
Abstract
The default mode network (DMN) is typically associated with off-task internal mentation, or with goal-oriented tasks that require self-referential processing such as autobiographical planning. However, recent reports suggest a broader involvement of the DMN in higher cognition. In line with this view, we report global connectivity changes that are centered on the main DMN hubs of precuneus and posterior cingulate cortex during a functional magnetic resonance imaging-based visuospatial version of the Tower of London planning task. Importantly, functional connectivity of these regions with the left caudate shows a significant relationship with faster reaction time to correct responses only during the high-demand planning condition, thus offering further evidence for the DMN's engagement during visuospatial planning. The results of this study not only provide robust evidence against the widely held notion of DMN disengagement during goal-oriented, attention-demanding, externally directed tasks but also support its involvement in a broader cognitive context with a memory-related role that extends beyond self-referential, internally directed mentation.
Introduction
A
One such mental process associated with the DMN that carries particular importance in our everyday life activities is autobiographical planning, that is, our ability to form, sequence, test, and retain strategies to achieve a given goal (Unterrainer and Owen, 2006). The most common test that is used to study this cognitive process in an experimental setting is the Tower of London (TOL) task (Shallice, 1982). In this test, participants are asked to move colored balls across three rods to achieve a goal configuration and are assessed for their visuospatial planning abilities. Positron emission tomography and functional magnetic resonance imaging (fMRI) studies of healthy control subjects have suggested that a distributed set of brain regions, including the dorsolateral prefrontal, anterior cingulate, premotor, posterior parietal, precuneal, striatal, cerebellar, and occipital cortices, show greater activation during TOL task performance relative to baseline conditions (Baker et al. 1996; Rowe et al. 2001; Schall et al. 2003). Further, left-hemispheric activity increases were observed in the dorsolateral prefrontal cortex, striatum, premotor cortex, supplementary motor area, precuneus, and inferior parietal cortices with increasing TOL task difficulty (Schall et al. 2003; van den Heuvel et al. 2003).
A considerable portion of these brain regions, which are associated with TOL execution, overlap with large-scale brain networks, including the fronto-parietal (FPN) and dorsal attention networks (DAN) that exist not only at rest but also during task performance (Cole et al., 2014). Moreover, planning tasks have also been linked to the DMN, but mainly in versions that require self-referential mental scene constructions (Andrews-Hanna et al. 2014; Buckner et al. 2008). In fact, Spreng et al. have observed greater connectivity between the DMN and FPN during a modified, real-life autobiographical planning version of the TOL, arguing that the DMN may be involved in goal-directed paradigms, but only when the task goals require access to long-term memory and self-referential processing associated with this network (Spreng et al., 2010).
However, a collection of studies also describe task relevant activity in the main DMN hubs (medial prefrontal cortex, posterior cingulate cortex, precuneus, and angular gyri) during the original, visuospatial version of the TOL task (Boghi et al. 2006; Schall et al. 2003; Unterrainer and Owen 2006; van den Heuvel et al. 2003). For example, activity in the precuneus/posterior cingulate cortex (PCUN/PCC) has been associated with TOL task difficulty (van den Heuvel et al., 2003). Thus, the contribution of the DMN to the higher executive function of planning may not be confined to self-referential cognition but may also involve processing of non-self-referential, external information. In other words, there might be simpler, sub-processes carried out by the DMN that give rise to superordinate complex functions such as autobiographical planning. In line with this hypothesis, recent studies propose that the DMN is contributing to tasks that require access to working memory (Esposito et al. 2009; Hampson et al. 2006; Konishi et al. 2015; Vatansever et al. 2015a, 2016), a key component of planning abilities (Unterrainer and Owen, 2006).
Given this evidence, as well as the DMN's extensive structural and functional connections with remote brain regions (Buckner et al. 2009; Hagmann et al. 2008), we argue for a broader memory-related role for the DMN that extends beyond its link with spontaneous cognition during unconstrained rest or activity limited to self-referential paradigms. We, therefore, propose that DMN processes may continue to exist during a variety of higher cognitive states, including planning, but at different levels of attenuation based on the task demands for access to working memory stores (in contrast to perceptual processing). In this regard, the connectivity mechanisms through which specific DMN regions switch from a high-demand visuospatial planning to a baseline condition have not been previously assessed. In addition, the behavioral significance of such alterations in DMN connectivity in response to changing task demands remains to be elucidated.
For this purpose, in an fMRI version of the classic TOL paradigm, we assessed the potential contribution of the DMN to the higher cognitive process of planning and related task performance to functional connectivity. For an objective approach, we analyzed these data with a technique that does not require a priori selection of a seed region, called the intrinsic connectivity contrast (ICC) (Martuzzi et al., 2011). The major aim was to characterize global connectivity differences between planning and baseline conditions to delineate the possible contribution of specific DMN regions to higher cognitive processing. Given previous findings of a greater PCUN/PCC engagement observed with increasing TOL task load (Schall et al. 2003; van den Heuvel et al. 2003), we hypothesized that the connectivity of the PCUN/PCC during planning, which requires the generation and online retention of testable strategies to achieve task goals, would be associated with task performance.
Materials and Methods
Participants
This study was approved by the local Cambridge University Hospitals NHS Trust Ethics Committee, and informed consent was obtained from all participants. A total of 22 healthy subjects (19–57 years old, mean = 35.0, SD = 11.2, 9:13 female to male ratio) took part in this study. They had an average National Adult Reading Test (NART) score of 117.1 (SD = 5.76) and a mean Mini Mental State Examination score of 29.33 (SD = 0.85). According to the exclusion criteria, no history of drug or alcohol abuse, psychiatric and neurological disorders, head injury, medication use that may have affected cognitive performance (e.g., tricyclic antidepressants), physical handicap that may have prevented completion of testing, left-handedness, contraindication of magnetic resonance imaging scanning, or severe claustrophobia were reported in any of the participants.
Experimental paradigm specifications
The experimental paradigm was based on the original TOL task (Shallice, 1982), modified for fMRI with an event-related design (Baker et al., 1996) (Fig. 1A). In this task, the participants were presented with two pictures on a single screen containing “start” and “target” configurations of colored balls (blue, yellow, red, green) on three vertical rods, descending in heights from left to right with a capacity to hold 3, 2, and 1 balls, respectively. The participants were asked to determine the minimum number of moves required to match the bottom (start) with the top (target) picture. The balls had to be moved one at a time, but they could jump a rod, that is, a green ball from the first rod could move to the third rod, which would count as 1 move. In the control condition, the participants were asked to count the number of balls on the screen, thus matching the task condition in regards to the basic visuospatial information and response selection, but lacking the aspect of planning. It is important to note that “start” and “target” cues were not presented in the control condition to avoid confusion. The participants responded by pressing a button pad with four buttons, numbered from one to four to indicate their answers. The required number of moves, varied between one and four, were presented in a random sequence and interleaved with the control condition.
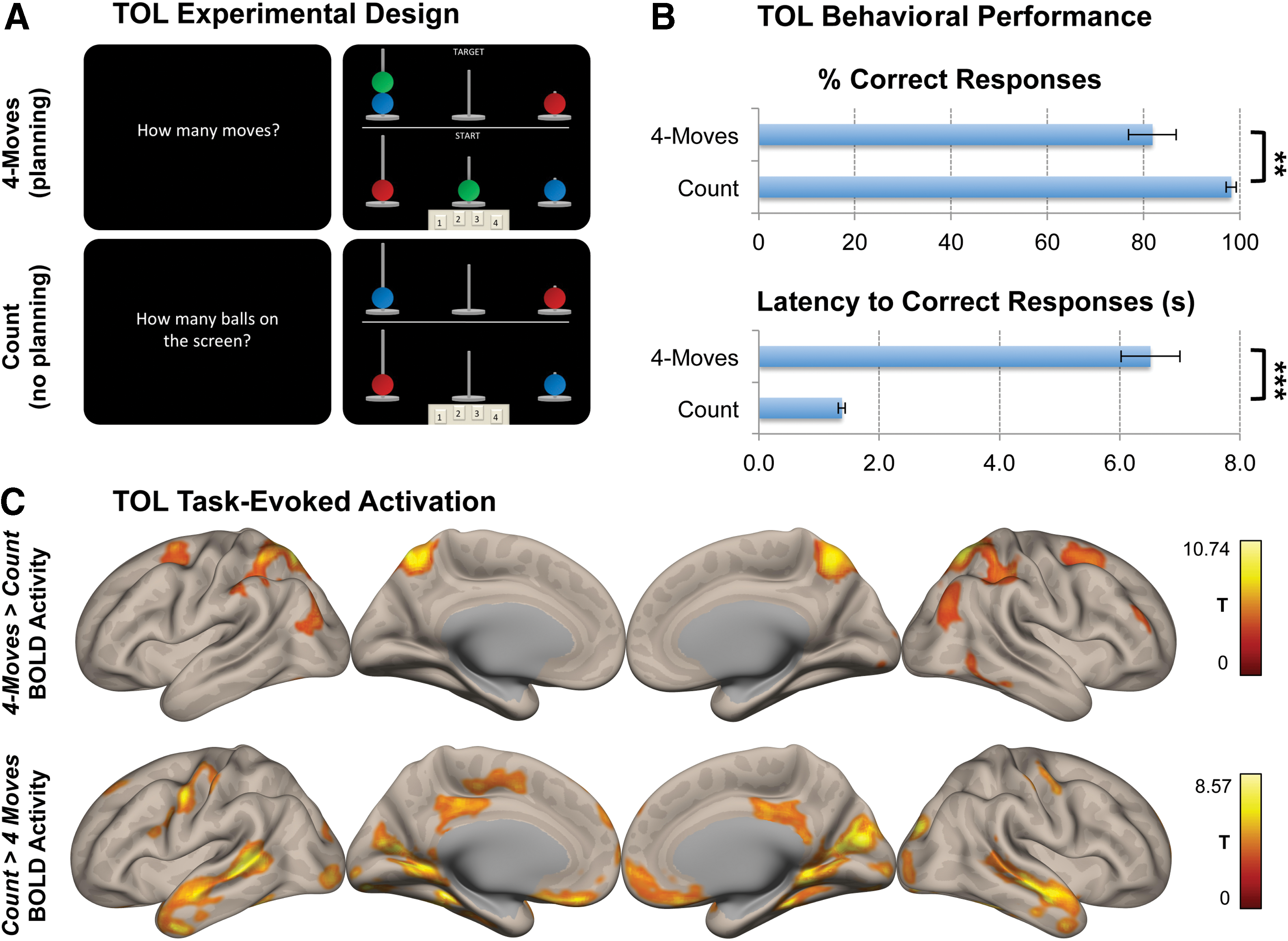
Experimental specifications, behavioral performance, and task-evoked brain activity in the TOL paradigm.
Instructions indicating “How many moves?” or “How many balls on the screen?” were presented on the screen for 2500 msec. The participants were allowed 15,000 msec to record their answers by pressing the button pad, after which the next trial appeared. If the subjects did not respond by the allotted time, the trial was marked as a missed response. The counting trials on the other hand were divided into 2500 msec of instructions and 5000 msec allotted time for a response. In total, there were 10 trials for each of the 1, 2, 3, 4-moves and 10 count trials, with 50 trials in total.
Neuroimaging data acquisition
Using a Siemens MAGNETOM Tim Trio 3T scanner at the Wolfson Brain Imaging Centre, Cambridge, the imaging protocol began with a localizer followed by a high-resolution T1-weighted, magnetization-prepared rapid gradient echo (MPRAGE) structural scan (repetition time [TR] = 2300 ms; echo time [TE] = 2.98 ms; acquisition time [TA] = 9.14 min; flip angle = 9°; field of view read = 256 mm; voxel size = 1.0 × 1.0 × 1.0 mm, slices per slab = 176). After a session of eyes-closed resting-state fMRI scanning, a set of randomized task acquisitions was carried out, including the TOL paradigm. The echo-planar imaging parameters for the task scans were as follows: TR = 2000 msec; TE = 30 msec; flip angle = 78°; field of view read = 192 mm; voxel size = 3.0 × 3.0 × 3.0 mm; slices per volume = 32; volumes = 390.
Neuroimaging data preprocessing
The image analyses were carried out by using SPM Version 8.0, based on the MATLAB Version 12a platform. After the removal of the first five volumes to achieve steady-state magnetization, the data were slice-time adjusted and corrected for motion artifacts by using six degrees of freedom (x, y, and z rotations as translations). Structural images were co-registered to the mean fMRI image via rigid transformation and subsequently were segmented into gray/white matter and cerebrospinal fluid probability maps. All functional data were then spatially normalized to Montreal Neurological Institute (MNI) space by using the segmented high-resolution structural gray matter image and an a priori template. This procedure utilized the unified segmentation–normalization framework, which combines tissue segmentation, bias correction, and spatial normalization in a single unified model (Ashburner and Friston, 2005). Finally, the data were smoothed with an 8 mm FWHM Gaussian kernel and were carried on to further statistical analysis.
The functional connectivity analyses were carried out by using the Conn functional connectivity toolbox (Whitfield-Gabrieli and Nieto-Castanon, 2012). For this, the condition-specific bloody-oxygenation-level-dependent (BOLD) signal time series first had to be separated into event-specific scans by using the onsets and response time durations of each task condition (i.e., variable epoch model). These regressors were then convolved with a rectified hemodynamic response function to account for the delay in hemodynamic response. Finally, time series for each event type were concatenated with the scans associated with nonzero effects, and they were appropriately weighted. Such an analysis pipeline de-weights the beginning and end of each task condition to eliminate the signal from non-biological origins at the points of concatenation, minimizing the potential cross-talk between adjacent task conditions. Further details of this procedure are explained in Vatansever et al. (2015b) and Whitfield-Gabrieli and Nieto-Castanon (2012).
Next, variance attributable to nuisance variables was removed by using linear regression, which included six motion artifact parameters and their first-degree derivatives, a linear detrending term, and five CompCor principle components representing the white matter and cerebrospinal fluid signals, extracted by using individual masks obtained from the T1-weighted segmentation (Behzadi et al., 2007). This procedure eliminated the need for global signal normalization that is reported to introduce spurious anti-correlations (Chai et al., 2012; Murphy et al., 2009). The main task effects and their first-order derivatives were also included in the statistical analysis as potential confounds with the aim of removing task-specific coactivation effects (Fair et al., 2007). In addition to the low-frequency fluctuations that characterized early resting-state investigations (Fox et al., 2005), recent studies indicate the detectable presence of large-scale brain networks at high frequencies during task conditions (Boubela et al., 2013; Cole et al., 2014). Thus, given the event-related design of the TOL task, no low-pass filtering was employed.
Task-activation analysis
With the aim of confirming that the task has produced results expected from previous TOL studies, our initial objective was to contrast 4-moves high-demand planning and count control conditions using the general linear model. Subject-specific 4-moves > count and count > 4-moves contrasts were carried over to group-level one-sample t-tests, with age introduced as a potential confound. All reported clusters were corrected for multiple comparisons by using the Family-Wise Error (FWE) detection technique at the p = 0.05 level of significance (p = 0.001 voxel level, uncorrected).
ICC analysis
The main objective in this analysis was to examine whether DMN areas are involved in visuospatial planning. With that goal in mind, the connectivity analysis was initiated with an objective, whole-brain voxel-based metric called the ICC that does not require a priori selection of a seed region. This novel measure utilizes graph theoretical metrics to objectively define how well each voxel is connected to the rest of the brain.
An earlier version of this metric that solely relied on the graph measure of degree has been employed in an Alzheimer's disease study to investigate changes resulting from the disease process in the brain's cortical hubs (Buckner et al., 2009). Further extending this metric, Martuzzi et al. (2011) introduced a measure of connectivity strength by weighting the connections of each voxel with their average r 2 value, therefore eliminating the need for an arbitrary threshold. Specifically, the ICC score denotes the squared sum of mean correlations, with a greater ICC score representing greater average strength of the correlations in a given voxel (Martuzzi et al., 2011). Two recent publications have used this method to characterize whole-brain voxel-wise connectivity: One examined angular DMN connectivity in an n-back working memory task (Vatansever et al., 2016), and another looked at the effect of short-term gravitational changes on brain connectivity (Van Ombergen et al., 2017).
Using this method, the ICC was calculated for each voxel, thus producing subject-level, condition-specific (count, 1, 2, 3, and 4-moves) whole-brain ICC maps that were normalized by subtracting the mean and dividing by the standard deviation. Given our interest in DMN regions that showed the greatest alterations in their global connectivity during the TOL paradigm, we focused only on the high-demand planning (4-moves) and control (count) conditions. To assess differences in global connectivity between these two conditions, the corresponding ICC maps were entered into a paired t-test (results thresholded at voxel-level uncorrected p < 0.001, cluster-level FWE corrected for multiple comparisons p < 0.05).
Region of interest-to-voxel functional connectivity analysis
Moreover, with the aim of assessing the spatial extent of the functional connectivity of the cluster with the highest change in global connectivity, we also employed conventional seed-based functional connectivity analysis. For that purpose, the global peak cluster from the ICC paired t-test (denoting the most significant global connectivity changes across the two task conditions) was used as a seed region of interest (ROI) in two further seed-based functional connectivity analyses: one for 4-moves planning and another for the count control condition. Group-level inferences were made by using a one-sample t-test for each condition. A paired t-test between the two conditions was also employed to identify any connectivity changes with altered task demands.
Behavioral correlation analysis
The final question to answer was whether the extent of the functional connectivity from the selected ROI would show a relationship with behavior during two distinct stages of the TOL task, namely the 4-moves high-demand planning and count conditions. The behavioral measure we used was reaction time to correct responses, which has been previously shown to be a reliable performance indicator in working memory and TOL paradigms (Kitzbichler et al., 2011; Newman et al., 2003; van den Heuvel et al., 2003). For this analysis, we employed a multiple linear regression by using the beta maps obtained from the seed-based functional connectivity analyses and our selected behavioral measure, that is, reaction times to correct responses. Age was included as a potential confound in this analysis, and all results were corrected for multiple comparisons by using the FWE method at the p = 0.05 level of significance (p = 0.001, voxel wise, uncorrected).
Results
Behavioral performance and task-evoked brain activity in the TOL paradigm
The main objective of this study was to detect any global changes in functional connectivity centered on the DMN regions in the high-demand planning condition in comparison with the count control condition, and to establish a possible relationship between connectivity and performance. For that purpose, we first assessed behavioral results to confirm the differences in performance between the 4-moves (high-demand planning) and count conditions (low-demand control). The percent correct responses decreased from 98.2% (SD = 5.0) in the count to 81.8% (SD = 23.2) in the 4-moves (p = 0.003) condition, and the reaction time to correct responses increased from 1380.8 msec (SD = 267.1) in the count to 6510.2 msec (SD = 2310.4) in the 4-moves condition (p < 0.001) (Fig. 1B). These results confirm that the participants performed the task as expected. Moreover, in line with previous TOL studies (Newman et al., 2003; Newman et al., 2009; Schall et al., 2003), the task activation analysis for the 4-moves > count contrast revealed regions commonly associated with planning tasks, including the precuneus, bilateral inferior/superior parietal lobes, bilateral frontal eye fields, the right middle frontal gyrus, and right inferior temporal lobe with a global peak located on the left cerebellar crus I (p < 0.0001, MNI: −42 − 56 − 36). The reverse contrast, on the other hand, revealed greater activity on the cuneus, posterior cingulate and medial prefrontal cortices, calcarine and lingual gyri, bilateral pre/postcentral gyri, and the inferior/middle temporal gyri with a global peak centered on the left fusiform gyrus (p < 0.0001, MNI: −34 − 42 − 18) (Fig. 1C).
Global connectivity changes with altered task demands
With the aim of objectively identifying brain regions that alter their connectivity with planning-based demands, we first employed a whole-brain metric called ICC, a normalized voxel-based measure denoting the average strength of global brain connectivity.
The group-level 4-moves > count contrast revealed greater global connectivity during the 4-moves planning condition in four clusters: one with a global peak in the left precuneus extending to the posterior cingulate cortex (p < 0.0001, MNI: −4 − 52 36), a local peak in the left superior parietal lobe (p = 0.002, MNI: −22 − 72 54), as well as clusters in the right supramarginal gyrus (p = 0.029, MNI: 54 − 30 46) and left cerebellar crus II (p = 0.022, MNI: −6 − 80 − 30) (Fig. 2).
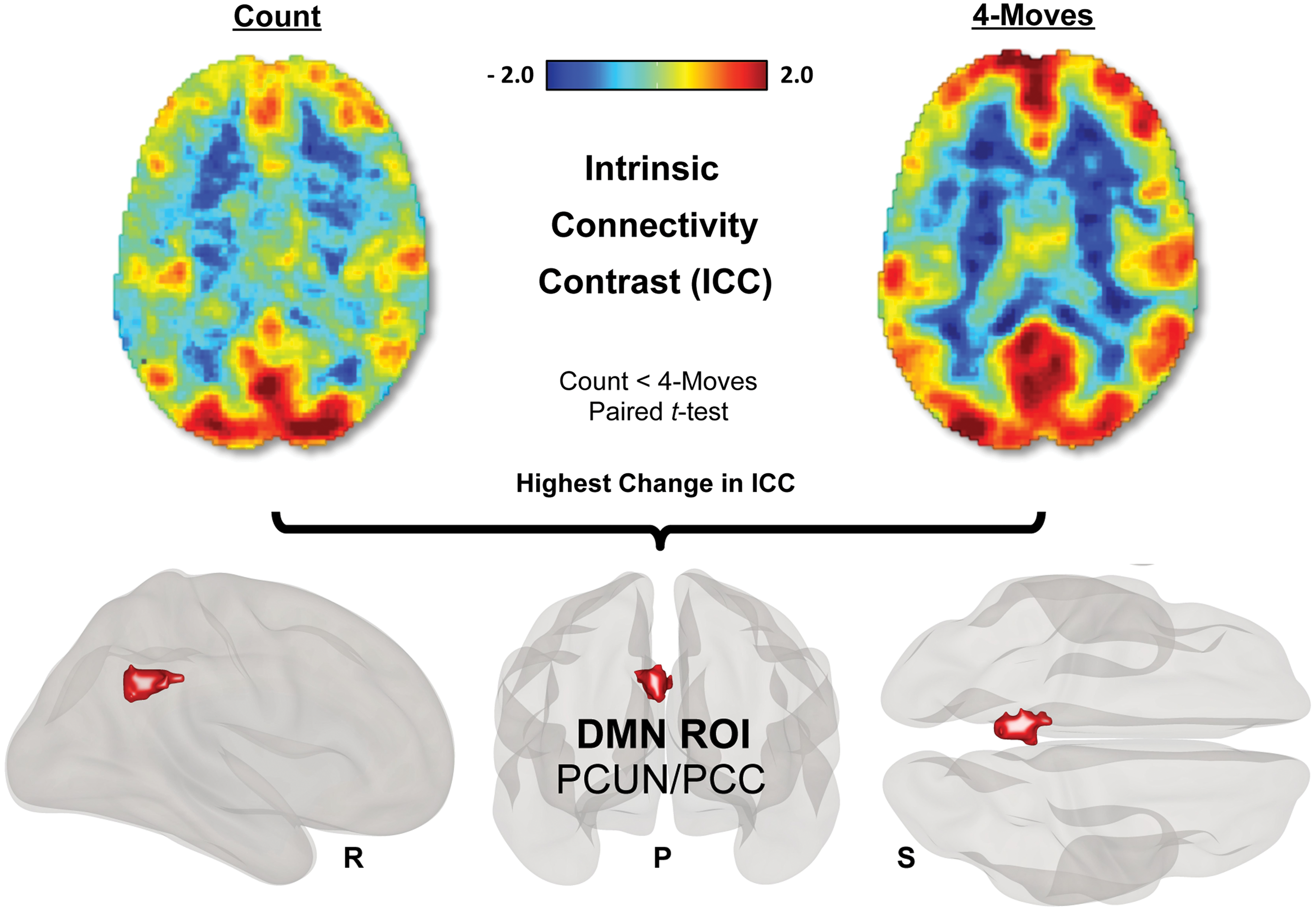
Global brain connectivity alterations between 4-moves (high-demand planning) and count (low-demand control) conditions in the TOL paradigm. The red to blue ICC maps show the most and the least globally connected regions of the brain. A paired t-test for the 4-moves > count contrast revealed a PCUN/PCC cluster as the global peak (p < 0.0001, MNI: −4 − 52 36), as well as (not shown) clusters in the left superior parietal gyrus (p = 0.002, MNI: −22 − 72 54), right supramarginal gyrus (SMG) (p = 0.029, MNI: 54 − 30 46), and cerebellar crus II (Cer Crus II) (p = 0.022, MNI: −6 − 80 − 30), cluster corrected for multiple comparisons by using Family-Wise Error at the 0.05 level of significance (p = 0.001 voxel level, uncorrected) (R = right hemisphere, P = posterior, S = superior view). The PCUN/PCC cluster is considered a major DMN hub and was subsequently used as an ROI for further functional connectivity analyses. ICC, intrinsic connectivity contrast; PCUN/PCC, precuneus/posterior cingulate cortex; ROI, region of interest. Color images available online at
All of these brain regions were previously reported to show greater engagement with higher task demands in TOL paradigms (Schall et al., 2003; van den Heuvel et al., 2003). From a large-scale brain networks perspective, the PCUN/PCC areas are considered a major hub of the DMN (Fransson, 2005; Fransson and Marrelec, 2008), whereas the superior parietal cortex is considered part of the DAN (Fox et al., 2005), and the supramarginal gyrus part of the FPN (Vincent et al., 2008). The cerebellum is rarely discussed in the context of large-scale brain networks; however, recent reports indicate that the cerebellar crus II may be part of the FPN (Buckner et al., 2011; Habas et al., 2009).
Functional connectivity of the DMN persists across TOL
After gaining valuable information from the ICC analysis on the changes in global brain connectivity strength associated with task demands, further seed-based functional connectivity analysis was carried out to reveal the spatial extent of the temporal connectivity patterns during the planning and control conditions. For this purpose, the cluster centered on the PCUN/PCC (i.e., the highest change in ICC) was chosen as a seed.
The results indicated that the PCUN/PCC seed was functionally connected with established DMN regions (e.g., medial prefrontal, angular gyri and medial temporal lobe) and negatively connected with parts of the DAN (e.g., superior parietal cortex) regions in both the planning and control conditions (Fig. 3A, B). A paired t-test between the two conditions revealed no significant differences.
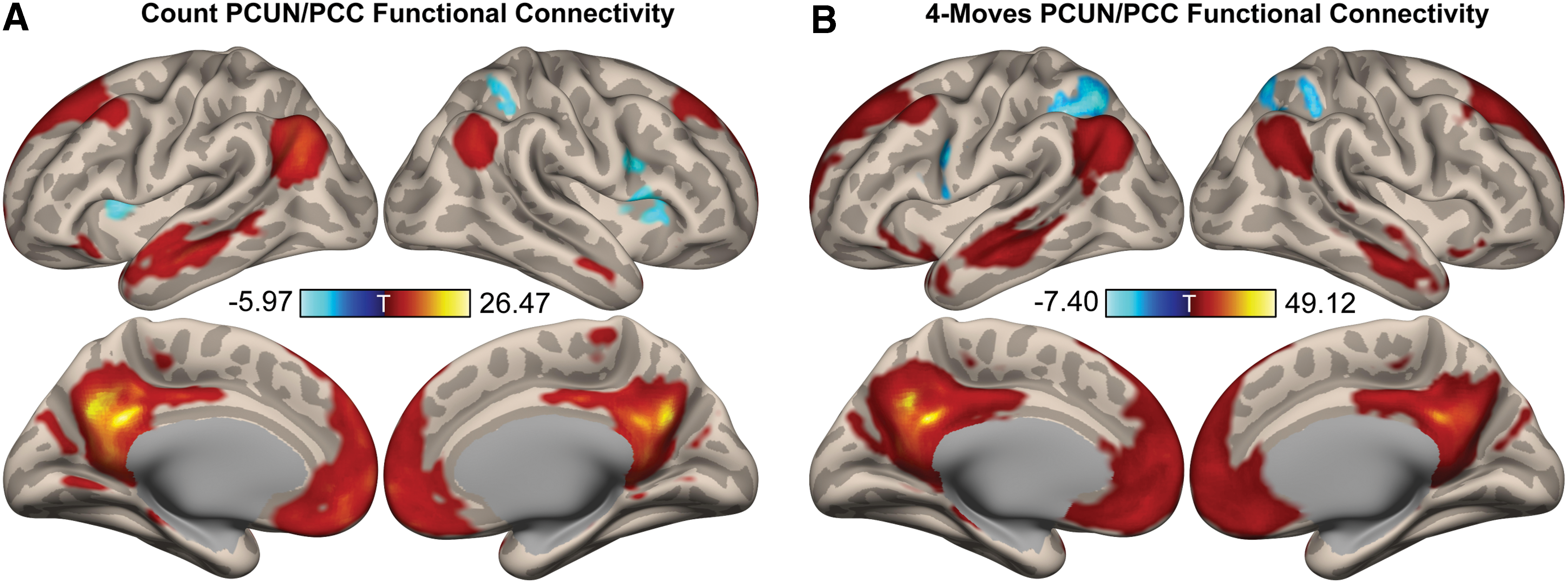
PCUN/PCC functional connectivity in the count and 4-moves TOL conditions. In both the
DMN-caudate connectivity correlates with task performance
The results so far have indicated that the PCUN/PCC region increases its global connectivity strength with the rest of the brain in the 4-moves planning condition. However, no differences were observed in the spatial extent of its functional connectivity, suggesting this network's persistence in the two selected conditions of the TOL paradigm. The next question was whether behavioral performance observed during the task correlates with PCUN/PCC connectivity to the rest of the brain in either the planning or the control conditions.
The whole-brain multiple linear regression analysis indicated that stronger PCUN/PCC connectivity with a left caudate cluster correlated with faster reaction times to correct responses, illustrating a clear separation between the 4-moves planning and count control conditions (Fig. 4). In the 4-moves high-demand planning condition, the variability in task performance could be explained by individual differences in the PCUN/PCC and caudate connectivity (p < 0.0001, R2 = 0.89); however, this correlation did not reach significance in the count condition (p = 0.051, R2 = 0.18).
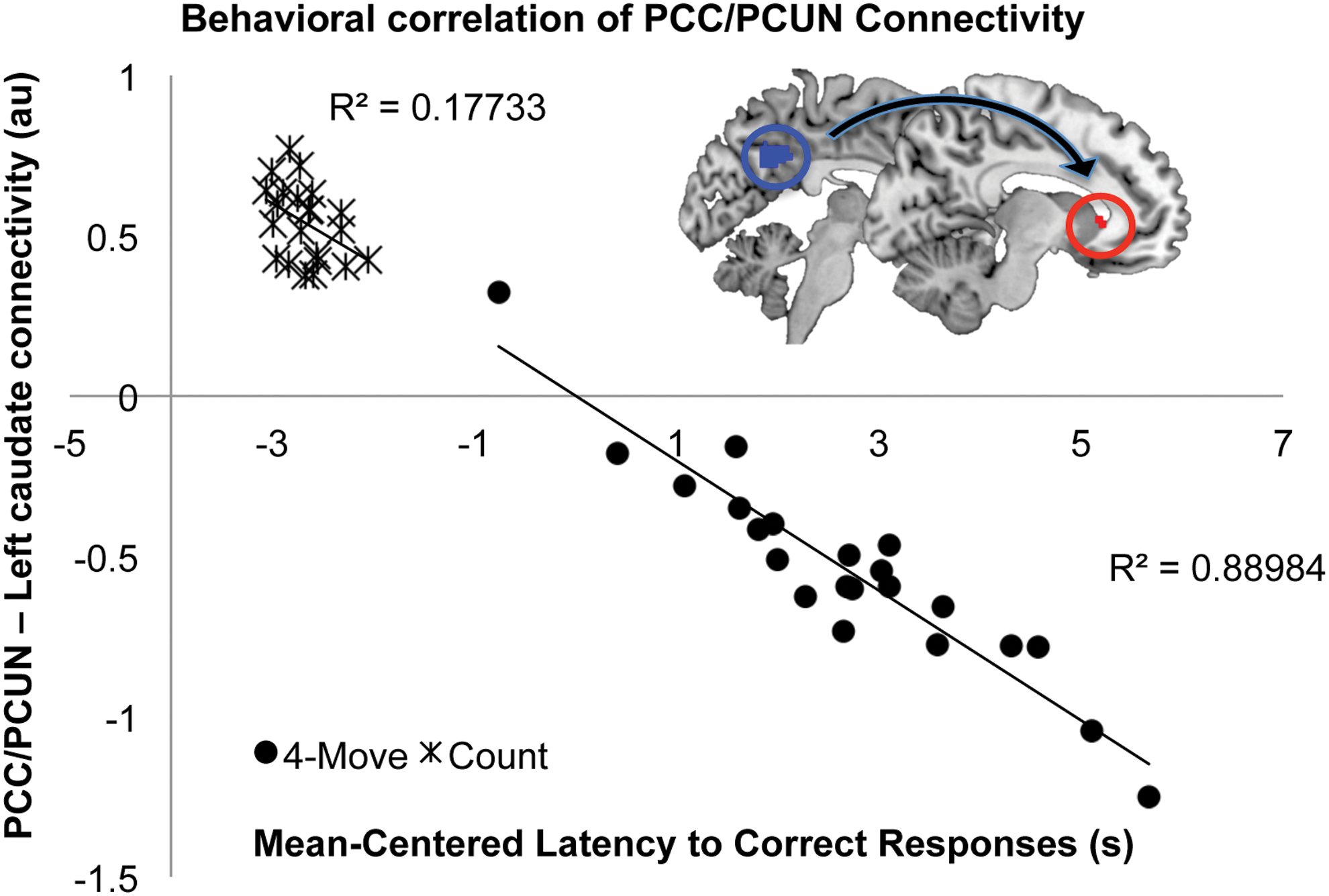
PCUN/PCC functional connectivity during TOL and its relationship with task performance. In the high-demand 4-moves planning condition, the variability in task performance (mean-centered latency to correct responses) could be explained by individual differences in the DMN-left caudate connectivity (mean-centered beta values) (p < 0.0001), whereas this correlation did not reach significance in the count control condition (p = 0.051). While the seed region is represented in blue, the target region is colored red. Color images available online at
Previous studies have indicated basal ganglia involvement in the performance of the TOL task (Newman et al., 2009) and suggested a potential role of the caudate head in this executive function, especially when the task involved sequencing of strategies to achieve a desired goal (Melrose et al., 2007). Further, other reports have indicated sustained left caudate head activity during a modified version of the TOL task, assessing the effects of learning (Beauchamp et al., 2003). The results presented in this study indicate that greater coupling between a major DMN hub and the caudate supports better performance in the high-demand planning condition of the visuospatial TOL paradigm.
Discussion
The aim of this study was to investigate the contribution of DMN regions to the higher executive function of visuospatial planning. We employed an ICC technique to map the changes in global connectivity between the count control and 4-moves planning conditions, attempting to provide evidence for a DMN role that extends beyond autobiographical, self-referential planning. The most significant change in global connectivity was detected in a PCUN/PCC cluster. Further seed-based functional connectivity analysis revealed the spatial extent of this region's connections. From a systems perspective, areas implicated strongly included the DMN, as well as parts of the DAN. There were no significant differences in the spatial extent of this region's connectivity between the two selected conditions, suggesting the network's persistence throughout the TOL paradigm. However, only when participants were engaging with the high-demand planning condition, greater connectivity between the PCUN/PCC and left caudate head was related to faster task performance. This indicated that the relationship between these two regions, one of which constitutes a major DMN hub, is important for visuospatial planning.
Both high metabolic activity (Raichle et al., 2001) and functional connectivity at rest (Greicius et al., 2003) have contributed to the definition of an intrinsic default mode of brain function (Gusnard et al., 2001). However, one central question that still remains to be answered is the exact contribution of the DMN to human cognition. In this regard, tasks that involved self-referential and autobiographical processes were all shown to activate DMN regions (Andrews-Hanna et al., 2014). This has led to the internal mentation hypothesis in which the DMN was suggested to reconstruct mental scenes for an adaptive advantage (Buckner and Carroll, 2007). Though such spontaneous cognition based on internally generated information may occur during unconstrained states of rest (Christoff et al., 2009) and paradigms involving self-relevant processing (Addis et al., 2007), if they, indeed, represent a “default” mode of brain function, DMN processes are likely to exist in an ongoing manner, even during non-self-referential, externally directed tasks (Vatansever et al., 2015b). To this end, better performance in a higher cognitive, attention-demanding n-back working memory task was previously associated with greater DMN connectivity (Esposito et al., 2009; Hampson et al., 2006), suggesting a more general memory-related role for the DMN that extends beyond long-term memory-based self-referential internal mentation.
Supporting this hypothesis, the initial analysis in this study illustrated that a major DMN hub, the PCUN/PCC, showed the greatest change in connectivity with the rest of the brain in response to the change in task demands. Previous studies have also implicated DMN regions such as PCUN/PCC in TOL planning paradigms (Schall et al., 2003), and revealed an association between greater task load and increased activity (van den Heuvel et al., 2003). Although the involvement of precuneus in either the FPN or the DMN (Buckner et al., 2008; Margulies et al., 2009) has been debated, recent reports have established it as a core DMN region (Utevsky et al., 2014) contributing to task performance in attention-demanding paradigms such as working memory (Fransson and Marrelec, 2008). From a higher cognitive perspective, PCUN/PCC is believed to be involved in the mentalization of potential strategies, sequencing, as well as the tracking of response outcomes (Cavanna and Trimble, 2006), processes that are all relevant to the TOL paradigm employed in this study.
Further, although the spatial extent of the PCUN/PCC connectivity remained stable across the selected control and planning conditions, specifically in the 4-moves, our findings point to a relationship between connectivity of the PCUN/PCC to the left caudate head and TOL task performance. Previous studies have shown that the left caudate activity increased with greater task difficulty (van den Heuvel et al., 2003), and activity in the caudate was reported to persist throughout a learning version of the TOL paradigm (Beauchamp et al., 2003). Hence, these findings suggested that the caudate might be important in visuospatial planning through acquisition and automatization of information processing.
Taken together, the results of this study indicate that the PCUN/PCC and caudate might support TOL task performance by allowing the sequencing of sub-goals and mentalization of potential outcomes to reach the goal state. During a TOL paradigm, to achieve task goals, the participants have to identify the current and goal configurations, generate sub-goals with testable hypotheses, and execute decisions based on predicted and observed outcomes. These memory-related processes all require online access to learned information, in line with previous studies, thus suggesting a positive correlation between DMN connectivity and working memory task performance (Esposito et al., 2009; Hampson et al., 2010; Hampson et al., 2006).
Similar to the episodic simulations attributed to the DMN during internal mentation tasks (Buckner and Carroll, 2007), the visuospatial planning here may also rely on associative learning and event simulation processes that are attenuated during the control condition. Thus, the behavioral correlation in our study may represent a broader version of this mechanism that requires the associative integration of information for conscious processing (Baars, 2002; Smallwood et al., 2012; Vatansever et al., 2015a). However, further research delineating the different stages of planning will be necessary to decipher the detailed contribution of the DMN and caudate to this higher cognitive process. Nevertheless, the results in this study support the notion that the DMN might be involved in cognition that extends beyond self-referential processing, and may contribute to externally directed tasks.
Conclusion
In conclusion, the results of this study further challenge the status quo on the disengagement of the DMN during higher cognitive tasks, and they provide evidence for its engagement during task performance in a TOL paradigm that assessed visuospatial planning abilities. Moreover, the contribution of such memory-related DMN functions in non-self-referential tasks indicates that the DMN may play a more fundamental role, which can be further explored by utilizing a variety of higher cognitive paradigms. Such task-based manipulations would provide more information on the exact DMN contribution to human cognition.
Footnotes
Acknowledgments
The funding for this study was provided by the Evelyn Trust (RUAG/018). In addition, D.V. received funding from the Yousef Jameel Academic Program, D.K.M. was funded by the MRC, the NIHR Cambridge Biomedical Centre (RCZB/004), and an NIHR Senior Investigator Award (RCZB/014), and E.A.S. was supported by the Stephen Erskine Fellowship Queens' College Cambridge and FP7-funded BioCog. The authors would also like to thank Dr. Sanja Abbott for programming the stimulus delivery, as well as Dr. Guy Williams, Victoria Lupson, and the rest of the staff in the Wolfson Brain Imaging Centre (WBIC) at Addenbrooke's Hospital for their assistance in scanning. Last but not least, the authors thank all the participants for their contribution to this study.
Author Disclosure Statement
No competing financial interests exist.