
Abstract
When the brain is not engaged in goal-directed activities and at rest, there are still measureable patterns of activity. One resting-state network, the default mode network (DMN) is responsible for a self-referential introspective state. There are many factors that influence normal changes in brain activity. The purpose of this review is to summarize differences in DMN functional connectivity in healthy individuals by age, sex, cognitive function, and analysis type to characterize what is “normal.” Studies were systematically selected up to August 2016. Two reviewers independently used predetermined inclusion and exclusion criteria to identify relevant studies. Studies that provided sufficient information were included in a subsequent voxel-wise meta-analysis. Strength of DMN functional connectivity follows an inverse U-shape, where it is strongest in adulthood and lowest in children and elderly. Cognitive function is positively correlated with DMN functional connectivity. Females exhibit stronger intranetwork connectivity compared with males. Effects of analysis type were inconclusive and more studies need to incorporate complementing techniques. The voxel-wise meta-analysis was only conducted for the age factor. Findings supported an immature network in children compared with adults and a stronger network in adults compared with elderly. This is the first study to review differences of DMN functional connectivity in healthy individuals by age, sex, cognitive function, and analysis type. Findings add to the understanding of normal variance. Furthermore, defining a normal comparative base may allow for the identification of DMN change into pathology. This is important since it may allow for the detection of an intermediate risk phenotype and could serve as a biomarker for treatment response.
Introduction
Default mode network
W
Further work observed dampening of DMN activation upon initiation of a task performance leading to the coining of “task-negative network,” which postulates that the DMN is responsible for a self-referential and introspective state (Andrews-Hanna et al., 2010; Greicius et al., 2003; Long et al., 2008; McKiernan et al., 2003). This functional theory has been supported with studies showing that DMN suppression will increase with task difficulty suggesting that introspective attentional resources must be reallocated to focus on an extrinsic task (Singh and Fawcett, 2008). Evidence also suggests that the DMN mediates the process of one's thoughts and feelings, which is an important factor for an individual to remember one's past, form beliefs, and plan for the future (Buckner et al., 2008; Raichle and Snyder, 2007). Studies requiring participants to recollect a memory show activation of the DMN, which has been postulated to show the individual “retrieving” the past experience (Daselaar et al., 2009).
Factors accounting for differences observed through neuroimaging
Age and cognitive function
Aging is a normal physiological process and understanding its influence on the brain's architecture and functional connectivity is essential. Pathways involved with emotion processing such as prefrontal control over the amygdala differ in healthy children and adults. In particular, amygdala activation is increased in adults, but decreased in children in response to a frustrating event (Perlman and Pelphrey, 2011). Interestingly, when comparing healthy adults and older adults there is a consistent age-related decrease in amygdala activation in response to negative stimuli (Nashiro et al, 2011). Furthermore, understanding and subsequently defining normal age progression from adult to elderly populations is crucial. With healthy aging there is a natural loss of gray matter especially in the lateral prefrontal cortex, hippocampus, cerebellum, and caudate nucleus and white matter in the prefrontal cortex (Reuter-Lorenz and Lustig, 2005). However, there is an interesting phenomenon, where there is an age-dependent region-specific overactivation for processes involved with executive function, motor control, and episodic and autobiographical memory, where activation levels and performance are positively correlated (Reuter-Lorenz and Lustig, 2005). Healthy young adults exhibit functional connectivity between the hippocampus and ventral prefrontal cortex predict performance on a memory encoding task, whereas in older adults between the hippocampus and dorsolateral prefrontal cortex suggesting a shift in cognitive resources with age (Grady et al., 2003). Therefore, there is a need to better understand differences in functional connectivity and in particular at rest for normal aging processes.
Sex
The brain is sexually dimorphic in morphology, BOLD activation, and functional connectivity. For example, regions involved in the DMN such as the PCC are larger in females than males (Goldstein et al., 2001). Furthermore, there are sex differences in response to positive mood induction such that females experience greater BOLD signals in the right PCC compared with males (Hofer et al., 2006). In a large study of 603 subjects it was found that there is stronger intranetwork connectivity in females and stronger internetwork connectivity in males especially in resting-state sensorimotor networks (Allen et al., 2011). Current findings that support sex differences in brain function and morphology provide strong evidence to further investigate if these differences exist in the DMN.
Methods of analysis
There are four main approaches to analyze resting-state functional connectivity—seed-driven analysis (Fox et al., 2005), independent component analysis (ICA) (Beckmann et al., 2005; Greicius et al., 2007), regional homogeneity (ReHo) (Zang et al., 2004), and functional connectivity density mapping (FCDM) (Tomasi and Volkow, 2010).
Seed-driven analysis requires an a priori selection of a voxel, cluster, or anatomical brain atlas region and then calculates the Pearson's correlation coefficients between the seed brain region of interest and all other voxels in the brain, thus producing correlation maps (Fox et al., 2005). The correlation coefficients are then converted to normally distributed Z-scores using a Fisher's transform to allow for second-level general linear model analysis (Fox et al., 2005; Whitfield-Gabrieli and Nieto-Castanon, 2012). The main advantage over other methods is that it provides a direct and easily interpretable answer since it shows the network of regions most strongly functionally connected with the seed. The main disadvantage with seed-based analyses is the influence of noise (scanner or participant based) (Cole et al., 2010). Furthermore, seed-based analysis is a data-driven model that assumes that each brain region will have the same hemodynamic response, which is the physiological basis behind fMRI BOLD responses (Moritz et al., 2000).
ICA assumes that there is an archetypal spatial pattern seen in resting state allowing it to be reliably detected using a template-matching procedure for each subject (Greicius et al., 2007). Without any a priori assumption on the hemodynamic model, ICA is a data-driven method that can gather more information than a model-driven method. In particular, it can identify nonanticipated or transient task-related components. ICA avoids prior spatial and noise assumptions associated with seed-based analysis and has the ability to simultaneously compare the coherence of activity in multiple distributed voxels (Cole et al., 2010). However, there are indications that ICA cannot discriminate between inter- and intraregional functional connectivity (Onoda et al., 2012).
ReHo is a voxel-based analysis measure of brain activity, which evaluates the similarity or synchronization between the time series of a given voxel and its nearest neighbors (Zang et al., 2004). The main advantage of ReHo analysis is that it overcomes the need for knowledge of the experimental design since the correlation of activation at a specific time is simply compared individually to its neighboring voxels that gives the size of activation. This is accomplished by calculating the Kendall's coefficient of concordance of activated clusters (Zang et al., 2004). Often, ReHo is not used as a substitute for data-driven models, but rather as a complementary method since it may be better at inferring short-distance connectivity within a brain region (Jiang and Zuo, 2016).
FCDM is a new data-driven voxel-wise method that can very quickly identify hubs in resting-state networks (Tomasi and Volkow, 2010). FCDM was not designed to be an independent measure of functional connectivity, but rather one that is complementary to seed-driven analyses. FCDM provides a voxel-wise measure of the number of functional connections, but cannot provide directionality (Tomasi and Volkow, 2010). Thus, the proposed use of FCDM is to identify hubs that can be subsequently used as seed locations for model-driven analyses (Tomasi and Volkow, 2010).
The purpose of this systematic review is to summarize DMN activity and functional connectivity in healthy subjects based on factors such as age, sex, cognitive function, and analysis type.
Methods
Search strategy
Relevant studies were searched systematically up to August 2016. Studies were identified through MEDLINE, PsycINFO, EMBASE, and the grey literature. Reference lists from identified articles and relevant reviews were searched for articles not found in the above databases. Search terms were: magnetic resonance imaging, DMN, sex characteristics, and limits were the following age groups: 0–18 years, 19–44 years, and 45 plus years. A total of 963 titles were initially identified and following predetermined inclusion and exclusion criteria, two reviewers (L.M. and R.M.) determined that 30 were suitable to be included in the review.
Selection criteria and quality assessment
Two reviewers L.M. and R.M. independently used predetermined inclusion and exclusion criteria to review the identified titles and abstracts. Disagreement between reviewers was 0.93%, discrepancies were further discussed with a 100% agreement rate. Studies were included if they (1) used a nonclinical sample, (2) included comparators such as age or sex or analysis type or cognitive function, (3) looked at the DMN measured by BOLD response using functional MRI, (4) observational studies that were either cross-sectional or longitudinal in design, (5) used seed-based analysis or voxel-wise driven analysis or ICA or ReHo, and (6) published in a peer-reviewed journal. Studies were excluded if they included a clinical sample, did not include any comparator factors, investigated resting-state functional connectivity that was not the DMN, was an interventional study, did not report brain regions that differed by comparator factor, or was not in English. The process of study inclusion and exclusion is summarized in the PRISMA flow chart (Fig. 1).
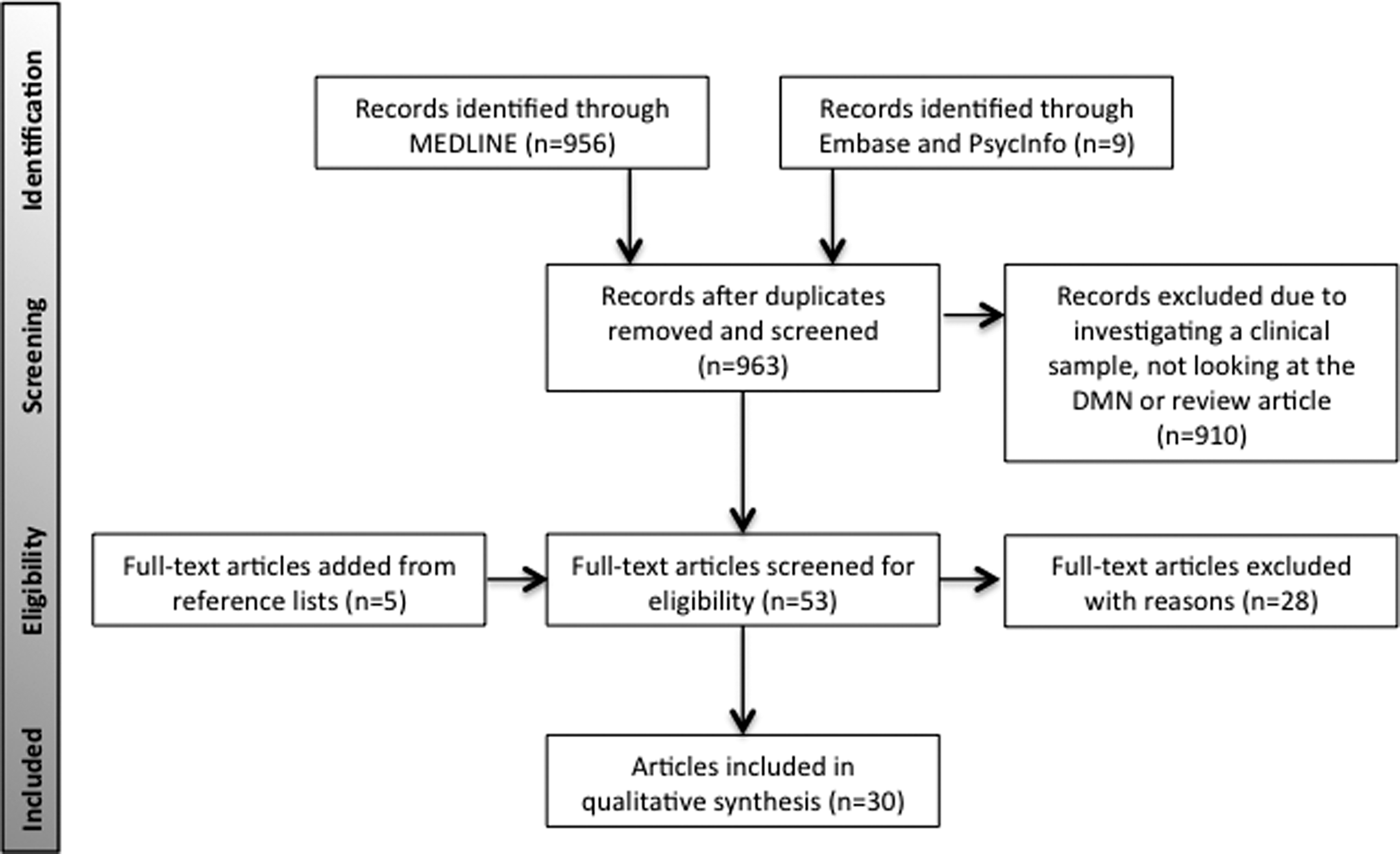
PRISMA flowchart depicting selection of included studies in final analysis.
Voxel-wise meta-analysis
Studies with reported Montreal Neuroimaging Institute (MNI) or Talairach coordinates and t-values or z-scores were included in the meta-analysis. Coordinates were all converted to MNI coordinates and z-scores were converted to t-values. Seed-based d Mapping (SDM) was used to analyze DMN by groups and between groups. SDM methods are described in detail elsewhere (Radua and Mataix-Cols, 2009). Once coordinates are selected and converted, a map of functional differences is recreated for each study. A unique component of this program is it combines both positive and negative coordinates in the same map resulting in a signed differential mapping. Once each map is created, a meta-analytic map is calculated. Threshold for significance was set at p < 0.005 with a cluster size greater than 10 voxels. These values were based on the standard values of the SDM program and were determined to optimize sensitivity and specificity (Radua et al., 2012). Studies were combined only if they employed the same analysis technique. For example, seed-based analysis studies were not combined with ICA studies.
Results
Age
Preterm to adolescence and children versus adult
Evidence supports the presence of premature resting-state networks that are not the DMN in infants as young as 25 weeks (Fransson et al., 2007). Preterm infants present with resting-state networks that are less mature and/or abnormally formed (Smyser et al., 2010). By 4 years of age, there is marked increase in connectivity and the DMN becomes similar to what is observed in adults (Damaraju et al., 2014; Gao et al., 2009; Lee et al., 2013). Despite the similarity, the DMN in children between the ages of 5 and 13 is present, but in an immature state (de Bie et al., 2012; Thomason et al., 2008), where there is increasing integration between regions of the DMN over time (Sherman et al., 2014). Only two studies (Sherman et al., 2014; Thomason et al., 2008) were included in the further meta-analysis and only the first time point was used in the Sherman and colleagues (2014) study. Findings support the premature state of the DMN (Fig. 2; Table 1).
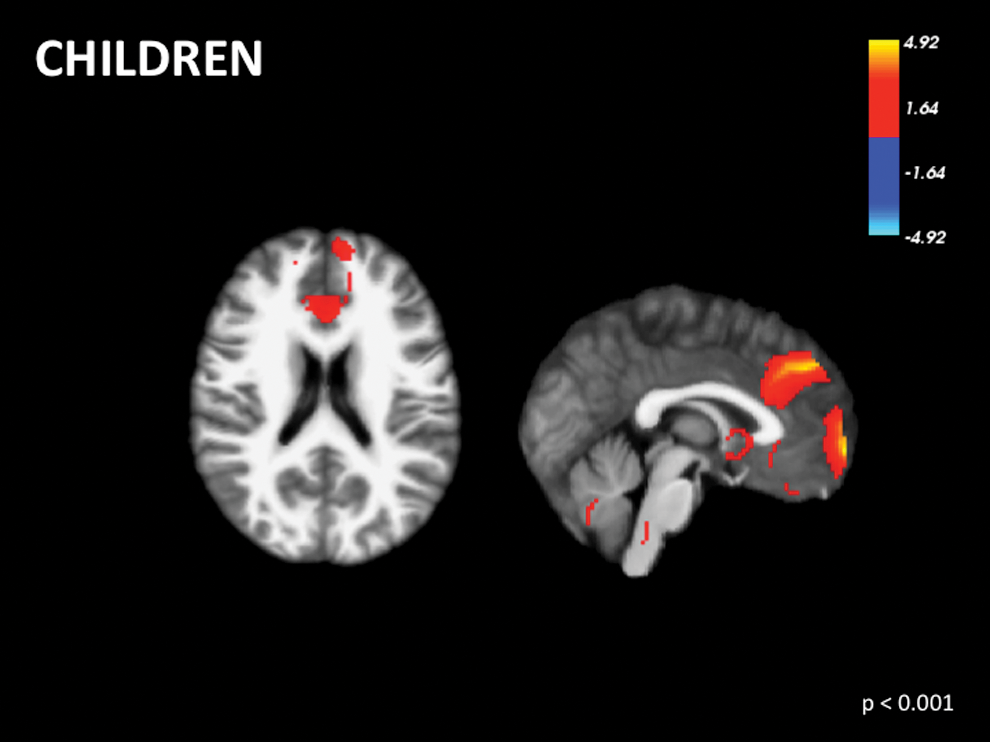
Voxel-wise signed differential mapping meta-analysis of the default mode network in children. Color images available online at
Reported coordinates are in Montreal Neuroimaging Institute space.
SDM, signed differential mapping.
There is conflicting evidence in the literature comparing children and adults. Some studies suggest that DMN regions are more sparsely functionally connected in children (Fair et al., 2009; Supekar et al., 2010). In contrast, other studies suggest similar patterns of functional connectivity and ReHo in adults and children, but strength is greater in children (Jolles et al., 2011; Lopez-Larson et al., 2011) (Supplementary Table S1; Supplementary Data are available online at
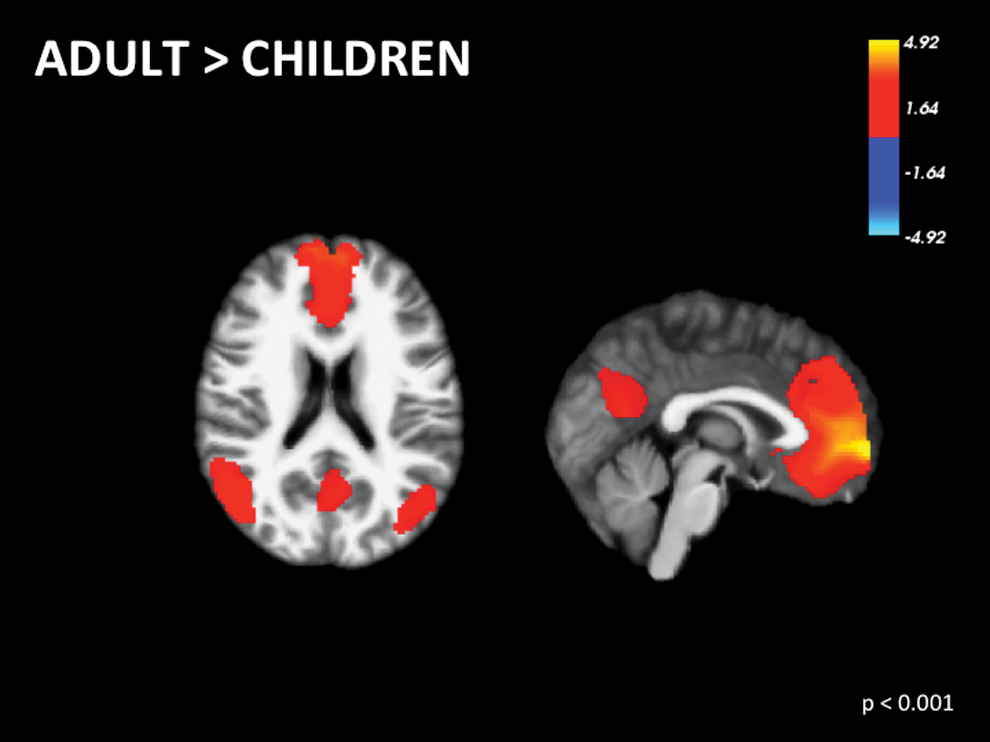
Voxel-wise signed differential mapping meta-analysis of functional differences in the default mode network between adults versus children. Color images available online at
Adult versus elderly
Even in middle-aged adults, DMN functional connectivity is inversely correlated with increasing age (Bluhm et al., 2008; He et al., 2013). However, certain areas such as the thalamus and left superior frontal gyrus are positively correlated with age (Bluhm et al., 2008). There is decreased activity (Damoiseaux et al., 2008; Koch et al., 2010; Persson et al., 2014; Sambataro et al., 2010; Wu et al., 2011) and functional connectivity between regions of the DMN with age (Grady et al., 2010; Madhyastha and Grabowski, 2014; Mevel et al., 2013; Ng et al., 2016; Tomasi and Volkow, 2012) (Supplementary Table S2). Two studies (Bluhm et al., 2008; Persson et al., 2014) were included in the meta-analysis assessing adults. Findings support a fully formed and integrated DMN by adulthood (Fig. 4; Table 1). Furthermore, three studies (Grady et al., 2010; Mevel et al., 2013; Wu et al., 2011) were included in the meta-analysis comparing adults and elderly individuals. The right precuneus and left superior frontal gyrus show greater activation in adults versus elderly adults (Fig. 5; Table 1).
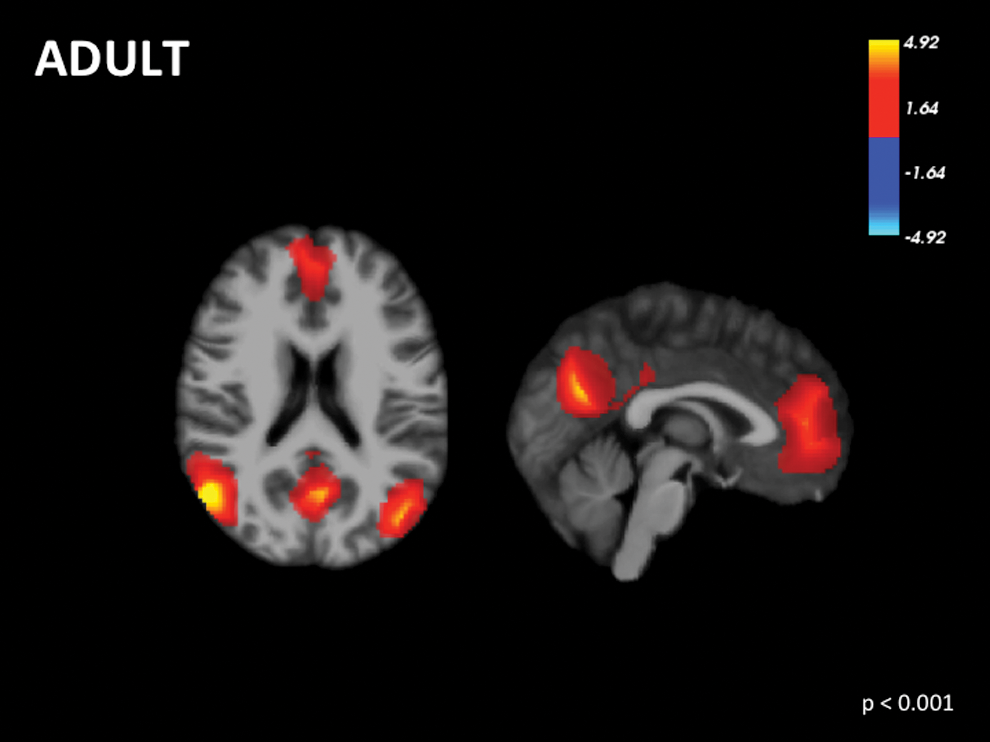
Voxel-wise signed differential mapping meta-analysis of the default mode network in adults. Color images available online at
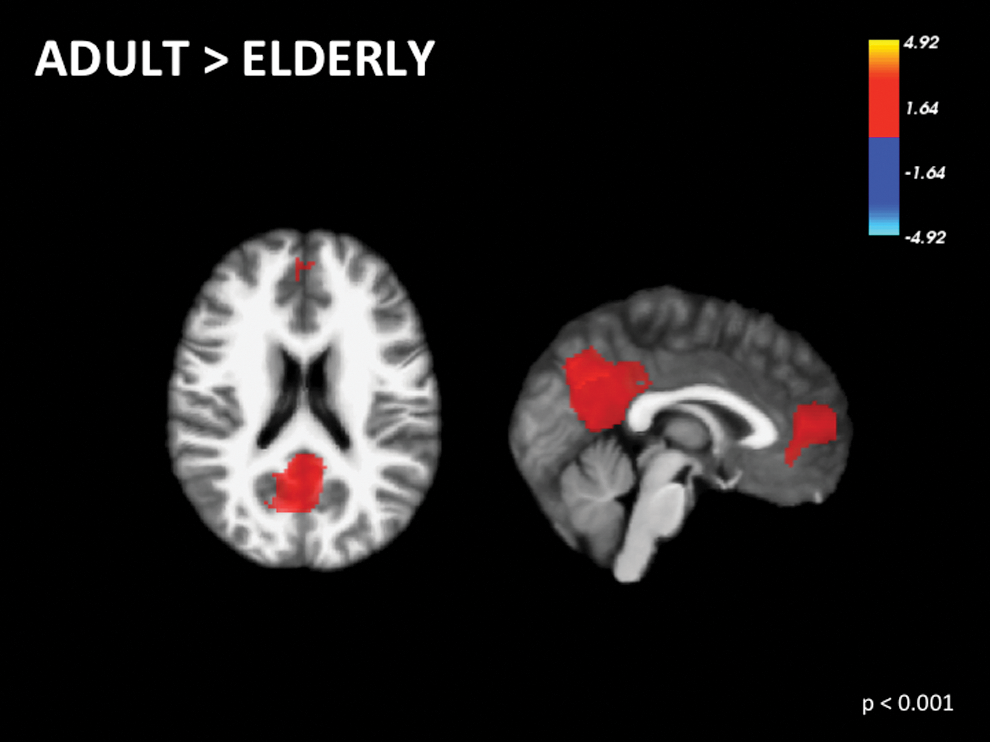
Voxel-wise signed differential mapping meta-analysis of the functional differences in the default mode network between adults versus elderly. Color images available online at
Sex
Some studies described a higher degree of functional connectivity in women between the PCC/precuneus and prefrontal cortex (Bluhm et al., 2008; Tomasi and Volkow, 2012). Regardless of age, females have greater local connectivity in the medial temporal regions (Lopez-Larson et al., 2011). However, one study found no significant functional connectivity differences in the DMN by sex (Weissman-Fogel et al., 2010) (Supplementary Table S3).
Cognitive function
Overall, studies suggest that cognitive performance is positively correlated with DMN functional connectivity (Hampson et al., 2006; Mevel et al., 2013; Persson et al., 2014; Prakash et al., 2012; Sambataro et al., 2010; Yang et al., 2013), although one study found no significant correlation between functional connectivity within the DMN and cognitive performance (Onoda et al., 2012). A longitudinal study in a Danish birth cohort reported that reduced task-induced deactivation in the posterior region of the DMN is only seen in a subclinical cognitive decline group and not in the control group (Hansen et al., 2014). Overall, the evidence lends support that changes in DMN activation and functional connectivity might be explained by a combination of cognitive abilities and age effects (Supplementary Table S4).
Analysis type
There is a paucity of work comparing and contrasting analysis type in DMN studies. One study demonstrated that in healthy adults DMN activation was seen in the same brain regions across three types of analysis—ReHo, region-of-interest (ROI), and ICA (Long et al., 2008). Another study found that the effects of normal aging, such as the loss of PCC coactivity could only be detected by ICA, but not by volume of interest signal time course correlation analyses of DMN interconnectivity (Koch et al., 2010) (Supplementary Table S5).
In preterm to adolescence, three of the eight studies analyzed their data using seed-based analyses, whereas the rest employed ICA. Analysis techniques comparing adolescence to adults yield conflicting results. The two ICA studies had conflicting findings, where Jolles et al., (2011) found greater connectivity in children versus adults, whereas the other (Supekar et al., 2010) found that children can attain adult-like levels of connectivity. Findings were also different comparing seed-based analysis (Fair et al., 2009) to ReHo by Lopez-Larson et al., (2011). Seed-based analysis found that the DMN is sparsely functionally connected in children, but develops over time, whereas ReHo found general decreases with age and the greatest reduction being seen in the anterior cingulate and temporal lobe. The two studies completed in adults both employed a seed-based ROI analysis and one also used ICA. He and colleagues (2013) found that functional connectivity was decreased even in middle-aged adults. Using ROI analysis, Bluhm and Associates (2008) found that similar findings with PCC/precuneus functional connectivity inversely correlated with age. However, ICA indicates a positive correlation with thalamus and left superior frontal gyrus connectivity and age. Comparing adults and elderly individuals, all studies regardless of analysis type support an overall decrease in activity and/or correlational strength within the DMN. Four studies in particular support decreased functional connectivity between the PCC and other regions of the DMN such as the ventral medial prefrontal cortex (vmPFC) (Grady et al., 2010; Koch et al., 2010; Mevel et al., 2013; Sambataro et al., 2010; Tomasi and Volkow, 2012).
Of the studies investigating sex differences, studies using ICA found no significant differences by sex in one study (Weissman-Fogel et al., 2010), whereas another study found that women show greater activation in the bilateral superior frontal gyrus and right angular gyrus. Two studies used ROI analysis and they both found that women have higher functional connectivity in regions such as the PCC/precuneus and prefrontal cortex (Bluhm et al., 2008; Tomasi and Volkow, 2012). The one study using ReHo found greater local connectivity in the right medial temporal regions in females only (Lopez-Larson et al., 2011).
Studies employing ICA, seed-based analysis, or multiscale entropy found support that functional connectivity is correlated with performance on cognitive tasks. However, the one study that used both ICA and seed-based analysis found no significant correlation between inter-DMN functional connectivity and cognitive performance (Onoda et al., 2012).
Discussion
Summary of results
Resting-state networks and in particular the DMN are important, since they reveal intrinsic and spontaneous networks that provide insight into the functional architecture of the brain (Whitfield-Gabrieli and Nieto-Castanon, 2012). Understanding their variance across natural factors, such as age, sex, cognitive function, and analysis type is crucial to characterize what is normal (Buckner and Vincent, 2007). The DMN changes in appearance and the strength of functional connectivity across age. It is less mature and abnormally formed in preterm infants. It continues to develop into adolescence. However, when comparing the strength of functional connectivity between children and adults, some studies indicate sparser and others stronger connectivity in children. This discrepancy in the formation and strength of the DMN in infants and children may be better explained by how data are collected. To account for children moving in the scanner, researchers often scan this demographic when they are asleep. There are natural sleep-induced alternations of the DMN, where coupling between the mPFC and anterior cingulate cortex with the PCC become nonsignificant (Horovitz et al., 2009; Sämann et al., 2011). Unfortunately, the studies included in this review did not comment if their participants fell asleep and if this impacted their analyses. Furthermore, many of the articles did not appropriately account or correct for motion artifacts. Thus, findings of decreased functional connectivity in children may be due to increased movement. However, studies that did have conservative motion correction parameters did find lower functional connectivity between the mPFC and PCC of the DMN in children compared with adults (Supekar et al., 2010). Therefore, the variance of functional connectivity in children might be better explained by sleep or motion rather than immature network formation. Similar to as discussed above, the pattern in preterm infants requires more evidence to support whether the immature network is due to the lack of development or due to imaging during different stages of sleep.
In middle-aged adults, the strength of functional connectivity in the DMN begins to decrease and continues into older age. Thus, it appears that the strength of DMN functional connectivity follows an inverse U-shape where it is greatest in adulthood. The findings from the meta-analysis support that adults have greater activation in many regions of the DMN in comparison to children and older adults. More studies are needed that combine analysis techniques, strict protocol measures such as accounting for sleep, and motion correction to better characterize and support the current findings.
Cognitive function has an effect on functional connectivity, where it is correlated with performance on cognitive tasks. Further studies should assess whether the age-related decline in functional connectivity is mediated by cognitive function. This finding would be of interest since it would help explain if variance is explained by increasing age or rather by cognitive abilities.
There are sex differences in the DMN, where females tend to exhibit stronger intranetwork connectivity compared with males. However, one study found no significant differences. Despite obvious influences of sex on DMN functional connectivity, very few studies investigate sex differences. Furthermore, included studies did not take into account the effects of premenstrual symptoms on BOLD activations. In women using hormonal contraceptives, somatic and psychological premenstrual symptoms are positively correlated with connectivity in the PCC and precuneus (De Bondt et al., 2015).
Only two studies explicitly compared DMN functional connectivity and analysis type. Future studies should incorporate complementing techniques such as seed-based analyses and ICA to provide more robust findings and to better determine if variance is truly present.
Limitations
This study has three major limitations. First is the relatively small number of included studies, and furthermore, the meta-analysis only included seven of the twenty-nine studies. Despite the small number of included studies in the meta-analysis and the limited power, the findings provide an impetus for future research. Second is the broad definition of “healthy.” Some studies employed very strict criteria defining healthy as the absence of neurological and psychiatric disorders, whereas others did not specify what they defined healthy as. Lastly, different analysis types were used among the included studies and, therefore, the findings from combining their results have limited power.
DMN and pathology
Identifying the progression of DMN activation and connectivity change in pathology is important since it provides useful information in identifying an intermediate risk phenotype and could serve as a biomarker for treatment response.
Future directions
Emerging research
New areas of research are emerging and findings add to the better characterization of this default mode of brain function. A recent study comparing homosexual and heterosexual men found differing DMN activities based on sexual orientation. In particular, homosexual men had decreased activity in regions such as the left precuneus and the post central gyrus, an area of the parietal lobe (Hu et al., 2014). Their work provides further information to characterize what “normal” DMN activity is by factors like sexual orientation. In a study investigating healthy subjects there was an increase in activity within the DMN associated with decreased emotion perception and increased hopelessness (Wiebking et al., 2011). The findings provide interesting information on how we define “healthy” and the need to have a broad definition that accounts for normal variation. Furthermore, recent evidence supports that the DMN consists of several subnetworks with varying specialized roles. This includes self-referential or social cognitive processing (Menon, 2011; Spreng et al., 2009) or differentiating between oneself from others (van Veluw and Chance, 2014). It would be of great interest to investigate if these subnetworks in the DMN differ by age, sex, cognitive function, and analysis type.
Recent work investigating the effects of external scanning parameters on DMN functional connectivity has highlighted the importance of recognizing their contributions. Scanner background noise alters attentional demands and consequently has been shown to suppress components of the DMN, including the mPFC, posterior cingulate, and precuneus (Gaab et al., 2008). Another study corroborated and supports findings that neuroimaging studies investigating resting-state functional connectivity need to adjust and control for individual differences in in-scanner motion (Mowinckel et al., 2012).
Emerging work investigating genetic contribution to DMN variability found that between 9% and 41% of the variance is attributed to genetic contribution (Glahn et al., 2010). Where the greatest variance explained by heritability was found for functional connectivity between the posterior cingulate and right inferior parietal nodes (Korgaonkar et al., 2014). The findings provide evidence that DMN connectivity variability could serve as an endophenotype for genetically predisposed psychiatry and neurological disorders (Korgaonkar et al., 2014).
Environmental factors
In healthy individuals, gray matter volume is significantly decreased in the hippocampus and significantly increased in the dorsomedial prefrontal cortex and the orbitofrontal cortex in participants who had experienced childhood maltreatment compared with those who had not (Chaney et al., 2014). In a longitudinal study, early life stress within the first year of life predicts higher cortisol levels at 4 years of age and at 18 years of age, the childhood cortisol levels were significantly correlated with amygdala vmPFC resting-state functional connectivity (Burghy et al., 2012). Childhood maltreatment influences functional connectivity and brain morphology; future work should assess its impact on DMN variation.
Conclusion
In conclusion, our study is the first to review differences in the DMN in healthy individuals. Further findings were stratified by age, sex, cognitive function, and analysis type.
Footnotes
Acknowledgments
The authors would like to thank Karen Gagnon for her assistance with the search strategy. This study was supported in part by the Ontario Brain Institute and Canadian Biomarker Integration Network for Depression.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.