
Abstract
The present study aimed to explore the association between resting-state functional connectivity and creativity ability. Toward this end, the figural Torrance Tests of Creative Thinking (TTCT) scores were collected from 180 participants. Based on the figural TTCT measures, we collected resting-state functional magnetic resonance imaging data for participants with two different levels of creativity ability (a high-creativity group [HG, n = 22] and a low-creativity group [LG, n = 20]). For the aspect of group difference, this study combined voxel-wise functional connectivity strength (FCS) and seed-based functional connectivity to identify brain regions with group-change functional connectivity. Furthermore, the connectome properties of the identified regions and their associations with creativity were investigated using the permutation test, discriminative analysis, and brain–behavior correlation analysis. The results indicated that there were 4 regions with group differences in FCS, and these regions were linked to 30 other regions, demonstrating different functional connectivity between the groups. Together, these regions form a creativity-related network, and we observed higher network efficiency in the HG compared with the LG. The regions involved in the creativity network were widely distributed across the modality-specific/supramodality cerebral cortex, subcortex, and cerebellum. Notably, properties of regions in the supramodality networks (i.e., the default mode network and attention network) carried creativity-level discriminative information and were significantly correlated with the creativity performance. Together, these findings demonstrate a link between intrinsic brain connectivity and creative ability, which should provide new insights into the neural basis of creativity.
Introduction
C
The spontaneous activity of the human brain is self-organized within a network and can be revealed by resting-state functional magnetic resonance imaging (rs-fMRI) (Beckmann et al., 2005; Biswal et al., 1995; Mitra et al., 2014). The concept of graph theory is well suited for characterizing and depicting the topological organization of brain networks (Bullmore and Sporns, 2009; Lee et al., 2016; Rubinov and Sporns, 2010). Graph-based network analyses have demonstrated the existence of an optimized topology in brain networks with efficient information transmission and exchange (Fallani et al., 2014; Zuo et al., 2012). An optimized brain network is the neural basis of cognitive behavior (Achard and Bullmore, 2007), and network analysis provides a new avenue for elaborating the neural basis underlying cognition.
Many studies have shown that the majority of neural responses to cognition tasks can be shaped by intrinsic brain activity (Aziz-Zadeh et al., 2013; Dietrich and Kanso, 2010; Wei et al., 2014; Zhao et al., 2014), or in other words, intrinsic brain activity could be the neural network preparing resources for cognition performance. Therefore, the intrinsic activity modality could be closely associated with behavior performance (Bolay et al., 2002; Fox and Raichle, 2007; Zou et al., 2013). Correspondingly, researchers have shown that the interindividual creativity divergence is highly related to resting-state functional connectivity (De et al., 2016; Kounios et al., 2008; Takeuchi et al., 2012). It also appears that the regions involved in creativity-related functional connectivity are widely distributed across the default mode network (DMN; e.g., the posterior cingulated), the medial prefrontal cortex, and dorsolateral superior frontal gyrus (DLSFG) (Takeuchi et al., 2012, 2017; Wei et al., 2014). The middle temporal gyrus (MTG) may be central to the generation of novel ideas which are key to creativity (Ellamil et al., 2012). In addition, the inferior temporal gyrus (ITG) and cerebellum were also found to be highly associated with creativity behavior performance (Chávez-Eakle et al., 2007; Kounios et al., 2008; Mayseless et al., 2014). These observations imply that widespread brain regions and their interconnectivity constitute a brain network that sustains creativity performance. Therefore, a network perspective should provide a fuller view of the neural substrates underlying creativity behavior performance.
In this study, we used a data-driven approach to explore the resting-state brain network relevant to creativity behavior. Toward this end, participants were recruited for high-creativity group (HG, n = 22) and low-creativity group (LG, n = 20) based on the figural Torrance Tests of Creative Thinking (TTCT). Then, we collected rs-fMRI data for each participant in the two groups. Based on these data, the voxel-wise functional connectivity strength (FCS) of the whole brain was calculated for each participant, and the brain regions with different FCS between the two groups were identified. The above-identified brain regions were then further analyzed as regions of interest (ROIs), and an ROI-based functional connectivity analysis method was used to trace the regions that originated the FCS changes in the two groups. Finally, the ROIs and traced regions were analyzed together, and the interconnections were measured using graph-based network analysis.
Materials and Methods
Participants
For the present study, a total of 180 right-handed healthy undergraduates (90 males and 90 females, age range from 18 to 22 years) were recruited from the campus of South China Normal University (SCNU, Guangdong, China). The creativity score of each participant was measured using the figural TTCT questionnaire. The TTCT, which includes verbal, figural, and auditory tests, was designed to be a measure divergent thinking, a central aspect of creativity (Kleibeuker et al., 2017; Takeuchi et al., 2017; Wei et al., 2014). Visual creativity is of importance that it reflects the character of divergent thinking and the figure creativity test is found to be effective to measure the visual creativity. Thus, in this study, we used the figural TTCT to identify the participants with high visual creativity performance, which has been widely used in many studies (Chávez-Eakle et al., 2007; Huang et al., 2012; Kim et al., 2006; Pidgeon et al., 2016). The figural TTCT comprises three tasks. One of the tasks required participants to imagine a picture or a story based on an egg-shaped line figure that was pictured in a paper. One of the tasks required participants to draw some interesting things based on 10 unfinished pictures. The last task required participants to add some lines to draw some different objects in 30 pairs of parallel lines. For each task, scoring comprised four components: fluency (the number of relevant responses, which is related to the ability to consider different possibilities), flexibility (the ability to shift between two conceptual fields), originality (the degree of unusual responses, which is associated with breaking away from the common and obvious thinking habits), and elaboration (the amount of details on each response). The TTCT has suggested high predictive validity (r > 0.57) and high reliability (r > 0.90) (Torrance, 1988, 1990, 1993). The current study used the total creativity scores (sum of fluency, flexibility, originality, and elaboration scores). In addition, we also examined the intellectual ability of each participant using the Combined Raven's Test (CRT), which is a widely recognized intelligence test with high reliability and validity (Tang et al., 2012). The previous findings suggested that gender might influence creativity ability (Abraham et al., 2014; Lin et al., 2012; Ryman et al., 2014). Based on the figural TTCT scores, we selected 22 participants from the top 12% of figural TTCT scores (11 males, 18.86 ± 1.08 years) as the HG and 22 participants (11 males, 19.13 ± 0.99 years) from the bottom 12% of figural TTCT scores as the LG. The previous studies have proposed that gender might influence creativity ability (Abraham et al., 2014; Lin et al., 2012). Thus, we selected the same number of females and males to minimize the influence of gender differences. To maintain the homogeneity of each group, we excluded two subjects whose TTCT scores more than 50 in LG (9 males, 19.10 ± 1.02 years). Then, we collected rs-fMRI scans for all selected participants in the two groups. This study was approved by the Institutional Review Board of SCNU. Each participant provided informed written consent for the present study. Table 1 lists the detailed demographic data of the two groups.
The t values were obtained using a two-sample t-test.
The t value was obtained using a chi-square test.
p < 0.01.
CRT, Combined Raven's Test; figural TTCT, the figural Torrance Tests of Creative Thinking; HG, high-creativity group; LG, low-creativity group.
Image acquisition
All participants were scanned using a 3T Siemens Trio Tim MR scanner at the Brain Imaging Center of SCNU. The rs-fMRI data were collected using a GE-echo planar imaging (EPI) sequence: 32 axial slices; repetition time (TR) = 2000 msec; echo time (TE) = 30 msec; slice thickness = 3.5 mm; no gap; flip angle (FA) = 90°; matrix = 64 × 64; field of view (FOV) = 192 × 192 mm. The participants were instructed to lay quietly with their eyes closed during the scans. After scanning, a total of 240 volumes were obtained for each participant. In addition, individual high-resolution three-dimensional structural images were also acquired using a T1-weighted MP-RAGE sequence: TR = 1900 msec; TE = 2.52 msec; slice thickness = 1 mm; no gap; FA = 9°; matrix = 256 × 256; FOV = 256 × 256 mm.
Data preprocessing
The rs-fMRI data were preprocessed using DPARSF (Yan et al., 2009) based on SPM8 (
Voxel-wise FCS
In this study, the degree centrality was considered to be an important property in whole-brain FCS (Buckner et al., 2009; Zuo et al., 2012). In calculations, the time series of each voxel of the preprocessed fMRI data were extracted for each participant using a GM probability map (threshold = 0.2) to constrain all the analyses within the GM. We obtained Pearson's correlations between the time series of all pairs of voxels in the entire brain, generating a whole-brain connectivity matrix for each participant. The individual correlation matrices were further translated to a z-score matrix using the Fisher's r-to-z approach. In the z-value matrix, the FCS of each voxel was calculated by computing the sum of the connections for a given voxel with all other voxels. In this study, functional connectivity was restricted to positive scores due to the ambiguous interpretation of negative correlations (Wang et al., 2014). Finally, an FCS of each participant was normalized by translating to z-score values within each participant of the two groups.
Seed-based functional connectivity
Using the FCS analysis, we were able to identify the brain sites that showed differences in degree centrality between the two groups based on a two-sample t-test (for details, see the Statistical Analyses section). The degree centrality is an important indicator of the FCS. In this study, we wished to obtain greater insight into the regions showing different FCS. Therefore, the regions that showed differences in degree centrality between two groups were further analyzed as ROIs, and we chose seed regions (defined as a sphere centered at the ROI coordinate with a 6-mm radius) to calculate the voxel-wise functional connectivity of the whole brain using the preprocessed data. The resulting FCS data were further subjected to Fisher's r-to-z transformation to improve normality. The above manipulations were performed using REST (
Network analysis
The functional connections of the regions with different FCS and the subsequently traced regions were further analyzed due to their high likelihood of being involved in creativity performance. To reveal the organization among these regions, we constructed a connectivity matrix individually and then analyzed them using graph-based analysis. Network analysis was implemented in MATLAB using GRETNA (Wang et al., 2015). Briefly, a spherical ROI was generated for each of these regions, with the centroid located at the corresponding peak voxel (radius = 6 mm). We found out in total of 36 regions by combining the FCS method and the ROI-based functional connectivity approach, which also have been used in a Parkinson's disease study and the study suggested a meaningful result (Zhang et al., 2015a). In order to overcome the issue of the overlap among these regions, we excluded potentially overlapping ROIs based on the Euclidean distance if the Euclidean distance <6 mm (Zhang et al., 2015a). Based on the above calculation, we finally excluded one brain region. These remaining regions were treated as the nodes of the brain network, and we further extracted the mean time series of each node for each participant and calculated the Pearson's correlation between any two of them as edges. In this way, we obtained an n × n correlation matrix (n is the number of remaining regions) for each participant. To exclude possible effects due to spurious correlations on network topology, we chose a sparsity threshold (i.e., the ratio of the number of existing edges divided by the maximum possible number of edges in a network) to convert the individual correlation matrices, such that only the highest correlations remained. The sparsity approach normalized all resultant networks to have the same number of nodes and edges and also minimized the effects of discrepancies in the overall correlation strength between groups. We empirically thresholded each correlation matrix multiple times over a wide range (0.1059 < sparsity <0.5859; interval = 0.02) to obtain sparse and weighted networks.
We calculated multiple network measures, including the clustering coefficient (Cp ), shortest path length (Lp ), local efficiency (Eloc ), and global efficiency (Eglob ) (Achard and Bullmore, 2007; He et al., 2009a). The area under the curve (AUC) method was used for each network metric to provide a summarized scalar independent of single threshold selection (Watts and Strogatz, 1998; Zhang et al., 2011).
Statistical analyses
A two-sample t-test was performed on the individual FCS data from the rs-fMRI data and the FCS data for each ROI to identify brain regions that exhibited differences between HG and LG. Statistical analyses were corrected using the Monte Carlo simulations approach implemented in REST. Before calculations, the FCS data were z-score transformed to improve normalization and spatially smoothed (FWHM = 8 mm).
Nonparametric permutation tests (10,000 permutations) were used to identify between-group differences in network parameters (Cp , Lp , Eloc , and Eglob ). Age, gender, and CRT scores were treated as unconcerned covariates for all comparisons.
Afterward, a discriminant analysis (
Finally, we explored a partial correlation (Pearson correlation) between the figural TTCT scores and network/regional efficiency (Cp , Lp , Eloc , and Eglob ) in each group, in which age, gender, and CRT scores were treated as unconcerned covariates. When we explored the correlation between the figural TTCT scores and regional (nodal) efficiency, we used the 1/n (n = 34) criterion to perform multiple comparison correction (Bassett et al., 2009; Lynall et al., 2010; Zhang et al., 2015b). Then we computed the effect size (Cohen d) and statistical power which was determined by Cohen d (Cohen, 1977, 2013). The Cohen d of Cp , Lp , Eloc , and Eglob were 0.8, 1.1, 0.9, and 0.8, respectively.
Results
Voxel-wise FCS
We explored the voxel-wise FCS of the whole brain in HG and LG, and a similar pattern was observed for the group-mean FCS between HG (Fig. 1A) and LG (Fig. 1B). The group-mean FCS was distributed across the entire brain, and the regions with the strongest FCS were predominately distributed across the bilateral precuneus, bilateral cingulated cortex, left cuneus gyrus, bilateral superior occipital gyrus, bilateral calcarine fissure, bilateral surrounding cortex, bilateral lingual gyrus, and bilateral cerebellum. A similar FCS distribution pattern was observed in both the HG and LG.
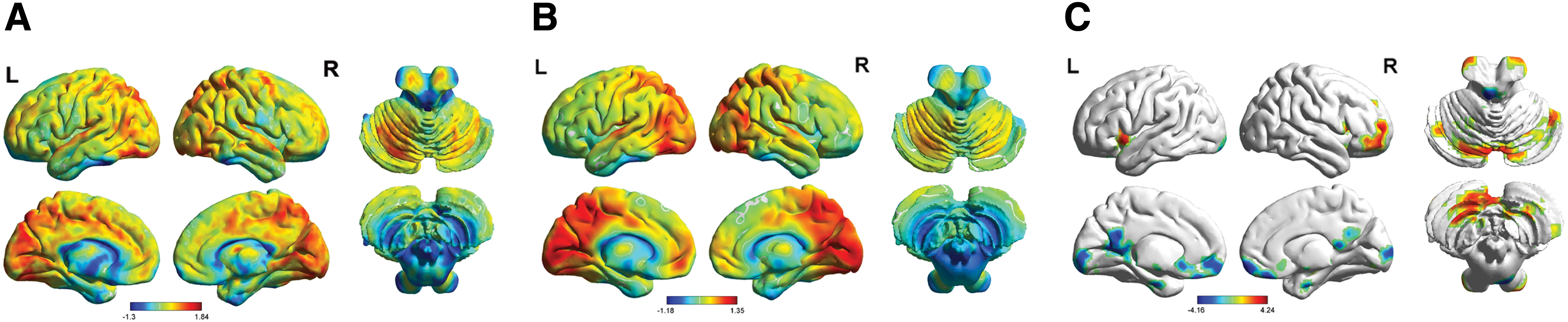
The results of the within/between-group analyses of FCS.
However, further analysis revealed that four FCS regions showed differences between the two groups (Fig. 1C and Table 2). Among these regions, the FCS of the right cerebellum was significantly increased in HG compared with LG. We also found significant decreases in FCS in the right rectus gyrus, the right calcarine fissure and the left cerebellum in HG compared with LG.
The ROIs of identifying the brain sites that showed difference of FCS between the two groups.
Regions from the right inferior cerebellum, center = [27 − 66 − 57].
Regions from the right gyrus rectus, center = [6 39 − 24].
Regions from the right calcarine fissure, center = [12 − 96 6].
Regions from the left superior cerebellum, center = [−6 − 39 − 6].
ATN, the attention network; AUN, the auditory network; DMN, the default mode network; FCS, functional connectivity strength; L, left hemisphere; MNI, Montreal Neurological Institute; R, right hemisphere; ROIs, regions of interest; SCN, the subcortical network; SEN, the sensorimotor network; VIN, the visual network.
Figure 2 shows the between-group differences in seed-based functional connectivity in HG and LG. The details of the traced regions with respect to functional connectivity are listed in Table 2. The ROIs and the traced regions could together represent a creativity-related network (Fig. 3). The involved regions of the creativity-related network were widely distributed across the cerebral cortex, subcortex regions, and cerebellum. Perhaps unsurprisingly, most of the relevant regions were located at the cerebral cortex, including the modality-specific networks (e.g., the visual network [VIN], the auditory network [AUN], and the sensorimotor network [SEN]) and supramodality networks (e.g., the DMN and the attention network [ATN]). Notably, these brain networks (Table 2) were identified based on the findings of resting-state functional connectivity studies (He et al., 2009b; Heine et al., 2012; Zhang et al., 2010).
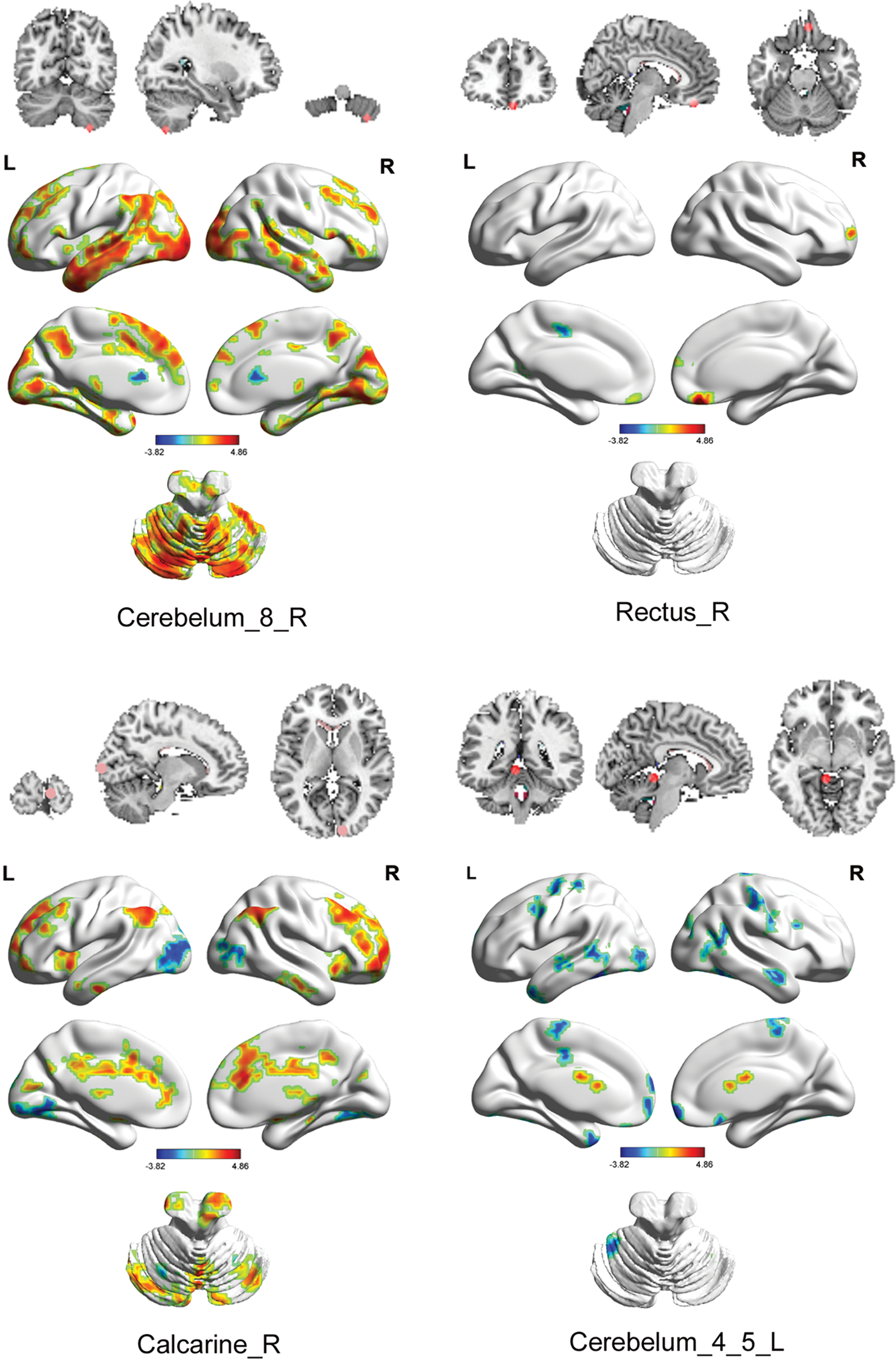
Between-group differences in seed-based functional connectivity (p < 0.05, corrected). Color images available online at
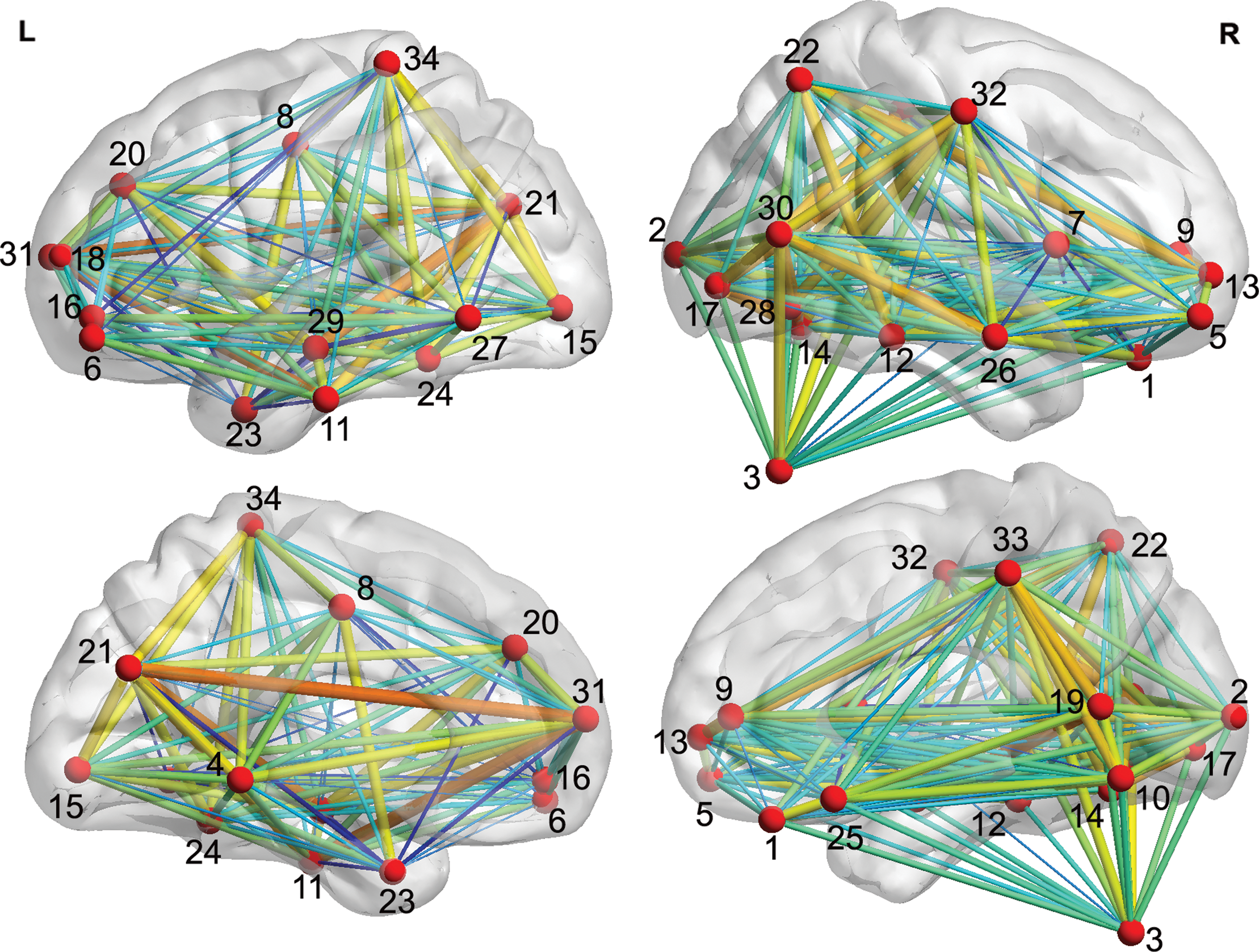
A brain surface representation of the 34 ROIs and their connections within the creativity-related network. ROIs, regions of interest. Color images available online at
Network parameters
Results revealed that the HG had shorter Lp (p = 0.0003) and higher Eglob (p = 0.0006) values than the LG (Fig. 4A). Furthermore, we found that there was negative correlation between Lp and the figural TTCT scores (r = −0.5082, p = 0.0010) (Fig. 4B). Similarly, we found a positive correlation between Eglob and the figural TTCT scores (r = 0.5132, p = 0.0008) (Fig. 4C).
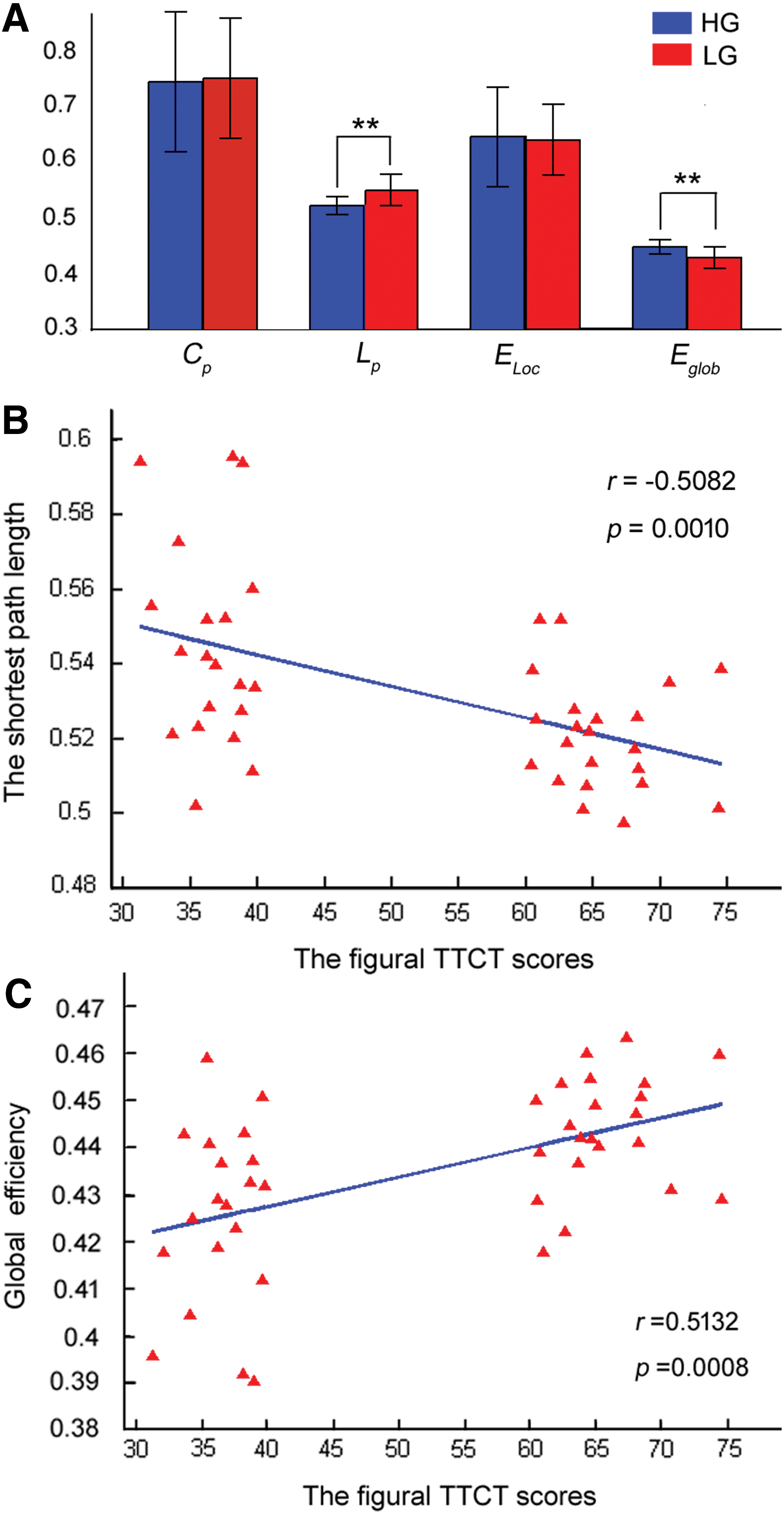
Regional network parameters
We found that the properties of the regions related to the supramodality network obtained discriminative power in distinguishing HG and LG. These results showed these parameters had good distinction power. First, we found that the FCS between the right MTG (DMN) and the right middle frontal gyrus (MFG) (ATN) exhibited the highest discriminative power (AUC = 0.8309, SE = 0.06276, 95% CI area = 0.70788 – 0.9539) (Fig. 5A). Second, we found that the right DLSFG (DMN) exhibited discriminative power and that the Cp (AUC = 0.77572, SE = 0.07210, 95% CI area = 0.63441 – 0.91704) and Eloc (AUC = 0.8091, SE = 0.06720, 95% CI area = 0.67742 – 0.94082) values from this region could be used to effectively classify the two groups (Fig. 5B–C). Moreover, the Eglob value of the left DLSFG (DMN) also exhibited discriminative power (AUC = 0.8956, SE = 0.05042, 95% CI area = 0.79679 – 0.99442; Fig. 5D).
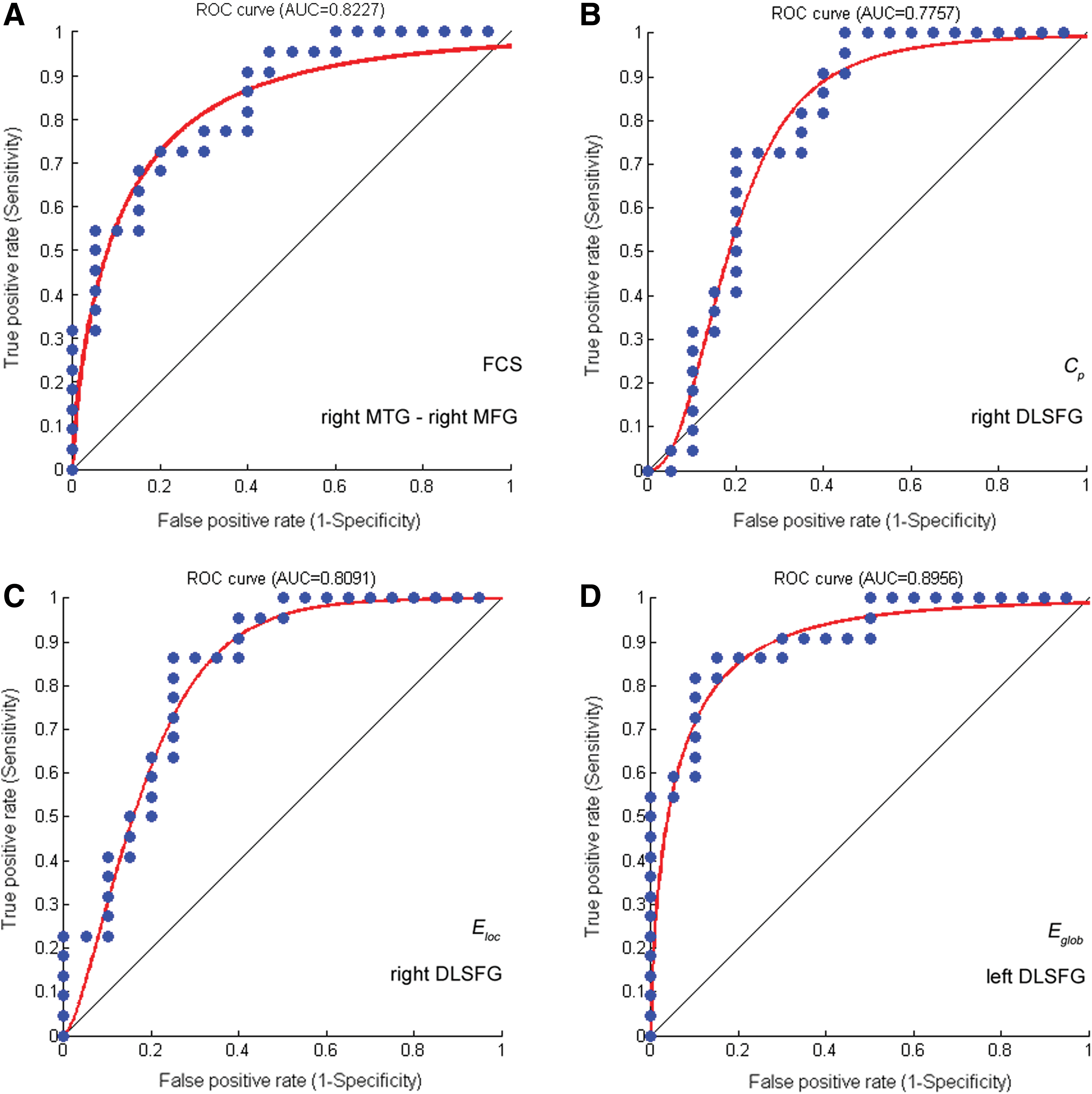
ROC curve for distinguishing HG and LG using network properties. ROC, receiver operating characteristic. Color images available online at
In addition, there were significant correlations for Cp , Eloc , and Eglob with the figural TTCT in many regions in the two groups (Table 2 and Fig. 6). The results showed a significant negative correlation between regional efficiency (Cp ) and the figural TTCT scores in the right postcentral gyrus (PCG) (SEN) in HG. Similarly, in the same region there was a significant negative correlation between regional efficiency (Eloc ) and the figural TTCT scores in HG. Moreover, the relationship between global efficiency (Elglob ) and the figural TTCT scores in HG showed a significant positive correlation in the left ITG (ATN).
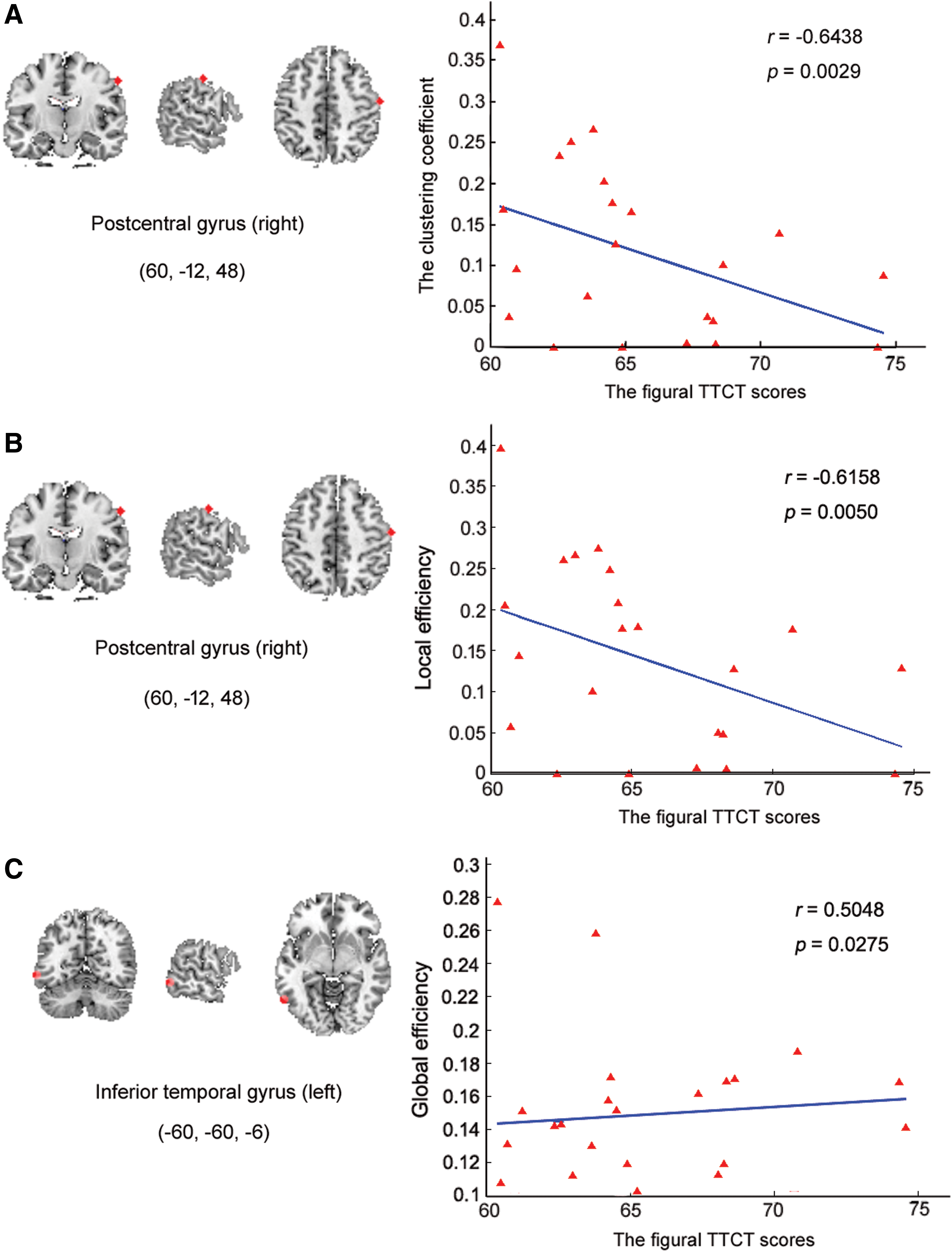
Significant correlations between Cp
, Eloc
, and Eglob
and the figural TTCT scores for the HG.
Discussion
This study used a data-driven approach to explore the functional brain connections underlying creativity ability in terms of group differences and brain–behavior correlations. The main results can be summarized as follows: (1) there were many functionally connected regions that showed group differences in the modality-specific/supramodality cerebral cortex, subcortex, and cerebellum, and together these regions form a creativity network in which information translation efficiency is higher in HG than in LG; and (2) the properties of the supramodality regions within the creativity network can be used to discriminate between HG and LG and are significantly correlated with creative performance.
We first used degree centrality to compute the differences between two groups. Degree centrality is the number of edges connecting to a node (Zuo et al., 2012). The degree centrality is an important indicator of the functional connectivity strength. It represents the most local and directly quantifiable centrality measure. The degree centrality was different between two groups, it means FCS has differences between high-creativity and low-creativity group. On this aspect, we think the ROIs which derived from different analysis (degree centrality and FCS analysis) played similar roles in the network in the current study.
Many previous studies have shown that the patterns of intrinsic brain activity are trait related (Heine et al., 2012; Mitra et al., 2014; Rubinov and Sporns, 2010) and that resting-state functional connectivity is highly associated with creativity ability (Chen et al., 2014; Takeuchi et al., 2012). To extend these previous findings using aspects of individual divergence, we explored voxel-wise FCS across the whole brain between groups of participants with high- and low-level creativity in a resting state, and we found changes in the FCS of the DMN (e.g., the left DLSFG and right rectus gyrus), attention-related regions (e.g., the right MFG), and cerebellum between the two groups. Previous studies focusing on single brain regions identified the location and function of specific regions in creativity performance (Aziz‐Zadeh et al., 2009; Fink et al., 2009; Li et al., 2014), demonstrating the importance of the right MFG in semantic divergence and creative story generation (Howard-Jones et al., 2005) and insight solutions (Aziz‐Zadeh et al., 2009). In addition to these above regions, activation of the right insula was observed during creative assessment tasks (Mayseless et al., 2014) and the insight and search solution (Aziz‐Zadeh et al., 2009). Notably, creativity is a complex cognitive process that includes multiple mental processes, including hypothesis generation, search solution, and insight (Aziz‐Zadeh et al., 2009; Dietrich and Kanso, 2010; Howard-Jones et al., 2005; Liu et al., 2015; Zhu et al., 2017). Thus, the results of this study with respect to spontaneous brain activity are consistent with the crucial roles of the aforementioned regions (e.g., the left DLSFG, right MFG, and the left ITG). Furthermore, the agreement between our findings and those of previous studies demonstrates the sensitivity and rationale of using voxel-wise FCS to explore the effects of individual divergences in intrinsic brain activity characteristics on creativity levels.
Many studies have shown that the intrinsic activity of brain regions creates a complex network of interactions (Bolay et al., 2002; Fox and Raichle, 2007). Consistently, it has been shown that creativity is not due to cognitive activities in isolated brain region, but rather is the result of interactions and interconnections between multiple brain regions (de Souza et al., 2010; Dietrich and Kanso, 2010; Fink et al., 2009) and multiple brain networks (Beaty et al., 2016, 2017). In this study, using a seed-based functional connectivity approach, we explored the regions of origin of the observed changes in FCS, and we found a number of regions that were widely distributed across the supramodality and modality-specific networks. According to the hypothesis of embodiment cognition, human cognition is not beyond perception, but is rather tightly related to the specific modality (Overwalle et al., 2015). In the present study, we also found that many modality-specific regions (e.g., SEN, VIN, and AUN), in addition to the supramodality regions (e.g., the DMN and ATN), together comprise an interacting functional brain network. Indeed, with respect to brain spontaneous activity, the intrinsic activity of the modality-specific regions (e.g., the primary visual cortex) was observed to be highly coordinated with that of the supramodality regions, perhaps creating a pattern for mental imagery (Wang et al., 2008), which was shown to be correlated with the creativity performance (LeBoutillier and Marks, 2003). The most important advance of the current work is that we identified a functional connectivity network for creativity by searching the brain connectome at the voxel level, suggesting the existence of a network associated with creativity.
It should be noted that graph-based network analysis provides one approach for depicting the topological organization of complex interactions between multiple regions within a network. With respect to aspects of information translation efficiency, the topological organization of brain networks was found to be different for certain mental traits (e.g., disease [Kullmann et al., 2014], intelligence [Sawaya et al., 2015], personality [Li et al., 2014], and mental state [Geerligs et al., 2015]). In this study, we explored network organization using two groups with different creativity levels to analyze not only the shortest path length but also the network efficiency on the group-difference aspect. Similar to our previous study of dynamic functional connectivity in human creative brain (Li et al., 2017), the results from both aspects showed that the brain networks of individuals with high creativity levels exhibited better optimized network organization compared with those from low creativity individuals. These findings provide evidence for the existence of an optimized, creativity-related functional brain network, and they also demonstrate the rationale of the data-driven approach used in the present study. In addition, the properties of the creativity-related network, such as the hub regions, should be further explored in future works.
In addition to our findings on the creativity-related brain network, the present study further explored the relationship between regional network properties and creative ability. For the group-difference aspect, we found that discriminative analysis could effectively classify the two groups using the network properties of the DMN (e.g., the right MTG and left DLSFG) and ATN (e.g., the right MFG) as features. These findings suggest that the most discriminative features between the two groups were attributes of the supramodality regions linked to the creativity-related network, although this network was widely distributed and included both the modality-specific and supramodality regions. We also explored correlations between the regional properties of the creativity-related network and creativity performance, and we found that that the information translation efficiency of the ATN (e.g., the left ITG) within the creativity-related network was significantly correlated with the creativity performance. At the same time, the regional translation efficiency of SEN (e.g., the right PCG) within the creativity-related network was significantly negative correlated with creative performance. These observations indicate that the variance of network properties in the supramodality regions can explain individual divergence in creative performance. With regard to intersubject variability, DMN demonstrates the highest level of functional variability, with the ATN a close second; in addition, the activity of both regions is significantly more variable than for the SEN and VIN (Mueller et al., 2013). In other words, individual differences are primarily reflected in the properties of high-level brain regions. Notably, for the group-difference and brain–behavior correlation aspects, we consistently found that the network properties of the supramodality regions within our identified creativity-related network were highly associated with creativity ability. In fact, many previous studies have shown the importance of intrinsic functional connectivity of the DMN in creativity (Beaty et al., 2014, 2015). Consistently, we observed a tight association between the supramodality regions (e.g., the DMN and ATN) and creativity performance. To our best knowledge, attention is one of the core elements of creativity. HG showed better connectivity between the DMN and ATN, suggesting that HG had greater top-down control over the imaginative processes stemming from the DMN (Beaty et al., 2015) and that HG can better control their attention being creative. Consequently, our findings provide further evidence for the role of attention in the creative process.
Limitations and Directions
There are several limitations that need to be addressed in future studies. First, creativity levels were measured based on divergent thinking using TTCT scores. Although divergent thinking is the key component of creativity ability and several previous studies have used the figural TTCT scores as a measure of creativity (Dietrich and Kanso, 2010; Torrance, 1998), creativity is a complex cognitive process with many other aspects besides divergent thinking. Thus, other dimensions of creativity should be considered in future works. Second, with respect to individual divergence, the present study showed significant differences in network connections between HG and LG. However, it should be noted that these findings were derived from the healthy undergraduate students, and it should be determined whether these conclusions extend to other populations. Third, by using resting-state data, we identified a creativity-related network whose properties were related to creativity levels. However, it remains unclear how this creativity network would perform during creativity tasks, which should be explored in future studies. Finally, the main results were derived from a small sample size dataset, which should be further validated using a generalized population in future works.
Conclusions
The present study used a data-driven approach to identify creativity-related intrinsic brain connections, including regions of the modality-specific/supramodality cerebral cortex, subcortex, and cerebellum. The topological organization of the functional connectome was dependent on creativity level. The properties of the supramodality regions (i.e., the DMN and ATN) exhibited discriminative power in classifying the creativity of participants and also displayed strong correlations with creative performance. These findings have important implications for understanding the neural substrates underlying creativity.
Footnotes
Acknowledgments
This work was supported by the Natural Science Foundation of China (No. 31600907 and No. 31371049), the Guangdong Provincial Natural Science Foundation of China (No. 2014A030310487), and the School of Psychology of SCNU (No. hsxly2016021).
Author Disclosure Statement
No competing financial interests exist.