
Abstract
Transcutaneous spinal direct current stimulation (tsDCS) is a noninvasive method that can modulate spinal reflexes, sensory afferent conduction, and even pain perception. Although neurophysiological evidence suggests that tsDCS alters somatosensory and nociceptive afferent conduction to the cortex, its supraspinal effects have not yet been investigated by using functional imaging to investigate tsDCS-induced alterations in intrinsic functional connectivity (FC). Therefore, we hypothesize that tsDCS-induced changes in neurophysiological measures might also be reflected in spontaneous brain activity. We investigated tsDCS-induced changes in somatosensory cortical connectivity by using seed-to-voxel-based analyses from the bilateral primary somatosensory cortex (S1) and the thalamus in a double-blind, crossover study design. Resting state FC was measured by using blood oxygenation level-dependent, functional magnetic resonance imaging (3T Philips) before and after anodal, cathodal, and sham tsDCS (20 min, 2.5 mA, active electrode centered over T11 spinous process, reference electrode over left shoulder blade) in a double-blind, crossover study of 20 healthy men (24 ± 0.7 years). As compared with sham, anodal tsDCS resulted in a decreased connectivity between the S1 and the ipsilateral posterior insula for both left and right hemispheres. Anodal tsDCS also resulted in decreased thalamic connectivity with the anterior cingulate cortex, and increased connectivity between S1 and the thalamus. Cathodal tsDCS showed increased FC between the right thalamus and both left and right posterior insulae, and decreased connectivity between the S1 seeds and the occipital cortex. Our results provide evidence of supraspinal effects of tsDCS and suggest that tsDCS may provide a noninvasive intervention that is able to target cortical sensory networks.
Introduction
T
In addition, stimulation effects are not limited to elicit responses. In rats, for example, the spontaneous activity in the nucleus gracilis as well as the primary somatosensory cortex (S1) was altered both during and immediately after tsDCS (Aguilar et al., 2011), suggesting not only that tsDCS effects extend to supraspinal structures but also that they influence task-independent activity. Direct current stimulation (DCS) applied to other areas have also shown effects on resting-state functional connectivity (FC) that extend past the site of stimulation. This has been shown by using the parent application of tsDCS, transcranial DCS (tDCS), where DCS is applied over the head. When applied over the primary motor cortex (M1), for example, anodal tDCS has been shown to affect not only intrinsic local connectivity within the M1 (Polanía et al., 2012a) but also connectivity between remote cortico-thalamic circuits (Polanía et al., 2012b). TDCS-induced influences on frontal connectivity have further been linked to effects on improved cognition (Yu et al., 2015). Therefore, it follows that tsDCS may similarly have consequences on intrinsic functional circuits that are located distant from the site of stimulation. The extent of the remote supraspinal effects associated with tsDCS, however, remains unclear.
To this end, we investigated FC changes throughout the cortex associated with tsDCS application. In particular, resting-state functional magnetic resonance imaging (fMRI) was used to identify tsDCS-induced changes in intrinsic FC patterns. Resting-state FC has been shown to resemble patterns of activity known from task-evoked blood oxygenation level-dependent (BOLD) fMRI (Biswal et al., 1995; Fox et al., 2005; Raichle et al., 2001) and to have direct links to behavior, perception, and learning (van den Heuvel and Hulshoff Pol, 2010), emphasizing a link between resting-brain activity and behavior. Since resting-state measurements are not constrained by any one task, they allow for a simultaneous investigation of multiple functional networks. This seems to be most appropriate for tsDCS, as effects have been shown in multiple modalities, for example, motor (Bocci et al., 2015a, 2015c), somatosensory (Cogiamanian et al., 2008), nociceptive (Cogiamanian et al., 2011; Truini et al., 2011), and visual (Bocci et al., 2015b) modalities. When also considering the tsDCS-induced changes in spontaneous activity in the S1 in rodents (Aguilar et al., 2011), it seems that tsDCS has broad effects on task-evoked responses, but that these effects may also extend to intrinsic activity, that is, in the absence of a task. Therefore, we hypothesize that tsDCS-induced changes in neurophysiological measures might also be reflected in spontaneous brain activity in the associated networks. Here, we specifically investigate tsDCS-induced changes in somatosensory cortical connectivity by using seed-to-voxel-based analyses from the S1 and the thalamus in a double-blind, pseudo-randomized, crossover study design. Twenty healthy male subjects were scanned six times over three sessions. Changes (post–pre differences) in resting-state FC within anodal and/or cathodal stimulation sessions were compared with changes in connectivity within the sham stimulation session.
Materials and Methods
Participants
To maintain a homogeneous cohort in this initial study, 20 healthy and right-handed men (age ± SD, 24 ± 0.7 years) participated in this study in the first half of 2013. All participants were recruited via local advertising, were free of neurological symptoms, unmedicated, and provided written informed consent. This study was approved by the Ethics Commission of the Ruhr-University Bochum Germany (No. 4421-12, date of approval: 20/08/2012) and was conducted in accordance with the ethical standards of the Declaration of Helsinki. All subjects participated in three pseudo-randomized sessions that were spaced at least 24 h apart. All were blinded to the study protocol and the hypothesis of investigation.
Transcutaneous spinal direct current stimulation
TsDCS was generated by a battery-driven DC-StimulatorPlus (neuroConn, Germany) and applied to the participant via a pair of rubber electrodes (each 7.3 × 5.0 cm; 36.5 cm2; 2 mm thick). Ten 20 conductive paste (Weaver and Company) was used to minimize skin resistance (one layer 1 mm thick). Using a maximum current of 2.5 mA (current density 0.068 mA/cm2), we delivered a total charge of 61.2 mC/cm2. TsDCS was applied for 20 min with the “active” electrode centered over T11 (T10–T12) spinous process and the reference electrode on the left shoulder with the right edge of the electrode aligned with the shoulder blade for consistent placement between sessions and for similar tissue properties between subjects. The stimulation was applied for 1170 s with an additional fade in of 10 s and a ramp out of 20 s (20 min of total stimulation). Polarity (anodal or cathodal) of tsDCS was referred to the electrode placed over T11. Sham tsDCS was delivered in cathodal montage with a current of 1.5 mA for 15 s with the same 10-s linear ramp up and 20-s linear ramp down as in anodal and cathodal polarities to mimic the initial tingling sensation while avoiding any stimulation-induced effects. In this way, subjects could not distinguish between the different stimulation procedures. A second investigator set the stimulation program so that the primary investigator was unaware of the stimulation applied during the session. Stimulation was pseudorandomized and balanced across and between subjects.
Stimulation was applied outside of the scanner immediately after the preresting state scan with the subjects in a seated position. Application within the scanner was avoided due to the suboptimal curvature of the back while lying in the supine position. For better control over electrode placement, we chose a seated position outside the scanner. After stimulation, subjects were immediately returned to the scanner for the postresting state scan (Fig. 1A). After each session, subjects completed a questionnaire to assess any pain associated with tsDCS as well as their guess as to which polarity of tsDCS they had received. This was to control for systematic sensory differences associated with any one polarity.
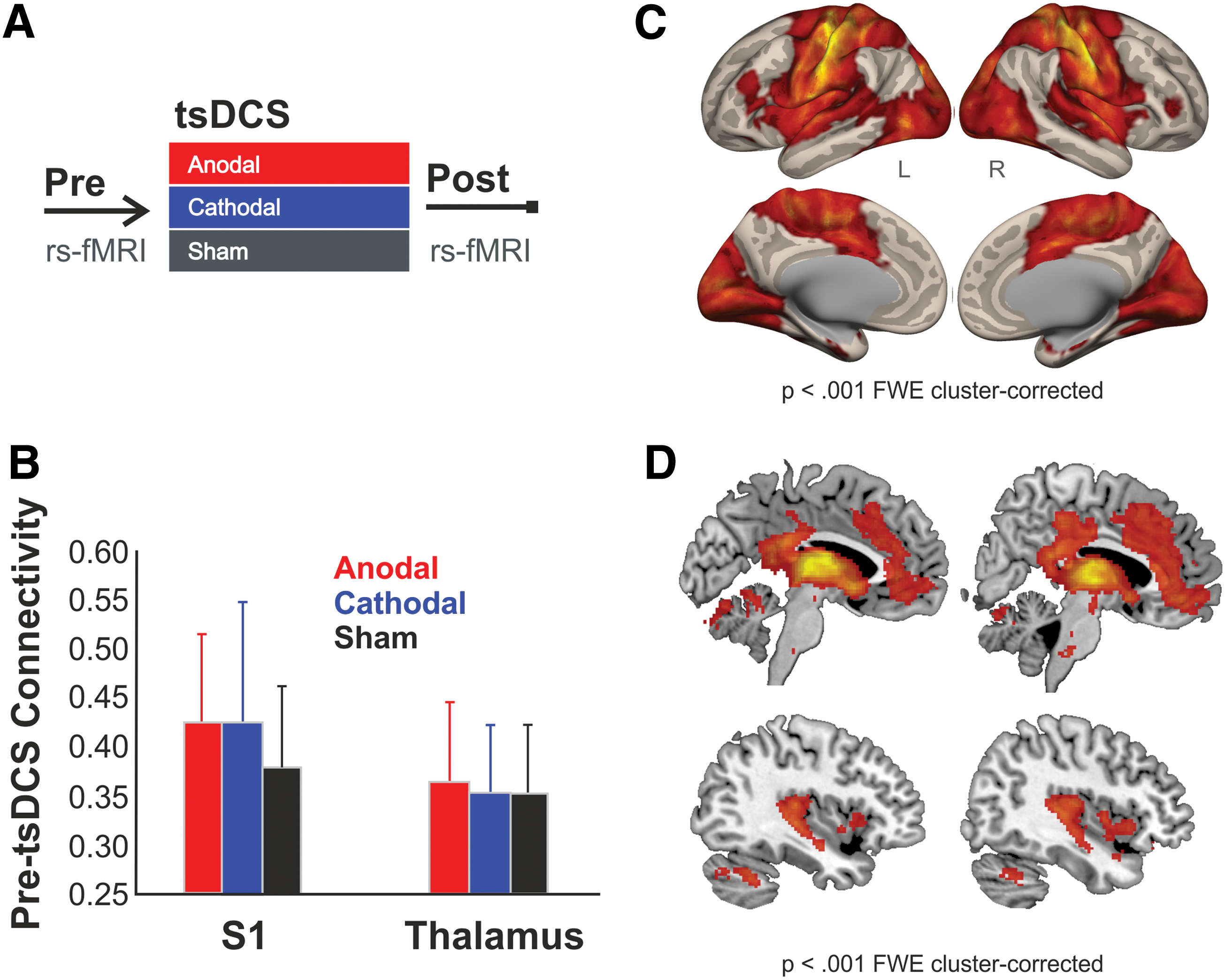
Pre-tsDCS S1 FC.
Image acquisition
Participants were scanned by using a Philips 3.0 T Achieva X scanner using a 32-channel head coil. Each session included a structural T1-weighted scan (MPRAGE, repetition time (TR)/ echo time (TE): 8.5/3.9 ms, voxel size (1 mm) three isotropic, field of view (FOV) 256 × 256 mm) followed by a T2-weighted resting-state scan (Gradient-echo planar imaging, TR = 2500 TE = 35 ms, Flip angle = 90°, FOV: 224 × 232 mm, 39 axial slices, slice thickness = 3 mm, no gap, 200 dynamic scans, five dummy scans, total acquisition time: 8 min 37 sec) where subjects were asked to keep their eyes closed, to not think about anything in particular, and to avoid falling asleep. After the tsDCS period outside the scanner, a structural reference scan (3 min) and the postresting state scan (with the same instructions as in the prerest) were acquired.
Functional connectivity
Preprocessing of functional images included slice time correction, realignment, and normalization to the Montreal neurological institute template. Structural images were normalized to the statistical parametric mapping T1 template without further spatial smoothing to define tissue boundaries. BOLD signal fluctuations from cerebrospinal fluid (CSF), white matter (WM), and the six head motion parameters from realignment preprocessing were regressed out of each individual (scan specific) BOLD time course by using the anatomical CompCorr method (Behzadi et al., 2007) provided by the Conn Toolbox (Whitfield-Gabrieli and Nieto-Castanon, 2012), after which bandpass filtering (0.008–0.1 Hz) was performed. Repeated-measures analysis of variance was performed to compare changes between sessions, that is, change during the anodal or cathodal session compared with the change during the sham session. Peak voxel threshold was set at p < 0.05 false discovery rate (FDR)-cluster correction (correction for multiple comparisons) was considered significant and are reported here.
Seeds of interest
A total of four seeds of interest were selected: (a) the left thalamus (automated anatomical labeling [AAL] atlas), (b) the right thalamus (AAL atlas), (c) the left S1 (Brodmann area [BA]1, BA2, BA3 combined), and the right S1 (BA1, BA2, BA3 combined). The thalamus was selected due to its role as the primary somatosensory relay region to the cortex, and the S1 was selected for its role as the primary target for incoming somatosensory stimuli. Since we did not differentiate between functional associations of the S1 seed, BA1, BA2, and BA3 regions were combined. S1 ROIs were defined by using the Brodmann Atlas rather than using the AAL atlas due to the BA's more complete incorporation of the postcentral gyrus in this cohort's normalized brains. The AAL-defined thalamic ROIs incorporated all parts of the thalamus.
Results
All 20 subjects were included in the data analysis. No adverse effects from the tsDCS electrodes or fMRI scans were reported and no polarity was consistently guessed correctly, nor did any subject correctly guess all stimulation polarities. Baseline connectivity for the four seed regions (S1 and thalamus, each left and right) did not differ at baseline (i.e., pre-tsDCS) between sessions (Fig. 1B). S1 connectivity pre-tsDCS is shown in Figure 1C for the combined left and right S1 seeds and included S1, M1, insular, as well as visual cortex areas. Thalamic connectivity (combined left and right seeds) included insular, anterior cingulate, and posterior cingulate cortices (Fig. 1D).
Anodal and cathodal effects of tsDCS–interaction analyses
After anodal tsDCS, a decrease in FC of the left and right S1 seeds with a cluster in the ipsilateral insula was observed, as compared with sham tsDCS. The left S1 showed an increase in connectivity with a cluster in the thalamus, and the right S1 showed an increase in connectivity with a cluster in the right parahippocampal gyrus (Fig. 2A). Connectivity between the left thalamic seed and a cluster in the anterior cingulate cortex (ACC) was decreased and increased with a cluster in the right posterior lobe of the cerebellum (Fig. 2B). No changes in connectivity for the right thalamus were observed.
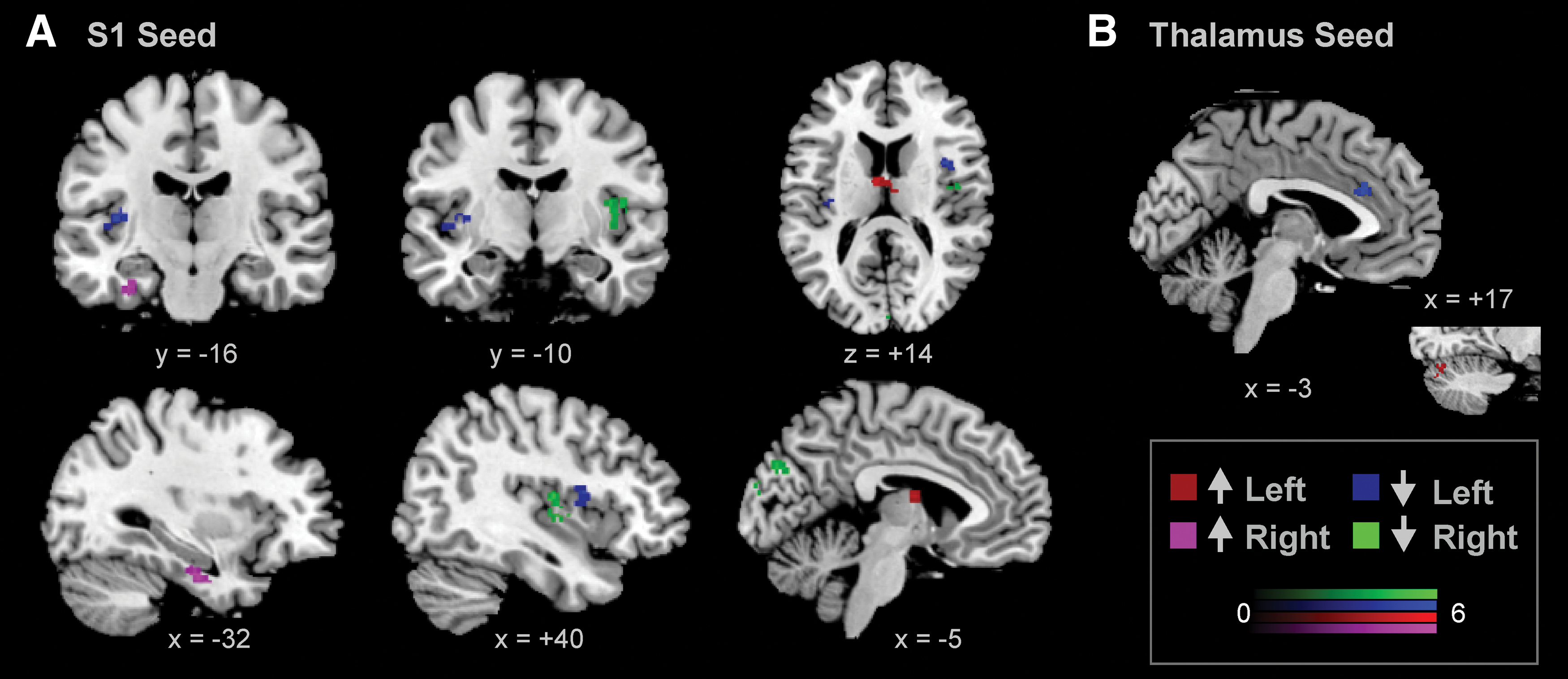
Altered FC after anodal tsDCS. Compared with sham tsDCS, anodal tsDCS decreases FC between
After cathodal tsDCS, S1 seeds (both left and right S1 seeds) showed decreased connectivity with a cluster in the occipital cortex, as compared with sham tsDCS (Fig. 3A). In addition, the left thalamus showed increased connectivity with a cluster in the left inferior frontal cortex/anterior insula and with a cluster in the left posterior insula (Fig. 3B). The right thalamus showed increased FC with clusters in the mid-posterior insulae bilaterally and with a cluster in the WM near the basal ganglia after cathodal tsDCS (Fig. 4). Detailed results of anodal and cathodal changes can be found in Table 1.
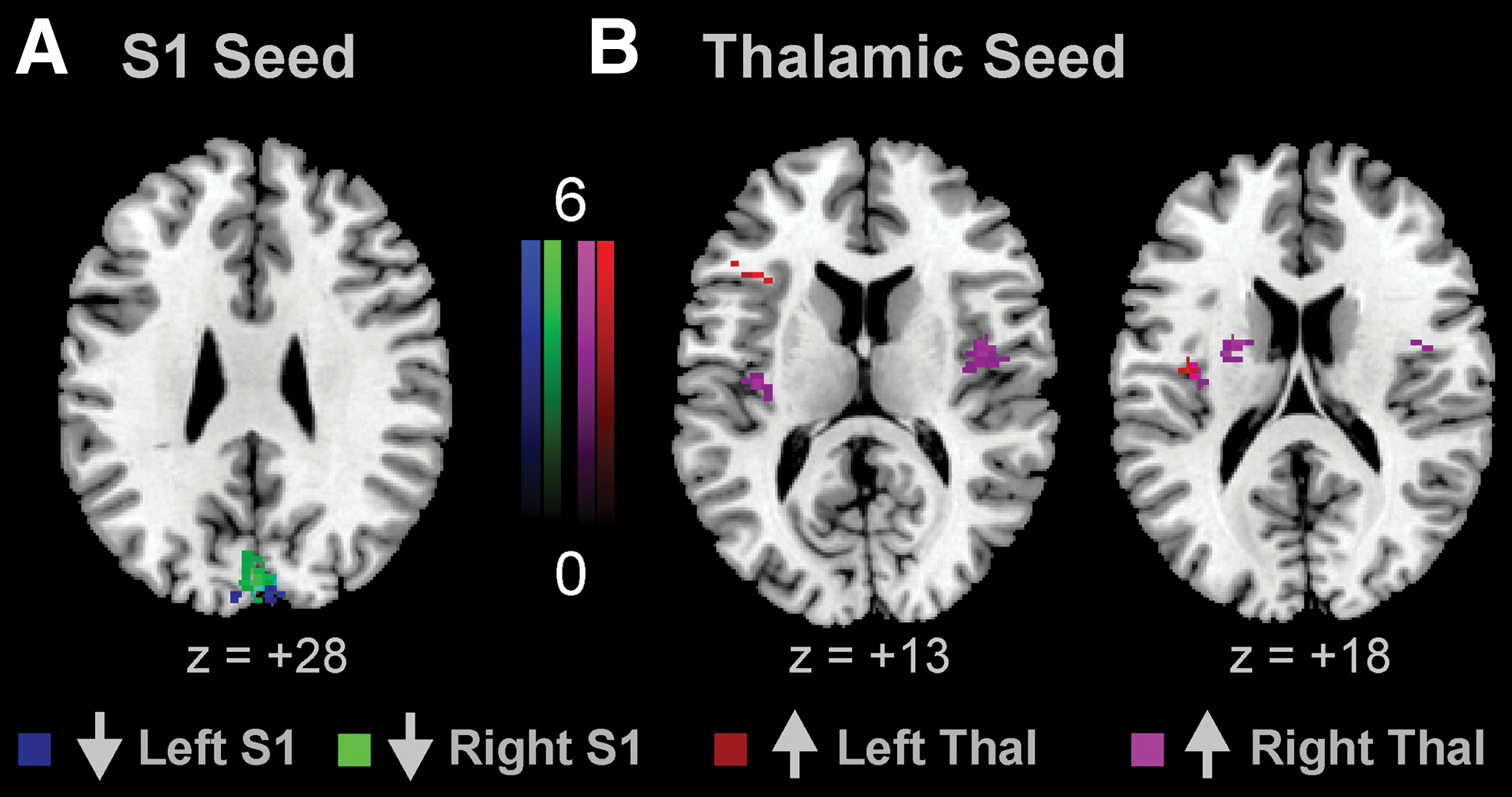
Changes in FC after cathodal tsDCS. Compared with sham, cathodal tsDCS decreases FC between both S1
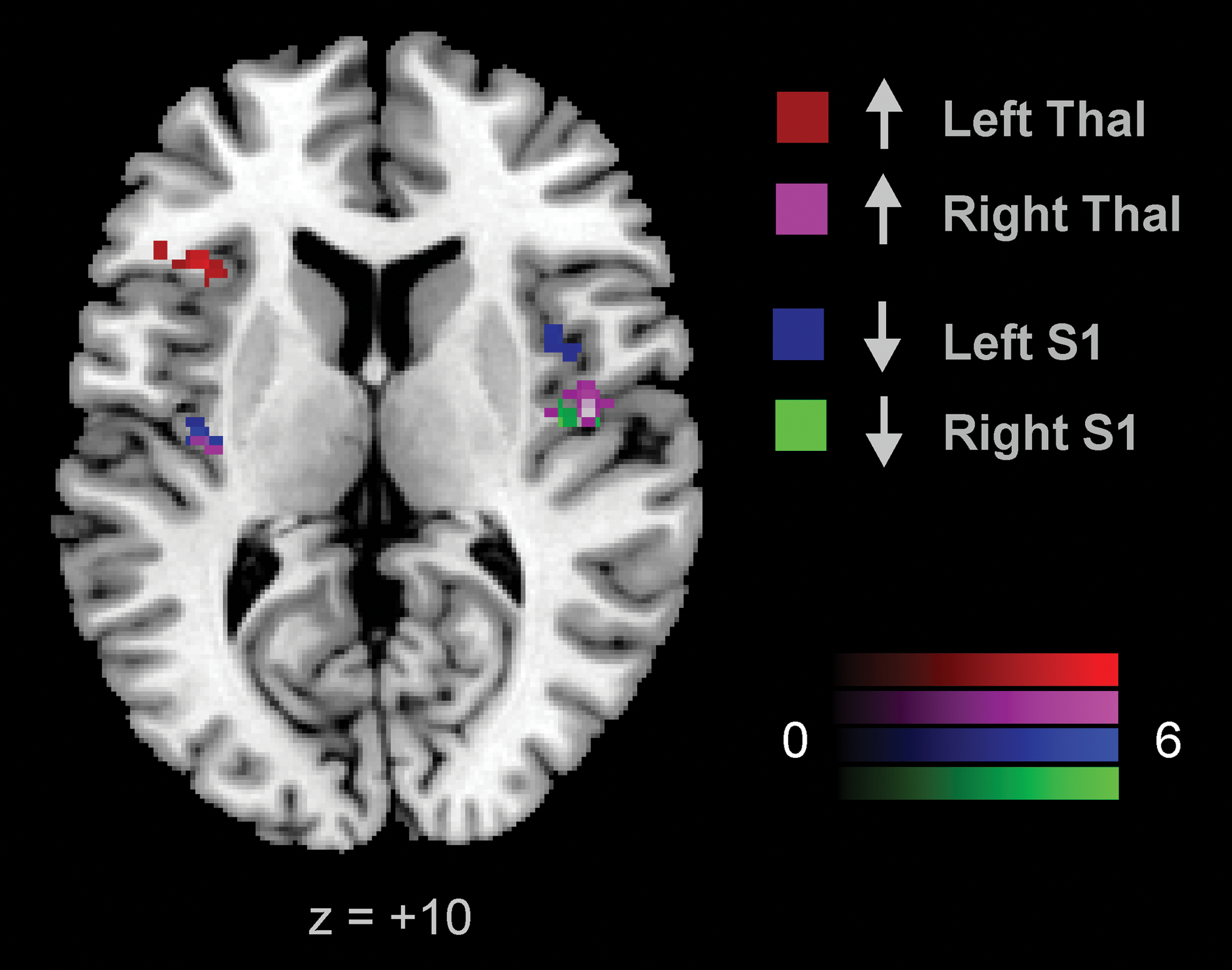
Overlapping insular areas. Decreases in connectivity with the insula after anodal tsDCS from the left (blue) and right (green) S1 seeds and increases after cathodal tsDCS from the left (red) and right (violet) thalamic seeds are shown. Voxels shown survive a p < 0.05 FDR-cluster correction (t-scores). Color images available online at
Seed-to-voxel connectivity changes using one of the four seed regions (see left column) are listed. Peak voxel coordinates, T-values, cluster sizes, and the direction of change (±, i.e., increased or decreased connectivity) denote the significant changes after either anodal (top portion) or cathodal (bottom portion) tsDCS compared with sham tsDCS.
AAL, automated anatomical lab; ACC, anterior cingulate cortex; L, left; R, right; S1, primary somatosensory cortex.
Discussion
This study investigated the supraspinal effects of tsDCS by using resting-state fMRI and could show that both anodal and cathodal tsDCS alter intrinsic FC within the somatomotor system. Compared with sham stimulation, anodal tsDCS reduced the FC bilaterally between S1 and the ipsilateral posterior insula whereas cathodal tsDCS strengthened the connectivity between the posterior insulae and the thalamus. These findings support previous evidence of cortical sensory effects measured after tsDCS.
TsDCS effects
Although a few studies have hypothesized a potential supraspinal effect of anodal tsDCS (Bocci et al., 2014; Cogiamanian et al., 2012; Heide et al., 2014), Truini and associates and Bocci and associates have been the only ones, to date, to report a change in a sensory cortical measure in human participants after anodal tsDCS, namely reduced LEP amplitudes and increased transcallosal time in the visual cortex, respectively (Bocci et al., 2015b; Truini et al., 2011). Similar to the two aforementioned studies, our results provide evidence for a supraspinal effect of tsDCS, specifically implicating sensory networks associated with S1 and multi-modal networks associated with the thalamus. After anodal stimulation, for example, our results identify that the FC between S1 and the ipsilateral posterior insula was reduced in both left and right hemispheres more so than after sham tsDCS. The same was true for the connectivity between the left thalamus and ipsilateral ACC. Interestingly, one of the LEP amplitudes that was shown to be suppressed was the N2 component (Truini et al., 2011), which reflects the activity in the ACC, secondary somatosensory cortex (S2, insula, and S1), as determined by intracortical recordings (Frot et al., 1999; Garcia-Larrea et al., 2003). Analogous to Truini et al. (2011), our data indicate the involvement of S1, insular, and ACC brain regions, but, in addition, suggest that the thalamus is implicated. Cathodal tsDCS effects targeted the insula, but rather than weakening insular-S1 connectivity, cathodal tsDCS strengthened insular-thalamic connectivity. The affected regions overlapped with those affected by anodal tsDCS (Fig. 4), suggesting that anodal and cathodal tsDCS have polarity-specific effects on insular connectivity; anodal decreases where cathodal increases insular connectivity.
The involvement of S1, insular, cingulate, and thalamic structures suggests either a multicircuit effect of tsDCS or possibly an effect on a functional network that involves all regions. All of these structures have been found to be consistently activated in response to painful stimuli (Apkarian et al., 2005, 2011; Peyron et al., 2000), forming a pain-related network (Melzack, 1999). Although these areas are not limited to a role in pain perception, their common involvement in pain and their common alterations after tsDCS suggest a potential for both anodal and cathodal tsDCS to affect pain-related networks. In other words, due to the presence of alterations in pain-related brain areas, tsDCS may be of interest to use to reduce (or simply alter) pain. In line with this, tsDCS has shown suppressive effects on pain-related behavioral measures, where anodal tsDCS increased cold pain tolerance (Truini et al., 2011), increased temporal summation thresholds (Perrotta et al., 2016), and reduced mechanical pain sensitivity (Meyer-Frießem et al., 2015), in addition to the aforementioned suppressive effects on LEP amplitudes evoked by painful heat stimuli (Truini et al., 2011). The insula has been previously shown to be affected by painful stimuli. For example, electrically induced pain has been previously shown to alter insular connectivity—in particular, posterior insular connectivity to the posterior cingulate cortex and precuneus (Vogt et al., 2016) and mechanical pinpricks have been shown by using task fMRI to evoke localized activation in the posterior insula (Baumgärtner et al., 2010). Since the posterior insula was also shown to be influenced by tsDCS in this study, it may be the case that insular connectivity changes, at least after anodal tsDCS, may be an underlying mechanism of tsDCS effects on pain and should be the focal point of further investigations.
As a behavioral correlate to these changes was not tested in the current study, it remains speculative that the FC changes reported here directly reflect the previously reported changes in pain measures. Nonetheless, the involvement of the insula, S1, thalamus, and ACC, all regions involved in pain processing, suggests that the resting connectivity within the pain network is altered by tsDCS, and, in turn, that tsDCS effects on pain processing may be linked to these changes. Further testing with combined FC and behavioral measures will elucidate how interconnected the two tsDCS effects are.
S1 connectivity was also noted to be altered between the S1 and the left occipital cortex (see Figs. 2A and 3A). Currently, we can only speculate on potential mechanisms underlying the observed decrease in connectivity between the sensory cortices; however, we would assume that this is an effect mediated by the thalamus. The thalamus modulates cortical activity at rest; given that there are thalamo-insular and thalamo-cortical loops, with cathodal stimulation increasing thalamo-insular connectivity, we would assume that a relative decrease in thalamo-cortical connectivity (not directly observed in our data) is associated with a (pseudo-) desynchronization of cortico-cortical connectivity. However, due to the lack of parallel studies supporting any tsDCS effects, it remains unclear at this time what the consequences of or mechanistic explanation for this change may be.
Resting-state fMRI
It is important to highlight the fact that the supraspinal changes reported here were measured in the absence of any specific task. It is well known that the brain is constantly active, regardless of whether a predefined task is presented or not, and over the past decade it has become increasingly clear that the underlying activity not only affects the evoked activity (Mennes et al., 2010) but is also itself directly related to behavior. Behavioral links to resting-state activity have been shown by using various sensory (Baldassarre et al., 2012), motor (Fox et al., 2007), and cognitive tasks (Hao et al., 2013; Xu et al., 2014). For example, we could recently show that a higher intrinsic connectivity in the S1 hand region is related to higher two-point discrimination acuity (Haag et al., 2015a), and also that the intrinsic connectivity at the network level can be used to predict accuracy in a cognitive conflict task (Haag et al., 2015b). Thus, the consequences of tsDCS-induced changes in resting-state connectivity are likely to have behavioral correlates. Whether this directly relates to the magnitude of reported effects remains to be seen.
Mechanisms
Our data support a clear supraspinal effect of thoracially applied tsDCS of both anodal and cathodal polarities, suggesting that tsDCS effects extend past the level of DC application. These supraspinal changes are in line with previous evidence of tsDCS effects on parameters measured rostral to the level of stimulation. Cathodal tsDCS-induced facilitation in somatosensory afferent conduction, for example, was measured at the cervical level of the spinal cord (Cogiamanian et al., 2008), and at the level of the cortex, anodal tsDCS was shown to suppress the amplitude of laser-evoked potentials (Truini et al., 2011). Anodal tsDCS was also shown to alter the interhemispheric conduction of visual-evoked potentials (Bocci et al., 2015b), suggesting that tsDCS effects do not only extend beyond more rostral levels of the spinal cord, but some may also even be independent of spinal afferents.
A likely mechanistic explanation of more rostral effects is that they are secondary to plastic alterations occurring at the level of stimulation, as supported by data consistently showing tsDCS effects on spinal reflexes. Anodal tsDCS, for example, reduced the amplitude of the RIII component of the lower limb flexion reflex (Cogiamanian et al., 2011), which is associated with the nociceptive component of the reflex and is related to pain (Sandrini et al., 2005), and, in addition, affected the monosynaptic H-reflex (Heide et al., 2014; Lamy and Boakye, 2013; Lamy et al., 2012; Winkler et al., 2010), which is unaffected by cortical modulation due to its monosynaptic nature. It is important to note that although anodal effects either increase or suppress a parameter, it is not yet clear that the anodal application itself is purely excitatory in nature, as it has been shown to be when applied over M1 (Nitsche et al., 2008). Animal studies have, in addition, noted alterations in local neurotransmitter concentrations within the spinal cord at the level of stimulation (Ahmed and Wieraszko, 2012), further supporting a primary spinally driven mechanism of action.
Nonetheless, due to the dense reciprocal connections between the spinal cord and cortex, alterations at one level are difficult to delineate from changes at the other level. A more general effect of tsDCS, which is one that targets both spinal and supraspinal connections, has been shown by using motor-evoked potentials (Bocci et al., 2015c) and motor unit recruitment (Bocci et al., 2014) to assess the corticospinal system. Interestingly, the tsDCS-induced alterations corticospinal tract excitability (Bocci et al., 2015c) were further accompanied by an increased inhibitory GABAergic drive at the level of the cortex (Bocci et al., 2015a), suggesting that an altered corticospinal excitability could be via a nonspinal mechanism (Bocci et al., 2015a). Although the delineation of cortical from spinal effects remains challenging, it has become evident that tsDCS has effects on multiple levels, for example, at the local spinal and rostral spinal segment levels, as well as supraspinally. Whether this is primarily due to changes at the level of the spinal cord remains to be determined. Our results point to a reorganization at the level of the cortex, suggesting that at least sensory afferent conduction to the cortex may be received differently after tsDCS, due to changes in the underlying activity of the S1 cortex and thalamus. With improved models of the current density after application of tsDCS (Parazzini et al., 2014; Toshev et al., 2014), a better understanding of the fibers primarily targeted by stimulation can be acquired (Danner et al., 2011), which will allow for a deeper understanding of which cortical connections may be influenced. This may also allow for optimized electrode montages and stimulation protocols.
Limitations and outlook
A notable limitation of this study is the lack of behavioral measures. Although we encourage future studies to look at the relationship between cortical and behavioral measures, for the initial study, we consciously did not include these or other measures that could alter the cognitive state before or between scans (Waites et al., 2005). In this way, we maintained a rather simplistic study design consisting of six resting-state scans in the same 20 male participants on three different days, with the only intervention between the intra-session scans being 20 min of tsDCS applied in a double-blind manner. Since other groups have reported tsDCS effects immediately after stimulation offset, it was also important to identify the “immediate” cortical effects of tsDCS. It will be necessary in future studies to determine whether the cortical effects, in addition, relate to behavioral changes after tsDCS, and moreover, whether this also holds true for female participants, since they are more often affected by pain conditions than male participants (Fillingim et al., 2009).
Recently, the brain-derived neurotrophic factor val158met polymorphism was identified to be involved in tsDCS effectiveness in altering the motor component of the H-reflex (Lamy and Boakye, 2013). Likewise, it will be interesting to see whether other factors, for example, personality traits, regional neurochemical levels, etc., also contribute to tsDCS effectiveness on sensory changes. With a better understanding of factors denoting subject responsiveness to stimulation, we can better determine target populations (healthy and patient populations) that could most benefit from this noninvasive electrical stimulation method. Recent evidence in spinal cord injury patients and restless leg syndrome patients highlights tsDCS’ potential to improve motor recovery by using cathodal tsDCS (Hubli et al., 2013) and to reduce symptom severity by using anodal tsDCS (Heide et al., 2014). It is currently unclear as to what extent tsDCS can serve as a therapeutic intervention, but our results, in particular insular changes (Vogt et al., 2016), suggest that tsDCS holds potential to affect somatosensory and possibly pain domains and, therefore, might be useful in pain conditions. This should be the focal point of further investigations (e.g., Schweizer et al., 2017).
In addition, further investigations using varied electrode placement, stimulation patterns, and length of stimulation of tsDCS are necessary. Up to now, in the field of spinal stimulation, there has been a lack of systematic investigation of dose titration. According to tDCS over M1, after-effects may be inverted depending on the length of stimulation (13 min vs 26 min; Monte-Silva et al., 2013) and cathodal inhibition may even switch to excitation when stimulation intensity is doubled to 2 mA (Batsikadze et al., 2013).
Finally, FC between regions was assessed by using bidirectional correlations. As such, the direction of connectivity, that is, whether the activity of Region A influenced the activity of Region B or vice versa, remains unclear. It is even conceivable that the connectivity changes were driven by a third region not identified in the analyses here. More sophisticated approaches exploring effective connectivity as well as the relationship between functional and structural connectivity (Damoiseaux and Greicius, 2009) will help to overcome these methodological shortcomings in future studies.
Conclusion
In conclusion, we have focused on the cortical effects of tsDCS measured by intrinsic BOLD signal fluctuations. We find that anodal tsDCS decreased connectivity between S1 and the posterior insulae, whereas cathodal tsDCS increased the connectivity between the thalamus and the posterior insulae. Our results, therefore, contribute to ongoing evidence that tsDCS extends to supraspinal levels and encourage more widespread applications of this method in sensory and potentially pain domains.
Footnotes
Acknowledgments
M.T. (SFB 874 TP A1) and T.S.W. received funding from DFG (SFB 874 TP A8). L.S. received a stipend from the International Graduate School of Neuroscience, Ruhr-University Bochum (BoNeuroMed). The authors would also like to extend their personal gratitude to the PHILIPS Germany personnel for their ongoing support.
Author Disclosure Statement
No competing financial interests exist.