
Abstract
Previous functional magnetic resonance imaging (fMRI) investigations suggest that the intrinsically organized large-scale networks and the interaction between them might be crucial for cognitive activities. A triple network model, which consists of the default-mode network, salience network, and central-executive network, has been recently used to understand the connectivity patterns of the cognitively normal brains versus the brains with disorders. This model suggests that the salience network dynamically controls the default-mode and central-executive networks in healthy young individuals. However, the patterns of interactions have remained largely unknown in healthy aging or those with cognitive decline. In this study, we assess the patterns of interactions between the three networks using dynamical causal modeling in resting state fMRI data and compare them between subjects with normal cognition and mild cognitive impairment (MCI). In healthy elderly subjects, our analysis showed that the salience network, especially its dorsal subnetwork, modulates the interaction between the default-mode network and the central-executive network (Mann–Whitney U test; p < 0.05), which was consistent with the pattern of interaction reported in young adults. In contrast, this pattern of modulation by salience network was disrupted in MCI (p < 0.05). Furthermore, the degree of disruption in salience network control correlated significantly with lower overall cognitive performance measured by Montreal Cognitive Assessment (r = 0.295; p < 0.05). This study suggests that a disruption of the salience network control, especially the dorsal salience network, over other networks provides a neuronal basis for cognitive decline and may be a candidate neuroimaging biomarker of cognitive impairment.
Introduction
N
A triple network model, comprising of these two antagonistic networks and the third network called the salience network (Chand and Dhamala, 2016b; Seeley et al., 2007; Uddin, 2015), has been recently proposed to understand the connectivity patterns of cognitively normal brain and the altered patterns in cognitive impairments (Chand et al., 2017; Menon, 2011). In cognitively normal young brains, the salience network was found to play a role in the dynamic switching of antagonistic activity between the default-mode and central-executive networks (Chand and Dhamala, 2016a; Goulden et al., 2014; Sridharan et al., 2008). However, whether such dynamic switching retains in normal aging and/or alters in mild cognitive impairment (MCI) is unknown.
Recent studies further subdivide the salience network into the dorsal and ventral salience networks (Touroutoglou et al., 2012, 2016). Dorsal salience network is implicated to be important in attention and switching between cognitive resources, while the ventral system is suggested to be crucial during affections or emotions (Kurth et al., 2010; Touroutoglou et al., 2012, 2016). Whether the dorsal or ventral salience network subsystem controls over the default-mode and central-executive networks is largely unknown. Evaluating the network interactions in diseased or aging population could help test the hypothesis already mentioned and provide insights into the relationship between network interactions and cognitive functions. Despite a large body of literature evaluating single network activity that reported reduced default-mode network activity in MCI and Alzheimer's disease (Brier et al., 2014; Greicius and Kimmel, 2012; Greicius et al., 2004), the patterns of interactions between the triple networks have remained largely underinvestigated in normal healthy aging and MCI. Studying network interactions may provide insight into the neural mechanism in the progression of MCI to dementia or Alzheimer's disease (Hajjar et al., 2016; Iadecola, 2014; Oveisgharna and Hachinski, 2010).
Therefore, our objectives were to investigate the salience network control over the default-mode and central-executive networks in a group of patients with MCI using rsfMRI data in comparison to normal subjects and correlate the degree of disruption of this interaction with their global cognitive scores as measured by Montreal Cognitive Assessment (MoCA). As the salience network control is argued to be involved in maintaining task-related cognitive performance and/or resting state in cognitively healthy individuals (Menon, 2011; Sridharan et al., 2008), we hypothesized that this control will retain in normal cognitive aging, whereas the MCI group will demonstrate disruption in the control of salience network over the default-mode and central-executive networks and that the degree of disruption correlates with cognitive performance.
Materials and Methods
Participants
The study protocol was approved by the Institutional Review Board of Emory University, Atlanta, Georgia. The rsfMRI data from 54 MCI individuals were obtained. The MCI participants inclusion criteria were as follows: (1) age ≥55 years, (2) systolic blood pressure ≥140 mm Hg or diastolic blood pressure ≥90 mm Hg or receiving antihypertensive medications, and (3) executive MCI assessed by cognitive testing: MoCA ≤26, cognitive performance at the 10th percentile or below in trail-mark or stroop interference or verbal fluency or abstractions, and minimal functional limitation as reflected by the functional assessment questionnaire ≤7. The participants exclusion criteria were as follows: (1) systolic blood pressure >200 mm Hg or diastolic blood pressure >110 mm Hg, (2) renal disease or hyperkalemia, (3) active medical or psychiatric problems, (4) uncontrolled congestive heart failure (shortness of breath at rest or evidence of pulmonary edema on examination), (5) history of stroke in the past 3 years, (6) ineligibility for MRI (metal implants or cardiac pacemaker), (7) inability to complete cognitive test and MRI scan, (8) women of childbearing potential, and (9) diagnosis of dementia (self-reported or caregiver reported).
The MRI data from 20 normal participants were included based on the following criteria: age ≥50 years, MoCA ≥26, clinical dementia rating scale (CDR) = 0, and normal logical memory subscale defined as ≥11 for 16 or more years of education, ≥9 for 8–15 years of education, and ≥6 for <7 years of education. The exclusion criteria were history of stroke in the past 3 years, ineligibility for MRI (metal implants or cardiac pacemaker), inability to complete cognitive test and MRI scan, and diagnosis of dementia of any type Abnormal thyroid-stimulating hormone (>10) or B12 (<250). All subjects were scanned on the same MRI scanner using the same imaging sequences. A written informed consent was collected from each participant before data collection.
MRI acquisition
MRI data were acquired on a SIEMENS Trio 3-Tesla scanner available at Center for Systems Imaging (CSI) of Emory University, Atlanta, Georgia. Foam padding and ear forms were used to limit head motion and reduce scanner noise to the subjects. High-resolution three-dimensional anatomical images were acquired using T1-weighted magnetization-prepared rapid gradient echo (MP-RAGE) sagittal with repetition time (TR) = 2300 ms, echo time (TE) = 2.89 ms, inversion time = 800 ms, flip angle (FA) = 8, resolution = 256 × 256 matrix, slices = 176, thickness = 1 mm. Blood oxygenation level dependent images for rsfMRI were collected axially using an echo-planar imaging sequence with TR = 2500 ms, TE = 27 ms, FA = 90, field of view = 22 cm, resolution = 74 × 74 matrix, slices = 48, thickness = 3 mm, and bandwidth = 2598 Hz/Pixel. The subjects were instructed to hold still, keep their eyes open, and think nothing during the rsfMRI scan.
Image preprocessing
We preprocessed the MRI data using SPM12 (Wellcome Trust Centre for Neuroimaging, London, United Kingdom;
Independent component analysis
We used constraint independent component analysis (ICA) to extract temporal signal for subsequence dynamical causal modeling (DCM) analysis. ICA has been widely used to investigate the functional brain network activities (Damoiseaux et al., 2012). A spatially constrained ICA approach has been introduced (Lin et al., 2010), which overcomes the difficulties in identifying components of interest and in determining the optimum number of components of conventional ICA. Constrained ICA has been highlighted as a useful tool, particularly if one is interested in specific brain areas or network(s). We used the Group ICA of fMRI Toolbox (GIFT;
We used the templates of each network (default-mode, salience, and central-executive networks) reported in a previous study (Shirer et al., 2012). For the dorsal and ventral salience networks, the coordinates from previous studies (Chand and Dhamala, 2016a; Sridharan et al., 2008; Touroutoglou et al., 2012) were used in the MarsBaR package (
Dynamical causal modeling
We used DCM to evaluate the dynamic interaction between the three networks. DCM analysis (Friston et al., 2003) assesses the directed connectivity between different brain areas or networks. In DCM analysis, several hypothesized possible models are specified, and Bayesian model selection (BMS) is then used to infer the model that best fits the measured data (Stephan et al., 2010). Recent methodological advances in DCM facilitate the implementation of random effects (RFX) for model selection (Stephan et al., 2009), which allows the specification of nonlinear modulations to investigate how a brain area (or network) influences connection strengths between other brain areas (or networks; Li et al., 2011; Stephan et al., 2008) and make use of stochastic models (Daunizeau et al., 2012). The stochastic model accounts for noise more accurately and, therefore, allows the application of DCM to rsfMRI data (Daunizeau et al., 2012; Friston et al., 2014). We carried out DCM analysis using SPM12 (Wellcome Trust Centre for Neuroimaging).
We specified three models for each subject with fully connected intrinsic connections (Friston et al., 2003, 2014) between the networks (Fig. 3). Model 1 specified nonlinear modulation by the default-mode network on both reciprocal connections between the salience and central-executive networks. Model 2 specified nonlinear modulation by the salience network on both connections between the default-mode and central-executive networks. Finally, model 3 specified nonlinear modulation by the central executive on both connections between the default-mode and salience networks. In this study, we used nonlinear differential equation based stochastic DCM already described to estimate the models. For both MCI and control groups, we identified the model with the highest probability using both the fixed effects (FFX) and the RFX-BMS methods, respectively. Both methods provide complementary information, but differ on the underlying assumptions. The FFX-BMS assumes that the optimal model is homogeneous across subjects and uses the group log-evidence, which measures the balance between fit and complexity of models and quantifies the relative goodness of models. In contrast, the RFX-BMS accounts for heterogeneity of model structure across subjects. The RFX-BMS provides posterior model probability, which measures how likely a specific model generated the data of randomly selected subject, and exceedance probability, which measures how one model is more likely than any other model, in the group data (Stephan et al., 2010).
Statistical analysis
Baseline factors were compared between the normal control (NC) and MCI groups using Mann–Whitney U test. Pearson's correlation analysis was performed to assess the association between connectivity strength of network modulations with MoCA for each model in MCI subjects only. We limited this analysis to MCI because all subjects in the control subjects had normal MoCA. All data were analyzed using MATLAB (Natick, MA;
Results
Fifty-three MCI and 20 normal subjects were included in the analysis. Sample characteristics are provided in Table 1. In the MCI group, mean age was 66.9 years (standard deviation [SD] 8.1), 60% were African American, 52.8% were women, mean education was 15 years (SD 2.4), mean systolic blood pressure 150.7 mm of Hg (SD 21.3), and mean diastolic blood pressure 90.9 mm of Hg (SD 13.5). In normal group, mean age was 65.8 years (SD 8.8), 20% were African American, 70% were women, and eight had hypertension. The mean education was 16 years (SD 2.9), mean systolic blood pressure 128.8 mm of Hg (SD 23.1), and mean diastolic blood pressure 71.7 mm of Hg (SD 11.7). Figure 1 shows the results of the constrained ICA for the (Fig. 1A) default-mode network, (Fig. 1B) salience network, and (Fig. 1C) central-executive network components for MCI. Figure 2 displays the results of constrained ICA for normal group.

The t-value maps of
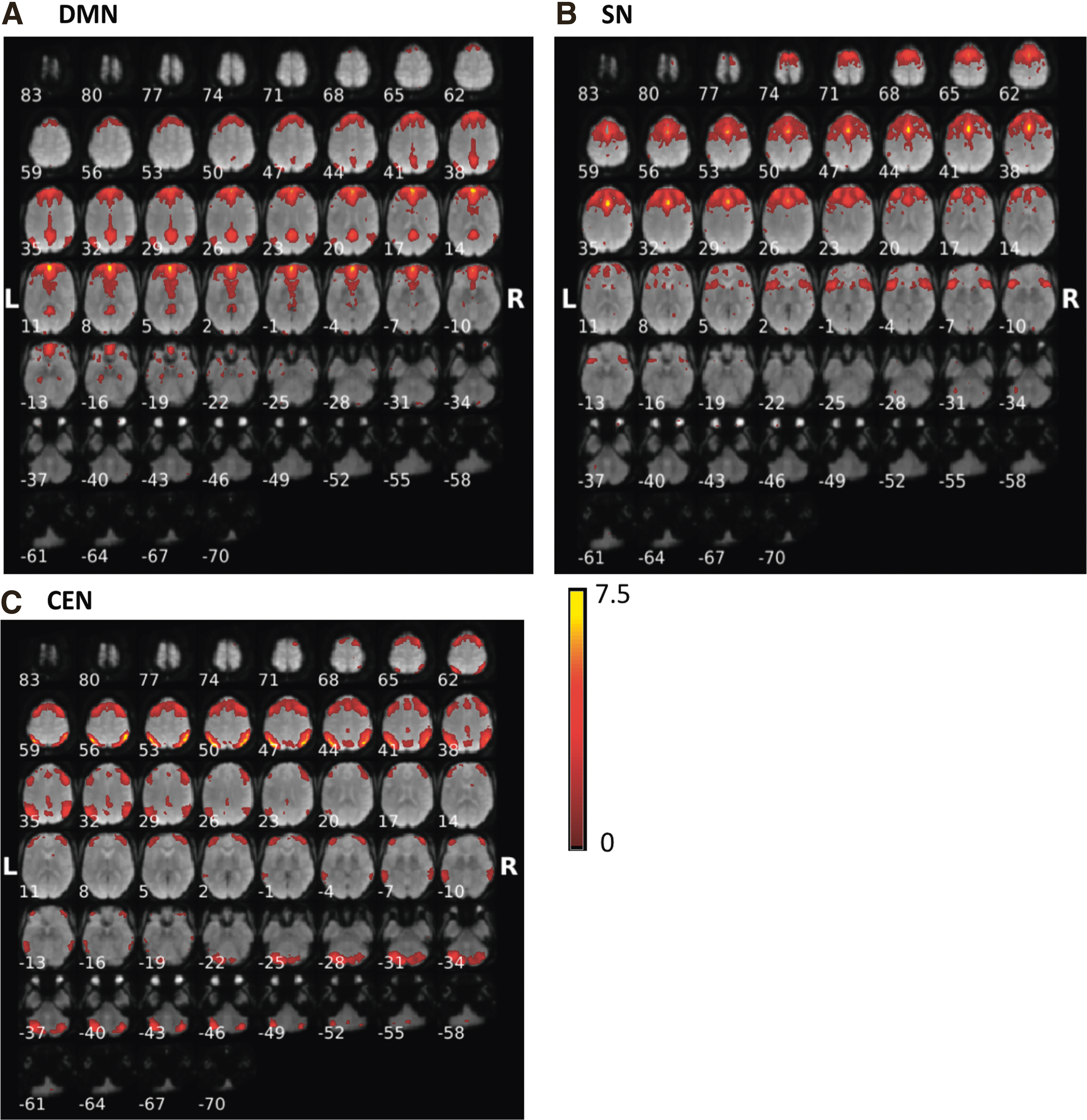
The t-value maps of
Number represents mean (SD) or count (%); p-value represents MCI versus NC using Mann–Whitney U test or chi-square test; and p < 0.05 is considered statistically significant.
BP, blood pressure; MCI, mild cognitive impairment; NC, normal control; SD, standard deviation.
Three network connection models were considered and compared statistically and are illustrated in Figure 3. For model 1 (Fig. 3A), the default-mode network extends modulations over the connections between the salience network and the central-executive network; for model 2 (Fig. 3B), the salience network extends modulations over the connections between the default-mode network and the central-executive network suggesting controlling/switching role of the salience network; for model 3 (Fig. 3C), the central-executive network extends modulations over the connection between the default-mode network and the salience network.
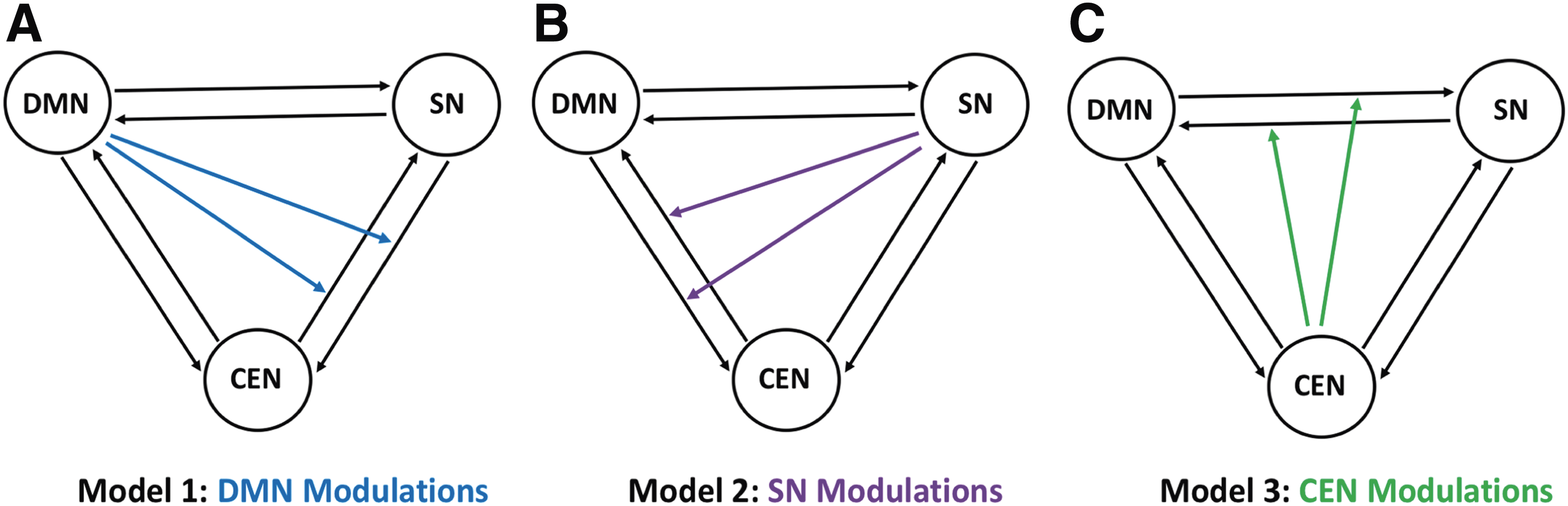
Model representation:
We carried out the RFX-BMS analysis (Fig. 4). The first column (Fig. 4A, B) shows the expected posterior model probability and exceedance probability for normal group. Similarly, the second column (Fig. 4C, D) shows the expected posterior probability and exceedance probability for MCI group. The RFX analysis revealed that model 2 has higher probability value than other models in normal group. It also revealed that model 2 no longer has higher probability in MCI group, instead the overall probability values shifted toward model 3. The three models were further compared within the normal control, MCI, and between the normal control and MCI groups, respectively (Fig. 5). In normal control (Fig. 5A), we found that model 2 had significantly higher probability than model 1 (p = 0.046; Mann–Whitney U test) and model 3 (p = 0.002), but there was no significant difference between models 1 and 3 (p = 0.531). In MCI group, we found that model 3 had significantly higher probability than model 1 (p = 8.53 × 10−19) and model 2 (p = 7.81 × 10−18), and model 2 has higher probability than model 1 (p = 1.91 × 10−4). Comparison between normal and MCI groups demonstrated a significant difference between two groups in each model (Table 2).

The first column

Comparison of models within groups and between groups:
To validate our findings, the FFX analysis was also performed and we found similar patterns of network interactions that model 2 had higher probability in normal controls and model 3 in MCI group (Supplementary Fig. S1; Supplementary Data are available online at
We further investigated the interactions of the salience network subsystems—the dorsal salience network and ventral salience network—with the default-mode and central-executive networks. We observed that the dorsal salience network exhibits control over the default-mode and central-executive networks in the normal group, but not in the MCI group (Fig. 6 for RFX results; Supplementary Fig. S3 for FFX results). In contrast, we also found that the ventral salience network does not exhibit control over the default-mode and central-executive networks in neither the normal group nor the MCI group (Fig. 7 for RFX results; Supplementary Fig. S4 for FFX results).


rsfMRI-cognition relation
We investigated the association between connectivity strength and MoCA score in patients with MCI (since normal group all had normal scores). Lower modulation strength from salience network on the other two networks derived from DCM analysis was associated with worse performance on cognitive testing as reflected by the MoCA scores (p = 0.034; r = 0.295). These results are presented in Figure 8.
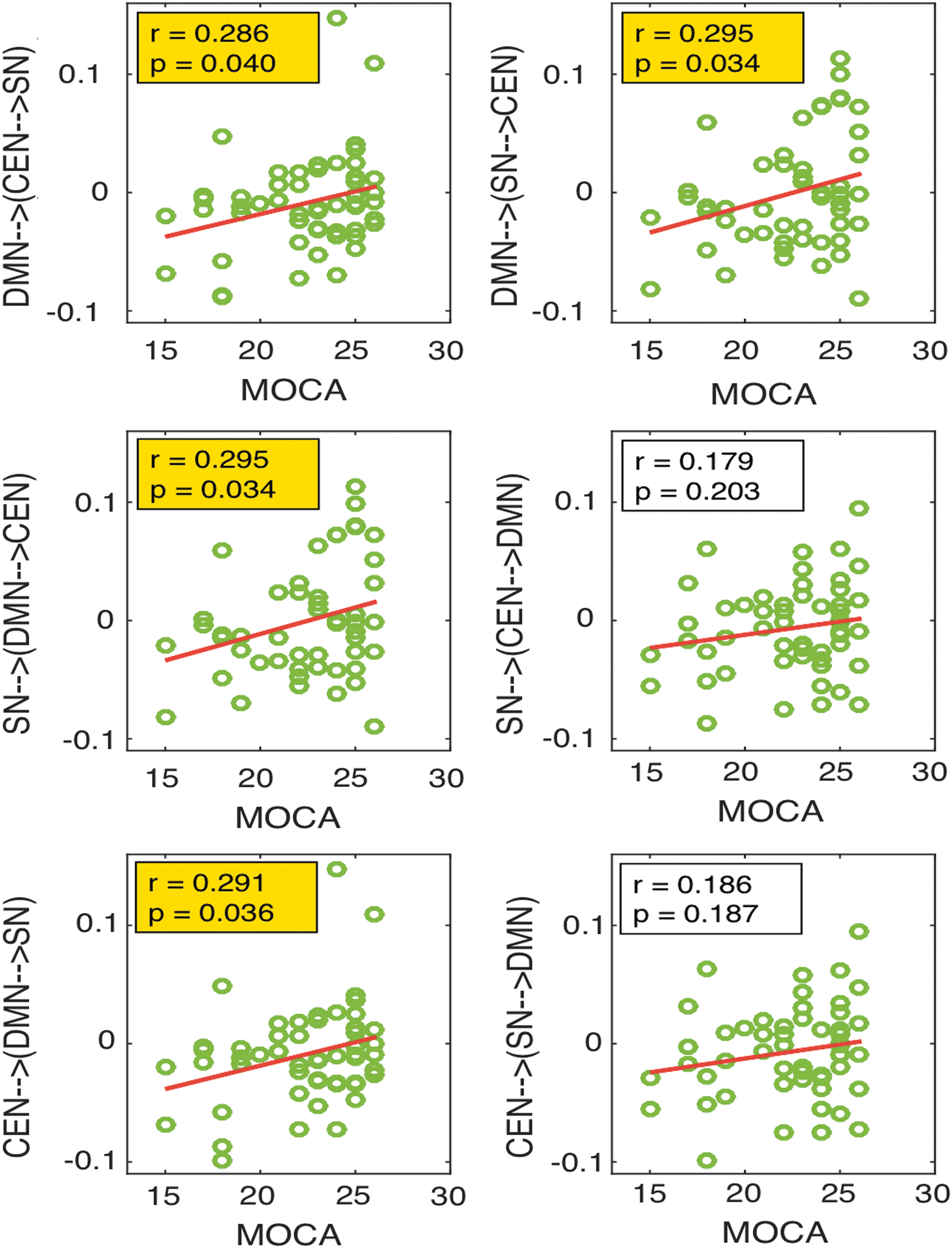
The correlation analysis between the network connections and MoCA scores in MCI group. The first row shows the correlation of connectivity of DMN on the connections between SN and CEN with MoCA, the second row shows the correlation of connectivity of SN on the connections between DMN and CEN with MoCA, and the third row shows the correlation of connectivity of CEN on the connections between DMN and SN with MoCA. MoCA, Montreal Cognitive Assessment. Color images available online at
Discussion
The present study showed that MCI is associated with a disrupted interaction among the default-mode, salience (dorsal salience), and central-executive networks, whereas it is maintained in cognitively normal elderly subjects. Interactions between the networks have been argued to be important for effective maintenance of normal cognitive status (Bressler and Menon, 2010; Chand and Dhamala, 2016a; Menon, 2015; Uddin, 2015). It is consistently reported that the salience network drives the default-mode and central-executive networks during both resting state and tasks in healthy younger population (Chand and Dhamala, 2016a; Goulden et al., 2014; Sridharan et al., 2008). Our results for healthy older (normal) group are consistent with those findings, suggesting that the dynamic switching role of salience network is intact in healthy aging. In addition, a comparison between hypertensive and normotensive cognitively normal individuals showed that there is no significant difference in the DCM model probability (Supplementary Fig. S1). We also found significant correlation between the modulation strength of the salience network over the other two networks and the neuropsychological test. These findings suggest that the difference in network interaction found in our MCI group is likely primarily driven by neuronal changes associated with cognitive impairment rather than vascular contributions due to hypertension in the MCI group.
Previous research has established that the default-mode network activity is impaired in MCI and Alzheimer's disease (Brier et al., 2014; Greicius and Kimmel, 2012; Greicius et al., 2004). Our results for MCI further demonstrated impaired control of the salience network over the default-mode and central-executive networks in this population, which may underlie reduced default-mode network activity. Our study was inspired by several studies that implemented network analysis in aging research. Recent study by Tsvetanov et al. (2016) reports the directed connectivity within and between the salience, dorsal attention, and default-mode networks. Our study consisted of central-executive network instead of dorsal attention network and tested a different hypothesis that whether the salience network is responsible in switching between the default-mode and central-executive networks, which was tested for healthy younger adults in previous studies (Goulden et al., 2014; Sridharan et al., 2008). In addition, we included the nonlinear modulations in interactions as implemented in prior investigations (Goulden et al., 2014; Li et al., 2011; Stephan et al., 2008).
It is interesting to note that the salience network-based control feature might be facilitated by unique cytoarchitecture of its key regions, the anterior insula and the dorsal anterior cingulate cortex (Bonnelle et al., 2012). These regions consist of von Economo neurons. These specific types of neurons are present in larger number in humans than in great apes, but not at all in other primates, and are suggested to relay information processed in these regions to the other parts of the brain (Allman et al., 2005, 2010; Watson et al., 2006). Whether these neurons are specific to the switching function of the salience network remains to be established. Previous studies have consistently reported co-activation of these regions (Chand and Dhamala, 2016b, 2017; Dosenbach et al., 2006; Seeley et al., 2007). The anterior insula is functionally connected to the networks responsible for adaptive behavior, including the salience network (Seeley et al., 2007) and the frontoparietal control network (Vincent et al., 2008). This area has direct white matter connections to other key regions, including the dorsal anterior cingulate cortex (Jilka et al., 2014), the inferior parietal lobe (Uddin et al., 2010), and the temporoparietal junction (Kucyi et al., 2012), making the insula well placed to perform its putative role of evaluating the environment and the cognitive state (Uddin et al., 2010), reorienting of attention (Ullsperger et al., 2010), and switching between cognitive resources (Uddin and Menon, 2009).
The dorsal anterior insula—a key node of the dorsal salience network—is reciprocally connected with multiple regions in prefrontal cortex, precentral operculum, parietal, and temporal cortices (Cerliani et al., 2012; Mesulam and Mufson, 1982a, b; Mufson and Mesulam, 1982) and is implicated to be important in attention and switching between cognitive resources (Sridharan et al., 2008; Touroutoglou et al., 2012). In contrast, the ventral anterior insula—a key node of the ventral salience network—is reciprocally connected with multiple limbic and paralimbic structures (Cerliani et al., 2012; Mesulam and Mufson, 1982a, b; Mufson and Mesulam, 1982) and is suggested to be crucial during affections or emotions (Kurth et al., 2010; Touroutoglou et al., 2012, 2016). The activity in the dorsal anterior cingulate cortex is known to signal the need for enhanced cognitive control and to implement behavioral changes (Egner, 2009). Our results for normal elderly control therefore provide strong evidence for the neural basis of the control feature of the salience network (Allman et al., 2005, 2010; Watson et al., 2006), consistent with previous reports in normal young subjects (Chand and Dhamala, 2016a; Goulden et al., 2014; Sridharan et al., 2008). Emerging evidence shows atypical engagement of the salience network in several neuropsychiatric disorders, such as in autism spectrum disorder, schizophrenia, and frontotemporal dementia (Menon, 2015; Uddin, 2015).
Prior studies report that the default-mode network activity decreases in MCI and/or with progression of Alzheimer's disease (Brier et al., 2014; Greicius and Kimmel, 2012; Greicius et al., 2004). Decreased probability of model 1 (i.e., modulations by default-mode network) in MCI compared to that of NC might indicate such changes as suggested by those studies. Dorsolateral prefrontal cortex—a key node of the central-executive network—has rich connections with several areas in the brain such as visual, somatosensory, and auditory areas and receives the information from occipital, parietal, and temporal cortices and is shown to be important in several cognitive functions, including executive function (Chand and Dhamala, 2017; Chand et al., 2016; Miller and Cohen, 2001; Petrides and Pandya, 1999). The central-executive network activity has been conflicted with disease (Diener et al., 2012). These works suggest this network, especially the dorsolateral prefrontal cortex node, to hypoactivate or hyperactivate with disease. Our results in MCI suggested that the probability of model 3 (i.e., modulations by central-executive network) is greater in MCI compared to that of NC implying hypermodulations by central-executive network. Our findings and existing literature taken together thus suggest that the salience network's control mechanism over default-mode and central-executive networks is achieved in healthy aging; however, this control is impaired and the network modulations are altered in MCI.
In conclusion, this study evaluated the resting-state patterns of directed interactions among the default-mode, salience, and central-executive networks in aging population with normal cognition and those with MCI. We found that the salience network, especially the dorsal salience network, played causal control over the default-mode and central-executive networks for normal control and that this feature was disrupted in MCI. These findings advance our current understanding of how the default-mode, salience, and central-executive networks coordinate neural information in healthy aging and in MCI.
Footnotes
Acknowledgments
The authors thank the members of our team and the subjects who volunteered for these studies. This research was conducted with NIA/NIH grants RF1AG051633 and R01AG042127 to I.H. D.Q. is supported by NIH grants AG25688, AG42127, AG49752, and AG51633 and has received research support from Medtronic and Siemens Medical Solutions.
Author Disclosure Statement
All authors have approved the article and agree with submission to Brain Connectivity. We have read and have abided by the statement of ethical standards for manuscripts submitted to Brain Connectivity.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.