
Abstract
The relationship between post-traumatic stress disorder (PTSD) and chronic symptoms of mild traumatic brain injury (mTBI) is difficult to discern and poorly understood. An accurate differential diagnosis, assessment, and treatment of mTBI and PTSD are challenging due to significant symptom overlap and the absence of clearly established biomarkers. The objective of this work is to examine how post-traumatic stress influences task-free default mode network in chronic mTBI subjects. Control subjects (N = 44) were compared with chronic mTBI subjects with low (N = 58, PTSD Checklist–Civilian Version [PCL-C] total < 30), medium (N = 124, PCL-C total = 31–49), and high (N = 105, PCL-C total ≥ 60) post-traumatic stress symptoms (PTSS). The results indicate significant differences in Brodmann area 10 for all mTBI subject groups, indicating potential mTBI-related disruptions with regulation of emotions and decision-making. The effects of PTSS were observed in the anterior cingulate and parahippocampus, suggesting possible disruptions pertaining to memory regulation, encoding, and retrieval. The overall results indicate the presence of aberrant connectivity patterns between controls and chronic mTBI subjects with low, medium, and high PTSS. Furthermore, the findings suggest a disruption in attention relating to a network of brain regions involved with emotional regulation and memory coding, rather than a fear-related response. Taken together, the results suggest these regions form a network that could be a target for future research pertaining to PTSD and chronic mTBI. Furthermore, the use of clinical measures, task-based imaging studies, or multimodal imaging could help further elucidate specific neural correlates of PTSS and mTBI.
Introduction
I
The presence of postconcussive symptoms in mTBI significantly increases the risk of developing other disorders, one of which is post-traumatic stress disorder (PTSD) (Schneiderman et al., 2008; Taylor et al., 2012). The clinical diagnosis of PTSD (Diagnostic and Statistical Manual of Mental Disorders version 4-TR [DSM-IV-TR]) is based on persistent symptoms exceeding 1 month that encompass three main symptom clusters: hyperarousal, re-experiencing, and avoidant behavior pertaining to events that are extremely terrifying, involve grievous injury, or actual or threatened death directed to the individual or another person either by first-hand witness or indirect communication (American Psychiatric Association, 2000). Furthermore, the individual would have to react to the traumatic event with intense fear, helplessness, or horror, and the PTSD symptoms must significantly impair social and occupational functioning or create clinically significant distress (American Psychiatric Association, 2000).
PTSD is defined as psychological trauma. Indeed, the manner in which a person sustains an mTBI (i.e., exposure to explosive ordnance, falls, motor vehicle accidents, and blunt force trauma) (Silver et al., 2011; Tanielian and Jaycox, 2008; Warden, 2006) may be associated with traumatic events in their own right, capable of inducing fear, helplessness, or shock. Studies have also shown that mTBI can exacerbate post-traumatic stress symptoms (PTSS), therefore increasing the risk of developing PTSD (Brenner et al., 2011; Bryant and Harvey, 1998; Schneiderman et al., 2008; Taylor et al., 2012). Researchers at the Department of Veterans Affairs report that in a large sample of mTBI subjects, 73% had PTSD (Taylor et al., 2012). There is significant overlap between persistent postconcussive symptoms and PTSS such as hyperarousal, sleep disturbance, headache, impaired concentration, fatigue, and somatosensory and cognitive impairment (Bryant and Harvey, 1998; Greenspan et al., 2006). The high comorbidity of mTBI and PTSS often hinders the performance of activities of daily living leading to high rates of disability and decreased quality of life. In addition, comorbid manifestation of mTBI and PTSD may increase the risk for suicide (Brenner et al., 2011).
However, not all mTBI patients go on to develop PTSD, and the exact mechanisms that drive the development of PTSD in some mTBI patients and not others are unknown. It is thought that post-traumatic stress can develop from mTBI based on four possible methods (King, 2008). First, because mTBI patients have little or no amnesia (as opposed to those with most severe TBI with extended loss of consciousness [LOC]), they are able to recall all or parts of the traumatic event (Creamer et al., 2005). Second, instead of veridical memory encoding, subconscious mechanisms may encode the fear response, which is then triggered when the patient is subsequently exposed to specific stimuli reminiscent of the traumatic event (Creamer et al., 2005; Harvey and Bryant, 2001; Roitman et al., 2013). Third, the formation of “pseudomemories” by either imagined or reconstructed events could manifest into PTSD (Harvey and Bryant, 2001). Finally, events that are not directly related to the cause of the mTBI, such as painful medical procedures, loss of control, dignity, and livelihood, could lead to PTSD symptomatology (King, 2008).
Functional imaging tools provide a noninvasive method to understand the mechanisms of mTBI and PTSD in human subjects in vivo. Task-based imaging studies report increased activation in the amygdala, insula, hippocampus, anterior cingulate, and regions of the frontal gyrus in patients with PTSD (Bruce et al., 2013; Sripada et al., 2013). These are key regions that form a brain network involved in fear response and encoding of fearful memories (Bruce et al., 2013; Sripada et al., 2013). In contrast, other research suggests reduced amygdala and hippocampus activation in PTSD subjects due to misrepresentation of stimuli that are encoded incorrectly by the brain (Hayes et al., 2011). These discrepancies could be attributed to the diverse methods used for fear conditioning (e.g., narrated scripts, trauma recollection, or images used to invoke PTSD flashbacks) and individual differences in extinction for specific stimuli. It is unclear if the observed activations are the result of selective response to the specific experimental stimuli or a general response indicative of PTSD symptomology (Yan et al., 2013a).
Task-free paradigms offer a method to understand the regulatory mechanisms of consciousness in the absence of experimental manipulation. In the absence of explicit stimuli, connectivity between specific brain regions form distinct topological networks (Beckmann et al., 2005). The default mode network (DMN) is a prominent pattern of brain connectivity observed during the absence of explicit stimuli; however, connectivity of the DMN has been observed to diminish with cognitively demanding tasks (Raichle et al., 2001). The presence of aberrant connectivity within the DMN has been reported in both mTBI and PTSD studies (Burgess et al., 2007; Bush et al., 2000; Gilbert et al., 2006; Vlassenko et al., 2004; Yan et al., 2013a). Furthermore, some of these connectivity differences are found to correlate with symptoms such as increased fatigue, anxiety, and cognitive impairment (Burgess et al., 2007; Gilbert et al., 2006; Vlassenko et al., 2004). However, a key limitation of many TBI studies is broad inclusion criteria that often involve a combination of multiple severity levels (mix of mild, moderate, and severe). In addition, many TBI studies focus on the acute phase of injury or combine both acute and chronic subjects. Finally, very few studies use functional imaging tools to study chronic mTBI patients with comorbid PTSD.
The main goal of this study is to understand how networks of the brain are affected in chronic mTBI patients with comorbid PTSS. We focus on the task-free DMN to determine if there are specific features of network connectivity between brain regions that distinguish chronic mTBI from comorbid chronic mTBI with PTSS. The purpose in measuring the task-free DMN is to minimize stimulus bias and subjectivity of response across subjects. Furthermore, the task-free DMN provides a uniform method for studying salient cognitive functions that pertain to conscious states of mind and has been shown to be sensitive to pathology (Beckmann et al., 2005; Broyd et al., 2009; Damoiseaux et al., 2006; Van de Ven et al., 2004).
In this study, independent component analysis (ICA) decomposition was used because this is a multivariate approach with nonparametric association and without any a priori assumptions (Hyvärinen et al., 2004). When compared to seed-based correlation analysis, ICA is an unbiased data-driven method for exploratory data analysis (Hyvärinen et al., 2004). Furthermore, seed-based correlations have limitations pertaining to the selection of seed regions, consistency of seed region connectivity across subjects, and the assumption that the seed voxel time course is an accurate representation of other voxels within brain regions of the functional network (Beckmann et al., 2005; Damoiseaux et al., 2006; Van de Ven et al., 2004). These assumptions are arbitrary and do not account for potential artifacts from various sources of indeterminate noise in the MRI scanner (Beckmann et al., 2005; Damoiseaux et al., 2006; Van de Ven et al., 2004). Furthermore, seed-based correlation methods are susceptible to spurious correlations (false positive) or conversely missing important patterns or coactivity (false negative) in addition to not being able to account for time course composites within the functional MRI (fMRI) blood oxygen level–dependent (BOLD) signal (Beckmann et al., 2005; Damoiseaux et al., 2006; Van de Ven et al., 2004).
Methods
Subjects
Control subjects recruited for this study consisted of service members and seven civilians enrolled in the Defense Enrollment Eligibility Reporting System, who were free from TBI, previous diagnosis of psychiatric disorders, head injuries, neurological injuries, and were neither pregnant nor breastfeeding. The TBI subjects consisted of active duty service members with a diagnosis of mTBI based on routine comprehensive clinical screening evaluations by healthcare professionals at the Walter Reed National Military Medical Center (WRNMMC), using U.S. Department of Veterans Affairs and U.S. Department of Defense (2016) guidelines for TBI. The determination of mTBI severity was based on the presence and duration of post-traumatic amnesia (PTA; 0–1 days), LOC (0–30 min), alteration of consciousness (AOC; <24 h), and the lack of radiological findings as reported by a licensed clinical radiologist. Formal assessment of PTA, LOC, and AOC was performed during clinical TBI evaluation at WRNMMC, and additional information regarding the duration of these measures was obtained from medical records of the initial treatment facility during the acute phase of injury. In this study, the mTBI subjects were not distinguished based on mechanism of injury (i.e., blast compared to nonblast). At present, definitive biomarkers and indicators of blast on the human brain and long-term sequelae are not clearly understood, and disentangling this information from behavioral deficits or multiple incidences of injury remains an area of extensive research (Belanger et al., 2009; Lange et al., 2012).
The mTBI subjects were divided into three groups based on the total PCL-C scores: low PTSS (PCL-C total < 30), medium PTSS (PCL-C total = 31–49), and high PTSS (PCL-C total ≥ 60) groups. At present, there is no agreed upon cut score for the PCL-C, with studies reporting cut scores ranging from 30 to 60 (McDonald and Calhoun, 2010). For this study, we selected a cut point of ≥60 for high PTSD symptoms and a cut point of <30 for low or no PTSD symptoms (McDonald and Calhoun, 2010). We chose a “medium” PTSD group based on scores of 31–49. All mTBI subjects recruited for this study were at least 6 months postinjury and were excluded if they were unable to consent themselves, were actively participating in other research studies, or were pregnant or breastfeeding. On the day of the scan, all mTBI subjects completed self-report assessment forms. All subjects provided written informed consent and this study was approved by the Institutional Review Board of WRNMMC. Complete demographic information for each of the mTBI subject groups is summarized in Table 1. A one-way Welch analysis of variance (ANOVA; Bonferroni corrected for multiple comparisons) performed with Statistical Software Package (SPSS; IBM, Inc., Armonk, NY) revealed no significant differences between all groups for age (F[3, 325] = 0.772, p = 0.51) and no significant difference between mTBI groups (F[2, 271] = 0.551, p = 0.577) for time since injury. Years of education were significantly different (F[3, 325] = 33.91, p < 0.001) between groups, with Games–Howell post-hoc comparisons indicating significantly higher (p < 0.001) years of education between the control group and all three PTSS mTBI groups; and significantly higher (p < 0.015) years of education between the low PTSS mTBI group and the high PTSS mTBI group; years of education did not differ between the low and medium PTSS mTBI groups (p = 0.057) and between the medium and high PTSS mTBI groups (p = 0.911).
PCL-C, PTSD Checklist—Civilian Version; PTSS, post-traumatic stress symptoms.
Self-report assessments
The PCL-C (Blanchard et al., 1996; Keen et al., 2008), based on the DSM-IV-TR, was used to screen subjects for PTSS. The PCL-C is a 17-question self-report with item responses measured on a 5-point Likert scale ranging from 1 (not at all) to 5 (extremely) (American Psychiatric Association, 2000; Blanchard et al., 1996; Keen et al., 2008). The PCL-C provides an overall measure of PTSD symptomatology as well as three subcluster scores. Overall PTSD symptomatology is scored by taking the sum of all questions with the result ranging from 17 to 85. The three symptom subcluster scores, based on DSM-IV-TR criteria (American Psychiatric Association, 2000; Blanchard et al., 1996; Keen et al., 2008), are re-experiencing (cluster “B,” questions 1–5), avoidance or numbing symptoms (cluster “C,” questions 6–12), and hyperarousal (cluster “D,” questions 13–17) (Blanchard et al., 1996; Keen et al., 2008). The PCL-C has been extensively validated in both civilian and military populations with test–retest reliability dictated by Pearson's r between 0.66 and 0.96, and internal consistency dictated by Cronbach's α ranging from 0.71 to 0.97 based on comparison to the “gold standard” of PTSD diagnosis, which is the Clinician-Administered PTSD Scale (CAPS) (Blanchard et al., 1996; Keen et al., 2008). However, because PTSD is a clinical diagnosis and cannot be obtained solely from self-report measures such as the PCL-C, in this article, the data obtained from the PCL-C measures are referred to as PTSS. Our use of the civilian version of the PCL is based on our desire to capture all potential emotional trauma and not restrict the query to only military emotional trauma as would be captured in the military version of the PCL.
MRI data acquisition
Image data in this study consisted of whole-brain T1-weighted anatomical scans (3D BRAVO sequence: TR/TE = 6.7/2.5 ms, FA = 12°, resolution = 1.0 × 1.0 × 1.2 mm) and fMRI task-free, “resting-state” data (echo planar imaging [EPI] pulse sequence, sagittal plane, TE/TR = 25/2000 ms, FA = 60°, in plane resolution = 3.75 × 3.75 × 4 mm, matrix size 64 × 64, and FOV 240 × 240) acquired using a 3T MRI scanner (GE Healthcare, Waukesha, WI) with a 32-channel phased-array head coil (MR Instruments, Inc., Minnetonka, MI). The task-free scan was 6-min long and was the first in the series of fMRI paradigms in the experiment protocol. All subjects were presented with the sequences in the same order and were instructed to close their eyes for consistency, to reduce distractions and potential eyestrain from static fixation for the entire duration of the scan. No survey was administered before or after the scan to assess the mental or emotional state of the subjects.
Data analysis
The Advanced Normalization Tools (ANTS) (Avants et al., 2011) software package was used to transform the anatomical T1-weighted images to the Montreal Neurological Institute (MNI) 152 template (Mazziotta et al., 1995). The Analysis of Functional NeuroImages (AFNI) software package (Cox, 1996) was used to correct head movement using three-dimensional (3D) volume registration with respect to the middle volume of the resting-state series as the reference; subsequently, the first three time points were discarded to account for magnetization steady state, and temporal interpolation was performed to correct for slice-time acquisition differences. Respiratory and cardiac signal effects were removed using the Physiological Noise Modeling (PNM) package (Brooks et al., 2008). Subsequently, the fMRI data were corrected using a B0-field map, aligned to the T1-weighted anatomical data set and then warped to the MNI-152 template (Mazziotta et al., 1995). A 6 mm full width at half maximum (FWHM) Gaussian kernel was used to smooth the data (Cox, 1996). Each individual data set was manually inspected for poor spatial alignment, improper B-field map correction, excessive motion, and inaccurate physiological correction. Motion parameters and their derivatives were regressed out from the EPI data, and a high-pass filter was applied (cutoff frequency >0.005 Hz) using the 3dDeconvolve program (Cox, 1996; Ward, 2002). The data were only high pass filtered to account for noise associated with scanner drift and some physiological processes (i.e., cardiac) (Niazy et al., 2011; Van Dijk et al., 2010). The data were not low pass filtered. While BOLD signals are often assumed to have largely low-frequency components, however, the true filter characteristics are not known at this moment and there could be contribution from higher frequency components to the BOLD signal (Niazy et al., 2011; Van Dijk et al., 2010), as determined after adequately accounting for artifacts (respiration, cardiac, motion, etc.). Furthermore, certain regions have been observed to have increased power (>0.005 Hz) at varying durations and frequencies (Handwerker et al., 2012).
Temporally concatenated probabilistic independent component analysis (PICA) was performed using the Multivariate Exploratory Linear Decomposition into Independent Components (MELODIC) (Beckmann and Smith, 2004) tool in the FSL software package (Smith et al., 2004) on the control subjects to obtain a control group ICA template to which the mTBI subjects could be compared. A brain mask was created and time series for each voxel within the mask were de-meaned and normalized by the voxelwise variance, followed by whitening of the data sets. The data were then decomposed into 25 independent components based on principal component analysis (PCA) estimates from the Eigenspectrum analysis. Each component map was subsequently normalized with respect to the standard deviation of the residuals (Beckmann and Smith, 2004). The component maps were examined, and the component matching the DMN based on the literature was extracted (Beckmann et al., 2005, 2009). Spatial and temporal maps for each individual control and mTBI subject were obtained using the dual-regression ICA tool employing multiple linear regression to estimate spatial and temporal interactions at the individual subject level using regression performed against the original data (Beckmann et al., 2009). The correlation values for each individual subject data set were converted to z-scores and resampled to a resolution of 1.0 × 1.0 × 1.0 mm. Subsequently, voxelwise t-tests (Cox, 1996) in which age and years of education were covaried out were used to compare differences between controls, the high, medium, and low PTSS mTBI groups. The comparisons between low, medium, and high PTSS mTBI groups also included time since injury as an additional nuisance regressor. The noise smoothness of each individual subject data set was estimated using the 3dFWHMx AFNI program (Cox, 1996) with the spatial autocorrelation function or “acf” option (Cox et al., 2016). Subsequently, the average of the estimated autocorrelation function parameters for all subjects obtained using the 3dFWHMx program was entered into the 3dClustSim, which was used with the “acf” option (Cox, 1996; Cox et al., 2016). The estimated effective FWHM was 12.35 mm. The group difference maps were thresholded using a corrected p-value <0.05 and a cluster size >560 voxels (1 mm3 each) as determined by parametric and spatial thresholds using 10,000 Monte Carlo simulations (Cox, 1996; Cox et al., 2016; Forman et al., 1995).
Results
The results presented are all voxelwise corrected at p < 0.05, based on initial p < 0.005 and a cluster size >500 voxels. In reporting our results, we focus on presenting “connectivity” as the average Z-scores of correlation values within regions significantly different in the group comparisons.
Comparison of controls and PTSS mTBI groups
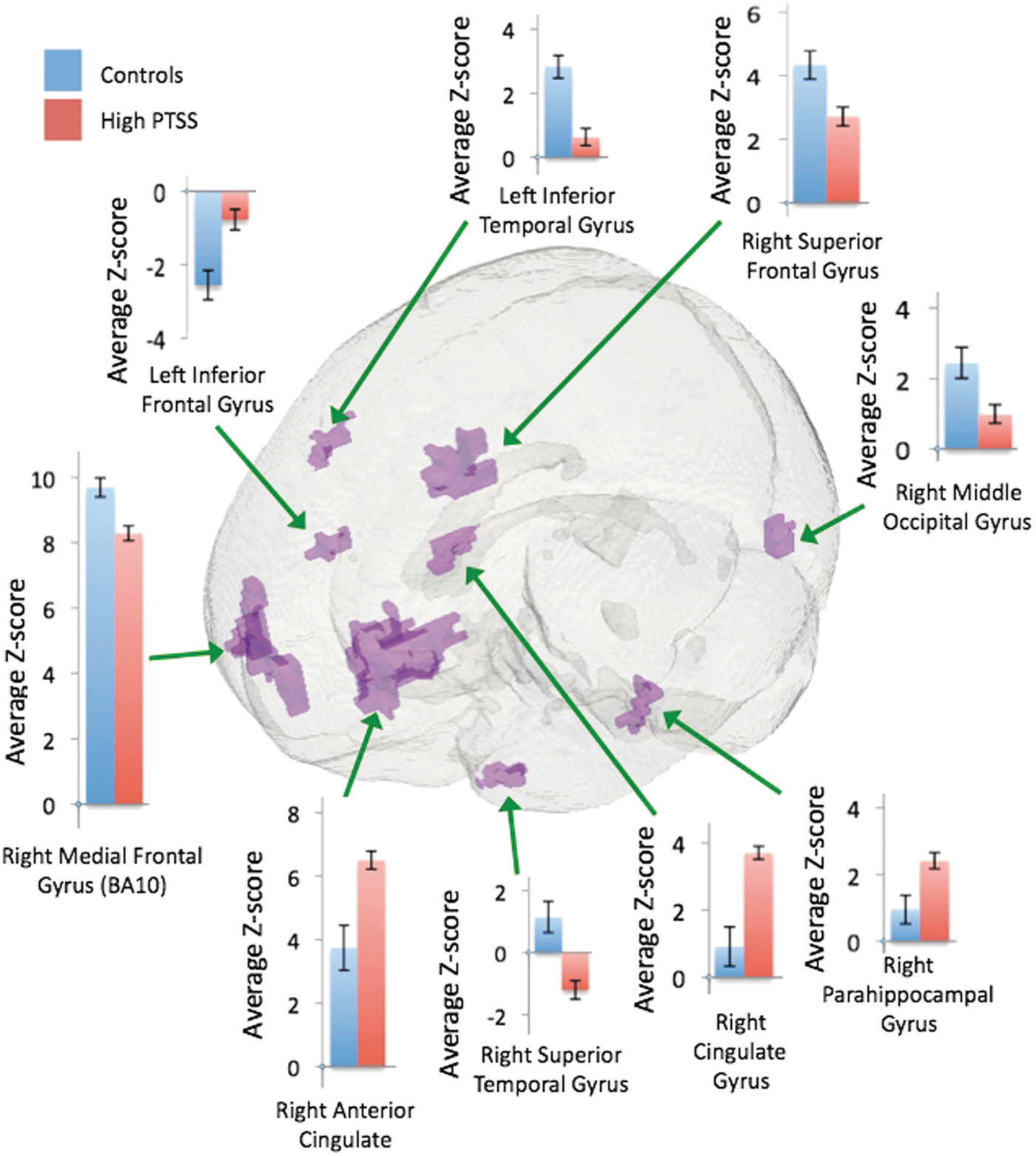
Distinct connectivity patterns were observed when comparing the control group to each PTSS mTBI group. The results indicate that connectivity in the medial frontal gyrus (Brodmann area 10 [BA10]) was higher in the control group compared to each of the PTSS mTBI groups. The center of mass for these regions was mostly in the right cortex, except for the low PTSS mTBI subjects; however, the region of significant difference in connectivity encompassed both left and right hemispheres. In addition, the anterior cingulate had higher connectivity in each of the PTSS mTBI groups in contrast to the control group. The fusiform gyrus and a region within the cingulate cortex were also observed to have higher connectivity in the high and medium PTSS mTBI groups when compared to the control group. These regions were different between the controls and the low PTSS mTBI group but did not survive statistical significance. Comprehensive differences (cluster size and t-statistic) from the voxelwise t-test between subject groups are presented in Table 2 and illustrated in Figures 1 –3.
Corrected voxelwise t-test comparing the control group and high PTSS mTBI subjects. mTBI, mild traumatic brain injury; PTSS, post-traumatic stress symptoms. Color images available online at

Corrected voxelwise t-test comparing control group and medium PTSS mTBI subjects. Color images available online at

Corrected voxelwise t-test comparing the control group and low PTSS mTBI subjects. Color images available online at
BA10, Brodmann area 10; mTBI, mild traumatic brain injury.
Comparison of mTBI PTSS groups
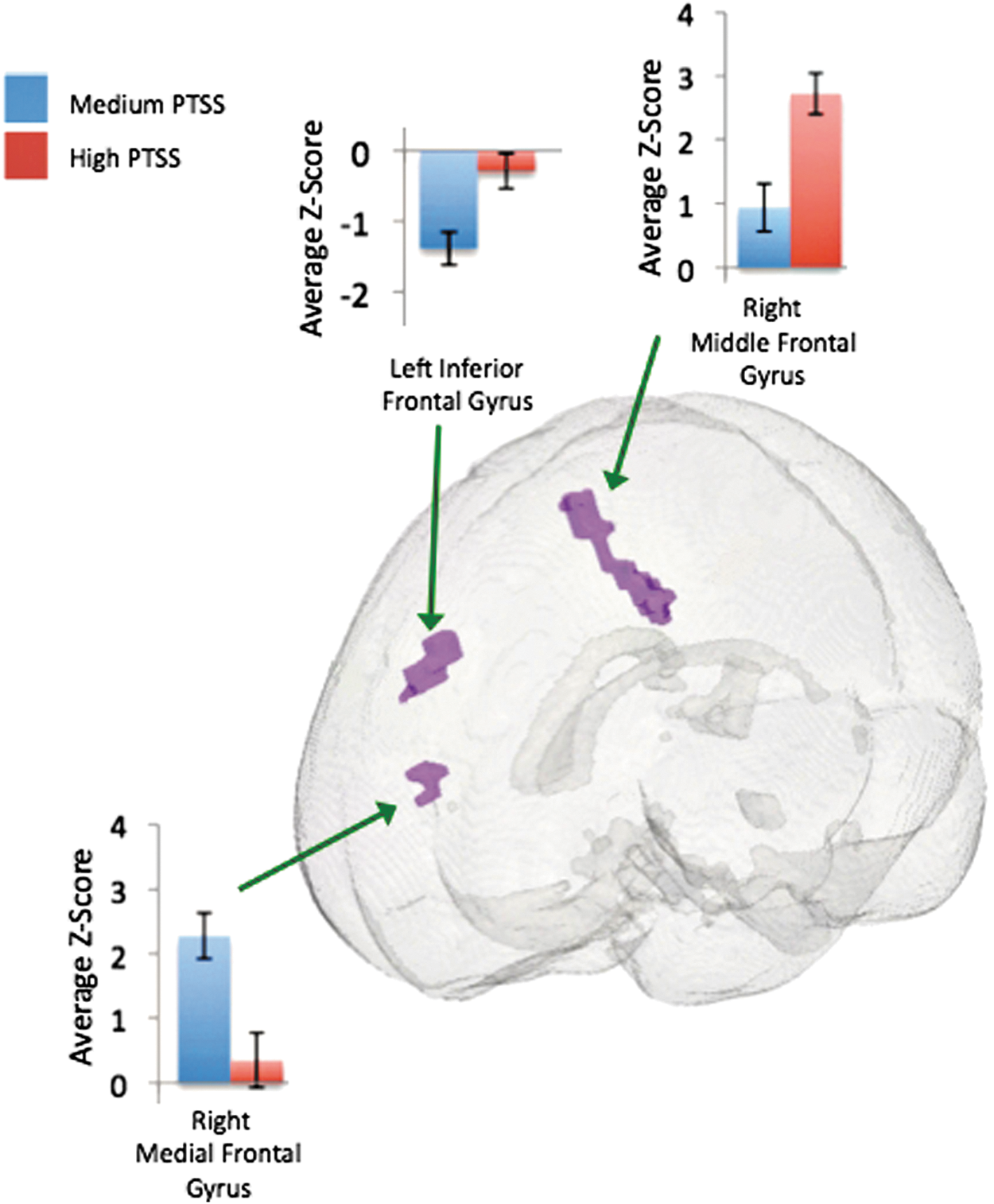
The results indicate that regions of the left parahippocampus, left middle frontal gyrus, and left cuneus had higher connectivity within the high PTSS mTBI group in contrast to the low PTSS mTBI group. The low PTSS mTBI group had higher connectivity in the right middle temporal gyrus and left middle occipital gyrus in contrast to the medium PTSS mTBI group that had higher connectivity in the right cerebellar lobule VIIa Crus 1. Finally, higher connectivity in the right medial frontal gyrus and left inferior frontal gyrus was observed in the medium PTSS group when compared to the high PTSS mTBI group. In contrast, the high PTSS group had higher connectivity in the right middle frontal gyrus when compared to the medium PTSS group. Comprehensive differences (cluster size, t-statistic, and effect size) from the voxelwise t-test between PTSS subject groups are presented in Table 3 and illustrated in Figures 4 –6.

Corrected voxelwise t-test comparing the low with the high PTSS mTBI subject group. Color images available online at

Corrected voxelwise t-test comparing the low with the medium PTSS mTBI subject group. Color images available online at
Corrected voxelwise t-test comparing the medium with the high PTSS mTBI subject group. Color images available online at
Discussion
The main objective of this study was to understand how task-free DMN connectivity is affected by PTSS in chronic mTBI subjects. The results indicate distinct connectivity patterns between each of the PTSS mTBI groups when compared to the control group, and differences also in DMN connectivity when comparing only the PTSS mTBI groups. The first key finding is all of the mTBI subjects, irrespective of PTSS severity when compared to the control group, had significantly lower connectivity in a region centered at the anterior prefrontal cortex (APC), in BA10, and extending into both the left and right medial frontal gyrus. The APC (BA10) is involved in a variety of cognitive functions that include the regulation of attention, emotion, memory, and mental states (Franklin et al., 2011). Damage to this area has been associated with compromised executive function pertaining to attention, concentration, decision-making, and response consistency (Beckmann et al., 2005, 2009; Gilbert et al., 2006). These are salient features that suggest the APC and parts of the frontal gyrus are involved in the regulation of state of mind and attention (Gilbert et al., 2006), and previously been observed as a key region of the task-free DMN (Beckmann et al., 2005, 2009).
The second key finding when comparing the control group to each of the PTSS groups is higher connectivity in the inferior portion of the anterior cingulate cortex (ACC). Specifically, connectivity in the ACC was highest in the high PTSS group, followed by the medium and low PTSS groups in contrast to the control group. The connectivity difference in the ACC is located in the subdivision of the ACC that is suggested to be responsible for affective processing and is heavily engaged in executive function, specifically in the regulation and evaluation of information and responses relating to emotion and motivation (Bush et al., 2000). Damage to the ACC has been reported to cause various emotional symptoms such as lability and apathy in addition to inattention and the dysregulation of autonomic functions (Bush et al., 2000), which are chronic symptoms present in some TBI patients (Cifu and Caruso, 2010).
The APC and ACC are anatomically connected and there is some functional overlap, however, the exact relationship between these two regions is not clearly understood at present. Prior studies show that the ACC and regions of the lateral prefrontal cortex and BA10 work in a concerted effort during various cognitive and mental processes (Burgess et al., 2007). It has been suggested that the APC may serve as a controller for response schema selection pertaining to conscious state of mind, specifically with respect to introspective stimuli such as memories, ideation, and perception (Burgess et al., 2007). In this capacity, it is hypothesized that the APC serves as a “gateway” that regulates information flow and resources of other regions and brain networks (Burgess et al., 2007). Depending on the stimuli, environmental conditions, and schema selected, the APC could regulate information flow to parallel and dispersed brain networks involved with attention and emotional processing, of which the ACC is an instrumental part (Burgess et al., 2007). The reduced connectivity in the APC among all the mTBI groups, irrespective of PTSS severity when compared to the control group, suggests an effect of mTBI that is disrupting the proposed “gateway” function of the APC leading to disparities in information flow to various brain networks and inhibiting proper “switching” in the context of conscious “relaxed” state of mind without explicit stimuli. In addition, the lack of control from the APC could lead to dysregulation of the ACC, resulting in hyperconnectivity that may be indicative of impairments. Hyperconnectivity in the ACC during task-free studies has been observed in subjects with cognitive disorders such as attention deficit hyperactivity disorder (ADHD) (Tian et al., 2006), major depression (Zhu et al., 2012), and in subjects who are at high risk for developing psychosis (Shim et al., 2010). In addition, hyperconnectivity has been also observed in subjects with PTSD and social anxiety during task performance and is thought to reflect increased efforts at task performance, maintaining attention or hypervigilance (Shaw et al., 2009; Simmons et al., 2008). Alternatively, the hyperconnectivity in the ACC could be indicative of a compensatory mechanism that seeks to modulate intrinsic representations of emotion and memory in the absence of explicit external triggers and could reflect a higher effort needed to maintain the “relaxed” state of mind.
However, the ACC and APC do not work in isolation, but rather as part of the DMN, and serve in some capacity to regulate emotional balance and state of mind by integrating current information pertaining to the environment, current state of mind, and past experiences, that is, memories. The temporal lobes are instrumental in memory formation and long-term storage related to associative learning, familiarity, and recollection (Eichenbaum et al., 2007; Squire et al., 2004). In this functional capacity, the temporal lobes belong to larger networks that include cortical and subcortical regions that are involved with cognitive functions relating to the parsing of memory elements that drive learning, recall, decision-making and task performance (Eichenbaum et al., 2007; Squire et al., 2004).
The third key finding pertains to distinct connectivity patterns within the temporal regions of the brain when comparing the control group to each of the PTSS mTBI groups. The control group had higher connectivity when compared to the high PTSS mTBI group and the low PTSS group, but the control group had lower connectivity in contrast to the medium PTSS group, except in the left middle temporal gyrus. In addition, some regions such as the right superior temporal gyrus, right inferior temporal gyrus, left middle temporal gyrus, and right medial temporal gyrus had opposite correlation polarities (anticorrelations) when comparing the PTSS mTBI groups with the controls. This opposite correlation pattern was also present when comparing the low and medium PTSS groups (other PTSS mTBI comparisons did not survive correction). Lower connectivity within the temporal lobes has been observed in resting-state networks of acute PTSD subjects (Lanius et al., 2010). In a task-based study comparing veterans with PTSD and controls, increased activation of the temporal lobes was observed in the PTSD veteran group during memory encoding, and conversely during voluntary retrieval, there was a reduction in activation of the temporal gyri (Geuze et al., 2008). The differences in temporal lobe connectivity between the controls and the PTSD subjects could be indicative of disruption pertaining to the ability to appropriately parse sensitive emotional stimuli with memory correlates of past traumatic events, therefore leading to altered encoding and retrieval. Specifically, it is proposed that trauma response can consist of two possible mechanisms, the first is dissociative and the second is hyperarousal (Bremner and Marmar, 2002). fMRI studies investigating these responses have reported that an increase in activation was present in dissociative subjects in regions of the temporal gyri and also including the medial frontal and parietal gyri and anterior cingulate, however, nondissociative PTSD subjects do not have these similar activation patterns (Lanius et al., 2002). Since we did not explicitly screen our subjects for PTSD response type, this and the chronic effects of PTSD could be a source of variation in the temporal lobe connectivity patterns present and warrant further research.
The fourth key finding is significantly higher connectivity in the parahippocampus of high PTSS mTBI subjects compared to the low PTSS mTBI subjects and the control subjects. The parahippocampus is closely associated with the temporal lobes and actively involved with memory encoding and subsequent retrieval (Eichenbaum et al., 2007; Lanius et al., 2002; Squire and Zola-Morgan, 1991). The present findings from task-free DMN complement prior research findings of increased activation observed in the parahippocampal region in PTSD subjects during direct exposure to negative stimuli (Lanius et al., 2002; Sakamoto et al., 2005) and both successful and false recall (Thomaes et al., 2009). Studies have reported that abnormal connectivity in the hippocampus and parahippocampus is the result of disruptions in episodic and autobiographical memory encoding and storage, and that this region may be a part of the temporal lobe network, therefore being abnormally active in PTSD (Eichenbaum et al., 2007; Lanius et al., 2002; Sakamoto et al., 2005; Squire and Zola-Morgan, 1991; Thomaes et al., 2009). We noticed several of the regions from the group having temporal anticorrelations. These temporal differences could be attributed to slower processing and information flow across brain regions due to the effects of chronic mTBI or, alternatively, could reflect frontal regions (BA10 and ACC) driving executive function for regulating information from regions such as the temporal lobes or visual cortex that may pertain to past or recurrent memories. However, the exact behavioral correlates of these anticorrelated regions are not clear at the moment and warrant more research.
The results do not indicate any significant involvement of the amygdala as previously reported in PTSD studies (Bruce et al., 2013; Hayes et al., 2011; Sripada et al., 2013). We attribute these differences to the fact that connectivity in the amygdala and regions pertaining to fear and PTSD responses reported in prior studies was elicited through experimental paradigms that employed explicit stimuli used to invoke emotional responses (Bruce et al., 2013; Hayes et al., 2011; Sripada et al., 2013). Conversely, task-free studies have also used seed-based correlations or region of interest analysis in which the amygdala was the point of focus, and this may have limited the reported results (Rabinak et al., 2011; Sripada et al., 2012). The amygdala has been reported to be instrumental in the development of PTSD and maintenance of PTSD symptoms (Liberzon and Sripada, 2007). PTSD disrupts several large distributed networks that are interconnected with the amygdala. It is proposed that these disruptions pertain to compromised “top–down” modulation of emotional, memory, and response systems, of which key components include the amygdala and other structures of the limbic system (Liberzon and Sripada, 2007; Nathan et al., 2015; Shin et al., 2006). Our results support this notion of disrupted “top–down” modulation, not limited to a single region, but rather a network of regions. Specifically, the differences attributed to PTSS such as varying connectivity in the temporal lobes, differences in the parahippocampal gyrus, ACC, and regions of the frontal gyrus could be attributed to cognitive deficits that pertain to networks involved with attention and memory encoding, which could be reflected in PTSS of increased re-experiencing and hyperarousal that hinder proper switching of conscious state of mind. Alternatively, the decrease in amygdala activation was also reported in dissociative PTSD subjects and it thought that the lack of “top–down” control from the frontal regions pertaining to emotional processing affects the limbic system in the brain and causes a state of hypoarousal (Lanius et al., 2002). On the contrary, these differences could be attributed to compensatory mechanisms in which increased connectivity in the task-free DMN of the high PTSS group could be indicative of increased effort needed by specific brain regions to suppress unwanted intrusive thoughts and to maintain a more “relaxed” state of mind in the absence of explicit stimuli. However, it is unclear to what extent mTBI affects regulatory cognitive mechanisms, chiefly “top–down” modulation involving frontal regions of the brain such as BA10, and medial frontal gyrus, and how information from these regions is compromised to exacerbate PTSS. The specific incorporation of task-based paradigms or the use of causal connectivity analysis such as Granger causality (Nathan et al., 2015) would complement these findings and help to explicitly probe information flow and subsequent regulation within these cognitive networks.
The preprocessing of our data sets did not include scrubbing, due to the potential risk of introducing high-frequency noise contamination that could lead to spurious correlations (Carp, 2013). Scrubbing has been found to remove some types of noise but not others and could result in the removal of a large number of time points (Yan et al., 2013b). The removal of varying portions of time points between subjects could lead to increased connectivity disparities across subjects (Yan et al., 2013b). Furthermore, the reported removal of negative correlations by scrubbing (Yan et al., 2013b) could bias the results due to removal of negative correlations, which may have potential neuronal origins; in addition, an objective manner to clearly distinguish low or high signal fluctuations of neuronal origin compared to artifact is still an area of intense research interest, and optimization. Most large motion artifacts that scrubbing aims to remove can be detected and removed with ICA without sacrificing temporal degrees of freedom and destroying the temporal autocorrelation structure of the data sets (Pruim et al., 2015), features that affect function connectivity measures. However, scrubbing may have utility in seed-based correlation analysis (Yan et al., 2013b), but caution should be exercised for the aforementioned reasons.
There are several important limitations to this study. First, the subjects used in this study were not “pure” mTBI subjects, but were obtained from a clinical population with heterogeneous symptoms (e.g., pain, sleep disruption). Second, the regulation of DMN activity may have been affected by sleep disruption; however, information about these symptoms obtained from the Neurobehavioral Symptom Inventory and PCL-C for the groups showed very mild sleep disturbance and did not approach significant levels. Third, subjects recruited in this study were receiving ongoing clinical care for injuries experienced in addition to the TBI and therefore were prescribed medications. Some medications could alter cerebral perfusion and DMN connectivity; however, the extent of alterations within the DMN is inconsistent across brain regions and understanding the effects of specific medications or the interaction of multiple medications is highly challenging, as various medications could have opposing effects when taken together (Franklin et al., 2011; Vlassenko et al., 2004). Fourth, substance use (i.e., nicotine, alcohol, or caffeine) could induce additional variance in the DMN data. Fifth, there is no objective way at present to quantify or control for the intensity of the injury mechanism or the severity of injury symptoms. These features make analysis with chronic mTBI subjects challenging and could have contributed toward the variability in our data set. Sixth, the control group had a higher number of female participants and some civilians. These two factors could potentially relate to some variability of the connectivity patterns within the control group, however, definitive differences between resting-state functional connectivity and gender are an ongoing research effort and future work would focus on better matching the gender of subjects recruited for the study, and the elimination of civilians. Seventh, the imaging results from pairwise group comparison t-tests were not corrected for multiple comparisons; perhaps the use of ANOVA or linear mixed effect models in future analysis with multiple comparisons corrections may help to identify within-group features in contrast to pairwise t-tests. Finally, the self-report measures could be susceptible to individual bias in symptom reporting. Therefore, future studies should incorporate more objective measures of symptoms.
Footnotes
Acknowledgments
The views expressed in this article are those of the authors and do not reflect the policy of the United States Department of the Army, Navy, Air Force, Department of Defense, or the United States Government. This study was supported through the Congressionally Directed Medical Research Program, grant number DM130132. The authors are grateful to Mr. David Joy, BS, Dr. Joseph Hennessey, PhD, Mr. Andrew Bryant, Mr. Alex Kubli, BS, Mr. John Morissette, Mrs. Jamie Harper, MPH, Mr. Justin Senseney, MS, Ms. Rachel Wolfowitz, BS, Mr. Arman Taheri, the participants and their families, and to the clinicians and staff at the National Intrepid Center of Excellence.
Author Disclosure Statement
No competing financial interests exist.