
Abstract
Pramipexole is widely prescribed to treat Parkinson's disease but has been reported to cause impulse control disorders such as pathological gambling. Recent neurocomputational models suggested that D2 agonists may distort functional connections between the striatum and the motor cortex, resulting in impaired reinforcement learning and pathological gambling. To examine how D2 agonists modulate the striatal-motor connectivity, we carried out a pharmacological resting-state functional magnetic resonance imaging study with a double-blind randomized within-subject crossover design. We analyzed the medication-induced changes of network connectivity and topology with two approaches, an independent component analysis (ICA) and a graph theoretical analysis (GTA). The ICA identified the sensorimotor network (SMN) as well as other classical resting-state networks. Within the SMN, the connectivity between the right caudate nucleus and other cortical regions was weaker under pramipexole than under placebo. The GTA measured the topological properties of the whole-brain network at global and regional levels. Both the whole-brain network under placebo and that under pramipexole were identified as small-world networks. The two whole-brain networks were similar in global efficiency, clustering coefficient, small-world index, and modularity. However, the degree of the right caudate nucleus decreased under pramipexole mainly due to the loss of the connectivity with the supplementary motor area, paracentral lobule, and precentral and postcentral gyrus of the SMN. The two network analyses consistently revealed that pramipexole weakened the functional connectivity between the caudate nucleus and the SMN regions.
Introduction
P
The basal ganglia receive cortical projections and project back to cortical regions via the thalamus, forming parallel (Alexander et al., 1986) and integrative circuits (Bar-Gad and Bergman, 2001) in support of motor, cognitive, and emotional processes. This circuitry has been emulated by recent neurocomputational models. In the basal ganglia Go/NoGo model (Frank et al., 2007), for example, the striatum is connected to the supplementary motor area (SMA) through a direct pathway, comprising so-called Go neurons that express mostly D1 receptors, and an indirect pathway, made up by NoGo neurons that express mostly D2 receptors. The Go neurons disinhibit the thalamus by suppressing the internal globus pallidus internal segment and substantia nigra pars reticulate (GPi/SNr), facilitating the execution of the appropriate action for the current state. The NoGo neurons inhibit the thalamus by removing the inhibition of the globus pallidus external segment (GPe) on the GPi/SNr, suppressing the competing actions in the motor cortex. By abnormally elevating the dopamine level of the indirect pathway, this model has replicated the impaired performance in response to negative reinforcers and has explained the neural mechanism of pathological gambling behavior in PD patients treated with D2 agonists.
However, it has not been empirically examined how D2 agonists distort functional connections between the basal ganglia and the motor cortex, or at a global level. To test this issue, we carried out a pharmacological resting-state functional magnetic resonance imaging (fMRI) study with a double-blind randomized within-subject crossover design. Each participant received pramipexole and placebo in two different scanning sessions. We used two independent approaches to target the issue, an independent component analysis (ICA) and a graph theoretical analysis (GTA). ICA has been used to decompose the whole-brain fMRI data into several resting-state networks, including the sensorimotor network (SMN) and default mode network (DMN) (Beckmann et al., 2005; Damoiseaux et al., 2006). Regions of the same network are functionally related and time series of those regions are highly correlated. We used ICA to examine whether pramipexole modulates the interregional connectivity within the SMN. The GTA is motivated by the idea that brain function may emerge from the topology of the brain as a whole rather than individual regions or connections. Applied to human anatomical and functional imaging data, it provides a concise quantification of the extraordinary complexity of interregional connectivity in human brain (Bullmore and Sporns, 2009). By measuring global and regional properties, we examined on which level pramipexole affects the topology of the whole-brain network.
Materials and Methods
All procedures had been cleared by the Ethics Review Board of the University of Magdeburg, the affiliation of the senior author at the time of the experiment. The experiment was carried out according to the Declaration of Helsinki.
Participants
Only male participants (n = 16, mean age 25 years, age range 21–28 years) were included in this study to avoid problems that could arise with unknown pregnancies and cycle-dependent changes in blood oxygen level dependent (BOLD)-response in women. None of the participants had a history of neurological or psychiatric disorder. All of them provided written informed consent before the experiment.
Design and drugs
This study used a double-blind randomized within-subject crossover design. Participants received pramipexole and placebo in two different sessions, which were at least 1 week apart. In each session, they either orally received 0.5 mg pramipexole or identically overcoated placebo according to a randomization table. They also orally received 20 mg domperidone in a nonblind manner in each session to antagonize potential nausea induced by pramipexole. Seven of the participants received pramipexole in the first session and the rest in the second session. Participants were instructed to neither engage in cognitive nor motor activity and to keep their eyes closed during scanning. Pramipexole reaches peak plasma concentration in ∼2 h and has a half life of about 8 h in healthy young adults. The scanning started 3 h after the administration of pramipexole and lasted 6 min.
Data acquisition
Data were collected in a 1.5-T GE Signa Horizon LX scanner. Functional images were acquired using a T2*-weighted echo planar imaging sequence, with 2000-ms time repetition, 35-ms time echo, and 80° flip angle. Each image consisted of 23 axial slices, with 64 × 64 matrix, 200 × 200-mm2 field of view, 4-mm thickness, 1-mm gap, and 3.125 × 3.125-mm2 in-plane resolution. Structural images were obtained using a T1-weighted 3D SPGR sequence. Each structural image consisted of 124 contiguous slices, with 256 × 256 matrix, 200 × 200-mm field of view (FOV), and 1.5-mm thickness.
Data preprocessing
Data were preprocessed with SPM8. The first two volumes were discarded owing to longitudinal magnetization equilibration effects. Functional images were time-shifted to adjust for staggered slice acquisition, realigned with a six-parameter rigid body transformation to correct movement artifacts, and normalized into MNI space (ERP-derived template ICBM 152, Montreal Neurological Institute). The data were smoothed with a Gaussian kernel of 6-mm full-width half-maximum for the ICA, but not for GTA. Because time series of all voxels of the same regions would be averaged in GTA (see Graph theoretical analysis), this analysis would benefit from clear boundaries between regions. White matter, cerebral spinal fluid, and motion signals were regressed out to reduce the influence of unspecific physiological effects for GTA, but not for ICA. This is because the artifacts could be captured and discarded in later steps of the ICA (see Independent component analysis).
Independent component analysis
The ICA was performed to examine how pramipexole modulates interregional connectivity within the SMN. To avoid the problem of matching components across participants, the ICA was computed directly on the group data by implementing temporal concatenation plus dual regression with the GIFT toolbox (Calhoun et al., 2001, 2009). Individual data were temporally concatenated and split into 14 independent components with the infomax algorithm (Bell and Sejnowski, 1995). Each component was presented as a spatial map along with an associated time course. The optimal number of components was estimated with a modified minimum description length algorithm (Li et al., 2007). Individual images were back-reconstructed to capture subject-specific variations.
To identify artifacts, the spatial map of each component was correlated with prior probabilistic maps of white matter and cerebral spinal fluid (MNI templates from SPM). If the spatial correlation for white matter was greater than R 2 = 0.02 or greater than R 2 = 0.05 for cerebral spinal fluid, the component was considered to represent artifacts and therefore should be discarded. Visual inspection suggested the discarded components represented eye movements, head motion, and other artifacts. The remaining components comprised all classically identified resting-state networks, including the SMN, DMN, visual system, and auditory system. We only expected the effect of pramipexole on the SMN, but not on the DMN, visual system, or auditory system. For the sensorimotor component, individual images were first entered into a one-sample t-test in SPM regardless of session to generate a group-level spatial map (p < 0.05 family-wise error [FWE]-corrected) and then into a paired-sample t-test to examine the effect of pramipexole administration (p < 0.05 FWE-corrected). Additional exploratory analyses were performed to ensure that there was no effect of pramipexole on the DMN and visual or auditory networks.
Graph theoretical analysis
The GTA was performed to examine how pramipexole modulates the topology of the whole-brain network. This analysis was implemented with the Brain Connectivity Toolbox (Rubinov and Sporns, 2010). Each cerebral hemisphere was segmented into 45 regions (total number, N = 90) using the Automated Anatomical Labeling (AAL; Tzourio-Mazoyer et al., 2002). Time series of each region were averaged across voxels and corrected for the effect of white matter, cerebral spinal fluid, and head movement. Regional mean time series were decomposed into wavelet coefficients over five scales (frequency intervals) with the maximum overlap discrete wavelet transforms (Bullmore et al., 2004). For each scale, correlations were computed for every possible pair of regional wavelet coefficients to generate an N-by-N matrix C, where C(i,j) represented the functional connectivity between regions i and j. The connectivity matrix was constructed separately under pramipexole and under placebo. We restricted the following analysis on the matrix of the scale 2 (0.06–0.12 Hz). This frequency interval was chosen because previous studies have typically reported strongest resting-state interregional correlations at low frequencies ∼0.1 Hz (Achard et al., 2006; Biswal et al., 1995; Lowe et al., 1998).
An adjacency matrix A can be derived from the connectivity matrix by applying a threshold τ. A(i,j) is unity (binary matrix) or original correlation (weighted matrix) if C(i,j) > τ and zero if C(i,j) ≤ τ. The adjacency matrix can further be represented as an undirected binary or weighted graph G, in which a node indicates a region and an edge drawn between two nodes indicates an interregional connection. In this study, we focused on the adjacency matrix of positive correlations as we obtained few negative correlations. The topological cost of a graph is defined as the number of edges in G divided by the maximum possible number of edges, N(N − 1)/2. Since the selection of threshold has a major impact on the topology of G, we monitored global and regional properties of weighted graphs (cost = 0.46) as well as binary graphs over the whole cost range of 0.01–0.46 at 0.01 intervals (Achard et al., 2006). Statistical tests were conducted by permutation (1000 times, p < 0.05) due to the lack of knowledge concerning the distribution of most network parameters (Bullmore and Sporns, 2009).
Four global parameters were measured, namely the global efficiency, clustering coefficient, small-word index, and modularity. The global efficiency measures the efficiency of information transfer throughout the whole-brain network (measure of integration). The clustering coefficient measures the degree to which regions of the whole-brain network tend to cluster together (measure of segregation). According to Watts and Strogatz (1998), small-world networks have a larger clustering coefficient but a small average shortest path length, compared to random networks. The small-world index captures the features of high segregation and high integration in a single statistic. The modularity reflects the degree to which the whole-brain network can be subdivided into nonoverlapping groups of regions (i.e., modules) in a way that maximizes the number of within-group connections and minimizes the number of between-group connections. The parameters are computed separately for binary graphs and for weighted graphs (for equations, see Rubinov and Sporns, 2010).
At region level, we focused on the degree, defined as the number of edges of a given node. The degree of the caudate nucleus was computed for weighted graphs and for binary graphs at each cost. Permutation tests were conducted to examine whether the degree of the caudate nucleus decreased and which caudate connectivity was reduced under pramipexole (1000 times, p < 0.05). The degree also indicates the relative importance of a particular region within brain networks. Regions having a degree of one standard deviation above the mean of all regions are defined as brain hubs and can further be classified as provincial hubs or connector hubs (Sporns et al., 2009). To examine whether the caudate nucleus is a hub, z-scores of degrees were calculated and compared with 1 (1000 times, p < 0.05).
Results
ICA results
The ICA identified four resting-state networks, including the SMN, DMN, visual system, and auditory system. The SMN comprised the post- and precentral gyri, SMA, paracentral lobules, superior and inferior parietal lobules, superior temporal gyrus, and caudate nucleus (Fig. 1A). Within the SMN, the right caudate nucleus showed weaker functional connectivity under pramipexole than under placebo (MNI [12 6 9], t = 7.83, p < 0.05 FWE-corrected; Fig. 1B). Additional analyses confirmed that the drug effect was absent in the DMN and visual and auditory networks.
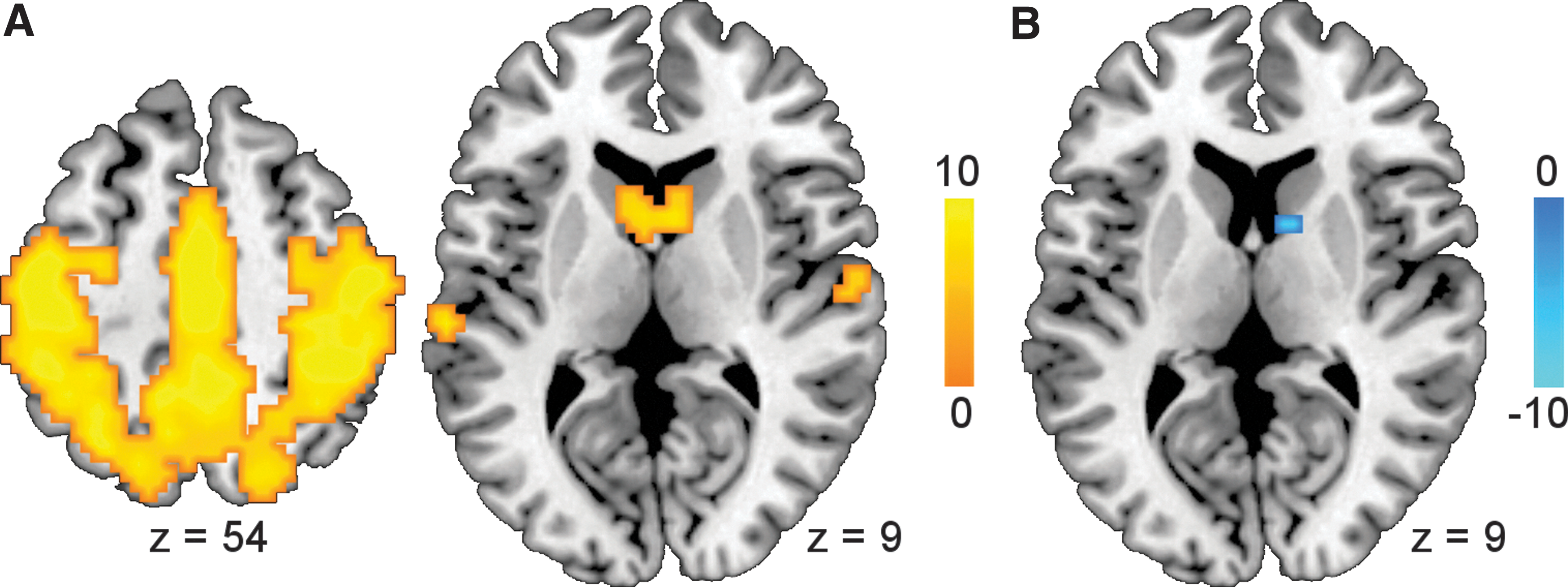
GTA results
At the global level, the whole-brain network under pramipexole was topologically similar to that under placebo (Fig. 2A–D). Global parameters such as global efficiency, clustering coefficient, small-world index, and modularity were measured for weighted graphs and for binary graphs over the whole cost range of 0.01–0.46 at 0.01 intervals and tested with permutation tests (1000 times, p < 0.05). None of these parameters differed significantly between medication conditions. The whole-brain network was identified as small-world network (small-world index >1), regardless of medication conditions.

At the regional level, degrees of the right caudate nucleus decreased under pramipexole in weighted graphs and in binary graphs over the cost range of 0.25–0.46 (1000 times, p < 0.05; Fig. 2E). However, the right caudate nucleus was identified as a nonhub region as the z-scores were significantly smaller than 1 in both weighted and binary graphs (1000 times, p < 0.05; Fig. 2F). Figure 2G illustrates the functional connectivity between the right caudate nucleus and regions of the four classical resting-state networks (priori definitions based on Beckmann et al., 2005; Damoiseaux et al., 2006) under placebo and pramipexole. The binary circle graphs were based on the group mean connectivity matrix with an arbitrary threshold of zero. The connectivity of the right caudate nucleus was weakened with the bilateral SMA, right precentral gyrus, right postcentral gyrus, and bilateral paracentral lobules of the SMN, and the right medial superior frontal gyrus of the DMN.
Discussion
This pharmacological resting-state fMRI study is an initial attempt to examine the action of D2 agonists from the perspective of network connectivity and topology. We found that pramipexole reduced the resting-state functional connectivity (from positive toward zero) between the caudate nucleus and the sensorimotor regions such as the SMA, precentral gyrus, postcentral gyrus, and paracentral lobules, but did not globally change the topology of the brain network. These findings were consistent with a recent resting-state fMRI study on Parkinson's patients ON and OFF levodopa (Berman et al., 2016). Like us, Berman and associates observed small-world properties of the whole-brain network regardless of medication condition. Increasing brain dopamine reduced local efficiency (a measure of segregation, such as cluster coefficient) of the dorsal attention network, but did not change the global efficiency of the whole-brain network. However, the chemical modulation of the topology of the resting brain is still an open question, given conflicting evidence from previous studies with D2 antagonist and noradrenergic reuptake inhibitor, which revealed both global and local changes of the network topology (Achard and Bullmore, 2007; van den Brink et al., 2016).
This observation dovetails nicely with recent neurocomputational models (Frank et al., 2004, 2007; Maia and Frank, 2011) according to which D2 agonists influence motor functions by distorting the striatal-motor pathway. Effects of D2 agonists in reality are quite diverse, however. Grace and colleagues (Bilder et al., 2004; Grace, 2000) stressed that dopamine dynamics in the striatum are driven by the interactions of phasic and tonic dopamine release. Phasic dopamine release is triggered by exogenous stimuli and transiently released dopamine is rapidly removed from the synaptic cleft via reuptake processes and is unlikely to influence extracellular dopamine levels. By contrast, tonic dopamine release is regulated by glutamatergic projections from the prefrontal cortex. The slowly but constantly released dopamine determines the general level of extracellular dopamine and therefore sets the rather stable baseline level of dopamine receptor stimulation. Alternations in tonic dopamine release may induce homeostatic processes that serve to restore the original stable level of dopamine receptor stimulation. This phasic–tonic interaction was recently observed in dopamine neurons and in neurons of the lateral habenula in monkey brains (Bromberg-Martin et al., 2010). In the resting state (with eye closed and no exogenous stimulus), pramipexole may lead to a decrease in extracellular dopamine level by downregulating tonic dopamine release. Animal work has shown that the stimulation of cortical D2 receptors may directly inhibit the activity of glutamate neurons in the prefrontal cortex and subsequently the activity of dopamine neurons in the striatum (Beyer and Steketee, 2000; Del Arco and Mora, 2005). Further studies need to clarify how the interaction of multiple medication-induced processes changes the striatal-motor connectivity in resting state and, more importantly, during motor or cognitive tasks.
In light of the known anatomical connectivity, it can be asked why a change of striatal-motor cortex connectivity rather than of striatal-thalamic-motor cortex connectivity was observed. The absence of the thalamus in the ICA and GTA may be due to the limitations of current imaging and analysis approaches. In this study, the ICA revealed the involvement of the thalamus in the DMN (left peak at [−6 −27 6], t = 7.12; right peak at [6 −21 9], t = 6.05), auditory system (MNI [6 −18 6], t = 11.68), and visual systems (MNI [−12 −21 6], t = 7.06) over a threshold of p < 0.05 FWE-corrected, although not in the SMN. However, additional analyses revealed that none of the default-mode, auditory, or visual networks showed a significant change under pramipexole. Visual inspection suggested that the thalamus also appeared in two artifact components revealed by the ICA. Statistical significance of the BOLD signals might be influenced by the insufficient signal-to-noise-ration (SNR) in the thalamus (at the 1.5T and with 6 min of scan time) and/or by the drug's effects on respiration and cardiac pulsatility (which are heterogeneous across the brain). In the GTA, no medication effects were observed for the degrees of the bilateral thalamus. In addition to the low SNR, the GTA may also be affected by the lack of thalamic subdivisions in current standard atlases. Subtle differences between subregional time series may be overlooked as the thalamus was defined as one single region in the AAL. Finally, it is worth noting that the wavelet correlation indicates the degree of possible dependence between two regional time series, not the structural connectivity between two regions. The change of striatal-motor functional connectivity may reflect the sum effect of connectivity changes in the direct and indirect pathways between the basal ganglia and the motor cortex.
In conclusion, we examined how the dopamine agonists such as pramipexole modulate the functional connections between the basal ganglia and the motor cortex by conducting a pharmacological resting-state fMRI with a double-blind randomized within-subject crossover design. We used two analysis approaches, the ICA and the GTA. The two analyses consistently revealed that pramipexole reduced the functional connectivity of the caudate nucleus in the SMA, paracentral lobule, and pre- and postcentral cortex. This observation was consistent with and contributed to the existing hypothesis concerning the mechanisms of dopamine agonists in the human brain.
Footnotes
Acknowledgments
T.F.M. is supported by grants of the DFG and the BMBF. Z.Y. is supported by the Thousand Young Talents Program.
Author Disclosure Statement
No competing financial interests exist.