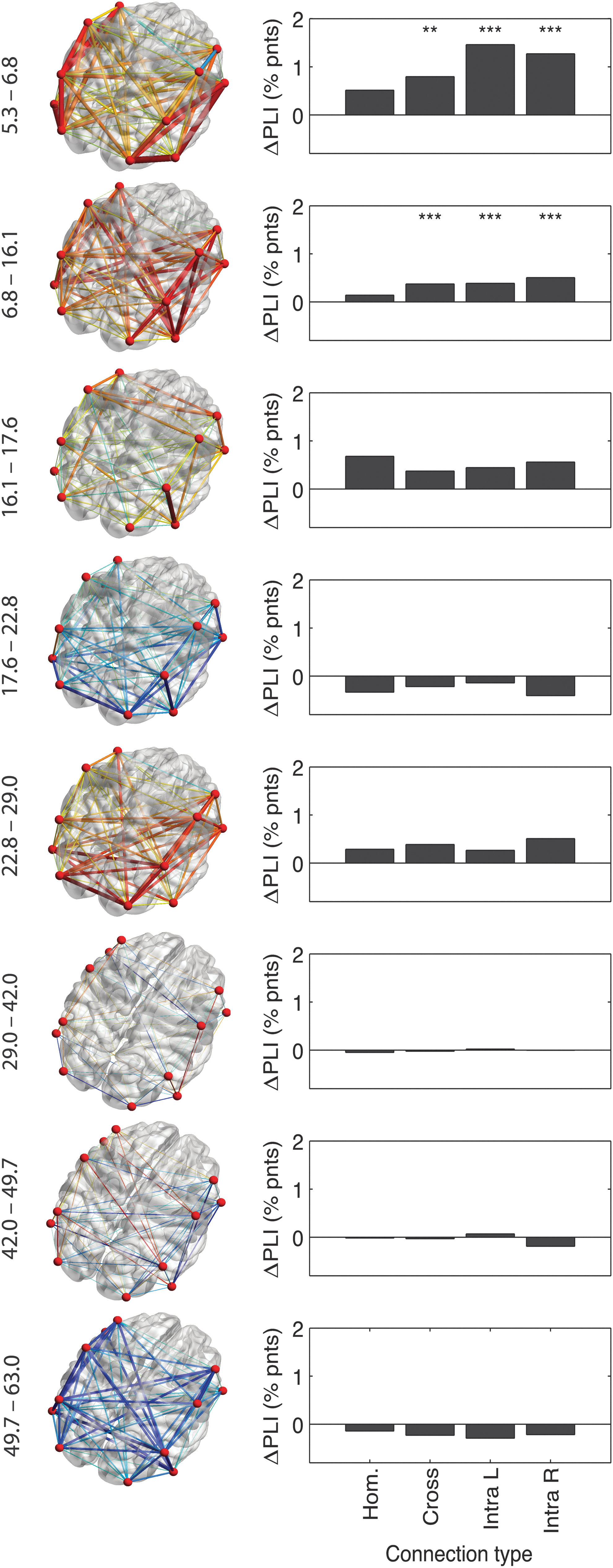
Localized development of connectivity strength. Left: 3D headplots of average change in PLI per year from one age group to the next (age groups 5, 7, 16, 18, ∼23, ∼30, ∼40, ∼50, 57.5+) from an elevated right posterior viewpoint. The location of maximal development is not stable, but changes with age. Actual ages of the age groups that are compared are shown on the left. Right: Separating edges into intrahemispheric left and right (Intra L, Intra R), contralateral homologues (Hom), and other cross-hemisphere (Cross) connections showed that childhood was marked by a clear intrahemispheric increase of connectivity, while later age groups showed no such strong prevalence or stronger increases in contralateral homologues (within adolescence). **p < 0.05, ***p < 0.001 after FDR correction at q = 0.05. Color images available online at
Changes within childhood were largely limited to intrahemispheric connections (Fig. 4 right column). Homologous (left–right lateralized) electrode pairs and other interhemispheric connections showed low PLI connectivity increase. From childhood to adolescence, both inter- and intrahemispheric connections showed increases, but homologues still showed less PLI change. In adolescence, a change is seen with homologues reaching the largest change. In later ages, reduction in connectivity strength is clearest in interhemispheric connections (other than homologues). In sum, the changes during childhood, adolescence, and middle-aged adulthood showed remarkable changes in topology. Clearly, the brain does not simply change connectivity, but changes the overall pattern of connectivity.
Reliability of MST parameters
Split-half reliability across two sets of eight epochs was corrected for the reduced time length in the eight compared to 16 epochs. Very high reliability for scalp-average PLI connectivity was obtained (0.96). Pairwise channel reliability ranged from r = 0.72 to 0.94. MST graph parameters were measured moderately reliably. Highest reliability was found for Kmax (0.741). The other centrality measures showed lower reliability (BCmax: 0.587, ECmax: 0.592). LN, DI, and DC showed reliability of 0.71, 0.628, and 0.605, respectively.
An increasingly integrated network
Figure 3 also shows the development of MST graph parameters as scatterplot with loess fit (Fig. 3A), bootstrap of the loess fit with 95% confidence intervals (Fig. 3B), and pairwise testing of significance across age groups (Fig. 3C). Network parameters showed developmental trends highly comparable to connectivity. Cubic curves were not significant (absolute robust z < 1.4, ns). All quadratic terms were significant (absolute robust z > 6.44, p < 1.2E-10) with all parameters showing inverted-U shapes—except DI showed a U curve as expected.
The brain network of children showed a lower LN, indicating a more random network and less integrated organization. Increasing age resulted in an increased LN and a correspondingly increased BCmax, ECmax, and Kmax. TH changed similarly in an inverted-u shape. DI and DC decreased. These findings are consistent with an increasing star-like organization and increased integration. The comparison of most measures showed significant change from 5 years of age to adolescence/adulthood with highly significant values (p < 0.0001, and p < 0.001 compared to age ∼40 for DC and ECmax). Age 7 showed a similar pattern. Both ages 5 and 7 generally showed no significant difference with the oldest age group (>57.5).
Older age (57.5+) was marked by a significant decrease for many parameters, although the effects were not very strong (p < 0.01). LN and TH decreased with older age compared to ages ∼30 to ∼50. Connectivity decreased only compared to age ∼40 (p < 0.01). The centrality measures showed less-consistent decrease, possibly due to a noisier variation.
Principal components reveal partly separate sources of variation
Since the developmental trends of connectivity and MST graph parameters showed markedly similar paths, we subjected the correlation matrix of four different measures for connectivity (homologous contralateral connectivity, other interhemispheric connectivity, and intrahemispheric connectivity left and right) and six MST-based graph parameters (LN, DI, BCmax, ECmax, Kmax, DC) to an eigenvalue decomposition after selecting one random subject per family and regressing out the effects of age and sex. TH was excluded since it was based on two other parameters and therefore does not add information to the correlation matrix. Scores for DI and DC were inverted so as to enforce positive correlations. Figure 4 shows the results. The correlation matrix shows a clear clustering of connectivity versus MST graph parameters. The highest eigenvalue of 5.85 explained 58.5% of the variance, the second highest was 2.09 (20.9% variation). Both the correlation matrix (Fig. 5A) and the scree plot (Fig. 5C) strongly suggest a two-factor solution. After varimax rotation, MST parameters loaded strongly on the first component and PLI connectivity measures on the second (Fig. 5B). Figure 5D shows that the two components show different developmental patterns, with a much clearer U-curve for MST graph parameters, while PLI connectivity shows a decrease from adolescence to young adulthood. For these reasons, we conclude that MST graph parameters and PLI-based connectivity largely reflect different sources of variation in brain function with different developmental curves.
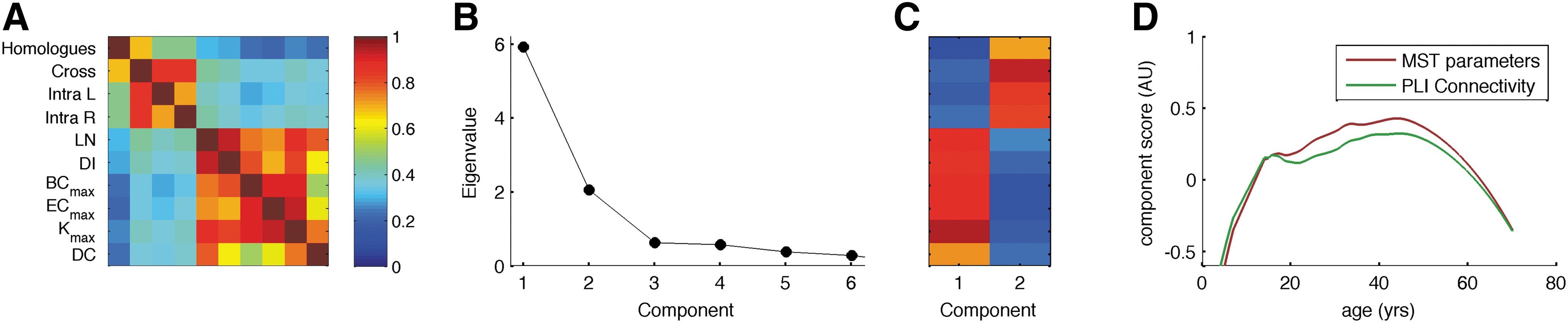
Principal Components Analysis of connectivity scores separated into contralateral homologues (Hom), other cross-hemispheric connections (Cross) intrahemisphere left (Intra L) and intrahemisphere right (Intra R), and MST graph measures. Note that TH was excluded as this measure is fully based on two other graph parameters (LN and BCmax). Positive correlations of DI and DC with other parameters were enforced by reversing scores.
Preservation of properties in MST graphs
Twelve-node graphs are arguably insufficient to detect properties of the underlying network that has spatially much higher dimensionality. We therefore conducted an analysis on 39 datasets with 128-channel 4-min eyes-open resting EEG available in our laboratory. We refer to Smit and associates (2013) for more specifics on data recording. The cleaned data were filtered (8–13 Hz), epoched into sixteen 8-sec epochs, PLI connectivity calculated, MST parameters calculated, and averaged over the epochs. After that, we selected the 12 electrodes nearest to those in the current dataset, and recalculated PLI and MST parameters. The average PLI correlated highly (r = 0.97). Kmax, DC, LN, and DI correlated moderately to highly (r = 0.48, r = 0.40, r = 0.70, and r = 0.61, respectively; all p < 0.011), indicating that a substantial proportion of interindividual variance is reflected in a very rudimentary 12-node representation of the initial 128-node network. BCmax and ECmax correlated nonsignificantly (r = 0.24), suggesting that maximum centrality measures are hardly preserved.
We additionally compared parameters derived from full 12-node graphs to those of MST graphs. Full graphs were thresholded graphs with a fixed average degree (K = 4.5) as calculated previously in this sample (Smit et al., 2012). The connectivity within the MST graphs correlated highly with connectivity in the full graphs (r = 0.93). Kmax correlated r = 0.63, ECmax 0.46, and LN 0.44 between full and MST graphs. This confirms previously reported results that spanning trees conserve important node properties and properties on information flow (Kim et al., 2004; Tewarie et al., 2015). However, it also confirms that the overlap might not be perfect due to the arbitrary threshold selection (van Wijk et al., 2010).
Discussion
Our aim was to investigate whether the increased integration of the network observed from 5 to 7 years of age extends into adolescence and adulthood. The large and highly significant differences found in graph parameters and connectivity between childhood and adolescence/adulthood suggest that this is the case. We established that the life-span development of average connectivity between pairs of scalp-recorded signals closely mimics those reported previously (Smit et al., 2012). By using the PLI (Stam et al., 2007)—a measure that was designed to ignore volume conduction—we have found support that our previous findings using SL (Stam and van Dijk, 2002) have not been spurious. We hypothesize that the sparse electrode layout in our previous report may have been protective against detecting false synchronization (Smit et al., 2012).
Average connectivity within the MST measured with PLI showed strong increases within childhood and from childhood to adolescence. Our previous report on the same sample used SL as a measure of functional brain connectivity. Each measure is sensitive to different types of functional connectivity as evidenced by the different connectivity matrices they produce (Fig. 1). Even so, the current results show remarkable similarities with previous reported results. Both PLI and SL showed a strong increase from childhood to adolescence with effect sizes over r > 0.40 comparing age group 5 with other ages. PLI showed peak value at age 40 (Fig. 3, right column), with SL peaking at around age 50. This suggests that the previous results were quite robust against effects of volume conduction and common reference. However, the current results also differed from those reported previously. Connectivity measured using SL showed continuous and significant increases into late adulthood, whereas PLI connectivity showed a nonsignificant change (and a decrease from age 18 to ∼23). Moreover, the correlations between SL and PLI were generally low to moderate, suggesting that they reflect different sources of variation.
Several findings in the extant literature suggest that this increase in EEG functional connectivity depends on maturation of white brain matter, including myelinization. For example, it has been found that interhemispheric EEG connectivity measured by coherence has been related to diffusion tensor imaging diffusivity (Teipel et al., 2009) and T2 relaxation times in white matter in head injury, which arguably is related to neuronal membrane lesion (Thatcher et al., 1998). In addition, we have previously found that developmental curves for connectivity are highly consistent with the protracted development of white matter development: both connectivity and WMV showed peaks in middle age (Allen et al., 2005; Bartzokis et al., 2001; Benes et al., 1994; Good et al., 2002; Walhovd et al., 2005a, 2005b; Westlye et al., 2010). Moreover, a moderate correlation was found between WMV and connectivity. Because PLI reduces the effects of spurious connectivity in the brain based on volume conduction and common reference, these results seem to further strengthen the idea that functional connectivity in the resting state reflects the strength of anatomical connectivity between distant brain areas. It is not expected that changes in head circumference are a spurious explanation for the observed changes in functional connectivity and resulting changes in the MST network. At the age of 5, the head has reached 90% of its final size (Rollins et al., 2010). Moreover, effects on volume conduction due to head growth are likely to scale with the brain and not produce relative changes in (relative) conductive properties of the brain/skull/scalp. Global changes in volume conduction induced by head circumference increase seem inconsistent with the regional changes in connectivity as revealed in Figure 4. Finally, we have previously found that signal strength (oscillatory power) is larger in childhood than in adulthood (Smit et al., 2012), which would indicate that S/N ratios decrease with age. Reduced S/N should result in noisier PLI estimates and graphs closer to noise, which is the opposite of what was observed. More likely, synaptic pruning and white matter development caused the increase of connectivity, resulting in greater connectivity and complexity even at larger distances. Functionally, brain regions come “closer” together during brain size increase.
Arguably, MST graphs are more comparable across groups than thresholded graphs (Stam, 2014; van Wijk et al., 2010). Graph parameters derived from the MST graph showed evidence for change in the level of integration. All MST parameters show an inverted-U curve (and a U curve for diameter). The backbone graph in the human brain activity moved from a line to a more star-like configuration during development. In later age, a return to a more line-like configuration was found. For all, but the centrality measures, these resulted in significant drops for age group 57.5+ compared to ages 30 and 50. Importantly, principal components analysis showed that MST graph parameters reflected different sources of variation compared to PLI connectivity. Clearly, not just the average connectivity, but the connectivity pattern changes. Note that we observed that MST graph parameters showed a more star-like configuration than random graphs (Fig. 1). In this sense, the observed developmental changes showed a move from random networks toward more integrated networks, and more random networks in later life. This, too, is consistent with previous observations of life-span development in the same sample (Boersma et al., 2010; Smit et al., 2010, 2012).
The current EEG dataset was limited to a small number of EEG signals (n = 12). The signals are linear combinations of the neural generators they project to the scalp location of the electrodes. Although the PLI algorithm disregards spurious connectivity of neural sources that project to multiple electrodes, it is clear that many sources contribute to a signal electrode. Therefore, each signal represents the activity of a large area in proximity of the electrode. We therefore analyzed high-density recordings for PLI and MST parameters, downsampled spatially to 12 channels, and analyzed PLI and MST parameters again. Average connectivity was well represented in the reduced 12-node network. In addition, many MST parameters were also (partially) preserved in the 10-fold reduced networks (notably, Kmax, DC, and LN). Note that this reduction includes an increased measurement error. Therefore, we conclude that the observed developmental changes in 12-node networks suggest that high-density measurements could, likely with much higher power, detect a similar change.
The results make MST graph parameters highly suitable as biomarkers for the development in early life and cognitive decline associated with older age. Follow-up studies could target the genetic variants that have been linked to neuronal change such as myelination. In addition, studies could investigate how genetic variants exert their influence in cognitive decline or Alzheimer's disease (e.g., apolipoprotein E gene [APOE], clusterin gene [CLU/APOJ], and phosphatidylinositol binding clathrin assembly protein [PICALM]) (Harold et al., 2009; Hollingworth et al., 2011; Lambert et al., 2013). Carriers of the APOE
In developmental neurobiology, the dichotomy into long and short projection distances may be essential. In an fMRI study, it was shown that decreased short-range connectivity concurs with increased long-range connectivity. Local connections in a cognitive control network become less diffuse with development from 10 to 22 years of age, which is accompanied by an increased long-distance functional connectivity (Kelly et al., 2009). Similar findings of changes in (long-distance) connectivity have been reported (Dosenbach et al., 2010; Fair et al., 2009; Supekar et al., 2009). The present results extend these findings in showing that from childhood to adulthood, brain networks move from less to more integrated graphs (Fig. 2). Since network parameters may be relevant predictors of cognitive performance (Micheloyannis et al., 2006; Tewarie et al., 2014; van den Heuvel et al., 2009) and are disrupted in neurological disorders (Stam et al., 2009, 2014; Tewarie et al., 2014; van Dellen et al., 2014), we can hypothesize that the increasingly integrated network topology is essential to the large developmental changes in human cognitive performance during the same period. Indeed, a more integrated network was predictive of better cognitive performance in MS patients (Tewarie et al., 2014). Cognitive performance correlated with a larger decrease in network integration in Parkinson's patients (Dubbelink et al., 2014). Whether these findings generalize to the normal population may be addressed in future investigations.
In conclusion, brain connectivity measured by the PLI shows large changes over the lifespan. These changes largely corroborate the earlier findings that the connection strength increases during development (Hagmann et al., 2010; Smit et al., 2010, 2012). Since PLI is less sensitive to volume conduction by ignoring the zero and π phase differences between signal pairs (Stam et al., 2007), developmental changes are therefore unlikely to reflect changes in conductive properties across age groups. The use of the MST backbone graph aimed to solve the problem that graph measures may not be compared across different sizes and degree distributions (van Wijk et al., 2010). However, MST graphs confirmed that brain matures across the lifespan and shows changes in structure both in the development in childhood and during aging in later life. These findings corroborate our earlier findings that the network shows reduced randomness from childhood to young adulthood (Boersma et al., 2010, 2013; Schutte et al., 2013; Smit et al., 2010, 2012).
Footnotes
Acknowledgments
This work was supported by grants Twin-family database for behavior genetics and genomics studies (NWO 480-04-004) to D.B., Genotype/phenotype database for behavior genetics and genetic epidemiological studies (NWO 911-09-032) to D.B., European Research Council (ERC-230374) to D.B., BBR Foundation (NARSAD) Young Investigator grant 21668 to D.S., NWO/MagW VENI-451-08-026 to D.S., VU University VU-USF 96/22 to D.B., Human Frontiers of Science Program RG0154/1998-B to D.B., and E.d.G., Netherlands Organization for Scientific Research, NWO/SPI 56-464-14192 to D.B.
Author Disclosure Statement
No competing financial interests exist.