
Abstract
Head motion is a known challenge in resting-state functional magnetic resonance imaging studies for biasing functional connectivity (FC) among distinct anatomical regions. These persist even with small motion, limiting comparisons of groups with different head-motion characteristics. This motivates an interest in the optimization of acquisition and correction strategies to minimize motion sensitivity. In this test–retest (TRT) study of healthy young volunteers (N = 23), we investigate the effects of slice-order acquisitions (sequential or interleaved) and head-motion correction methods (volume- or slice-based) on the TRT reproducibility of intrinsic connectivity of the default mode network (DMN). We evaluated the TRT reproducibility of the entire DMN and each main node using the absolute percentage error, intraclass correlation coefficient (ICC), and the Jaccard coefficient. Regardless of slice-order acquisition, the slice-based motion correction method systematically estimated larger motion and returned significantly higher temporal signal-to-noise ratio. Although consistently extracted across all acquisition and motion correction approaches, DMN connectivity was sensitive to these choices. However, the TRT reproducibility of the whole DMN was stable and showed no sensitivity to the methods tested (absolute reproducibility ∼7%, ICC = 0.47, and Jaccard = 40%). Percentage errors and ICCs were consistent across single nodes, but the Jaccard coefficients were not. The posterior cingulate was the most reproducible node (Jaccard = 52%), whereas the anterior cingulate was the least reproducible (Jaccard = 30%). Our study suggests that the slice-order and motion correction methods evaluated offer comparable sensitivity to detect DMN connectivity changes in a longitudinal study of individuals with low head-motion characteristics, but that controlling for the consistency in acquisition and correction protocols is important in cross-sectional studies.
Introduction
T
However, accurate and reproducible characterization of the DMN is challenging because measures of functional connectivity are susceptible to several confounding factors deriving from non-neural fluctuations in the BOLD signal, in particular in-scanner head motion (Murphy et al., 2013). The common presence of micromovements (<0.5 mm) exerts global effects throughout the entire brain, spuriously changing the BOLD content across voxels, causing signal distortions or loss (Van Dijk et al., 2012). It has been shown that head-motion artifacts can confound the intrinsic DMN connectivity, enhancing short range while diminishing long-range connections leading to reduced connectivity in the anterior cingulate cortex (ACC) (Murphy et al., 2013; Zeng et al., 2014). Furthermore, the intersubject motion variability hinders comparisons of intrinsic DMN connectivity across groups in cross-sectional designs (Power et al., 2012; Satterthwaite et al., 2012; Van Dijk et al., 2012; Zeng et al., 2014), whereas individual characteristic motion traits would limit the ultimate sensitivity to neural signals in longitudinal study designs (Guo et al., 2012; Zeng et al., 2014), precluding the development of DMN-based pre-clinical biomarkers of neurological conditions.
There are two general types of approaches aimed at addressing head-motion correction in MRI studies: one is based on prospective “real-time” measures of head motion with reorientation of the imaging slices on-the-fly (Godenschweger et al., 2016) and the other is based on retrospective estimation and reorientation of the data after acquisition (Jenkinson et al., 2002). The study of prospective measures is a promising active field of research, but such methods are not yet standardly available and are not further discussed.
In this study, we focus on retrospective head-motion correction methods, which are more commonly implemented in resting-state fMRI studies (Power et al., 2014). Among these methods, a typical approach consists of a volumetric registration of all brain time series to a reference volume along the entire train of acquisitions (Jenkinson et al., 2002). This procedure yields head translational and rotational parameters that are used to quantify motion, eventually excluding subjects, or only some volumes, compromised by excessive motion (Power et al., 2012). Other methods include the removal of the estimated motion parameters through multiple linear regressions (Satterthwaite et al., 2013) or exploit data-driven methods such as independent component analysis (ICA) to detect and remove motion signals (Griffanti et al., 2014; Pruim et al., 2014; Schöpf et al., 2010, 2011). Although widely implemented (Westbrook et al., 2005), one main limitation of the volumetric registration is that it treats the head as a rigid body and minimizes motion only between the acquired volumes (Murphy et al., 2013). However, in standard, fast two-dimensional (2D) MRI, each excitation pulse excites the magnetization of one slice at a time, while the head moves across all slices. This approach is therefore insensitive to the consequences of head motion on activation timing and patterns of excitation that occur across slices during volume acquisition (Cheng and Puce, 2014). More recently, a slice-based head-motion correction method was introduced to account also for these effects (Beall and Lowe, 2014) together with the introduction of novel acquisition sequences (Bright and Murphy, 2013; Kundu et al., 2012).
Besides retrospective head-motion correction methods, there are some fMRI acquisition factors that are relevant for head-motion during data acquisition, such as the time to acquire the full brain volume (repetition time [TR]) and the slice acquisition strategy (interleaved or sequential). A common approach is to acquire slices using an interleaved pattern, which allows good brain coverage having small or no gap between slices and yet avoid interference between adjacent slices that are not perfectly rectangular (cross talking). Interleaved slice acquisition, however, is more sensitive to head motion happening during each single volume acquisition, giving banding artifacts (Kim et al., 2008; Sladky et al., 2011). In contrast, sequential slice-order sequences present opposite characteristics and could minimize these motion-related artifacts during BOLD-weighted echo-planar imaging (EPI) volume acquisition, provided that a sufficient gap between adjacent slices is used (Kim et al., 2008; Sladky et al., 2011).
To the best of our knowledge, no studies have investigated whether there is an optimal combination of slice-order acquisition (sequential vs. interleaved) and retrospective head-motion correction (volume- vs. slice-based motion correction) methods that minimizes the sensitivity to in-scanner head motion, thus giving an optimal test–retest (TRT) reproducibility of intrinsic DMN connectivity. This is of interest as part of the optimization of acquisition and analysis protocols for longitudinal studies. In this study of healthy young adults, we evaluate how the combination between ascending slice acquisition methods, interleaved or sequential, and head-motion correction methods, volume based or slice based, influences the TRT reproducibility of intrinsic DMN connectivity metrics.
Materials and Methods
Subjects
Twenty-four healthy young volunteers (12 female, 27.0 ± 5.3 years) participated in this study approved by the Ethics Committee of the University of Trento. Subjects were scanned across two sessions, in average 4.2 ± 4.4 days apart. During each resting-state fMRI session, subjects were asked to relax, keep their eyes closed, stay still while avoiding to fall asleep, or engage in structured thoughts. Data from one participant were excluded from the analysis for being collected with a smaller number of TRs.
MR image acquisition
A 4T Bruker Medspec scanner (Bruker Medical, Ettlingen, Germany) with a birdcage-transmit and an 8-channel receive head coil (USA Instruments, Inc., Ohio) was used. In each session, one 3D (three-dimensional) T1-weighted magnetization-prepared rapid gradient echo (MP-RAGE) (TR/echo time [TE] 2700/4.18 ms; 1 mm3 voxel) and two resting state fMRI runs (TR/TE 2.2 s/30 ms, with 37 anterior–posterior commissure (AC-PC) parallel slices, voxel size 3 × 3 × 3 mm3, slice-gap 0.6 mm, 200 volumes) were acquired. The resting-state runs were acquired once with ascending interleaved and once with ascending sequential 2D slice acquisition orders. Slice-order acquisitions were not counterbalanced in the present study, with sequential acquisitions always being the last data to be acquired.
There is a risk that the last data to be acquired in each session (sequential slice acquisition) would show higher fatigue effects by introducing higher motion and potentially lower temporal signal-to-noise ratio (tSNR). This could introduce a bias for the sequential acquisition that could confound differences relative to the interleaved acquisition. Such biases were evaluated by within-session tSNR comparisons (see Preprocessing for more details).
Preprocessing
All DICOMs (Digital Imaging COmmunications in Medicine) were converted into Neuroimaging Informatics Technology Initiative (NIfTI) images using dicom2nii converter program in MRIcron. During conversion, two resting-state functional images that were acquired using the interleaved slice acquisition were inadequately flipped along the x-axis and were thus reoriented using the 3drotate AFNI tool.
The preprocessing of the functional EPI data was performed in the individual space of each subject using a combination of FSL (Jenkinson et al., 2012) and AFNI (Cox, 1996) programs (Fig. 1). The first six volumes were discarded (fslroi, FSL) to allow for steady-state stabilization of the BOLD fMRI signal. The tSNR from the full brain was in this study estimated (194 volumes) before any preprocessing corrections so that it would be in principle sensitive to head motion. The rationale was to evaluate whether potentially higher motion, due to higher fatigue during sequential acquisitions, would cause systematic reduction of tSNR.
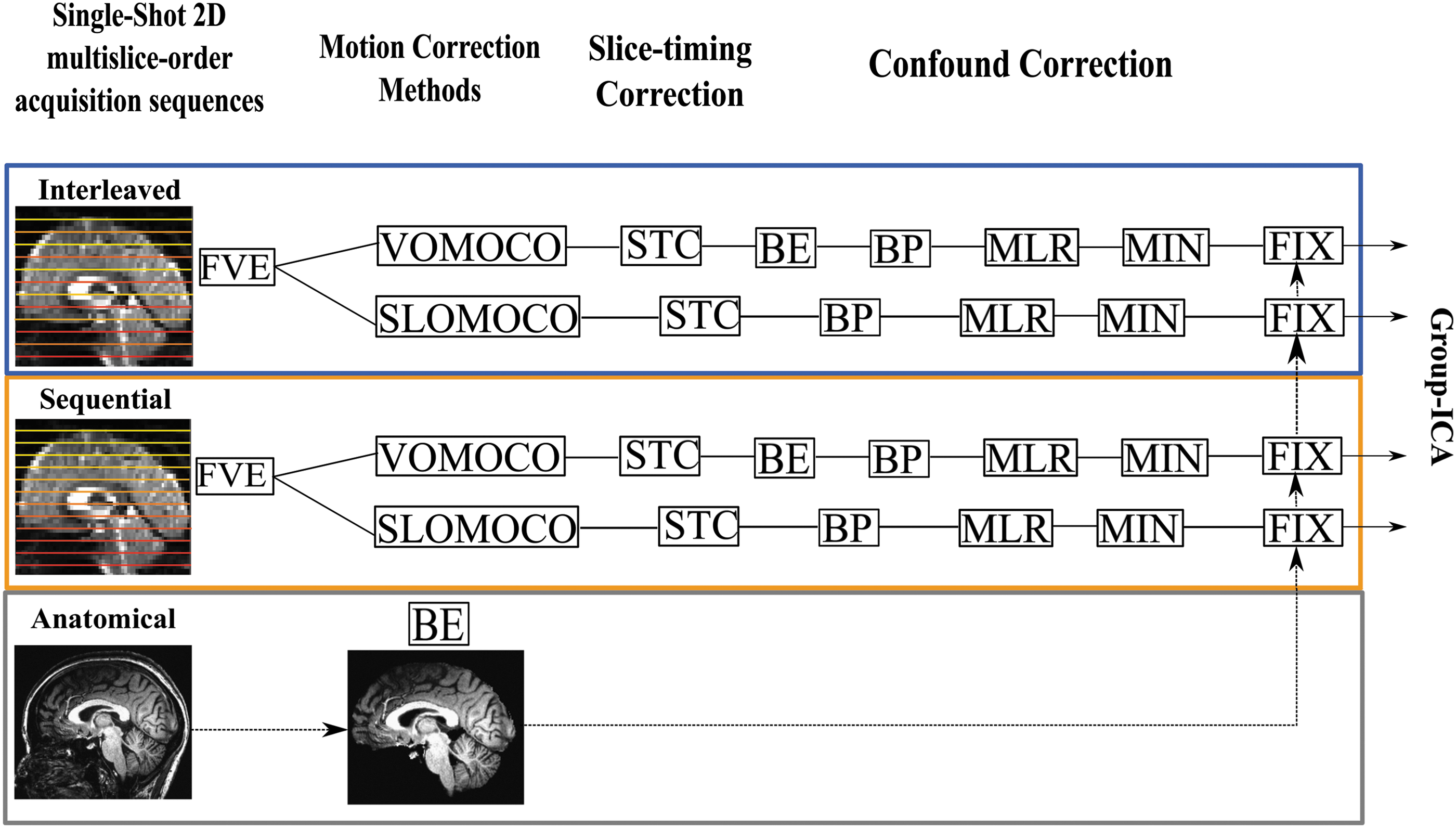
Preprocessing workflow. The figure illustrates the preprocessing pipeline under each slice-order acquisition sequence, either interleaved (blue rectangle) or sequential (orange rectangle) and motion correction method, either volume based (VOMOCO) or slice based (SLOMOCO). FVE, denotes the removal of the first six volumes from the BOLD time series; STC, slice-timing correction (interleaved/sequential); BE, brain extraction (included in SLOMOCO); BP, band-pass filtering (0.01–0.1 Hz); MLR, multiple linear regression (12 head movement time series (six head movement time series+six derivatives) filtered as BP; MIN, mean intensity normalization; FIX, FSL-FIX implementation. The bottom row indicates the anatomical brain extraction step needed for use in FSL-FIX (see Preprocessing for more details). SLOMOCO, slice-based head-motion correction; VOMOCO, volume-based head-motion correction. Color images available online at
Afterward, either volume-based or slice-based head-motion correction methods were implemented before slice-timing correction based on the slice-order acquisition method (slicetimer, FSL). The order of these corrections is controversial (Johnstone et al., 2006), but was in this study consistently adopted because it was predefined in SLOMOCO (see Slice-Based Head-Motion Correction section) (Beall and Lowe, 2014; Jones et al., 2008). From both head-motion correction methods, the six movement parameters were calculated.
Then, nonbrain voxels were removed from the EPI volume (bet2, FSL) and, to remove high-frequency signal fluctuations, a temporal filtering with a bandpass filter (0.01–0.1 Hz) was used on both the EPI volume (fslmaths, FSL) and the six head movement parameters (1dBandpass, AFNI) (Hallquist et al., 2013). Multiple linear regressions (3dDeconvolve, AFNI) were used to remove the six movement parameters and their relative derivatives (1d_tool.py, AFNI) plus second-order polynomials from the main signal. Individual brain volumes were normalized to mean signal intensity by a single factor of 1000 (fslmaths, FSL) and registered to brain-extracted anatomical volumes using Boundary-Based Registration (Greve and Fischl, 2009).
FMRIB's ICA-based Xnoiseifier (FSL-FIX) (Griffanti et al., 2014; Salimi-Khorshidi et al., 2014) was implemented to classify and remove the independent spatial components (25, MELODIC) associated with physiological noise and MRI hardware-related artifacts using a standard classifier (Standard.RData: TR = 3 s, Resolution = 3.5 × 3.5 × 3.5 mm3, session = 6 min, default FEAT preprocessing) and a medium classification threshold (50/100) as a criterion to distinguish between signal and noise associated components. To remove physiological noise confounds, FSL-FIX uses the nongray matter tissue masks (cerebral spinal fluid [CSF] and white matter [WM]) and the six motion parameters obtained after head-motion correction. The tissue masks were derived from the brain extracted anatomical volume, which was obtained using OptiBET for each single subject and session (Lutkenhoff et al., 2014).
For each condition, all subjects and sessions were spatially normalized to the MNI template through application of linear (affine) FLIRT (Jenkinson and Smith, 2001) and subsampled at a resolution of 3 mm isotropic voxels. To evaluate data preprocessing quality, the mean tSNR was estimated (194 volumes) on the three main tissue classes (gray matter [GM], WM, CSF) for each slice-order acquisition (interleaved and sequential) and each head-motion correction strategy (volume-based head-motion correction [VOMOCO] and slice-based head-motion correction [SLOMOCO]).
No spatial smoothing was applied in accordance to the recommendations by FSL-FIX developers (Salimi-Khorshidi et al., 2014). This choice has been seen not to affect the detectability of the DMN (Molloy et al., 2014). Given that ICA tends to separate motion components, we decided not to censor volumes in this study (Power et al., 2012). This also lets us use FSL-FIX and perform group analysis on time series of the same length (N = 194).
Head-motion correction methods
Below we outline the two head-motion correction methods applied and compared in this study: volume-based head-motion correction, in this study referred to as VOMOCO (Jenkinson et al., 2002), and slice-based head-motion correction, in this study referred to as SLOMOCO (Beall and Lowe, 2014).
Volume-based head-motion correction
VOMOCO was performed using a rigid-body motion correction method (
Slice-based head-motion correction
SLOMOCO was performed using SLice-Oriented MOtion COrrection (
Head-motion derived metrics
To evaluate the amount of motion estimated by the two head-motion correction methods, the transformation matrices were analyzed for each subject and session. Head rotation parameters were converted from radians to millimeters in terms of the corresponding displacement on a r = 50 mm sphere, representing the average distance between cortex and head center (Siegel et al., 2014). The straight sum between the six parameters (in absolute value) was computed to generate a 1D time series in this study referred to as the volume-to-volume displacement, for each subject and session. To further summarize head motion in each subject, the median volume-to-volume displacement was calculated.
Characterization of group DMN for each condition
After data preprocessing, we performed group independent component analysis (group-ICA) as implemented in the MELODIC ICA toolbox (Version 3.13) from FSL (Version 5.0.4) to extract the group DMN map relative to each combination of slice-order acquisition and head-motion correction method, respectively. All functional volumes across subjects (N = 23) and sessions (N = 2) were temporally concatenated, and the obtained signal was decomposed into 15 independent components (Infomax algorithm). Time courses were not variance-normalized, and default parameters were used for the mixture model fit. Dimensionality (15 ICs) was tailored to the empiric data to define the group DMN component across all conditions of interest (N = 4). Qualitative evaluations showed that higher dimensionality (16–20 ICs) gave splitting of the DMN across multiple components, while lower dimensionality (10–14 ICs) was insufficient to gain optimal IC estimation (Abou Elseoud et al., 2011; Jovicich et al., 2016).
For each condition, the DMN was visually selected among the 15 components as the one that included the most common DMN nodes: medial prefrontal and anterior cingulate cortex, posterior cingulate cortex and precuneus, and left/right lateroparietal cortex, using a threshold of z-score > 2.3 (Franco et al., 2010) (Fig. 2A). A reference template (Rosazza et al., 2012) was also used to qualitatively verify the degree of overlap of the group component map.

Group ICA and Dual Regression Analysis. The figure illustrates
Characterization of single-subject DMN
For each slice-order acquisition and head-motion correction method, dual-regression was then used to derive the single subject and session DMN (Fig. 2B) from the selected group component (Beckmann et al., 2009). Individual DMN volume maps were thresholded at z > 2.3, p < 0.01 (Beckmann and Smith, 2004).
To account for spatial DMN variability across sessions and subjects, a cluster analysis was run to detect the four main functional coactivated clusters that characterize the DMN (3dclust, AFNI). These clusters are the posterior cingulate and precuneus (including BA31, BA30, BA29, and BA23), the left/right ventral and dorsal posterior parietal cortex (including BA39, BA40, BA22, and BA7), and prefrontal and anterior cingulate areas (including BA9, BA10, BA32, and BA24) (Franco et al., 2010). To avoid inaccuracies for the definition of DMN activation maps, clusters were defined as made of voxels no >4 mm apart and a cluster volume of at least 67 mm3 in volume size, per each region-of-interest (ROI) (Marchitelli et al., 2016).
These clusters were anatomically constrained by a reference DMN template (Rosazza et al., 2012). Visual inspection was done to verify that clusters overlapped with the main regions attributed to the DMN.
Mean z-scores in the DMN (i.e., the mean z-score across all voxels in the four main clusters) and relative activation cluster size (i.e., the total number of voxels in the four main nodes) were characterized for each subject, session, and condition. In addition, mean z-scores and relative cluster size were also calculated for the posterior cingulate, medial prefrontal cortex, and parietal cortex, separately.
In addition to the combined group-ICA and dual-regression analysis, we also evaluated intrinsic DMN connectivity using a ROI-based approach, considering its wide employment in functional connectivity studies (Biswal et al., 1995; Fox et al., 2006; Raichle et al., 2001; Vincent et al., 2008). The ROI-based approach used spherical ROIs (10 mm radius) derived from the same reference template used for the group-ICA and dual-regression analysis (Rosazza et al., 2012). These ROIs were located around the posterior cingulate cortex (PCC) (Right-Anterior-Inferior [RAI] order: 0, 50, 28), ACC (RAI: 0, −45, 17), and right/left lateroparietal cortex (LPC) (RAI: ±47, 60, 30) on the MNI template. Of note, right/left LPCs were considered as a single ROI to be as more consistent as possible with the main ICA analysis used in this study.
For each subject and session, completely preprocessed and coregistered to the MNI space, Pearson's correlation coefficients were calculated for each possible combination of ROI pairs and bias-corrected using a bootstrap procedure (1dCorrelate, AFNI). Correlation coefficients were then averaged within each subject and session to yield a single DMN value and finally normalized to z-scores using the Fisher's r-to-z equation.
TRT reproducibility of the DMN and its main nodes
The main goal of this study was to evaluate the effects of different slice-order acquisition and head-motion correction methods on the longitudinal reproducibility of intrinsic DMN connectivity. Three measures of TRT reproducibility of functional connectivity patterns were evaluated on suprathresholded z-scores DMN maps (z > 2.3) for each ROI, slice acquisition, and motion correction method, respectively:
(1) The absolute percentage error to measure the intersession error in ROI-wise mean z-scores for each subject (Bennett and Miller, 2010):
with
(2) ROI-wise intraclass correlation coefficient, ICC (2,1) (Shrout and Fleiss, 1979) of mean z-scores, and relative confidence intervals (C.I.) (McGraw and Wong, 1996) to measure the proportion of intersubject variance out of the total variance in the entire sample:
with σ2 (between) and σ2 (within) indicating the intersubject and intrasubject variance in mean z-scores, respectively and ɛ is the residual error;
(3) To measure voxel-wise spatial reproducibility of the DMN, we used for each subject the Jaccard index, in this study defined as the fraction between shared (intersection, ∩) active voxels out of all (union, ∪) active voxels across sessions:
The Jaccard index was computed for the DMN as a whole and for each single DMN node separately (PCC, ACC, LPC, and RPC), for each acquisition and head-motion correction method.
Statistical evaluations
Statistical evaluations were performed in IBM SPSS Statistics for Macintosh, Version 22.0. We used bivariate Pearson's correlations to evaluate the existence of relationships between head movement measures (i.e., median volume-to-volume displacements) and functional connectivity and its TRT reproducibility in all the ROIs (N = 4) and for each combination of slice-order acquisition and head-motion correction method (N = 4). Of note, when correlating TRT reproducibility scores and motion-derived metrics, median displacements were averaged across the two sessions.
We used a paired t-test to evaluate slice-order acquisition effects potentially induced by motion in the full-brain tSNR from the nonpreprocessed acquired data (N = 46). After data preprocessing, the nonparametric Friedman test was conducted across all subjects and sessions (N = 46) to assess for significant head-motion correction and/or slice-order acquisition method effects in the mean tSNR values in GM and the other brain tissues (WM, CSF).
We performed the nonparametric Friedman test to evaluate both slice-order acquisition and head-motion correction method effects in mean gray-matter tSNR values after data preprocessing and MNI normalization (N = 46). The same nonparametric test was conducted to evaluate the effects of interest on the median values of motion-derived metrics across all subjects and sessions (N = 46). The Friedman test was finally used also to assess slice-order acquisition and head-motion correction method effects in mean ICA z-scores (N = 46) and TRT reproducibility values (N = 23), for the entire DMN and its single nodes, respectively. Correspondingly, the test was also used for mean DMN (Fisher's z-normalized) correlation coefficients (N = 46) and associated TRT reproducibility scores (N = 23). A statistical comparison analysis between the group-ICA/dual regression and the ROI-based methods was also conducted for each slice-order acquisition and head-motion correction method using a paired-sample t-test.
Statistical significance level was set to p < 0.05. Statistics were corrected for multiple comparisons over all possible pairwise combinations using the method of Dunn-Bonferroni (Dunn, 1964) at α = 0.05 and C.I. = 95%.
Results
Head-motion estimates and tSNR evaluations
The overall estimated head-motion parameters from VOMOCO and SLOMOCO are illustrated in Figure 3A. The Friedman test performed across all slice-order acquisition and head-motion correction methods revealed significant differences in the amount of movement estimated (Table 1). Pairwise comparisons revealed that median displacements estimated using VOMOCO (0.19 ± 0.14 mm) were significantly lower than SLOMOCO (1.16 ± 1.44 mm) for both interleaved (Z = −2, p < 0.001) and sequential (Z = −2, p < 0.001) acquisitions.
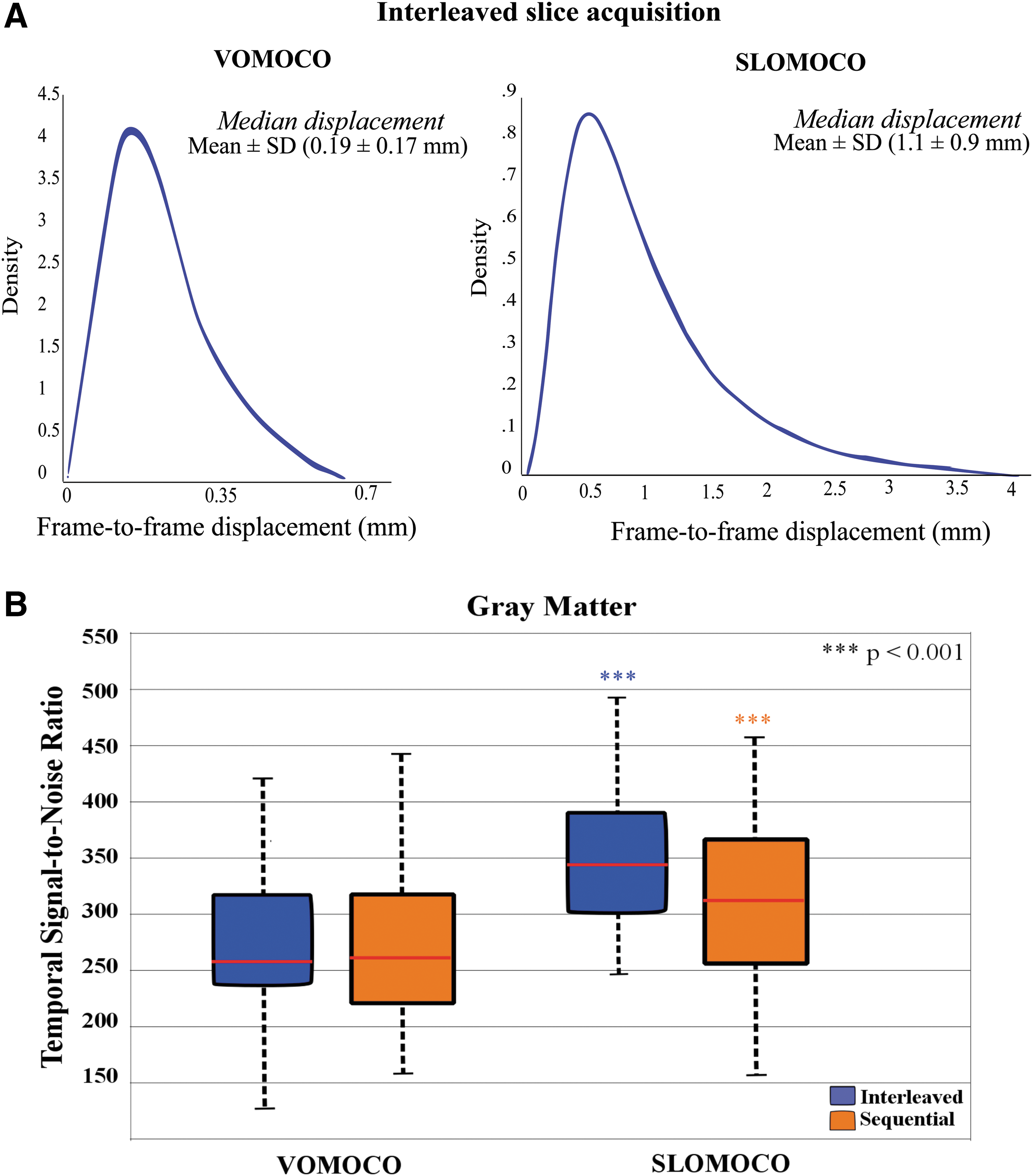
Overall head movement and tSNR estimates.
The table shows averaged median and maxima volume-to-volume head movement displacements across individuals for each slice-order acquisition protocol and head movement correction method. Second to last and bottom lines report the tSNR prior any head-motion correction and after data preprocessing, respectively. Head movement displacement time series were derived from the straight sum of the absolute value of six head movement parameters of 194 time points in length. Head movement estimates and the tSNR are similar across slice-order protocols, but differ across head-motion correction methods.
SLOMOCO, slice-based head-motion correction; tSNR, temporal signal-to-noise ratio; VOMOCO, volume-based head-motion correction.
Before data preprocessing, a paired sample t-test indicated no significant difference in tSNR measures for interleaved (22.2 ± 4.3) and sequential (22.6 ± 4.4) slice acquisition methods; t(45) = −4.9, p = 0.63. Since the sequential slice acquisition was always the last acquisition in each session, the lack of systematic tSNR differences between the two slice acquisition methods is consistent with no increased fatigue (motion) in the last run.
After data preprocessing, significant differences in tSNR were found within GM across methods (Table 1). Pairwise comparisons across conditions revealed that slice acquisition order had no effects on tSNR. However, SLOMOCO consistently gave higher tSNR than VOMOCO: SLOMOCO (334 ± 73) and VOMOCO (276 ± 65) for both interleaved (Z = −1.8, p < 0.001) and sequential (Z = −1.3, p < 0.001) acquisitions (Fig. 3B). Similar findings were obtained for all brain tissue classes examined (GM, WM, and CSF), with tSNR percent increases in the range of 23–26% for SLOMOCO relative to VOMOCO (Supplementary Fig. S1; Supplementary Data are available online at
In summary, in this group of subjects, the amount of movement estimated from both head-motion correction methods was relatively low compared to the voxel size. The movement estimates were not significantly affected by slice acquisition strategy, but they were significantly affected by the head-motion correction method used, with SLOMOCO giving larger movement estimates and higher tSNR after data preprocessing.
Intrinsic DMN connectivity: slice acquisition methods and head-motion correction effects
Despite differences in head movements estimated across retrospective motion correction methods, group-ICA revealed DMN coactivation patterns across all combinations of slice-order acquisition and head-motion correction methods (Fig. 4A). No subject was discarded during data preprocessing due to excessive motion or data postprocessing since the dual-regression methodology combined with the functional clustering analysis was able to detect individual DMN maps across all subjects, sessions, and conditions.

Group and individual intrinsic DMN connectivity.
Intrinsic DMN connectivity results are reported in Figure 4B. Significant statistical differences in mean z-scores across slice-order acquisition and head-motion correction methods were found in the entire DMN (Table 2). Pairwise comparisons revealed that mean z-scores in the entire DMN were statistically different between head-motion correction methods for interleaved acquisition (Z = 1.3, p < 0.001) and between slice-order acquisition methods, using both VOMOCO (Z = 0.08, p = 0.02) and SLOMOCO (Z = −0.09, p = 0.005).
Average (SD) mean z-scores across participants and sessions under each slice acquisition and motion correction method (columns) for each ROI (rows). The last column provides nonparametric analysis of variance output (uncorrected). Significant statistical effects were found across conditions in each single ROI (p < 0.05). Except for PCC (slice order effects only), for all other areas the effects were significant for both slice order and motion correction method. See text for more information.
ACC, anterior cingulate and medial prefrontal cortex; DMN, default mode network; LPC, latero-parietal corteces; PCC, precuneus and posterior cingulate cortex; SD, standard deviation; SLOMOCO, slice-based head-motion correction; VOMOCO, volume-based head-motion correction.
Statistical significant effects were also found in each single node of the DMN (Table 2). In the PCC, pairwise comparisons revealed that mean z-scores were statistically different between slice-order acquisition methods only when implementing SLOMOCO (Z = −0.76, p = 0.03). In the ACC, multiple pairwise comparisons revealed that mean z-scores in interleaved acquired and VOMOCO corrected data (7.0 ± 1.0) were statistically different from sequentially acquired (Z = 1.72, p < 0.001) and SLOMOCO corrected data (Z = 1.68, p < 0.001). Finally, in the LPC, mean z-scores were statistically different between slice-order acquisition methods (SLOMOCO: Z = −0.9, p = 0.005) and head-motion correction methods for interleaved acquisitions (Z = 1.26, p < 0.001).
Considering the whole DMN regions and the PCC node, no statistically significant correlation between mean z-scores and median volume-to-volume displacements were found for any slice-order acquisition and head-motion correction method. A weak correlation between these two metrics was found in both LPC and ACC (LPC/ACC: ρ = 0.3, p < 0.05) only for sequentially acquired data combined with VOMOCO (LPC) and SLOMOCO (ACC).
When considering the supplemental ROI-based analysis, by contrast to mean ICA z-scores, the mean z-normalized correlation coefficients associated with the entire DMN were not influenced by slice-order acquisition and head-motion correction methods [χ 2 (3, n = 46) = 0.47, p = 0.93].
To summarize, the main characteristic DMN nodes were found and had similar spatial locations in all slice acquisition and head-motion correction methods. However, the group connectivity estimations of the whole DMN and its separate main nodes were significantly affected by slice-order acquisition and head-motion correction methods.
TRT reproducibility of the DMN: slice acquisition and head-motion correction effects
The three TRT reproducibility measures of intrinsic DMN connectivity for each slice-order acquisition and head-motion correction method are shown in Figure 5. The absolute percentage error of mean z-scores, averaged across participants, and the ICC (2,1) scores is shown in the top and middle panels, respectively.

TRT reproducibility of FC-FMRI in the DMN. The figure shows
Descriptive and inferential statistical results are reported in Table 3. With regards to the reproducibility errors of the absolute percentage error, the overall error averaged across participants, slice acquisition methods, and head-motion correction methods was below 8% and stable among DMN nodes (DMN: 7%; PCC: 8%; ACC: 6%; LPC: 8%), while ICCs (2,1) were overall fair, scoring 0.46, consistent among nodes (DMN: 0.47; PCC: 0.42; ACC: 0.5; LPC: 0.47). There were no significant effects in either reproducibility errors or ICCs due to slice acquisition and head-motion correction methods (Table 3).
The table shows the average (SD) TRT reproducibility errors across participants and relative ICCs (C.I.) for each condition of interest (columns) and ROI (rows). The last column reports statistical evaluations using nonparametric Friedman test. No significant statistical differences were found in TRT reproducibility errors across conditions in any ROI.
ACC, anterior cingulate and medial prefrontal cortex; C.I., confidence intervals; DMN, default mode network; ICC, intraclass correlation coefficient; LPC, latero-parietal corteces; PCC, precuneus and posterior cingulate cortex; TRT, test–retest.
The supplemental ROI-based analysis revealed high-absolute TRT reproducibility error of intrinsic DMN connectivity (28.5% ± 24%) across all slice-order acquisition and motion correction methods. Although, alike ICA, no significant effects due to slice acquisition and head-motion correction methods were found; the TRT reproducibility of intrinsic DMN connectivity was significantly lower in the ROI-based approach compared with the ICA-based methodology, in any condition of interest (Supplementary Table S1).
The Jaccard index (bottom Fig. 5) was moderate for the overall DMN (40%), higher for the PCC node (51%) compared with LPC (39%), and the ACC node (30%) that scored the lowest reproducibility. The Friedman test performed across all subjects and conditions-of-interest revealed no statistically significant effects of either slice acquisition or head-motion correction method in the entire DMN, but revealed statistically significant effects in all its single nodes (Table 4). Slice-order acquisition or motion correction method effects were not systematic across DMN regions. In particular, sequential acquisitions combined with SLOMOCO induced slightly opposite effects between anterior (ACC) and posterior (PCC) nodes yielding the lowest (21% ± 7%) and highest (64% ± 8%) spatial overlap reproducibility, respectively. Noteworthy, these effects were still observed at α level = 0.01 and C.I. = 99%. In the LPC, interleaved acquisition combined with VOMOCO statistically outperformed all other combinations of slice-order acquisition and motion correction methods. However, at α level = 0.01 and C.I. = 99%, significant differences were observed only between VOMOCO (52% ± 10%) and SLOMOCO (32% ± 9%) under interleaved acquired data (Z = 1.17, p < 0.001) while no slice-order acquisition effects survived.
Average (SD) Jaccard index across participants is reported for each slice-order acquisition and motion correction method (columns) and ROI (rows). Nonparametric pairwise Friedman's tests testing effects among columns are reported in the last column (uncorrected). Even though no significant statistical effects were found when considering the entire DMN, slice-order protocol and/or motion correction method effects were found in each single DMN node, suggesting higher intranetwork susceptibility to these factors.
ACC, anterior cingulate and medial prefrontal cortex; DMN, default mode network; LPC, latero-parietal corteces; PCC, precuneus and posterior cingulate cortex.
Importantly, no statistically significant correlation was found between any TRT reproducibility measure and median motion displacement metrics, averaged across sessions, in any ROI and for any manipulation in the resting-state fMRI protocol.
To summarize, neither slice-order acquisition nor head-motion correction methods affected the TRT reproducibility of mean z-scores in the DMN, as well as among its nodes. Nor they affected the spatial overlap reproducibility of the entire DMN. However, the spatial overlap reproducibility was subjected to interregional variability, with frontal DMN node (ACC) as the least spatially reproducible. The choice of interleaved slice-order acquisition yielded the most reduced interregional DMN variability regardless of head-motion correction methods. Most importantly, no combination of slice acquisition and head-motion correction method was found that would offer significant TRT reproducibility improvements that were systematically consistent across DMN nodes.
Discussion
Main findings
The main goal of this TRT reproducibility study was to evaluate whether different combinations of slice-order fMRI acquisition protocols and head-motion correction techniques would differently affect the longitudinal stability of intrinsic DMN connectivity. Participants were instructed to remain as still as possible to investigate these effects in standard resting-state experimental conditions rather than with artificially directed movement. Our main findings are the following: (1) in this group of healthy young volunteers with low head-motion characteristics, group ICA and dual-regression methodology determined robust and consistent DMN activations across all subjects (N = 23) and sessions (N = 2) in all conditions (N = 4); (2) slice-based motion correction gave significantly higher motion estimates and also significantly higher gray matter tSNR after preprocessing; (3) slice-order acquisition and head-motion correction methods significantly affected mean z-scores in all DMN regions, but they did not influence their TRT connectivity reproducibility within the same regions; (4) the spatial overlap reproducibility of DMN was also insensitive to acquisition and analysis protocol manipulations, PCC being the most and ACC the least spatially reproducible nodes, respectively; and (5) most importantly, no combination of slice acquisition and head-motion correction methods improved the TRT reproducibility of intrinsic DMN connectivity in a systematic way.
Retrospective motion correction methods
In agreement with previous studies (Beall and Lowe, 2014), our results show that, on the same data, SLOMOCO systematically gives higher movement estimates compared with VOMOCO, which on average has been seen to miss at least the 50% of nonvolumetric motion in real data. In average, across all participants and sessions (N = 46), SLOMOCO parameters registered 64% and 61% higher motion than VOMOCO under interleaved and sequential slice-order acquisitions, respectively. Although both methods indicated overall low motion in the data, SLOMOCO yielded significantly higher tSNR in gray matter compared with VOMOCO under both slice-order acquisitions. This finding encourages the adoption and further development of intravolume motion correction methods to enhance existing intervolume retrospective correction approaches in cross-sectional resting-state FC-MRI studies.
Intrinsic DMN connectivity
Using group-ICA and dual-regression methods, we were able to identify the group DMN and reconstruct all individual DMN components under all slice-order acquisition and head-motion correction methods across fMRI sessions. These findings highlight the reliable nature of the ICA approach (Beckmann et al., 2009) to define stable DMN maps in longitudinal resting-state FC-MRI studies (Biswal et al., 2010; Chen et al., 2008; Damoiseaux et al., 2006; Jovicich et al., 2016).
Both slice-order acquisition and head-motion correction methods affected intrinsic DMN connectivity in the whole network and its single nodes. However, this effect was not equally distributed across all DMN nodes, being less prominent in the PCC region. In fact, in contrast to all other DMN nodes, we found that mean z-scores in the PCC were very robust against potential surviving motion artifacts, with no significant correlations with the estimated motion from both VOMOCO and SLOMOCO. These observations confirm the resilience of the main hub of the DMN (Fransson and Marrelec, 2008) and support research in PCC-based connectivity markers of dementia (Bai et al., 2009; Zhou et al., 2008).
TRT reproducibility of the DMN
On the contrary, neither slice-order acquisition nor head-motion correction methods affected TRT reproducibility errors or the ICCs. Both of these metrics gave fair-to-good reproducibility in mean z-scores, in agreement with previous DMN reproducibility studies (Braun et al., 2012; Shehzad et al., 2009). Importantly, both reproducibility metrics were not subjected to interregional variability across the main network's nodes. These findings suggest that while intrinsic DMN connectivity metrics are markedly sensitive to head-motion artifacts (Van Dijk et al., 2012) and relative correction choices (Power et al., 2014), the connectivity reproducibility results are comparable and robust. Despite circumscribed to a low-motion cohort, this might indicate an individual trait-like stable property of motion (Van Dijk et al., 2012) that cannot be accounted for by the evaluated slice-order acquisition and motion correction methods.
Compared with group-ICA and dual regression, the intrinsic DMN connectivity calculated through the ROI-based approach yielded overall poorer reproducibility. This generalizes previous comparative findings (Jovicich et al., 2016) and indicates that multivariate data-driven approaches show higher capability to discriminate neural from non-neural BOLD signal sources (Beckmann et al., 2005). In particular, the selection of ROI size, shape, and location across and within individuals remains challenging for longitudinal resting-state fMRI studies.
The spatial overlap reproducibility of the entire DMN was moderate and, unlike mean z-scores, insensitive to slice-order acquisition and motion correction methods. Nevertheless, it showed a certain degree of variability among DMN nodes, with the highest reproducibility in the PCC (52% ± 13%) and the lowest in the ACC (30% ± 11%). This finding is in agreement with previous 3T studies that found higher variability in frontal DMN areas across subjects and sessions (Damoiseaux et al., 2006; Van Dijk et al., 2010).
We found that none of the DMN reproducibility measures correlated with motion-derived parameters or were influenced by slice-order acquisition or head-motion correction methods. This null effect could be a consequence of the low head-motion parameters in our participants. A different protocol could have been considered in which subjects were asked to follow particular patterns of head motion to then parametrically characterize their effects on the acquisition and preprocessing schemes (Reuter et al., 2015). Such a study, however, would have no longer been a resting-state study and we decided to acquire our data using classical instructions as done in the vast majority of resting-state acquisitions. Other studies have shown that higher levels of head motion can negatively influence the TRT reproducibility of the DMN in healthy elders, acquired using interleaved sequences and applying VOMOCO methods (Guo et al., 2012).
Limitations and future directions
This study presents several limitations. As previously mentioned, the low levels of head motion in our population might have hidden significant acquisition and/or correction method effects in the reproducibility of the DMN. Populations prone to higher degree of head motion might be of interest for further studies. The data-driven strategy in this study implemented using ICA could be an additional reason for having found that reproducibility was invariant to the factors manipulated. Not only ICA is known to give reproducible components (Abou Elseoud et al., 2011; Calhoun et al., 2009; Zuo et al., 2010) but also it was used twice in our workflow, using the ICA-based method FSL-FIX to remove physiological noise and surviving motion-related signals (Griffanti et al., 2014; Salimi-Khorshidi et al., 2014). This double ICA decomposition could have helped reduce the amount of motion-related variability investigated. Besides ICA, there are alternative methods for estimating DMN connectivity. In particular, a widely used method is seed-based connectivity with a seed positioned in the PCC (Andrews-Hanna, 2012; Buckner et al., 2008; Fransson and Marrelec, 2008). Although known for its robust results, this method is prone to lower reproducibility because it relies on the use of the PCC anatomical prior, typically taken from an anatomical atlas. ICA, on the other hand, being data driven, is potentially better suited at removing signal fluctuations that are unrelated to the DMN activity that could reduce TRT reproducibility. In fact, a recent study directly compared ICA and seed-based DMN showing that the ICA TRT reproducibility was superior (Jovicich et al., 2016).
Other limitations are related to data acquisition: this DMN reproducibility study is that the order of sequential and interleaved slice acquisitions was not counterbalanced across subjects and sessions. Instead, sequential acquisition was systematically the last run in every session for all subjects. This raises the possibility of biases, given that fatigue effects could make the subjects systematically move more during the last session, giving both higher motion metrics and potentially lower tSNR for the sequential slice acquisition protocol. Our evaluations show, however, that there were neither significant systematic motion metrics nor tSNR differences between the two acquisitions in each run. In addition, we used an ascending protocol for both slice acquisition methods. Using descending protocols would have potentially made the data more robust against blood in-flow saturation effects in that direction (Turner et al., 1998), with overall lower sensitivity to motion spin-history artifacts.
Finally, this reproducibility study only investigates one resting-state network: the DMN. While other resting-state networks are of interest too, our data-driven approach was tailored to characterize this network and might show weaknesses when examining the entire functional connectome. While other group-ICA algorithms could be further evaluated to characterize reproducibility and head motion sensitivity (Esposito et al., 2005; Goebel et al., 2006), more and more FC-MRI studies are using graph-theoretical measurements (Sporns, 2011; van den Heuvel and Sporns, 2013) to investigate the architecture of resting-state networks and may offer improved reproducibility (Braun et al., 2012); future studies should therefore also investigate how acquisition and preprocessing choices might influence the TRT reproducibility of graph-derived metrics.
To address this limitation, we make the raw TRT data of this study (24 subjects, each with two slice acquisitions and two sessions, making a total of 96 resting-state fMRI datasets) publicly available at The Consortium for Reliability and Reproducibility (Zuo et al., 2014). Sharing the data may facilitate the evaluation of additional retrospective head-motion correction methods, additional resting-state networks, and additional connectivity metrics. Other analysis software and group-ICA algorithms could be further evaluated to characterize reproducibility and head-motion sensitivity (Esposito et al., 2005; Goebel et al., 2006).
Conclusions
This work evaluates how interleaved or sequential ascending slice-order acquisitions combined with volume-based or slice-based retrospective head-motion correction methods influence the short-term TRT reproducibility of the intrinsic DMN reproducibility in a group of healthy young adults. Both motion correction methods detected overall low motion, with slice-based correction detecting systematically higher motion than volumetric-based correction and higher gray matter tSNR (21%). Group-ICA and dual-regression methods were able to reliably characterize the DMN in the entire sample across all these fMRI protocol manipulations. The intrinsic DMN connectivity was sensitive to protocol manipulations, but the TRT reproducibility of the DMN was not. Both intersession percentage errors (8%) and the ICC analysis (ICC = 0.47) indicate fair-to-good reproducibility of mean z-scores, while the Jaccard index indicates moderate spatial overlap reproducibility (40%) of the DMN. The TRT reproducibility of mean z-scores is constant between the main DMN nodes (PCC, LPC, and ACC) across all slice acquisition and motion correction methods. In contrast, the spatial overlap reproducibility is the highest in the PCC (52% ± 13%) and the lowest in the ACC (30% ± 11%). None of the four protocols evaluated gave a systematic improvement of reproducibility relative to the others. Our findings, limited to a population with low head-motion characteristics, suggest that for longitudinal studies the four protocols evaluated offer comparable sensitivity to detect longitudinal effects. However, for cross-sectional studies our findings suggest that controlling for consistency in slice acquisition and motion correction methods is important because these can bias the intrinsic connectivity estimates.
Footnotes
Acknowledgments
Special thanks to Giulia Elli for resting-state data acquisition and to the CiMEC MRI laboratory for medical and technical support. Research was supported, in part, by a European Research Council starting grant (MADVIS; ERC-StG 337573) attributed to O.C.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.