
Abstract
Numerous studies have identified several large-scale networks within the brain of healthy individuals, some of which have been attributed to ongoing mental activity during the wakeful resting state. While engaged during specific resting-state functional magnetic resonance imaging (fMRI) paradigms, it remains unclear as to whether traditional block-design simple movement fMRI experiments significantly influence these mode networks or other areas. Using blood-oxygen level-dependent fMRI, we characterized the pattern of functional connectivity in healthy subjects during a resting-state paradigm and compared this with the same resting-state analysis performed on motor task data residual time courses after regressing out the task paradigm. Using seed-voxel analysis to define the default mode network, the executive control network (ECN), and sensorimotor, auditory, and visual networks, the resting-state analysis of the residual time courses demonstrated reduced functional connectivity in the motor network and reduced connectivity between the insula and the ECN compared with the standard resting-state data sets. Overall, performance of simple self-directed motor tasks does little to change the resting-state functional connectivity across the brain, especially in nonmotor areas. This would suggest that previously acquired fMRI studies incorporating simple block-design motor tasks could be mined retrospectively for assessment of the resting-state connectivity.
Introduction
The complex and dynamic system that is the human brain continually generates a myriad of oscillatory waves believed to reflect cyclic modulation of gross cortical excitability and long distance neuronal synchronization (Balduzzi et al., 2008; Buzsaki and Draguhn, 2004). The coordinated changes in this activity and the excitability of large number of neurons underlie these waves, seen as spontaneous fluctuations on the electroencephalogram, which range from <0.1 Hz to >30 Hz, and were first observed almost a century ago. The blood-oxygen level-dependent (BOLD) functional magnetic resonance imaging (fMRI) signal also exhibits spontaneous low frequency (< 0.1 Hz) fluctuations, which occur at all times—during quiet wakefulness, task-performance, and even sleep—and has been shown to be related to fluctuations in neural activity (He et al., 2008). In the absence of overt task performance or stimulation, the spatial distribution of temporal correlations among these spontaneous fluctuations in BOLD signal is referred to as resting-state functional connectivity (RSFC).
An understanding of the baseline or control state is fundamental for an overall appreciation of complex systems. Defining a baseline state in the human brain is, therefore, necessary to deepen our knowledge of neuroscience. Study of RSFC is still a relatively new area of BOLD fMRI investigation but has become increasingly popular. It provides insight into the intrinsic functional architecture of the human brain and the interdependency of different regions. The experimental design that dominated the early years of BOLD fMRI research was a block-design paradigm in which two alternating tasks were performed, often an active block, during which a movement or language task was performed, alternated with a period in which no task was performed. Paradigms using 15-sec (Jurkiewicz et al., 2007; Mikulis et al., 2002) or 30-sec (Brockway, 2000; Loubinoux et al., 2001) blocks produced robust activation maps, and were commonly used block durations in BOLD fMRI. Unlike conventional fMRI studies, no specific task is prescribed for the subject at any time during data collection in an RSFC experiment; however, the spontaneous firing of neurons, even in the resting state, is followed by regional cerebral blood flow increases (Golanov et al., 1994) that can be measured as spontaneous fluctuations in the fMRI signal. Brain regions whose spontaneous fluctuations covary with one another are interpreted as functionally connected.
In a simple and commonly used RSFC analysis, a map of the temporal correlations of MR signal time courses is computed by specifying a region of interest (a “seed”) in a functional area and cross-correlating it with all other voxel time courses in the brain. This type of analysis and others on BOLD fMRI data have identified multiple so-called large-scale brain networks, including the dorsal attention (Bell and Shine, 2015; Yuan et al., 2016), salience (Yuan et al., 2016), lateral visual (Bell and Shine, 2015; Yuan et al., 2016), and default mode networks (DMNs) (Bell and Shine, 2015; Meindl et al., 2009; Raichle et al., 2001; Yuan et al., 2016). Analysis of BOLD fMRI data acquired specifically for study of the resting state has uncovered a reproducible pattern of engaged areas that have been dubbed the “default mode network” (DMN) represented by the posterior cingulate cortex (PCC)/precuneus, medial prefrontal cortex, and the lateral parietal cortex (Meindl et al., 2009; Raichle et al., 2001).
Whether the connectivity map revealed by resting-state covariance analysis is confounded by specific constraints of a task is not well established. Studies examining how cognitive (Fair et al., 2007; Wang et al., 2012), auditory (Hasson et al., 2009), and visual (Arfanakis et al., 2000; Lewis et al., 2009) tasks modulate the resting state have in some cases found little task effect on the resting state, whereas in others, a marked effect was observed. In one study, Fair et al., 2007, used a block-design BOLD fMRI paradigm with subjects performing a simple lexical task alternating with rest periods. The authors removed the “task” blocks (100 sec each) and concatenated the “rest” blocks (32.5–42.5 sec each) to create continuous resting-state data, which was compared with a true resting-state task. Using a seed-based analysis, they identified few regions as differentially active between the two data sets, and concluded that it is unlikely that functional connectivity of the resting blocks is considerably changed by intermittent task performance. Even less work has been conducted looking at the impact of a motor task on the resting state. Using independent components analysis to compare a block-design bilateral finger tapping activation task (with 32 sec movement and rest blocks) with a resting-state task, Arfanakis et al. (2000) concluded that there was no significant difference between functional connectivity during the task and during the resting state in task-independent cortical regions. Peltier et al. (2005), however, demonstrated a significant decrease in the connectivity within the motor cortices during the resting state after 20 min of repetitive right hand gripping. There remains a paucity of studies on the potential modulation of the resting state by motor task performance and a lack of congruency in the findings to date.
Our hypothesis is that there is no difference in RSFC between a resting-state paradigm in which no overt task is performed and the residual time courses from a block-design motor task after regressing out the task paradigm. If equivalence of RSFC is demonstrated between resting-state and motor paradigms, it would create an opportunity for RSFC analysis to be performed on virtually all previously collected movement task data without the need for new acquisition. Clinical utility of fMRI has been limited to preoperative studies in individual patients to guide neurosurgical intervention. The development of RSFC analysis offers tremendous promise for improving the clinical applicability of fMRI with group differences in RSFC patterns observed in a variety of neuropsychological and neurodegenerative disorders (Auer, 2008; Fox and Greicius, 2010). RSFC analysis on block-design fMRI data could, therefore, lead to broader application of fMRI in the clinical realm by identifying how cortical areas work with one another, and providing insight into the pathology underlying various neurological disorders.
Materials and Methods
Subjects
Sixteen healthy subjects (eight male; 40.6 ± 13.4 years old) with no history of neurological or psychiatric disorder participated in the study. All subjects gave informed written consent to procedures approved by the University Health Network Research Ethics Board.
Data acquisition
All data were obtained on a 3T Signa HDxt MRI system (GE Medical Systems, Milwaukee, WI) using an eight-channel phased array head coil. Data were acquired using T2*-weighted echo planar imaging with the following imaging parameters: repetition time (TR) = 2 sec, echo time (TE) = 30 msec, flip angle (FA) = 85°, field of view (FOV) = 22 cm; 64 × 64 matrix with 3.44 × 3.44 mm2 in plane resolution; 28 transaxial slices with 4 mm thickness; and number of volumes = 154. Each functional session lasted 5 min and 8 sec. For each subject, a 7 min T1-weighted 3D spoiled gradient echo sequence was performed to obtain an anatomical reference (TR = 7.8 msec, TE = 2.9 msec, FA = 20°, FOV = 22 cm, 256 × 256 matrix that resulted in a voxel resolution of 0.859 × 0.859 × 1.5 mm). To prevent large head movements, the subject's head was fixed by placing foam pads on each side of the head within the head cage.
In the first experiment, subjects were instructed to keep their eyes closed, remain alert, and not to think of anything in particular. This was the specific resting-state paradigm (RS1). The second experiment was a standard block-design motor task paradigm (“RH15”) whereby each subject performed a simple, sequential, and repetitive pinching movement of each digit against the thumb (digit 2, 3, 4, 5, 2, 3, 4, 5, etc.) using the right hand at ∼1–2 Hz for 15 sec, then performed no task for 15 sec. Subjects were instructed to begin the movement period of the experiment by the voice command “Go” over the speaker system in the scanner and were similarly instructed to stop movement with the command “Stop.” The instructions given to each subject during the 15-sec period during which no task was performed were to lie still in the scanner, remain alert, and not to think of anything in particular. The third experiment was a similar standard block-design motor task paradigm (“RH30”) but the task was performed for 30 sec, alternating with 30 sec of rest. The fourth experiment was a similar standard block-design motor task paradigm (“BH30”) whereby the pinching movement was performed with both hands for 30 sec, alternating with 30 sec of rest. The resting-state paradigms were then repeated two additional times (RS2 and RS3).
Data analysis
Preprocessing
Data were preprocessed with Analysis of Functional Neuroimaging (version 2009_12_31_1431) and FMRIB Software Library (FSL, version 4.1.5). The first three volumes (6 sec) of each functional data set were discarded from the analysis to permit stabilization of magnetic saturation effects. Standard slice time correction was employed. Motion correction was performed by volume registering each 3D volume to a base volume. Motion time courses were obtained by estimating the value for translation (mm) and rotation (°) for each of the 151 consecutive volumes. Maximum head motion was <1 mm in translation and <1° in rotation for all subjects. The T1 anatomical image was segmented into gray matter, white matter, and cerebrospinal fluid (CSF) maps using the SPM5 package (Wellcome Institute of Cognitive Neurology, United Kingdom). The white matter probability map was thresholded at a probability of 0.5. This anatomic-resolution mask was overlaid on the coarser data-resolution sampling grid and only those data set voxels containing at least 50% white matter mask voxels were labeled white matter data voxels. This white matter data mask was then eroded such that only voxels with six nearest neighbor white matter voxels were retained. This procedure was repeated for the CSF probability map. The resulting white matter and CSF data masks should only contain voxels with no partial voluming.
Following the CompCorr method of Behzadi et al. (2007), a principal components analysis was performed on the time courses of all white matter and CSF voxels within these masks. The top five principal components for white matter and the top five for CSF were entered into a standard multiple regression analysis of the time course of each gray matter voxel, along with the six motion time courses. This procedure aims to remove as many nonneuronal sources of signal fluctuations as possible, such as cardiac, respiratory, and scanner variability. For each motor-task paradigm, the corresponding block design was convolved with a standard hemodynamic response function and added to the regression to additionally remove the effects of the task. The residual time courses were temporally band-pass filtered (0.01–0.1 Hz) and detrended to reduce low frequency noise (including slow scanner drifts) and influences of higher frequencies reflecting possible cardiac or respiratory oscillations (Greicius et al., 2003; Lowe et al., 1998). Both the true resting-state and the “pseudoresting-state data”—created by regressing out the usual model of the motor-task response from each of the task data sets—were entered into a standard seed-based connectivity analysis procedure.
The filtered residual time course data sets were first spatially transformed into Montreal Neurological Institute (MNI) space using parameters estimated during the anatomic segmentation procedure. Average time courses from spheres (radius 12 mm) centered on the following MNI coordinates were used in voxel-wise regression analysis to define the following functional brain networks: the PCC [0–42 39] defined the DMN, the right/left dorsolateral prefrontal cortex [±42 33 21] defined the executive control network (ECN), with auditory, visual, and sensorimotor (MOT) networks defined, respectively, by primary auditory cortex [±40–22 8], primary visual cortex [0–84 8], and primary motor cortex [±34–18 51]. Separate analyses were conducted using right and left seeds. For each data set, a spatial map was created consisting of correlation values calculated between the given seed time course and each voxel time course, after conversion into Fisher z values.
Spatial smoothing was applied to these maps using a Gaussian kernel with FWHM = 9 mm. For each resting-state network, comparisons were made between each of the pseudoresting-state connectivity maps (i.e., from task data sets RH15, RH30, and BH30) and the true resting-state connectivity map, using a simple paired t-test over subjects as random effects. FSL's threshold-free cluster enhancement was used to threshold differences between true and pseudoresting-state maps at map-wise (FWE) corrected pFWE <0.05.
Results
Resting-state paradigm
In our group of healthy subjects, all components of all of the studied brain networks were engaged in all 16 subjects during each resting-state experiment (mean DMN over all subjects is shown in Fig. 1). There were no areas of significant difference between mean DMN map across all subjects for RS1, RS2, and RS3.
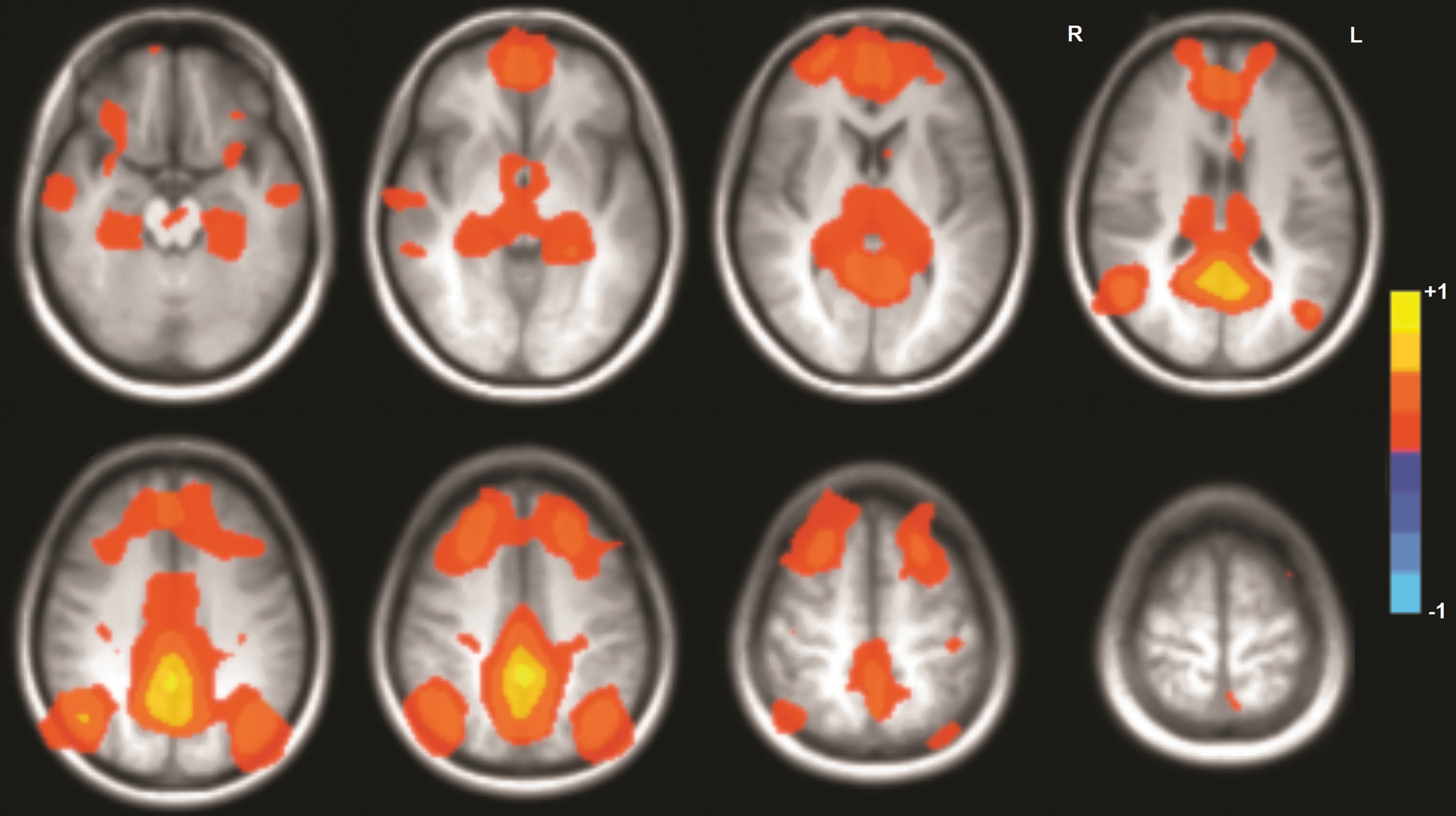
Mean default mode network activation across all 16 subjects during the resting-state paradigm. All expected areas were activated in each subject. Color images available online at
Movement paradigms
In our group of healthy subjects, all components of all of the studied brain networks were engaged in all 16 subjects during the pseudoresting state derived from each of the motor tasks (mean DMN over all subjects is shown in Fig. 2).
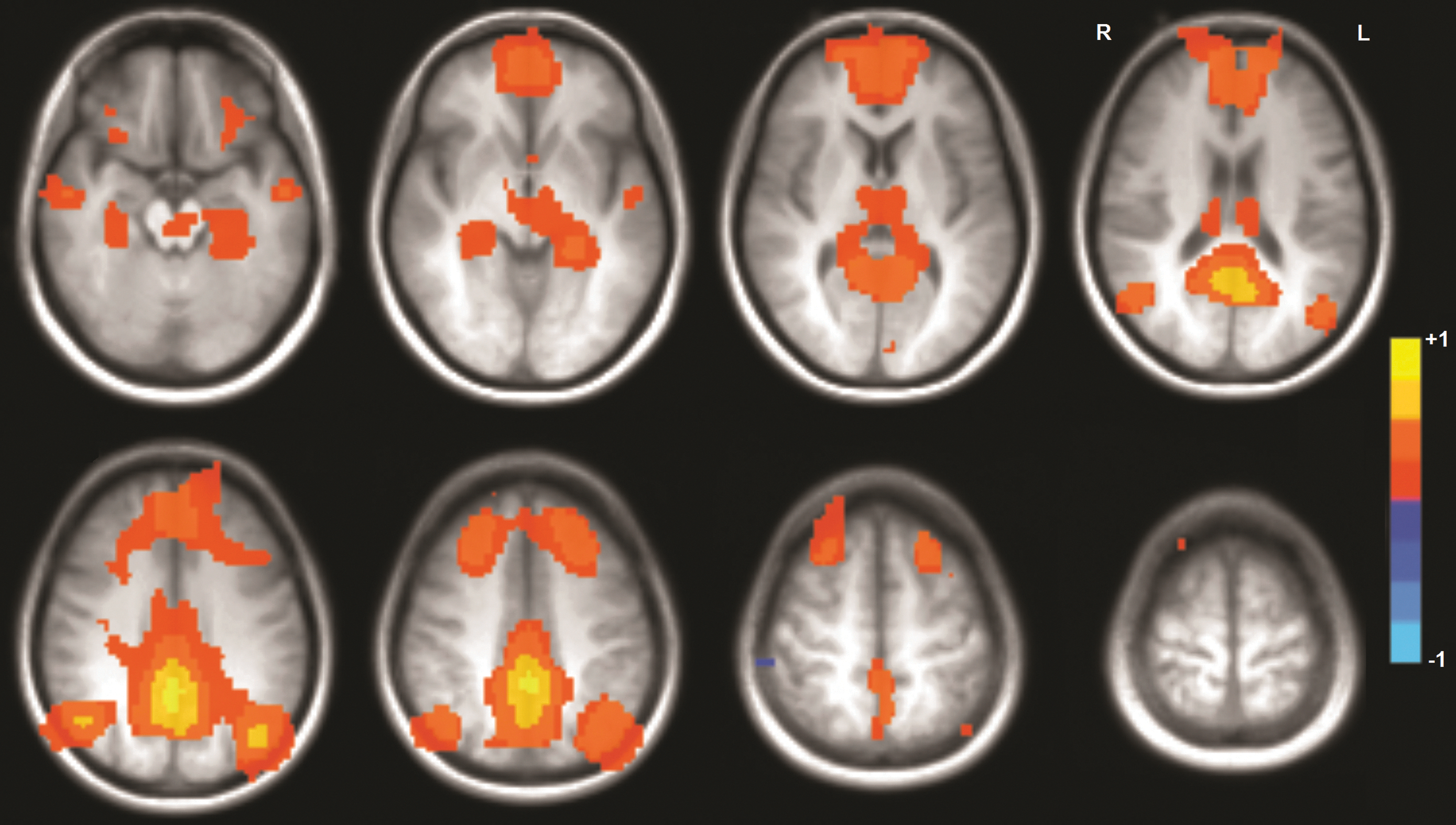
Mean default mode network activation across all 16 subjects during the pseudoresting state extracted from the RH30 paradigm. All expected areas were activated in each subject. Color images available online at
Resting-state versus movement paradigms
Seed connectivity analysis
For all three motor tasks, functional connectivity during the true resting state was greater than pseudoresting-state connectivity in all but one area identified as significantly different. For seeds located in the left hemisphere, differences were isolated to the sensorimotor cortex, mainly on the right (Fig. 3, Table 1). For only one task was there a significant difference between the resting-state and pseudoresting state in the ECN, and it was for the bilateral hand movement task, on the bilateral inferior frontal gyri and left superior temporal gyrus (Fig. 4, Table 1). For seeds located in the right hemisphere, significant differences were only observed between the resting state and bilateral hand movement pseudoresting state, in the bilateral sensorimotor cortex and bilateral cuneus, with the pseudoresting state greater than the resting state on the left precentral gyrus (Fig. 3; Table 1). For only one task was there a significant difference between the resting state and pseudoresting state in the ECN, and it was for the RH30 task, on the left superior frontal gyrus (Fig. 4, Table 1).
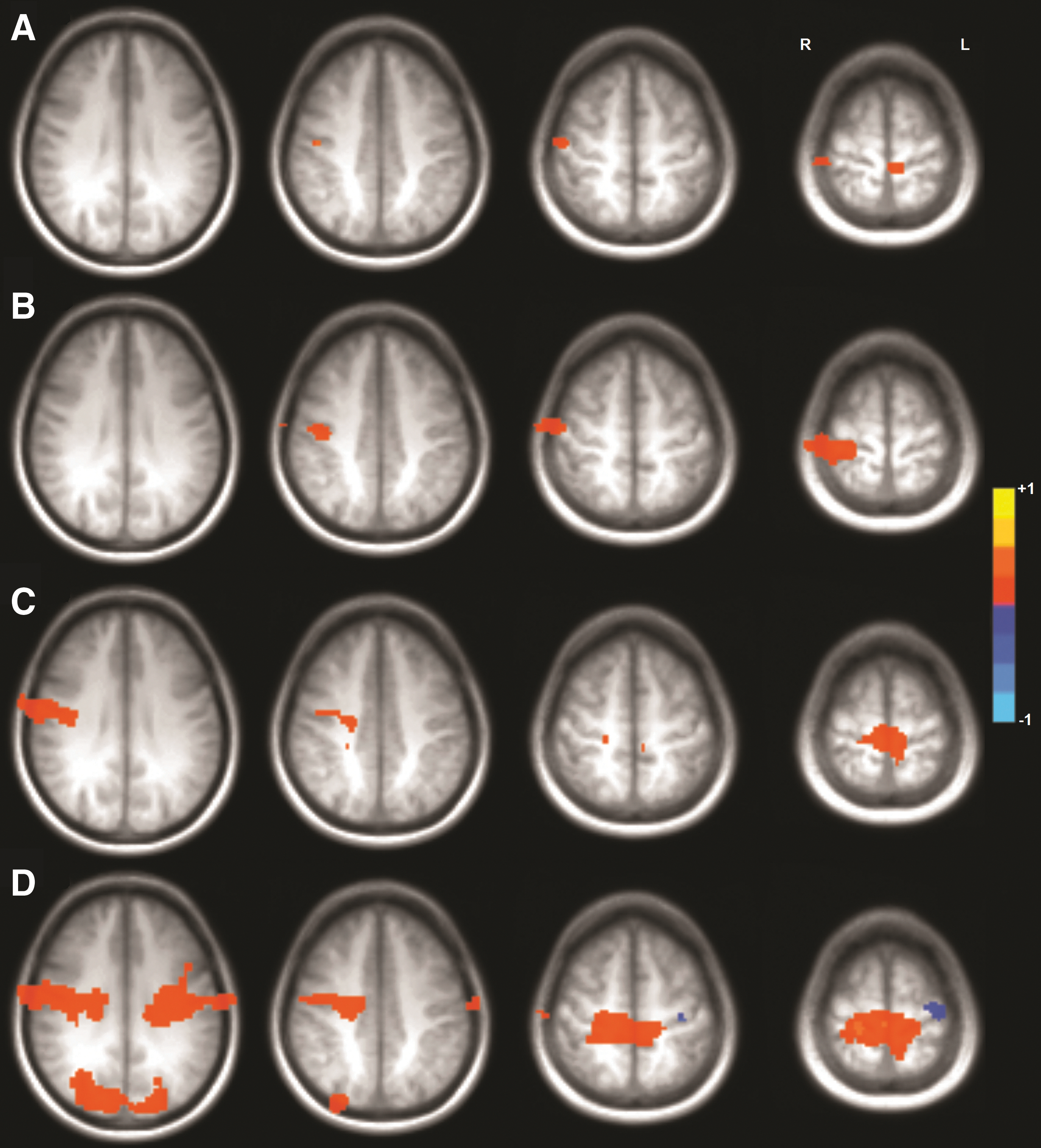
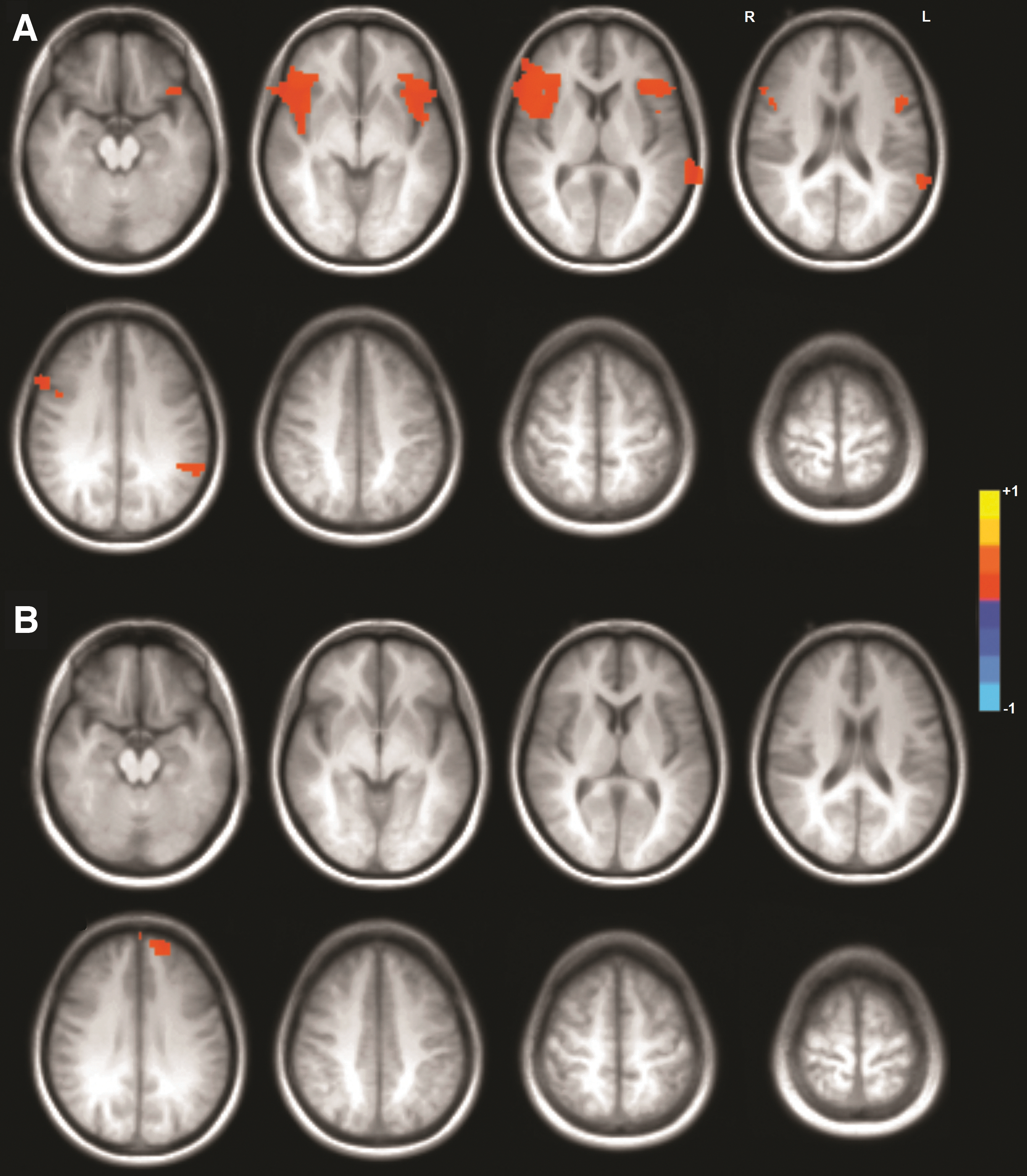
Clusters of Activation Significantly Different Between the Resting State and Pseudoresting State
FSL threshold-free cluster enhancement p < 0.05, clusters > two original voxel volumes.
Discussion
This study demonstrates that there is little difference in RSFC, in nonmotor task-related cortical areas, in healthy subjects as measured by BOLD fMRI during a specific resting-state paradigm and during simple block-design movement tasks. Furthermore, we found that the resting state extracted from dominant hand motor paradigms more closely resembled the actual resting state compared with that extracted from the bilateral motor paradigm.
Whether the connectivity map revealed by resting-state covariance analysis is confounded by specific constraints of a task is not well established as the work to date has been contradictory. In a study similar to ours, Arfanakis et al. (2000) used both a resting-state scan, where subjects performed no active task, and a block-design bilateral finger tapping activation task, in which they used independent components analysis to remove movement-related activation from the motor task data set to create activation-free pixel time courses. Although they limited their analysis to regions of the brain to nonmotor areas, they concluded there was no significant difference in functional connectivity during the task and during the resting state in the task-independent cortical regions. Their findings were, therefore, similar to ours in that we found no differences in the RSFC during the resting state and the pseudoresting state extracted from the motor paradigm across the DMN, the visual network, or the auditory network, and in only a few limited areas across the ECN. Our findings in motor areas of the brain for the true resting state were also congruent with prior studies (Biswal et al., 1995; Xiong et al., 1999). As in the current work, these prior studies showed functional connectivity in the bilateral primary motor cortices even during a resting-state task, although no comparison was made with an extracted resting state from the motor tasks they had their subjects perform. The regression step involved in creating the pseudoresting state could potentially remove network fluctuations but only at the precise fundamental frequency (and harmonics) of the motor task data sets, which, however, we would expect to affect all networks. That the motor network shows the greatest difference indicates to us that the motor task itself is influencing the amount of network fluctuations occurring during either the baseline of task block or both.
Interestingly, pseudo-RSFC was significantly lower than true RSFC in the right primary motor cortex during all three tasks—a movement-dependent cortical region. Peltier et al. (2005) studied the resting state before and following a repetitive unilateral hand grip task designed to induce fatigue. Their results suggest that interhemispheric functional connectivity between the motor cortices is reduced after the task, hypothesized to be a result of a temporary “disrupted state” either as a result or the cause of muscle fatigue. It is unlikely that fatigue contributed to the decreased connectivity in our subjects with the simple finger-tapping task we employed. Nonetheless, our results suggest that the right motor cortex is taken offline during a baseline state and engaged during simple self-directed unilateral and bilateral hand movements. This could represent enhanced ipsilateral motor cortical involvement for the right hand component of the motor task. It is well reported that the ipsilateral motor cortex is involved in simple unilateral hand movements, more so during nondominant hand movement. Increased unilateral hand movement complexity (van den Berg et al., 2011), force (Derosiere et al., 2014), and speed (Hayashi et al., 2008) affect the degree to which the ipsilateral motor cortex is involved. The impact of a synchronous bilateral hand motor task confounds the picture, as this movement would be expected to engage the motor cortex of interest (i.e., right motor cortex is ipsilateral during right hand movement, but is contralateral during left hand movement and is intimately involved in the control of left hand movement). As such, the finding of lower pseudo-RSFC in the right sensorimotor cortex during the bilateral finger-tapping task could alternatively represent increased contralateral sensorimotor cortical involvement for the left hand component of the bilateral motor task. Increased unilateral hand movement complexity (van den Berg et al., 2011), force (Derosiere et al., 2014), and speed (Hayashi et al., 2008) also affect the degree to which the contralateral motor cortex is involved, and again, the impact of a synchronous contralateral hand motor task confounds the picture. The movement employed in this study was performed at a slow speed (1–2 Hz) and at a low force (a light touch between opposing fingers), minimizing the effects of these parameters on cortical activation. The complexity of the movement, however, may have contributed to this finding as the sequential finger tap task employed, although relatively simple for the dominant right hand, may have been perceived as more complex for the nondominant left hand, engaging the right motor cortex to a degree, thus forcing these neurons from their baseline state, resulting in a lower pseudo-RSFC in this area. Yet another consideration is whether the resting state is really turned down when initiating the motor task. As it takes time for the movement to become “automated” and reach a point wherein one does not have to think about task performance, at which time the full resting state may become engaged—but this has yet to be determined.
The clinical significance of this work lies in the potential use of RSFC analysis on previously collected block-design motor task paradigm BOLD fMRI data. The finding that there is very little difference in RSFC, in nonmotor task-related areas, between specific resting state paradigms and certain motor task paradigms means that retrospective analysis of previously collected block-design motor task paradigm BOLD fMRI data can be performed to identify the resting state in each of these subjects. Despite its success and popularity as a research tool, fMRI has seen relatively little translation into the clinical realm. In general, clinical utility has been limited to preoperative fMRI in individual patients for guiding neurosurgical intervention. fMRI abnormalities seen in clinical research populations have not yet translated into diagnostic or prognostic tools for examining individual patients (Matthews et al., 2006). The development of RSFC analysis offers tremendous promise for extending the clinical applicability of fMRI into the realm of neurpsychological disorders—as group differences in RSFC patterns have been observed in a variety of neuropsychological disorders (Auer, 2008; Fox and Greicius, 2010)—and beyond. RSFC analysis will continue to advance fMRI research by providing broader knowledge of how cortical areas work with one another and insight into the pathology underlying various neurological disorders, and may eventually have significant impact through provision of useful diagnostic and prognostic information. This study shows that much could still be learned about the resting state using simple motor task fMRI data that have already been collected.
Conclusion
There is little difference in functional connectivity in nonmotor task-related cortical areas during a resting-state paradigm, in which no overt task is performed and the residual time courses from a simple, self-paced block-design motor task after regressing out the task paradigm. RSFC analysis can, therefore, be performed on virtually all previously acquired fMRI data incorporating these motor tasks without the need for new acquisition. Retrospective RSFC analysis on the fMRI motor task data could, therefore, lead to broader application of fMRI in the clinical realm by identifying how cortical areas work with one another and providing insight into the pathology underlying various neurological disorders.
Footnotes
Acknowledgment
Funding for this work was provided by a Toshiba America Medical Systems/RSNA Research Resident Grant No. RR1224.
Author Disclosure Statement
No competing financial interests exist.