
Abstract
Menstrual cycle-dependent changes have been reported for a variety of functions, including cognition, attention, emotion, inhibition, and perception. For several of these functions, an effect of hormonal contraceptives has also been discussed. Cognitive, attentional, emotional, inhibitory, and perceptual functions have been linked to distinct intrinsic connectivity networks during the resting state. However, changes in resting-state connectivity across the menstrual cycle phase and due to hormonal contraceptive use have only been investigated in two selected networks and without controlling for the type of hormonal contraceptives. In the present study, we demonstrate menstrual cycle and hormonal contraceptive-dependent changes in several intrinsic connectivity networks, including networks that have been related to emotion processing, olfaction, audition, vision, coordination, and two lateralized frontoparietal networks related to a variety of cognitive functions. These changes parallel behavioral changes in the functions associated with these networks. Changes in connectivity and changes in behavior occur during the same cycle phases. Furthermore, hormonal contraceptive-dependent effects were observed in the same networks and same target sites as menstrual cycle-related changes and were dependent on the androgenicity of the progestin component contained in the hormonal contraceptive.
Introduction
A
For instance, it has been demonstrated in the cognitive domain that verbal, spatial, and memory functions vary across the menstrual cycle with better performance in spatial tasks (particularly mental rotation) during the follicular and better performance in verbal and memory tasks (verbal fluency; verbal memory) during the luteal cycle phase (Dadin et al., 2009; Hampson, 1990a, b; Hausmann et al., 2000; McCormick and Teillon, 2001; Rosenberg and Park, 2002). However, there are inconsistencies in whether the improvement in spatial performance is attributable to reduced hormone levels in the early follicular phase (e.g., Hampson, 1990a) or elevated estradiol levels in the late follicular phase (Berry et al., 1997). For verbal, spatial, and memory functions, OC-dependent effects have also been demonstrated (Pletzer and Kerschbaum, 2014). Exploratory evidence suggests that the use of OCs containing androgenic progestins results in improved mental rotation performance, while the use of OCs containing antiandrogenic progestins results in impaired mental rotation performance (Wharton et al., 2008). While the results on effects on verbal abilities are still inconsistent, an improvement of various memory functions (verbal memory, emotional memory; face recognition) has repeatedly been demonstrated (e.g., Mordecai et al., 2008; Nielsen et al., 2011; Pletzer et al., 2015). However, the majority of these studies did not distinguish between androgenic and antiandrogenic progestins.
Furthermore, both menstrual cycle and OC-dependent effects have been demonstrated in different aspects of attention (Pletzer et al., 2014; Solis-Ortiz and Corsi-Cabrera, 2008; Thimm et al., 2014), and menstrual cycle effects have been discussed for a variety of higher cognitive functions, including working memory (n-back task, Jacobs and D'Esposito, 2011), cognitive control (Stroop task, Hatta & Nagaya, 2009), and inhibition (Stop-Signal task, Colzato et al., 2010). The latter appear to be locked to the preovulatory phase of the menstrual cycle and depend on the dopamine baseline levels of participants (Colzato and Hommel, 2014; Jacobs and D'Esposito, 2011).
A large number of studies have investigated both the effects of menstrual cycle and OC use on emotions (for reviews, see Oinonen and Mazmanian, 2002; Romans et al., 2013). Mood changes have been demonstrated during the luteal phase of the menstrual cycle, sometimes being attributed to elevated progesterone levels, other times, being attributed to the rapid decrease in progesterone and estradiol levels in the late luteal/premenstrual phase. Similarly, hormonal contraceptives appear to have positive effects on mood in some women, but negative effects in other women.
Perceptual changes, for example, in the sense of smell (e.g., Navarrete-Palacios et al., 2003; Watanabe et al., 2002) and appetite (e.g., McVay et al., 2012), have also been observed during the menstrual cycle.
Several of these cognitive functions and emotional or perceptual processes have been related to specific brain networks identified during the resting state through a variety of methods (e.g., Damoiseaux et al., 2006; Yeo et al., 2011). Especially subcortical networks have been related to functions for which relevant menstrual cycle-dependent changes have been reported, such as the Group 1 networks related to emotion and automatic processes identified by Laird and colleagues (2011). In the past, independent component analysis (ICA) has been a very useful tool to identify these networks (Beckmann et al., 2005; Damoiseaux et al., 2006). An extensive behavioral interpretation of resting-state networks obtained with an ICA approach, including their involvement in specific tasks, was given by Laird and colleagues (2011).
While an increasing number of studies try to identify differences in brain structure and function relating to observed cognitive and emotional changes across the menstrual cycle and due to hormonal contraceptive use (Pletzer et al., 2013, 2015; Schoning et al., 2007; Weis et al., 2008), only three previous studies have up to now investigated menstrual cycle and OC-dependent changes in resting-state network connectivity (De Bondt et al., 2015; Hjelmervik et al., 2014; Petersen et al., 2014). All of these studies used a group ICA approach, but focused solely on the default mode network (DMN) and executive control network, and report inconsistent results. While Petersen and colleagues (2014) found increased connectivity of the left angular gyrus within the DMN and increased connectivity of the left middle frontal gyrus within the executive control network during the follicular phase of the menstrual cycle, Hjelmervik and colleagues (2014) and De Bondt and colleagues (2015) were not able to replicate these findings. However, Petersen included only two cycle phases in a between-subjects design and did not distinguish between androgenic and antiandrogenic OC users, while the other two studies included three cycle phases. These are, however, only exemplary results on two of multiple resting-state networks (Beckmann et al., 2005; Damoiseaux et al., 2006) that can be identified during the resting state and linked to behavior and performance in certain tasks (Laird et al., 2011).
Therefore, the present study seeks to provide an explorative, but comprehensive, analysis of menstrual cycle and hormonal contraceptive-dependent changes in several resting-state networks. These resting-state networks have previously been linked to task-based networks related to certain cognitive and emotional behavior as revealed by Laird and colleagues (2011). Like Smith and colleagues (2009), they demonstrate that networks derived through ICA on peak coordinates of task-based fMRI studies archived in the BrainMap database strongly correspond to resting-state networks described by Biswal and colleagues (2010). The authors therefore argue that similar networks can be identified in task-based and resting-state studies. Using hierarchical clustering analysis on BrainMap MetaData, Laird and colleagues (2011) provide a detailed functional interpretation of these networks linking them to specific tasks. Based on their findings, the present study is conceived to relate menstrual cycle and OC effects on resting-state networks to previously reported menstrual cycle and OC effects on performance in certain tasks. We thereby provide an important basis for future research by establishing a direct link between menstrual cycle-dependent behavioral changes and changes in intrinsic connectivity in certain networks. We tested naturally cycling women at three time points during their menstrual cycle (menses; ovulation; mid-luteal) and compared them with two groups of hormonal contraceptive users (androgenic and antiandrogenic).
We hypothesize that menstrual cycle and hormonal contraceptive-dependent changes will be present in those networks supporting functions or tasks for which behavioral changes across the menstrual cycle and due to hormonal contraceptive use have previously been described, including the executive control network, the frontoparietal networks (changes in attention), the limbic system (changes in mood), and some of the perceptual networks.
Materials and Methods
Participants
Eighteen healthy young women (mean age: 26.61 ± 6.07 years) with a natural menstrual cycle (mean cycle duration: 29.08 ± 1.61 days) participated in the study. They were scanned at three time points throughout their menstrual cycle: during the early follicular phase (days 2–8; low estradiol and progesterone; mean cycle day: 4.61 ± 2.85), during the preovulatory phase (2–3 days before ovulation; high estradiol; mean cycle day: 14.22 ± 1.63), and during the mid-luteal phase (4–10 days after ovulation; high estradiol and progesterone; mean cycle day: 22.67 ± 3.29). Ovulation was calculated as lying 14 days before the estimated onset of the next period, and the preovulatory peak in estradiol was estimated through commercial ovulation tests. The order of cycle phases across scanning sessions was randomized.
Additionally, 32 users of OC participated in the study; 16 of them (mean age: 24.56 ± 3.03 years) were classified as users of OC containing androgenic progestins (Levonorgestrel, Gestodene). The remaining 16 (mean age 21.56 ± 2.92 years) were users of OC containing antiandrogenic progestins (Drospirenone, Dienogest, Chlormadinone Acetate). They were scanned during their active pill phase.
MRI data acquisition
Functional images as well as high-resolution structural images were acquired on a Siemens Magnetom TIM Trio 3 Tesla scanner (Siemens Healthcare). A T2*-weighted gradient echo planar (EPI) sequence (whole brain coverage, TE = 30 msec, TR = 2250 msec, flip angle 70°, slice thickness 3.0 mm, matrix 192 × 192, FOV 192 mm, in-plane resolution 2.6 × 2.6 mm) was employed for functional images. Thirty-six transversal slices were taken oriented parallel to the AC-PC line. Participants were instructed to close their eyes, relax, and let their mind flow. Total duration of resting-state scans was 20 min. A T1-weighted 3D MPRAGE sequence (160 sagittal slices, slice thickness = 1.2 mm, TE 2.9 msec, TR 2.3 msec, TI delay 900 msec, FA 0.95°, FOV 256 × 256 mm) was employed for structural images.
Preprocessing of fMRI data
Preprocessing of functional images was performed using SPM8 (
Group ICA
Following preprocessing, group ICA was performed across all participants and sessions using the INCASSO algorithm implemented in the GIFT toolbox (
The group ICA approach implemented in GIFT uses temporal concatenation of single-subject data, followed by an aggregate spatial ICA analysis, and includes a back-reconstruction step to compute individual subject maps and time courses (Calhoun et al., 2009). We used regular back reconstruction, intensity normalization, and expectation maximization. The individual subject maps that entered second-level analysis (see below) were then scaled to z-Scores. For each voxel, the z-score represents the strength of connectivity of this voxel with the rest of the network.
In accordance with previous group ICA approaches (Abou-Elseoud et al., 2010; Crone et al., 2015; Laird et al., 2011), a fixed number of 20 components were extracted. The extracted components 3, 7, 11, and 12 were discarded as artifactual due to predominant activation in peripheral areas, the cerebrospinal fluid, and white matter or high-frequency fluctuations and spikes. To identify the remaining components, spatial correlations with networks described by Laird et al. (2011) were performed using GIFT. The Laird and colleagues (2011) networks were chosen as a reference for two main reasons. First, the Laird and colleagues (2011) networks have been highly recognized and utilized as a reference in a variety of resting-state studies, including previous studies trying to identify menstrual cycle effects in specific resting-state networks, such as the default mode and executive control network (De Bondt et al., 2015; Hjelmervik et al., 2014; Petersen et al., 2014), which maintains comparability between studies. Second, they are based on an ICA approach as utilized in this study. Although they derive their networks from task-based metadata, the authors demonstrate a strong correspondence to networks obtained through ICA during the resting state. Accordingly, the authors are not only able to give a behavioral interpretation of the identified networks but also link them to specific tasks. This allows us to interpret our findings in the light of previous behavioral menstrual cycle studies utilizing these tasks.
Second-level analyses
Second-level analyses on the individual subject maps were performed in SPM8 (
Hormone levels
Hormone levels were assessed in naturally cycling women from saliva samples collected through the passive drool method using DeMediTec ELISAs for 17β-estradiol, progesterone, and testosterone. Hormone levels were not evaluated in OC users due to different binding affinities of different synthetic steroids to the receptors for natural hormones. Unfortunately, nine samples could not be analyzed due to insufficient sample volume.
Results
Hormone levels
Cycle phase of participants was confirmed by the expected variations in hormone levels across the test sessions (Fig. 1). Estradiol levels increased significantly at the time point of ovulation [t(15) = 3.09, p = 0.008] and decrease significantly during the luteal phase [t(10) = 2.66, p = 0.02]. Progesterone levels did not change at ovulation [t(17) = 0.65, p = 0.52], but increased significantly during the luteal phase [t(12) = 3.61, p = 0.008]. Testosterone levels showed a nonsignificant peak around ovulation (both t < 1.31, both p > 0.21).
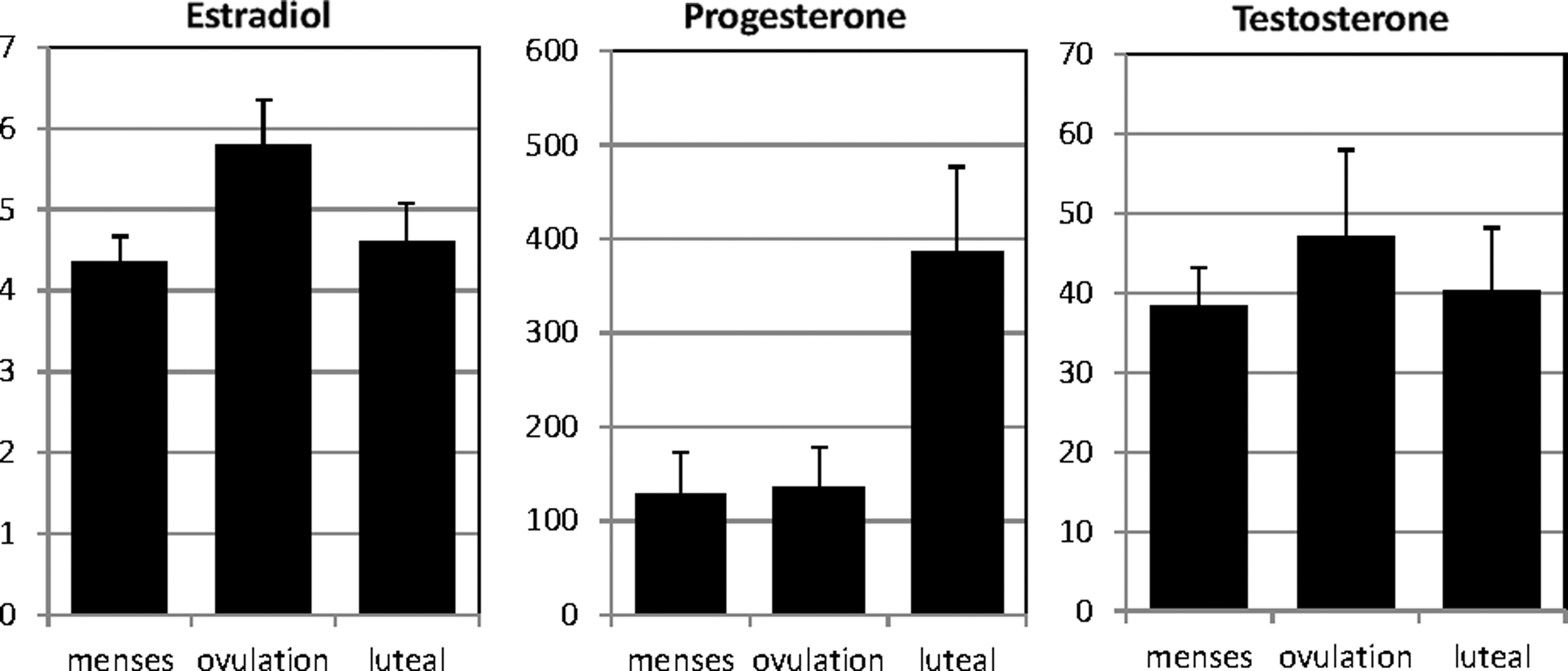
Average hormone levels across cycle phases in naturally cycling women (n = 181). Note: 1Due to insufficient sample volume, hormone values for the luteal phase were only analyzed in 12 subjects. Estradiol and testosterone values for menses and preovulatory phase were each only analyzed in 17 subjects.
Connectivity results
Laird and colleagues (2011) grouped the networks according to the functions they are associated with. The results here are presented in the same order. An overview of all results is summarized in Table 1.
Laird et al. (2011).
Compared with naturally cycling women during menses.
To highlight significant results, non-significant results are shaded in gray.
ACC, anterior cingulate cortex; Cpn, component; r, spatial correlation; ns, no significant differences; OC, oral hormonal contraceptives; OFC, orbitofrontal cortex; ICN, intrinsic connectivity network; BG, basal ganglia; SMA, supplementary motor area; mPFC, medial prefrontal cortex; SPL, superior parietal lobe; DLPFC, dorsolateral prefrontal cortex; R, right; L, left; post, posterior.
Group 1: networks related to emotional and autonomic processes
ICN1
We identified component 20 as corresponding to ICN1 with a spatial correlation of r = 0.50. Laird and colleagues (2011) describe ICN1 as a network including limbic and medial temporal areas. This system appears to be particularly involved in emotion discrimination tasks. Component 20 included the middle temporal gyri, hippocampus, amygdala, basal ganglia, parts of the fusiform and parahippocampal gyri, insula, opercular cortex, olfactory bulbs, and lingual gyri, as well as the cerebellum (Fig. 2A). Menstrual cycle-dependent changes in component 20 are displayed in Figure 2B. In the preovulatory phase, connectivity within component 20 was significantly decreased compared with menses in the basal ganglia ([−12, 12, 2], 128 voxels, T = 4.71, p = 0.02). In the luteal phase, connectivity within component 20 was significantly decreased compared with the preovulatory phase in the precuneus ([12, −78, 40], 236 voxels, T = 6.16, p < 0.001). Hormonal contraceptive-dependent changes in component 20 are displayed in Figure 2C. In users of antiandrogenic, but not androgenic, OCs, a significant increase in connectivity with component 20 compared with naturally cycling women was observed in the thalamus ([14, −18, 0], 105 voxels, T = 5.75, p = 0.05) and left temporal lobe ([−66,−36, 20], 101 voxels, T = 5.19, p = 0.05), while a significant decrease in connectivity with component 20 compared with naturally cycling women was observed in the supplementary motor area (SMA: [−8, 14, 56], 147 voxels, T = 4.30, p = 0.005).
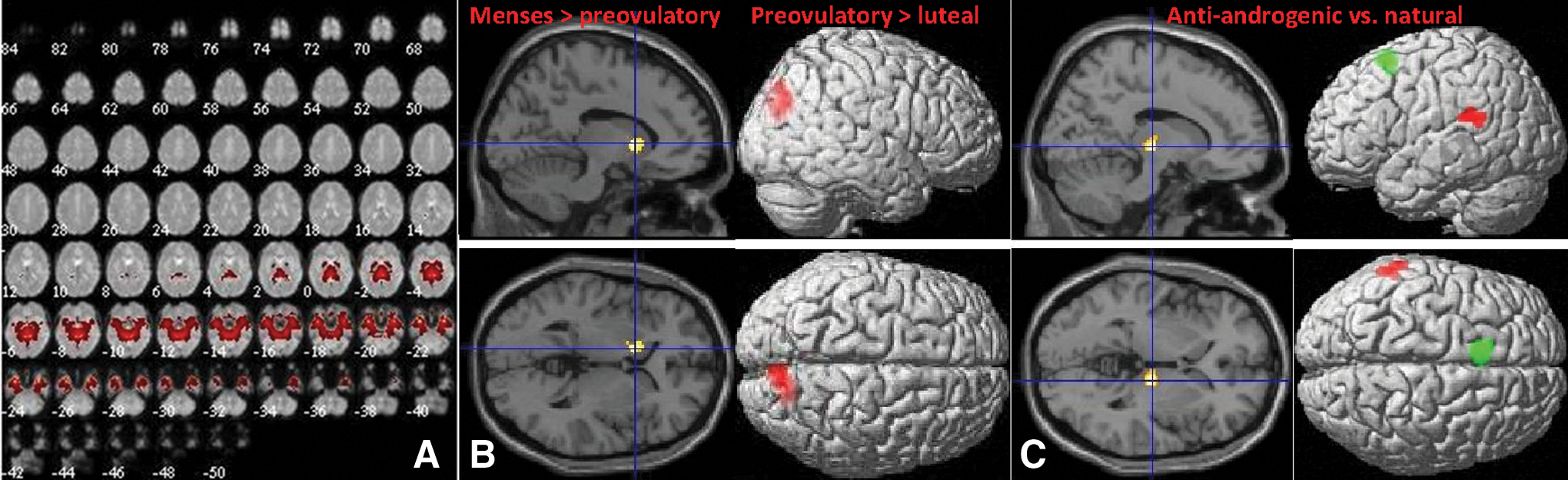
Component 20 (ICN1).
ICN2-3
We identified component 17 as corresponding to ICN3 with a high spatial correlation of r = 0.79. ICN3 was described by Laird and colleagues (2011) as including the bilateral basal ganglia and thalamus. The same component was also the only one, despite artifactual components, to show any correspondence (with a spatial correlation of 0.25) to ICN2. ICN2 was described by Laird and colleagues (2011) as including the anterior cingulate cortex (ACC) and OFC being functionally involved, for example, in olfaction. Both ICN2 and ICN3 are involved in olfactory and reward tasks, for example, in delay discounting (Laird et al., 2011). Accordingly, component 17 bilaterally included the whole thalamus and basal Ganglia, part of the hippocampus and amygdala, but extended toward Heschl's gyrus, the olfactory bulbs, and insula (Fig. 3A). No significant changes between menstrual cycle phases were observed in intrinsic connectivity within component 17, although a nonsignificant decrease in connectivity of the basal ganglia was visible in the luteal compared with the preovulatory phase ([18,6,4], 61 voxels, T = 5.01; Fig. 3B). However, a significant decrease in connectivity of the basal ganglia was observed in users of antiandrogenic OCs compared with naturally cycling women ([−26, −8, 2], 116 voxels, T = 5.07, p = 0.05; Fig. 3C). No connectivity changes within component 17 were observed in users of androgenic OCs.
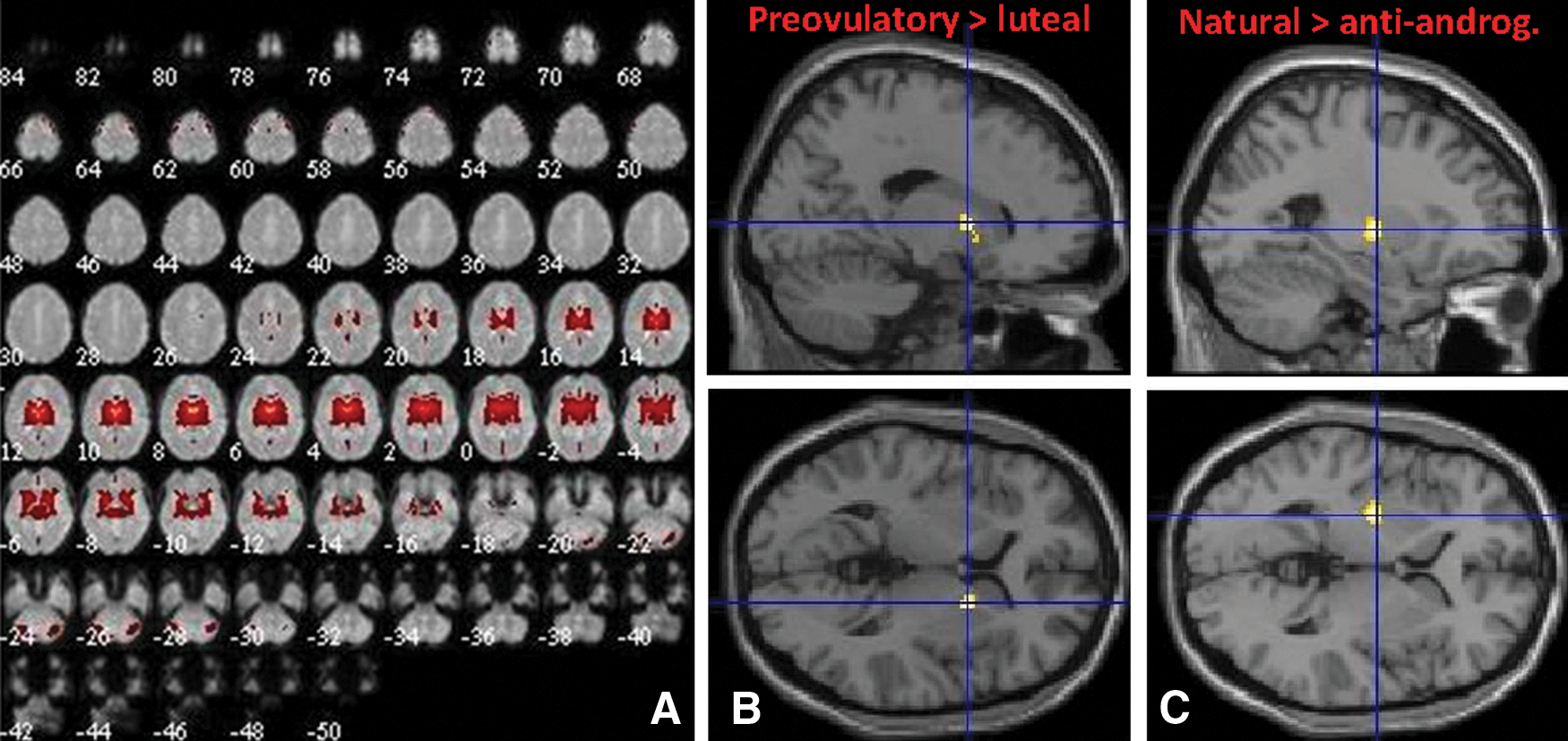
Component 17 (ICN2/3).
ICN4
We identified component 14 as most likely correspondent to ICN4 (r = 0.48), described by Laird (2011) as a network including the insula and cingulate cortex and responsible for linking cognition to emotion. ICN4 is typically linked to inhibitory control tasks such as the Stroop or Go-no-Go task (Laird et al., 2011). This network was chosen by previous menstrual cycle studies as executive control network (e.g., Petersen et al., 2014). The bilateral network included not only the insula, operculum, cingulate cortex, and SMA but also part of the basal ganglia and thalamus (Fig. 4A). No significant changes between menstrual cycle phases were observed in intrinsic connectivity within component 14. However, users of antiandrogenic contraceptives displayed decreased connectivity compared with naturally cycling women in the medial prefrontal cortex (mPFC: [−2, 52, 22], 186 voxels, T = 4.92, p = 0.008; see Fig. 4B).
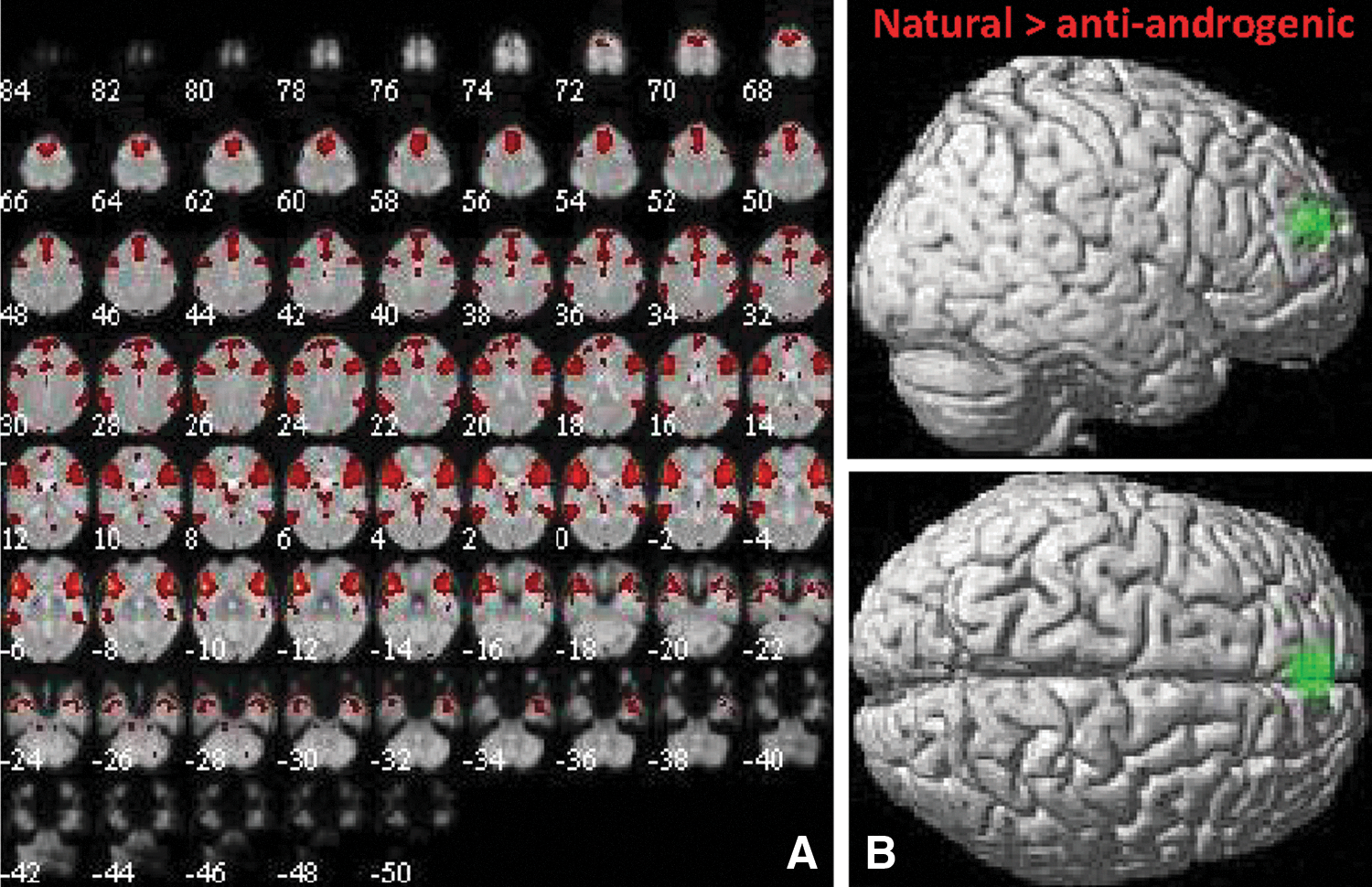
Component 14 (ICN4).
ICN5
We identified component 1 as corresponding to the ICN5 (r = 0.25), described by Laird and colleagues (2011) as including the midbrain and being involved in air hunger tasks. It included large parts of the cerebellum, hippocampus, and parahippocampal gyri. No significant changes between menstrual cycle phases and no differences between OC users and naturally cycling women were observed in connectivity within component 1.
Group 2: networks related to motor and visuospatial integration and coordination
ICN6/9
Component 10 was identified as showing strong correspondence to both ICN6 (r = 0.48) and ICN9 (r = 0.55). ICN6 includes the middle and superior frontal gyri and SMA and has been described as being involved in the preparation of movement, while ICN9 includes the superior parietal lobule (SPL) and has been implicated in hand-eye coordination (Laird et al., 2011). Component 10 included the SMA, middle and superior frontal gyri, pre- and postcentral gyri, and middle cingulum, as well as parts of the SPL bilaterally (Fig. 5A). Menstrual cycle-dependent changes in component 10 are displayed in Figure 5B. A significant increase in connectivity with component 10 was observed after menses in the calcarine gyrus (preovulatory: [18, −76, 8], 119 voxels, T = 4.36, p = 0.03; luteal: [−16, −84, 14], 118 voxels, T = 4.15, p = 0.03) and the SPL (preovulatory: [14, −54, 62], 101 voxels, T = 4.36, p = 0.04). In the luteal compared with the preovulatory phase, a significant decrease in connectivity was observed in the left inferior frontal gyrus ([−48, 14, 4], 116 voxels, T = 4.60, p = 0.03). No differences between OC users and naturally cycling women were observed in connectivity within component 10.
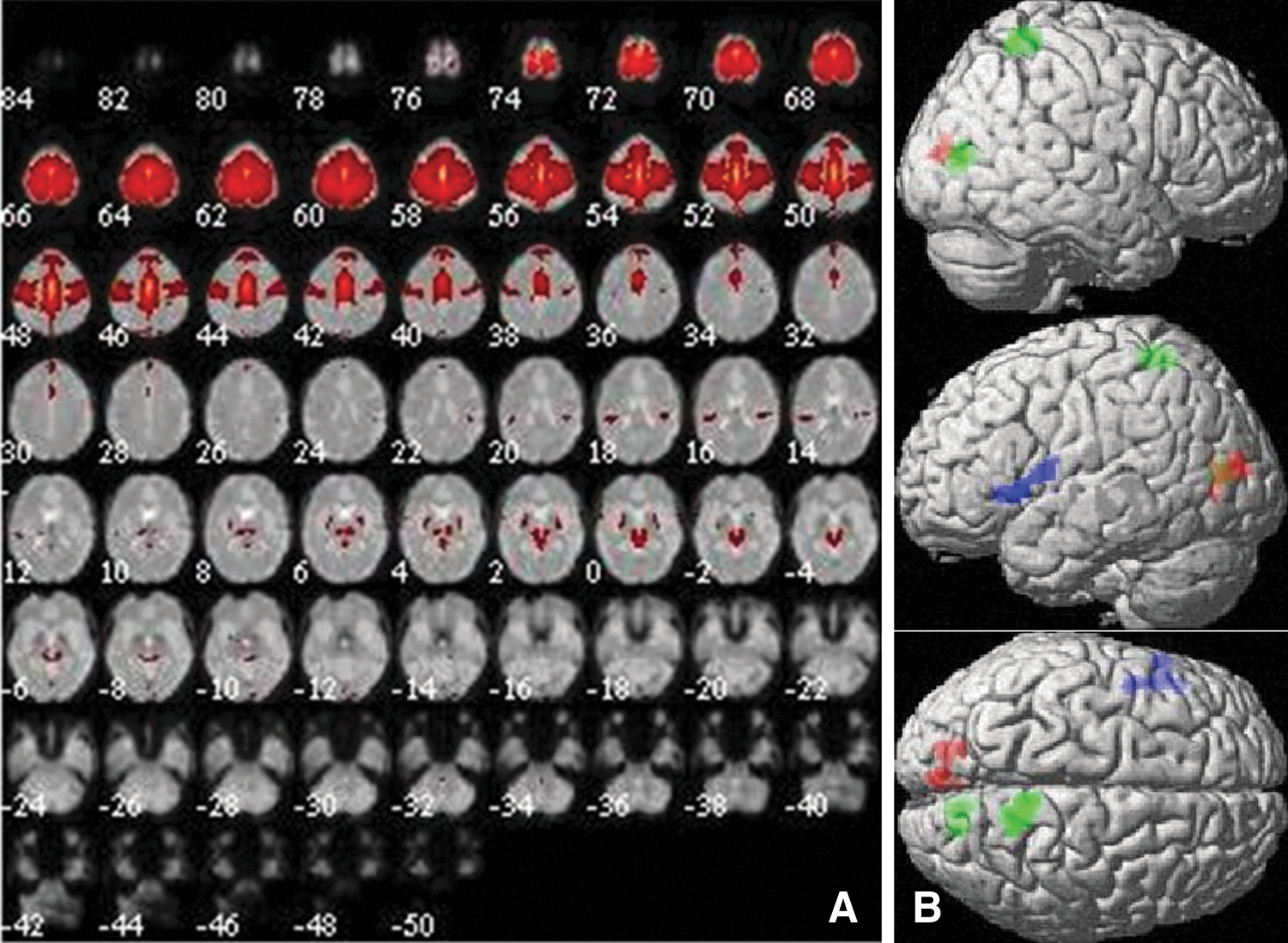
Component 10 (ICN6/9).
ICN7
Component 2 was identified as most likely correspondent to ICN7 (r = 0.41), including the DLPFC and posterior parietal cortices. ICN7 has—similar to ICN4—been proposed as the attentional control network, but more strongly involved in visuospatial tasks such as the Wisconsin Card Sorting task or mental rotation (Laird et al., 2011). Component 2 included the posterior parietal cortices, precuneus and angular gyri, as well as posterior and middle cingulum, and parts of the middle frontal gyri. No significant changes between menstrual cycle phases and no differences between OC users and naturally cycling women were observed in connectivity within this component.
ICN8
Component 8 displayed correspondence to ICN8 with a spatial correlation of 0.38. ICN8 was identified by Laird and colleagues (2011) as the ventral central cerebellar network responsible for hand movement, for example, as in finger-tapping tasks. A significant decrease in connectivity with component 8 was observed during the luteal compared with the preovulatory phase of the menstrual cycle in the right lateral frontal cortex ([20, 52, 4], 146 voxels, T = 4.71, p = 0.008, see Fig. 6). No differences between OC users and naturally cycling women were observed in connectivity within component 8.
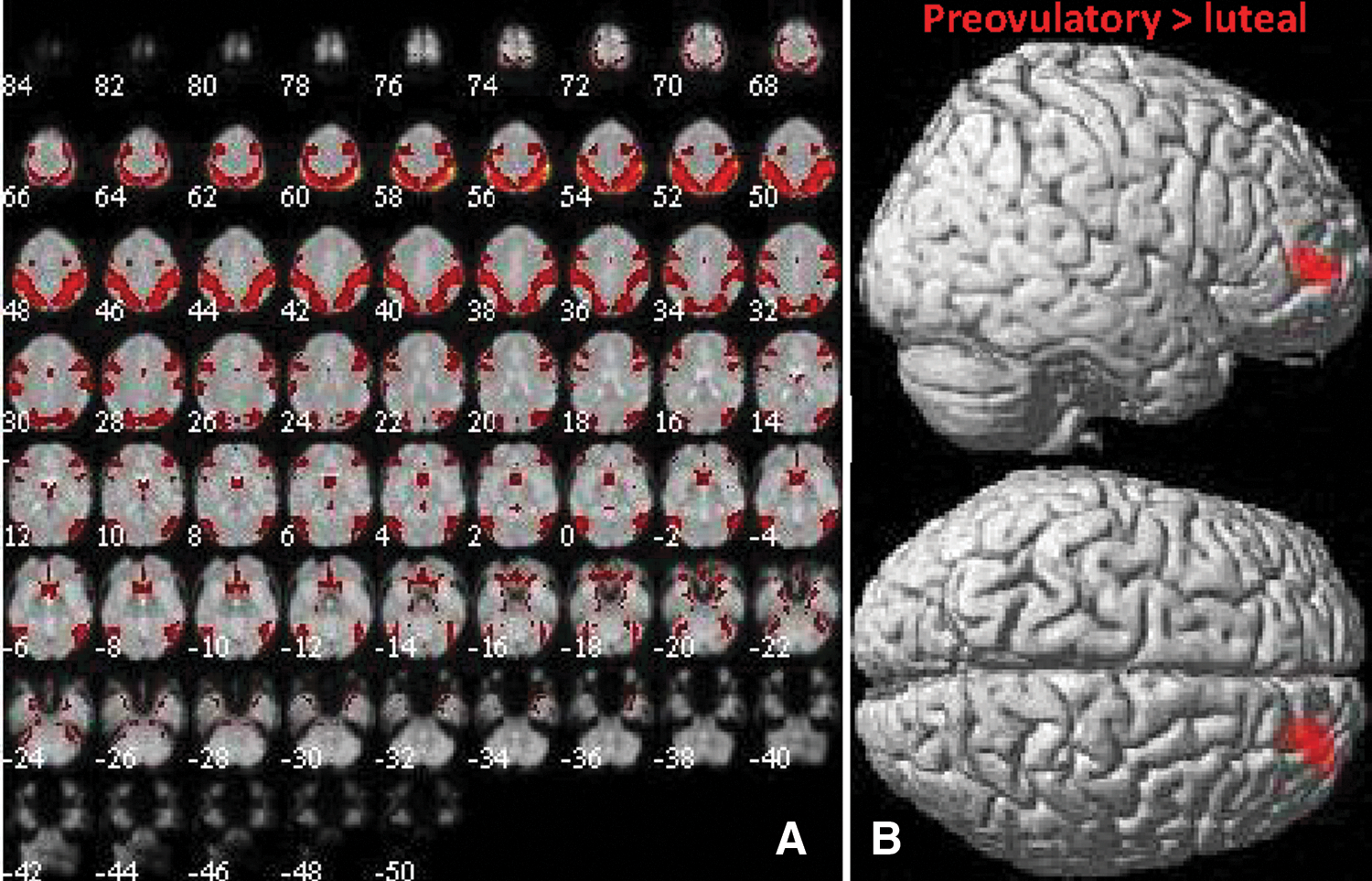
Component 8 (ICN8).
Group 3: networks related to visual perception
ICN10
No component could be identified as corresponding to the temporal visual network ICN10.
ICN11-12
Two components (6 and 13) were extracted corresponding to the two occipital visual networks, ICN11 and ICN12, with spatial correlations of r = 0.52 and r = 0.82, respectively (Fig. 7A, D). Significant changes in both components were observed during the luteal compared with the preovulatory cycle phase (Fig. 7B, E). After ovulation, connectivity with both components increased significantly in the precuneus (ICN11: [−14, −54, 26], 119 voxels, T = 5.66, p = 0.04; ICN12: [18, −36, 62], 193 voxels, T = 4.68, p = 0.001), while a decrease in connectivity with component 6 was observed in the SMA ([8, −28, 62], 128 voxels, T = 4.79, p = 0.02). Accordingly, users of antiandrogenic contraceptives showed increased connectivity compared with naturally cycling women with component 13 in the SMA ([0, 14, 48], 129 voxels, T = 5.31, p = 0.02) and also in the temporal lobe ([54, −28, −2], 167 voxels, T = 3.96, p = 0.01; see Fig. 7F). No differences between OC users and naturally cycling women were observed in connectivity within component 6 (Fig. 7C).
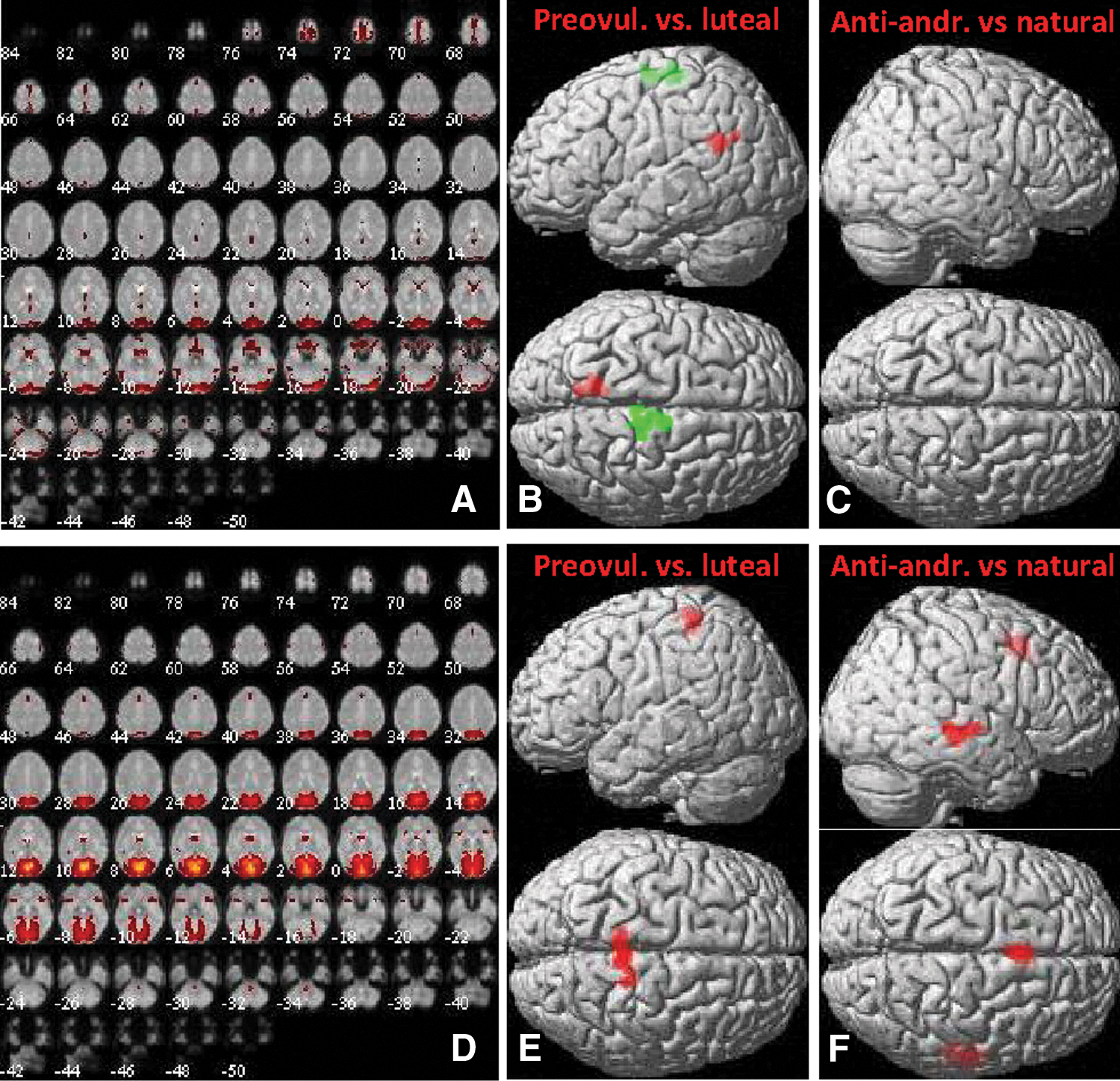
Component 6 (ICN11) and Component 13 (ICN12).
Divergent networks
ICN13 (Default mode network)
Component 19 showed the strongest correspondence to ICN13, described as the DMN (Laird et al., 2011), with a spatial correlation of r = 0.51 (Fig. 8). It included the middle temporal gyri, mPFC, middle and anterior cingulate gyri, angular gyri, and parts of the middle frontal gyri and insula (Fig. 8A). No significant changes between menstrual cycle phases were observed in intrinsic connectivity within component 19 (Fig. 8B). Users of androgenic, but not antiandrogenic, OCs showed a significant increase in connectivity with component 19 compared with naturally cycling women in the mPFC ([6, 48, 8], 148 voxels, T = 4.20, p = 0.03; Fig. 8C).
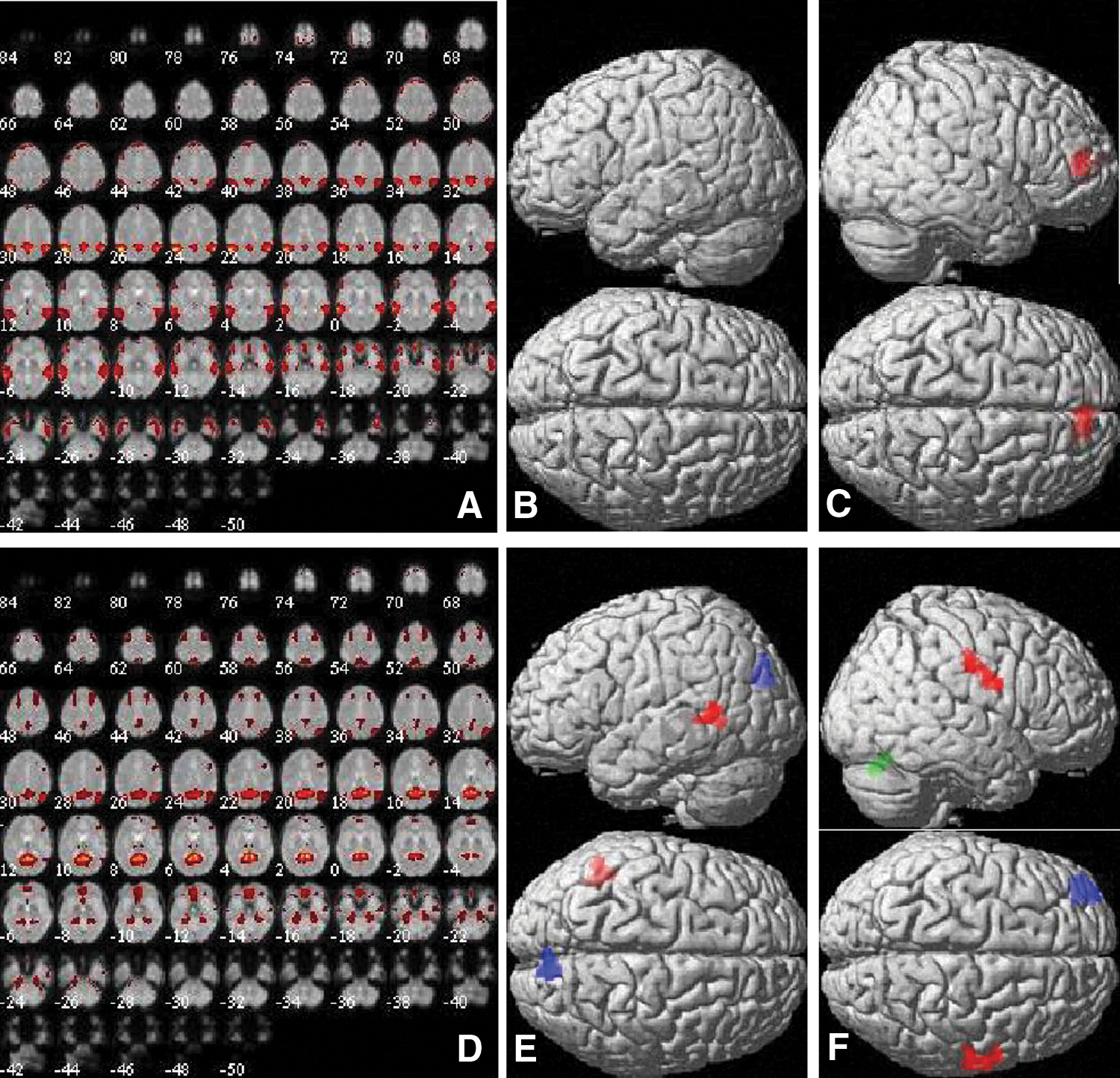
Component 19 and component 16 (ICN13).
However, as in previous resting-state studies (Biswal et al., 2010), a second component (component 16) was identified with high spatial correlation with ICN13 (r = 0.40), which also did not correspond to any of the other ICNs. Component 16 included the precuneus, cuneus, supramarginal gyri, and parts of the middle temporal and middle frontal gyri, as well as the cerebellum (Fig. 8D). This suggests a subdivision of the DMN as identified during task-based studies into a more anterior and more posterior subcomponent during the resting state (compare Laird et al., 2011). A similar subdivision has also been observed in previous ICA studies on menstrual cycle effects (Petersen et al., 2014). Connectivity with component 16 was increased during the preovulatory phase compared with menses in the left temporal cortex ([−46, −52, 6], 119 voxels, T = 4.36, p = 0.03) and during the luteal phase compared with menses in the cuneus ([8, 78, 28], 122 voxels, T = 4.31, p = 0.04; Fig. 8E). In users of androgenic OCs, connectivity with component 16 was increased compared with naturally cycling women in the right postcentral gyrus ([64, −6, 26], 126 voxels, T = 4.00, p = 0.05) and decreased in the cerebellum ([2, −72, −26], 150 voxels, T = 4.70, p = 0.02; Fig. 8F). In users of antiandrogenic OCs, connectivity with component 16 was decreased compared with naturally cycling women in the left superior frontal gyrus ([−38, 48, 8], 130 voxels, T = 4.78, p = 0.06).
ICN14 (Cerebellum)
Component 18 was identified as corresponding to ICN14, the cerebellum with a spatial correlation of 0.50. No significant changes between menstrual cycle phases and no differences between OC users and naturally cycling women were observed in connectivity within this component.
ICN16
Component 9 showed strong correspondence to ICN16 (r = 0.68) described by Laird and colleagues (2011) as including the primary auditory cortices and being involved in audition, for example, in tone discrimination. Component 9 included primarily not only the transverse temporal gyri but also lateral frontal areas (Fig. 9A). No significant changes between menstrual cycle phases were observed in connectivity within component 9. However, in users of antiandrogenic contraceptives, connectivity with component 9 was significantly increased compared with naturally cycling women in the precuneus ([8, −72, 34], 143 voxels, T = 4.99, p = 0.03) and significantly decreased in the right lateral frontal cortex ([14, 38, 38], 120 voxels, T = 4.17, p = 0.05, see Fig. 9B).
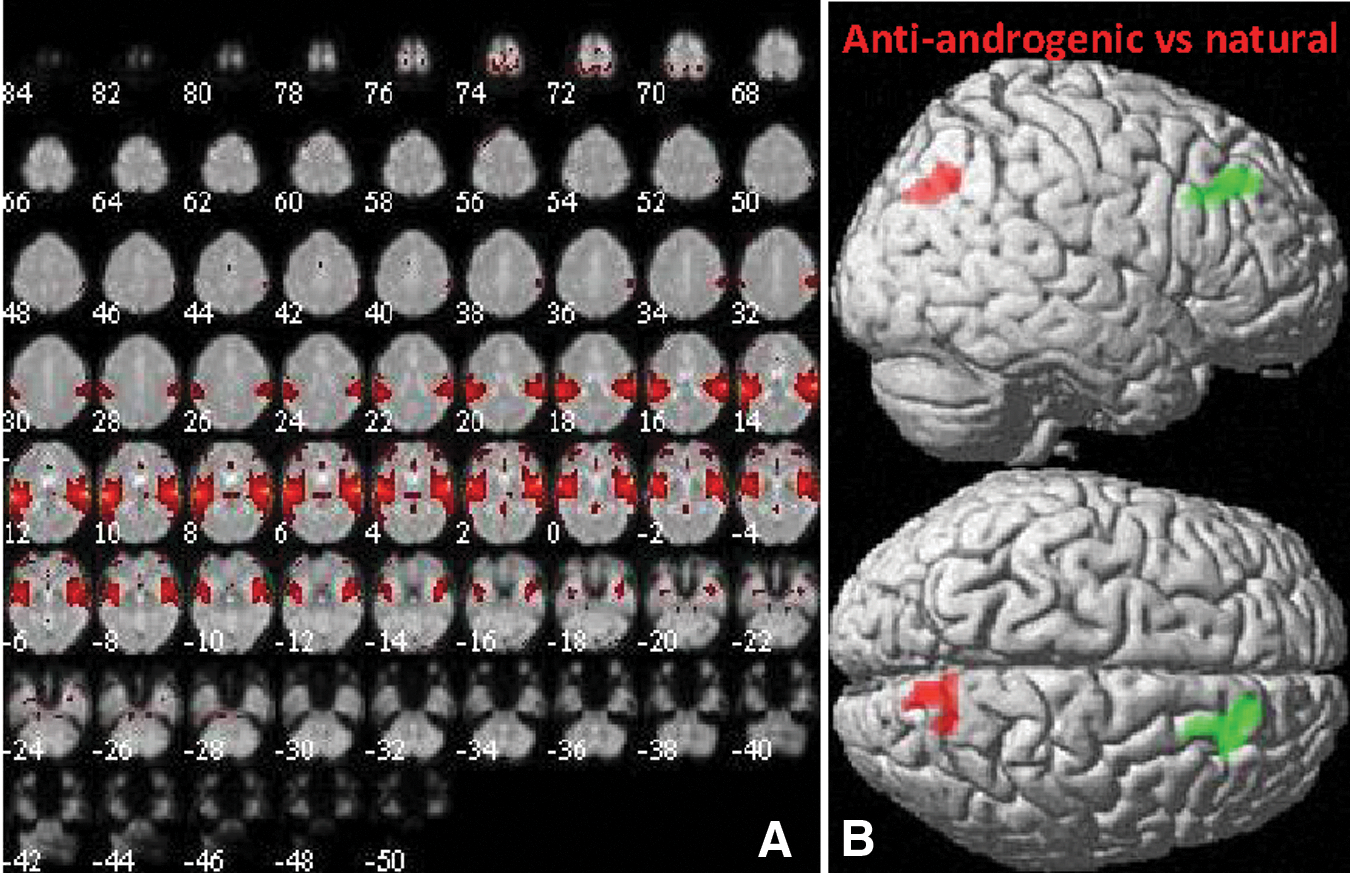
Component 9 (ICN16).
ICN17
Strong correspondence was observed between component 4 and ICN17 (r = 0.72), described by Laird and colleagues (2011) as the dorsal central cerebellar network involved in action and speech, such as during reading (Fig. 6). Component 4 included majorly the bilateral pre- and postcentral gyri and inferior frontal gyri. No significant changes between menstrual cycle phases and no differences between OC users and naturally cycling women were observed in intrinsic connectivity within this component.
ICN15 & ICN18 (Frontoparietal networks)
Component 5 showed strong correspondence to ICN15 (r = 0.54), described by Laird and colleagues (2011) as the primarily right-lateralized frontoparietal network (Fig. 10A) involved in multiple cognitive processes, including reasoning, working memory (e.g., n-back task), and divided attention. Component 15 corresponds to ICN18 (r = 0.44), described by Laird and colleagues (2011) as the primarily left-lateralized frontoparietal network (Fig. 10D), including Broca's and Wernicke's areas, involved in verbal tasks such as word generation, working memory, and explicit memory such as word recall. Significant changes within both networks were observed during the luteal phase of the menstrual cycle (Fig. 10B, E). After ovulation, that is, during the luteal phase, compared with menses, connectivity with component 5 increased in the mPFC ([0, 56, −6], 148 voxels, T = 4.50, p = 0.003) and the basal ganglia bilaterally (left: [−26, 4, 4], 102 voxels, T = 6.93, p = 0.02; right: [26, 10, 0], 157 voxels, T = 5.63, p = 0.003). Connectivity with component 15 decreased in the right precentral gyrus ([42, −22, 42], 124 voxels, T = 4.50, p = 0.02) and right operculum ([48, 12, 6] 147 voxels, T = 4.63, p = 0.009). Accordingly, in users of antiandrogenic OCs, compared with naturally cycling women, connectivity with component 5 was increased in the bilateral basal ganglia compared with naturally cycling women (left: [−26, 2, 12] 171 voxels, T = 4.04, p = 0.01; right: [22, 12, −4], 152 voxels, T = 5.16, p = 0.01; Fig. 10C). No differences between OC users and naturally cycling women were observed in connectivity within component 15 (Fig. 10F).
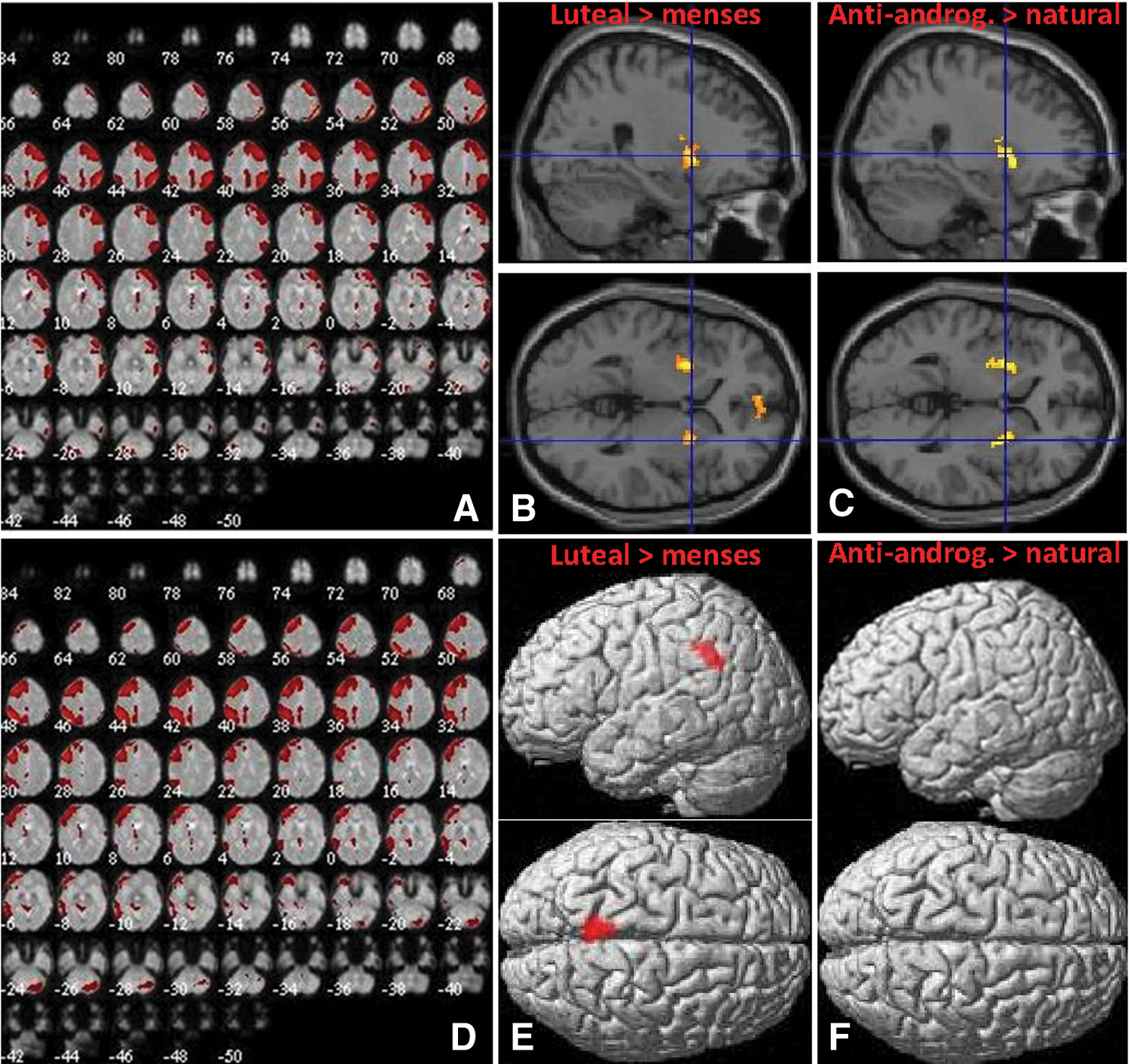
Component 5 (ICN15) and component 15 (ICN18).
Discussion
The aim of the present study was to investigate menstrual cycle and OC-dependent changes in intrinsic connectivity within resting-state networks, going beyond the previous examination of only two networks, that is, the default mode and executive control network. Menstrual cycle-dependent changes for each component were assessed across three cycle phases of naturally cycling women (menses—low progesterone and estradiol; preovulatory—high estradiol; luteal—high progesterone and estradiol). OC-dependent changes were assessed by comparing users of OCs containing androgenic or antiandrogenic progestins with the early follicular phase (menses) of naturally cycling women.
We found changes between menstrual cycle as well as differences between OC users and naturally cycling women in components corresponding to ICN1 (limbic areas: emotion discrimination), ICN 2 (thalamus, BG, and olfactory bulbs: olfaction), and ICN 12 (occipital: vision), as well as ICN15 (right frontoparietal). Changes between cycle phases, but no differences between OC users and naturally cycling women, were observed in two components corresponding to networks clustered in Group 2 (motor/coordination networks), as well as ICN18 (left frontoparietal). Differences between OC users and naturally cycling women, but no changes between cycle phases, were observed in a component corresponding to the DMN, the component corresponding to ICN16 related to audition, as well as the component corresponding to ICN4, referred to as the executive control network. No effects of hormonal status were observed in the components corresponding to the cerebellum (ICN14), the midbrain (ICN5), as well as ICN17 (dorsal central cerebellar network: action and speech) and ICN7 (DLPFC: attentional control). We would like to point out that the majority of OC-dependent effects concern the group of antiandrogenic OC users.
It is notable that five components affected by the menstrual cycle phase are also affected by hormonal contraceptive use. In two of these components (corresponding to ICN2/3 and ICN15, respectively), the differences between menstrual cycle phase and OC groups not only target the same brain areas, that is, the basal ganglia, but also go in the same direction, that is, decreased connectivity with ICN2/3, but increased connectivity with ICN15. The menstrual cycle change occurs in the luteal phase compared with menses and preovulatory phase, while the corresponding OC effects occur in users of antiandrogenic contraceptives compared with naturally cycling women. Thus, differences were observed between hormonal states with high and low estrogenic and progestogenic actions. This suggests that in these networks, endogenous and synthetic hormones target the same sites.
Interestingly, several of the regions affected by hormonal status in the present study have been identified as belonging to the so-called rich club, including the basal ganglia, thalamus, precuneus, and superior frontal cortex, which are highly connected throughout the brain (van den Heuvel and Sporns, 2011). These regions serve as intermediate connectors between different networks. For example, the basal ganglia have been shown to connect cortical and subcortical networks, while the precuneus has been shown to connect the DMN, the frontoparietal control network, and the dorsal attention network with very complex pattern of interaction (Leech and Sharp, 2014). It has been demonstrated that the precuneus shows increased connectivity with the right frontoparietal network during tasks, while it shows increased connectivity with the DMN during rest (Utevsky et al., 2014). Accordingly, in the present study, during high hormone phases, the basal ganglia as well as the precuneus increased their connectivity within one network, while decreasing their connectivity within other networks.
Specifically, connectivity of the basal ganglia during high hormone states decreased within a component corresponding to ICN2/3, two subcortical networks belonging to the emotion and automatic processes group, while there was a simultaneous increase in connectivity of the basal ganglia within ICN15, the right frontoparietal attention network. Changes in connectivity of the basal ganglia have also been observed in ICN1. Likewise, connectivity of the precuneus was decreased during the luteal phase in the components corresponding to ICN1 (emotion and automatic processes group) and increased in components corresponding to ICN11 and ICN12 (visual perception group). The use of antiandrogenic OC (estrogenic and progestogenic actions) was associated with increased connectivity of the precuneus in ICN16. Interestingly, however, in the present study, connectivity of the precuneus in the DMN, the frontoparietal control network, and the dorsal attention network was not affected by hormonal status. Rather, during high hormone states, connectivity of the precuneus during rest increased in networks involved in visual and auditory functions. Another region in the present study, for which such connectivity shifts between networks were observed, is the SMA, which is connected to the basal ganglia (Nambu, 2008). Connectivity of the SMA decreased within ICN1 belonging to the emotion and automatic processes group in users of antiandrogenic contraceptives, while it simultaneously increased within the visual network.
Furthermore, it is interesting to note that the connectivity changes observed within the resting-state networks occur in the same groups and during the same cycle phases as changes in behaviors that have been related to these networks (Laird et al., 2011). In the following, we would like to point out a few examples, which may serve as hypotheses for future studies exploring the behavioral effects of the connectivity changes observed in the present study. Note, however, that these interpretations are to be treated with caution. Although most differences in the present study occur in areas that are known to influence connectivity within and between distinct brain networks (regions belonging to the so-called rich club), it is still unknown whether brain function is similarly affected when OC-dependent changes are limited to certain areas compared with changes in connectivity of the whole network.
As can be viewed in Table 1, connectivity changes in the perceptual, emotional, and motor networks primarily occur around ovulation, while connectivity changes in the divergent networks primarily occur during the luteal phase. These changes in connectivity may in summary relate to a variety of perceptual and behavioral changes in the preovulatory and luteal phases targeted at enhancing the chances to find an ideal mate before ovulation and ensure the safety of potential offspring afterward. In particular, the menstrual cycle-dependent changes in connectivity within a component corresponding strongly to ICN2/3, which has been related to olfaction (Laird et al., 2011), may relate to an altered sense of smell around ovulation (Navarrete-Palacios et al., 2003; Watanabe et al., 2002). Menstrual cycle-dependent changes in the visual networks may correspond to increased visual sensitivity starting around ovulation (Diamond et al., 1972; Eisner et al., 2004). OC-dependent changes in ICN16, related to auditory functions (Laird et al., 2011), may correspond to hormonal contraceptive-dependent masculinization of otoacoustic emission patterns (McFadden, 2000; Snihur and Hampson, 2012). Menstrual cycle-dependent connectivity changes within a component corresponding to ICN1, which has been associated with emotion discrimination (Laird, 2011), may be related to previously reported menstrual cycle-dependent changes in emotion recognition (Derntl et al., 2008a, 2008b; Pearson and Lewis, 2005) during the preovulatory phase. Furthermore, like menstrual cycle-dependent changes in spatial abilities, verbal abilities, memory functions, and attention, as demonstrated by a large number of behavioral studies (Dadin et al., 2009; Hampson, 1990a; Mordecai et al., 2008; Nielsen et al., 2013; Pletzer et al., 2014; Rosenberg and Park, 2002; Solis-Ortiz and Corsi-Cabrera, 2008), connectivity changes within the left and right frontoparietal network also occur during the luteal phase of the menstrual cycle.
Finally, we would like to discuss similarities between our results and previous studies on the effects of menstrual cycle and hormonal contraceptives on resting-state networks. Like in the study of Petersen et al. (2014), we found OC-dependent effects on connectivity within the DMN and executive control network. In the present study, both affected the ACC/mPFC. The changes in the executive control network were in accordance with Petersen and colleagues (2014) (increased connectivity in OC users in the ACC). However, contrary to the study of Petersen and colleagues (2014), the effects of OC use in both components corresponding to the DMN were observed in different brain areas. These differences may be attributable to the fact that unlike Petersen and colleagues (2014), the present study differentiated between androgenic and antiandrogenic OCs. Note also that the DMN is the only network affected by use of androgenic OCs. Unlike Petersen and colleagues (2014), but comparable with Hjelmervik and colleagues (2014) and De Bondt and colleagues (2015), we did not observe any menstrual cycle-dependent effects on intrinsic connectivity within the DMN. However, differences in study designs may have contributed to these inconsistencies. While Petersen and colleagues (2014) used a between-subjects approach to assess cycle-dependent changes, the present as well as other previous studies (De Bondt et al., 2015; Hjelmervik et al., 2014) used a within-subjects design.
In addition, other than Petersen and colleagues (2014), but in accordance with Hjelmervik and colleagues (2014) and De Bondt and colleagues (2015), we did not observe any significant differences across the menstrual cycle or due to hormonal contraceptive use in the executive control network or the Stroop network. This result is in contrast to a number of behavioral studies indicating menstrual cycle-dependent changes in cognitive control and inhibitory functions across the menstrual cycle (Colzato et al., 2010, 2012; Hatta and Nagaya, 2009). However, it has already been discussed that these behavioral effects may be bidirectional depending on the baseline dopamine level (Colzato and Hommel, 2014; Jacobs and D'Esposito, 2011). Thus, women with effects in different directions may have been collapsed into one group in the present study, as in previous studies on resting-state connectivity of the executive control network using a within-subjects design (De Bondt et al., 2015; Hjelmervik et al., 2014).
In summary, the present study suggests that changes in intrinsic connectivity due to the menstrual cycle phase and hormonal contraceptive use affect resting-state networks associated with functions for which menstrual cycle and OC-dependent changes have also been reported. Moreover, the changes in connectivity and changes in behavior occur during the same cycle phases, specifically the luteal phase. Inconsistencies in connectivity changes arise in networks, that is, the control networks in which differential behavioral effects have been reported regarding the associated functions, that is, menstrual cycle-related changes in cognitive control. Furthermore, OC-dependent effects were observed in the same networks and same target sites as menstrual cycle-related changes and were dependent on the androgenicity of the progestin component contained in the OC.
Footnotes
Acknowledgments
The authors thank Christina Brötzner and Eva Reiter for help with data acquisition and all participants for their time and willingness to contribute to this study.
Author Disclosure Statement
No competing financial interests exist.