
Abstract
There exists diversity among individuals in difficulty controlling body weight. Body weight control, or obesity prevention, requires cognitive control over ingestive behavior, which may account for the diverse ability of body weight control. The caudate nuclei, especially the dorsal area, have been shown to play critical roles in ingestive behaviors, which significantly influences body weight control. However, the practice of body weight control is dependent on the body weight status, because the current obesity status determines the need for body weight control. To elucidate the underlying neural mechanism that accounts for individual differences in obesity prevention, we aimed to isolate functional caudate connectivity responsible for the underlying tendency of obesity prevention, independent of the current obesity status, using resting state fMRI data, body mass index (BMI), and assessment of ingestive behavior from 185 individuals from the NKI-Rockland sample. The underlying tendency of obesity prevention was estimated from BMI and behavioral and cognitive components of food intake. Functional connectivities between the caudate head and the whole brain were tested as a function of the estimated tendency in a voxel-wise manner. The bilateral precuneus showed inverse association between its connectivity to the caudate and the estimated tendency. Caudate–precuneus connectivity may have significant implications to understanding personal differences that accounts for the success in body weight control.
Introduction
A
Neuroimaging studies have demonstrated that regions associated with reward, motor, cognition, control, and attention are associated with obesity (Carnell et al., 2012; Pursey et al., 2014; Schlögl et al., 2016; Val-Laillet et al., 2015). Specifically, the caudate activity is associated with obesity. The caudate receives dopaminergic inputs from the substantia nigra and has extensive cortical projections (Dauer and Przedborski, 2003; Purves et al., 2001) through the thalamus (Grahn et al., 2009). The caudate is also implicated in a number of cognitive functions, including learning (Shohamy et al., 2008), memory (Frank et al., 2001), habit formation (Knowlton et al., 1996; Liljeholm and O'Doherty, 2012), working memory (Bäckman et al., 2011; Chatham et al., 2014), inhibitory control (Dalley et al., 2011; Tschernegg et al., 2015), and goal-directed behavior (Balleine and O'Doherty, 2010). Positron emission tomography studies demonstrated decreased receptor availability in the caudate of severely obese population (de Weijer et al., 2011; Wang et al., 2001), and increased availability (Cosgrove et al., 2015; Dunn et al., 2010; Guo et al., 2014) and meal-induced release (Kessler et al., 2014; Wang et al., 2014) in overweight and mild obesity. Caudate glucose uptake (Nummenmaa et al., 2012) and blood oxygen level dependent responses to the sight of calorie-predictive cues are also elevated in obese population (Ng et al., 2011; Nummenmaa et al., 2012; Rothemund et al., 2007; Stoeckel et al., 2008), whereas response to the receipt of a palatable and energy dense food is decreased (Burger and Stice, 2014; Frank et al., 2012; Green et al., 2011). Notably, the magnitude of the decreased caudate response predicts weight gain, in individuals who carry a copy of the A1 allele for the Taq1A polymorphism (Stice et al., 2015), which is associated with decreased dopamine receptors in the striatum (Jönsson et al., 1999). Moreover, the caudate nucleus facilitates ingestive behavior by anticipating reward cues (Burger and Stice, 2014; Pursey et al., 2014) and regulating reward-seeking behavior (Nishino et al., 1984; Ono et al., 2000).
The caudate nucleus comprises three regions: head, body, and tail. The caudate head is anatomically and functionally connected with emotive and cognitive regions, including amygdala, cingulate cortex, and prefrontal cortex such as orbitofrontal cortex, inferior frontal gyrus, and rostral anterior cingulate, and the caudate body is connected with self-control regions, such as the motor control region (superior and medial frontal gyrus), and cognitive control regions (e.g., dorsolateral prefrontal cortex) (Di Martino et al., 2008; Robinson et al., 2012). Specifically, the caudate head is closely linked to the frontal regions that facilitate reward, motivation, and affect regulation, and is also associated with pathological risk-taking and addictive behaviors, such as binge eating disorder (Kessler et al., 2016). Whereas the caudate tail receives inputs mainly from the inferotemporal cortex (Saint-Cyr et al., 1990). A majority of caudate tail neurons show differential responses to visual objects (Brown et al., 1995; Caan et al., 1984; Yamamoto et al., 2012), and is likely to be involved in gaze/attention capture (Chelazzi et al., 2013; Kim et al., 2015). Unlike the neurons in the rest of the caudate, neurons in the caudate tail were less influenced by reward values (Yamamoto et al., 2013). Altogether, previous findings raise the possibility that the caudate head, which may, in turn, have broader emotive and cognitive functional implications, would be associated with obesity preventive personal tendency.
Body weight is a relatively flexible human body phenotypical state, compared with height, for example. Body weight can be changed by the amount of food intake as well as energy metabolism. Most importantly, intentional body weight change, especially weight loss, is often attempted to avoid increased health risks by obesity and/or to achieve aesthetic ideals. Inhibiting body weight gain, however, is a major daily life concern to some but not others. This study aimed to isolate the neural correlate associated with this cognitive tendency to attend body weight.
Practices of cognitively restraining eating for body weight control (Herman and Mack, 1975) have been shown to be associated with adiposity (Aurélie et al., 2012; de Lauzon-Guillain et al., 2006; Dykes et al., 2004; Rideout and Barr, 2009). However, previous findings show that body mass index (BMI) and cognitive restraint (CR) of eating are positively associated in normal weight individuals and negatively in overweight and obese individuals (Cappelleri et al., 2009; de Lauzon-Guillain et al., 2006; Foster et al., 1998). Indeed, body weight status has been shown to predict CR of eating as a quadratic function in an inverted-U shape relationship (Dietrich et al., 2014). That is, CR of eating is the most practiced in the mid BMI range.
Given this BMI-dependent enforcement of food intake restriction, we hypothesized that (a) the obesity preventive tendency can be approximated by the residuals of the quadratic function that predicts CR from BMI, and that (b) this tendency would be associated with functional brain connectivity (Biswal et al., 2010; Fox and Raichle, 2007) to/from the caudate head that is responsible for regulating food intake. Specifically, associations were tested between the obesity preventive tendency and the functional connectivity from the caudate head and the rest of the brain in a voxel-wise manner.
Materials and Methods
Data acquisition
NKI-Rockland resting state fMRI (rs-fMRI) and SPGR data have been downloaded from fcon 1000 project website, along with demographic and other behavioral and cognitive assessments (
Obesity preventive eating tendency index
The CR of eating items (Factor I) from TFEQ has been used to assess current BMI-dependent ingestive behavior. Responses are scored 0 or 1 and summed. Higher scores denote higher levels of restrained eating. It has been shown that body weight status is associated with TFEQ-CR (Gibson, 2012) and also with the white matter structural connectivity, which were supposed to regulate ingestive behavior (Gupta et al., 2015). Using SPSS Version 21.0 (SPSS, Inc., Chicago, IL), a quadratic regression from BMI to TFEQ-CR was tested first to obtain studentized residual of the regression as the obesity preventive eating tendency (OPET) index.
Image processing
Script package from the 1000 Functional Connectomes Project (
The bilateral caudate nuclei were first extracted from Freesurfer (
Results
Sample demographics
Samples from 185 participants contained all of the rs-fMRI, MPRAGE, BMI, and TFEQ data and used for the analyses. Mean BMI (mean ± standard deviation [SD]) was 26.1 ± 5.48 kg/m2; mean age was 37.4 ± 19.4 years; 74 of the participants were female.
OPET scale and correlations between TFEQ scales and BMI
Mean TFEQ-CR (mean ± SD) was 7.5 ± 0.35. TFEQ-CR showed a robust inverted U-shaped relationship with BMI (squared partial correlation: 0.062, p = 0.006). The estimated peak of the TFEQ-CR was at BMI = 29.9 kg/m2.
Correlations between OPET index and connectivity maps
One cluster showed negative correlation between caudate connectivity and the OPET index ([1046 52], p = 0.004 FWE corrected, z = 3.51; Fig. 1). Other local peaks in the cluster are listed in Table 1. The left side of peak voxel in this cluster was placed in the precuneus and the left side of cluster was located from the anterior part of superior parietal lobe to inferior parietal lobe, including postcentral gyrus. The right side of the peak voxel was placed in the precuneus and the right side of cluster was located from the precuneus to middle cinglate gyrus, including paracentral lobe and superior marginal gyrus. There was no significant cluster that showed positive correlation between caudate connectivity and OPET index. The mean displacement within each individual rs-fMRI series was 0.13 (±0.21) mm. There was a significant correlation between OPET index and functional connectivity Z score within the cluster after controlling for the mean displacement (r = −0.25, p = 0.001) and for age (r = −0.24, p = 0.001).
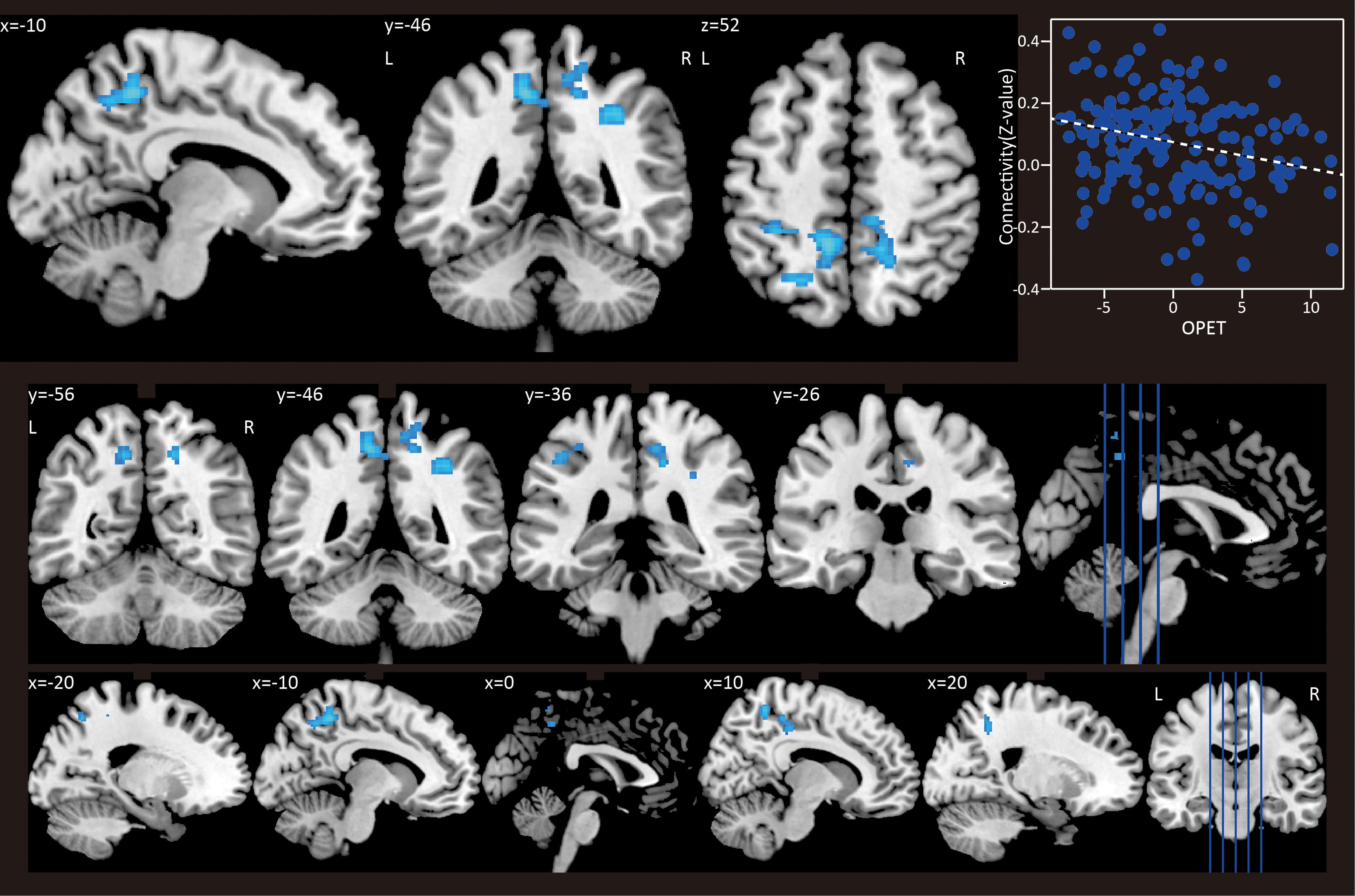
Correlations between OPET index and connectivity maps. One cluster showed negative correlation between caudate connectivity and the OPET index ([−10 − 46 52], p = 0.004 FWE corrected, z = 3.51). A scatter plot depicts connectivity values (z values) and OPET. FWE, family-wise error rate; OPET, obesity preventive eating tendency. Color images available online at
Cluster size in voxels.
Italics indicate that peaks fall under the same cluster as the preceding peak.
Discussion
To our best knowledge, this is the first study that tested functional brain connectivity and the personal tendency of obesity prevention. Caudate–precuneus functional connectivity showed inverse correlation to the underlying tendency of obesity prevention, estimated by OPET index. Higher caudate–precuneus connectivity was associated with lower obesity preventive tendency. The results can suggest that caudate–precuneus connectivity may inhibit the processes to cognitively prevent obesity.
The association between TFEQ-CR and BMI replicated previous finding that BMI predicts TFEQ-CR in an inverse-U manner (Dietrich et al., 2014). TFEQ-CR showed its peak (BMI = 29.9 kg/m2) within the overweight BMI range, very close to the obese threshold. This relationship can be interpreted that CR of eating is most practiced in the overweight to light obese range. Current body weight status has been shown to be associated with TFEQ-CR scores (Gibson, 2012) and also with the white matter anatomical connectivity, which is understood to regulate ingestive behavior (Gupta et al., 2015). Therefore, TFEQ-CR score would reflect the current status of CR practice, and the practice is reflected in individuals' current body weight. Here, we aimed to first eliminate this association so that we can isolate individuals' underlying tendency of paying attention to over eating, whether or not they are successful.
The precuneus has previously been implicated in various functions, for example, visuospatial imagery, episodic memory retrieval, and self-processing operations (Cavanna and Trimble, 2006), and has a significant role in the default mode network (Utevsky et al., 2014), attentional network (Soto et al., 2012), and self-consciousness (Kjaer et al., 2002). Anatomically, the caudate and precuneus have direct anatomical neural connections (Selemon and Goldman-Rakic, 1985; Yeterian and Pandya, 1995).
Previous studies support the association between the caudate–precuneus connectivity and obesity preventive cognitive processes. First, the anterior precuneus has been shown to be associated with obesity. In obese population, the precuneus has been found to have (a) greater activity at the baseline (Zhang et al., 2015), (b) higher functional connectivity to the default mode network (Kullmann et al., 2012), and (c) increased regional cerebral blood flow after satiation (Gautier et al., 2000). Second, the precuneus is implicated in body shape consciousness. The precuneus activity has been found to facilitate body shape estimation (Mohr et al., 2010). Body shape preoccupations have been found to correlate with the resting state functional connectivity between the anterior precuneus and dorsal anterior cingulate cortex (Lee et al., 2014). Third, the precuneus is implicated in eating disorders. Decreased gray matter volume in patients of anorexia nervosa has been reported (Joos et al., 2011). Patients of anorexia nervosa and bulimia nervosa showed greater functional connectivity to dorsal anterior cingulate cortex than the control group (Lee et al., 2014). More recently, the precuneus volume has been found to be associated with general self-efficacy (Sugiura et al., 2016). As general self-efficacy is an index of abilities to succeed in a various task by complying to one's own decision, success in body weight control may reflect the function of the precuneus. Although there was no direct association between obesity/eating behavior and the function connectivity between caudate and precuneus, aforementioned findings indirectly indicate that the precuneus is involved in obesity inducing or preventing behavior, through self-body consciousness as well as body weight control, whose failure can be manifested as obesity or eating disorders.
The current results supported the previous findings that the caudate modulates food reward processing. Cognitive restrained eating behavior has been shown to be associated with caudate reactivity to food reward stimuli. In addition, the activation in the dorsal striatum to downregulate or to admit one's desire for food showed correlations with the degree of dietary restraint (Hollmann et al., 2012). These results suggest that the dorsal striatum would regulate a desire for highly appetizing food, and modulate one's desire for such highly appetizing foods in concert with other brain regions including a gustatory cortex (e.g., the insula), and those caudate functions would be influenced by a cognitive restrained eating trait. This study strongly implicates that the functional connectivity to the precuneus is involved in food reward processing over obesity prevention.
In this study, the caudate–precuneus functional connectivity showed inverse association with the tendency of obesity prevention. The caudate, on one hand, is responsible for the food reward proceeding, whereas the precuneus is responsible for body weight consciousness. It is noteworthy that the precuneus and caudate have been shown to be anatomically and functionally associated (Cavanna and Trimble, 2006; Di Martino et al., 2008; Tomasi and Volkow, 2013). This implies that the caudate–precuneus finding would have an anatomical basis, rather than accidental coactivation.
The results of this study have to be considered in light of the following limitations. First, although we showed the association between the caudate–precuneus functional connectivity and the tendency of preventing obesity, we cannot address the causality of this relationship. Second, we would not be able to exclude the other factors that would modulate this relationship, such as education levels (Wagenknecht et al., 2007), social income (Wardle and Griffith, 2001), or cognitive functions (Cournot et al., 2006). Third, not only the caudate neural circuit but also other neural circuits would contribute to the tendency of preventing obesity. We need further research such as weight loss trials to bridge the lack of direct links between the caudate–precuneus functional connectivity and the tendency of preventing obesity.
We showed that the tendency of preventing obesity has negative association with caudate–precuneus functional connectivity. The personal characteristics of avoiding obesity-inducing behaviors were manifested as an inverse function of anticipation of food reward. The underlying neural circuit for attending and preventing obesity, if any, had not been identified. This study suggests that caudate–precuneus functional connectivity may facilitate such tendency and may play an important role in body weight control.
Footnotes
Acknowledgment
Image preprocessing was performed using the supercomputer cluster at the Mississippi Center for Supercomputing Research.
Authors' Contribution
Y.N. performed statistical analysis. T.I. performed preliminary processing of the resting state fMRI data. Both authors were involved in writing the article and had final approval of the submitted and published versions.
Author Disclosure Statement
No conflict of interest to disclose.