
Abstract
Fingerprint patterns derived from functional connectivity (FC) can be used to identify subjects across groups and sessions, indicating that the topology of the brain substantially differs between individuals. However, the source of FC variability inferred from resting-state functional magnetic resonance imaging remains unclear. One possibility is that these variations are related to individual differences in white matter structural connectivity (SC). However, directly comparing FC with SC is challenging given the many potential biases associated with quantifying their respective strengths. In an attempt to circumvent this, we employed a recently proposed test–retest approach that better quantifies inter-subject variability by first correcting for intra-subject nuisance variability (i.e., head motion, physiological differences in brain state, etc.) that can artificially influence FC and SC measures. Therefore, rather than directly comparing the strength of FC with SC, we asked whether brain regions with, for example, low inter-subject FC variability also exhibited low SC variability. From this, we report two main findings: First, at the whole-brain level, SC variability was significantly lower than FC variability, indicating that an individual's structural connectome is far more similar to another relative to their functional counterpart even after correcting for noise. Second, although FC and SC variability were mutually low in some brain areas (e.g., primary somatosensory cortex) and high in others (e.g., memory and language areas), the two were not significantly correlated across all cortical and sub-cortical regions. Taken together, these results indicate that even after correcting for factors that may differently affect FC and SC, the two, nonetheless, remain largely independent of one another. Further work is needed to understand the role that direct anatomical pathways play in supporting vascular-based measures of FC and to what extent these measures are dictated by anatomical connectivity.
Introduction
E
Recently, two magnetic resonance imaging (MRI)-based methodologies have emerged as non-invasive measures of the brain's functional connectivity (FC) and structural connectivity (SC). Functional MRI (fMRI) provides four-dimensional whole-brain images that reflect changes in cortical blood flow, volume, and oxygen as measured by the blood-oxygenation-level-dependent (BOLD) signal (Bandettini et al., 1993; Kwong et al., 1992; Turner, 1992). At rest, the spontaneous low-frequency fluctuations (<0.08–0.1 Hz) in the BOLD signal allow the detection of temporally correlated spatial patterns that are specific to each individual (Biswal et al., 1995; Damoiseaux et al., 2006) and are generally believed to reflect neuronal synchronization between brain areas [see Buckner et al. (2013) and references therein]. Diffusion MRI (dMRI), on the other hand, provides information about the structural architecture of the brain by probing the diffusion of water molecules in the white matter (WM) and is often used to infer anatomical or SC between brain regions.
It is well established that the strength of BOLD FC (often expressed as a correlation coefficient or z-score) varies across healthy individuals, though the source of this variability is unclear. Several studies have suggested that the “strength” or “integrity” of direct anatomical pathways between brain areas is related to the strength of their FC (Bowman et al., 2012; Goni et al., 2014; Hermundstad et al., 2013; Honey et al., 2009; Sporns, 2013; van den Heuvel et al., 2009; Varkuti et al., 2011; Ward et al., 2014; Zhu et al., 2014). However, other studies have reported a poor correlation between the two. For instance, FC is barely altered in patients with aggressive brain tumors (Ghumman et al., 2016), despite the fact that such insults can dramatically alter SC (Fornito et al., 2015; Leclercq et al., 2011; Nimsky et al., 2005, 2016). Similarly, individuals with complete agenesis of the corpus callosum (CC), nonetheless, display intact bilateral FC (Tyszka et al., 2011), demonstrating that FC between two brain areas is not necessarily dependent on a direct anatomical connection (Buckner et al., 2013).
It is also well established that BOLD FC varies more across subjects than repeated measures within the same subject (Finn et al., 2015), and Mueller and associates (2013) were the first groups to explicitly demonstrate that functional inter-subject variability
To investigate to what extent variations in SC are related to FC variability, we applied the approach proposed by Mueller and colleagues (2013) (originally intended for mapping functional variability) on SC matrices derived from dMRI tractography initiated from the WM and gray matter (GM) interface. The resulting structural variability indices were then compared with functional variability maps to determine whether those variances are predictive of one another. Using this approach, we report whole-brain SC variability measures that take into account the aforementioned biases and compare them with FC variability. We globally determined that structural variability is lower than functional variability. We also observed that regions of low (i.e., motor areas) and high (i.e., memory and language areas) functional variability also exhibited signs of low and high structural variability, respectively. Finally, our results suggest that the bulk of inter-individual variability in FC cannot be explained by corresponding variability in SC.
Methods
Anatomical, diffusion, and functional datasets were obtained on a 1.5 T Siemens Magnetom from nine subjects (s) over three sessions (t) separated by a 1 month interval (seven men; mean age 25 years ±3.1; range 21–30). An additional functional dataset was acquired during the last session for all participants. The study was performed according to the guidelines of the Internal Review Board of the Centre Hospitalier Universitaire de Sherbrooke. Acquisition details and processing steps are illustrated in Figure 1 and described later.

SC and FC pipeline. Brain parcellation was first performed on each subject-specific anatomical images, resulting in 159 cortical and subcortical regions. Top: FC matrices were generated by averaging the preprocessed signals within each of the brain regions and correlating them between each other. FC matrices were group-normalized by applying Fisher's r-to-z transform. Bottom: Probabilistic streamline tractography was done by using particle filtering tractography (Girard et al., 2014) with dense seeding from the white matter and gray matter interface (10 seeds/voxel). SC matrices were generated by computing the connectivity between pairs of cortical regions and normalized to account for various total streamline counts. FC, functional connectivity; SC, structural connectivity. Color images available online at
Structural connectivity
High angular resolution diffusion imaging (HARDI) data were acquired along 64 uniformly distributed directions by using a b-value of 1000 sec/mm2, a single-shot echo-planar imaging (EPI) sequence (128 × 128 matrix, 2 mm isotropic resolution, TR/TE 11,000/98 msec), and a GRAPPA factor of 2. The diffusion-weighted data of each subject were upsampled to the anatomical resolution (1 mm isotropic). Partial volume estimation (PVE) maps were obtained from the T1-weighted image (1 mm isotropic MPRAGE, TR/TE 6.57/2.52 msec) by using FSL-Fast (Zhang et al., 2001) and registered to the upsampled diffusion-weighted images. PVE were used in the particle filtering tractography algorithm by using anatomical priors (Girard et al., 2014) to provide a better tracking domain as opposed to fractional anisotropy-based mask, where streamline propagation is often prematurely halted in crossing regions (Descoteaux, 2015; Girard et al., 2014; Nimsky et al., 2016; Seunarine and Alexander, 2009; Smith et al., 2012). Fiber orientation distribution functions were obtained from constrained spherical deconvolution (Tournier et al., 2007, 2012), with a spherical harmonic order of 8. Probabilistic streamline tractography was done by using particle filtering tractography (Girard et al., 2014), seeding from the WM/GM interface (Girard et al., 2014; Smith et al., 2012) using 10 seeds per voxel (∼2,000,000 seeds leading to 1,400,000 surviving streamlines). The particle filtering tractography algorithm ensured that streamlines did not terminate prematurely in the WM by the application of a back-tracking rule to allow the tractography algorithm to find alternative pathways. Freesurfer (Fischl et al., 2004) was used to obtain a brain parcellation of 159 regions [Destrieux et al. (2010), see Supplementary Data for label description; Supplementary Data are available online at
where i, j, ∈ lower diagonal of
The result is a probability distribution function that expresses the probability of region A to be connected with all other regions (Ghaziri et al., 2017).
Functional connectivity
Continuous functional recordings were carried out by using a standard EPI sequence (eyes closed). For each run, 108 functional volumes consisting of 35 axial slices were obtained with a 64 × 64 matrix, field of view 220 mm, TR/TE 2730/40 msec, for a voxel size of 3.4 × 3.4 × 4.2 mm3. Preprocessing steps were performed in AFNI (Cox, 1996) and consisted of slice timing and motion correction, spatial smoothing based on non-local means (NLM) denoising (Bernier et al., 2014; Coupe et al., 2008), and band-pass temporal filtering (0.008–0.1 Hz). The use of NLM denoising allowed an improved identification of the BOLD signal in subcortical areas at 1.5 T, thus permitting a better comparison with diffusion data. Preprocessed signals were averaged within each of the aforementioned 159 regions (see Supplementary Data) after upsampling to the T1-space. Finally, the connectivity between each possible pair was assessed by computing their mutual covariance using Pearson's product-moment correlation. Finally, these correlation coefficients were converted to z-scores by using Fisher's r-to-z transform for standardization across subjects.
Inter-subject variability
Variability indices were computed by using the approach proposed by Mueller and associates (2013). The outline of the method is described as follows and was repeated for both FC and SC matrices: For each of the 159 brain regions, an inter-subject variability index (V inter ) was derived by regression of the intra-subject variability from an estimated group-variance index. Methodological details describing the estimations of variability indices are given later and are illustrated in Figure 2.
• Group similarity (
• Intra-subject variability (
• Inter-subject variability (
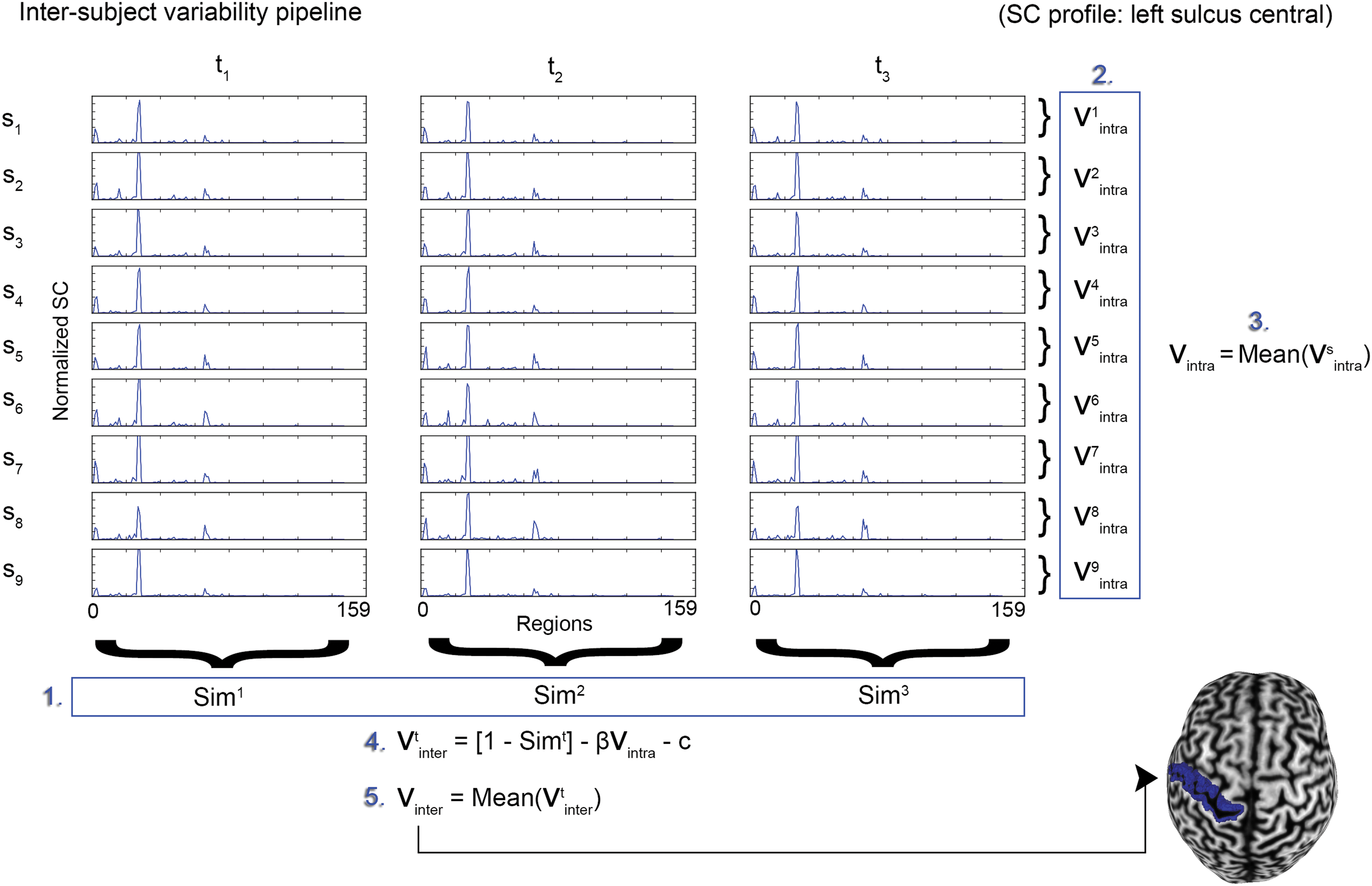
Inter-subject variability pipeline illustrated for a single brain region (left central sulcus, SC profile). Starting with s subjects and t sessions, the similarity between subjects was estimated for each session t by averaging the correlations between corresponding brain regions (1). Next (2), the across session variability was estimated for each subject s by averaging the correlation between homologous regions and subtracting from (1). Intra-subject variability values were then averaged across all subjects (3). Inter-subject variability was estimated for each session by t using ordinary least-squares regression (4), and it was finally averaged to obtain a single value for each region (5). The resulting indices were then back-projected to a brain surface for visualization. The pipeline was repeated independently for both FC and SC matrices. Color images available online at
With tractography, larger regions may be overly connected as opposed to regions with a lower volume or surface area. Thus, to exclude any confounding factors potentially related to volume differences across the 159 regions,
Subsequently, structural and functional V
inter
indices were back-projected to a brain surface for better spatial characterization. Statistical analysis included a direct comparison of functional and structural variability scores using Pearson's product-moment correlation. Finally, mutual regions of low and high variability were determined by isolating regions located in the top and bottom 15% (z = 1) of
Results
Functional variability
Whole-brain
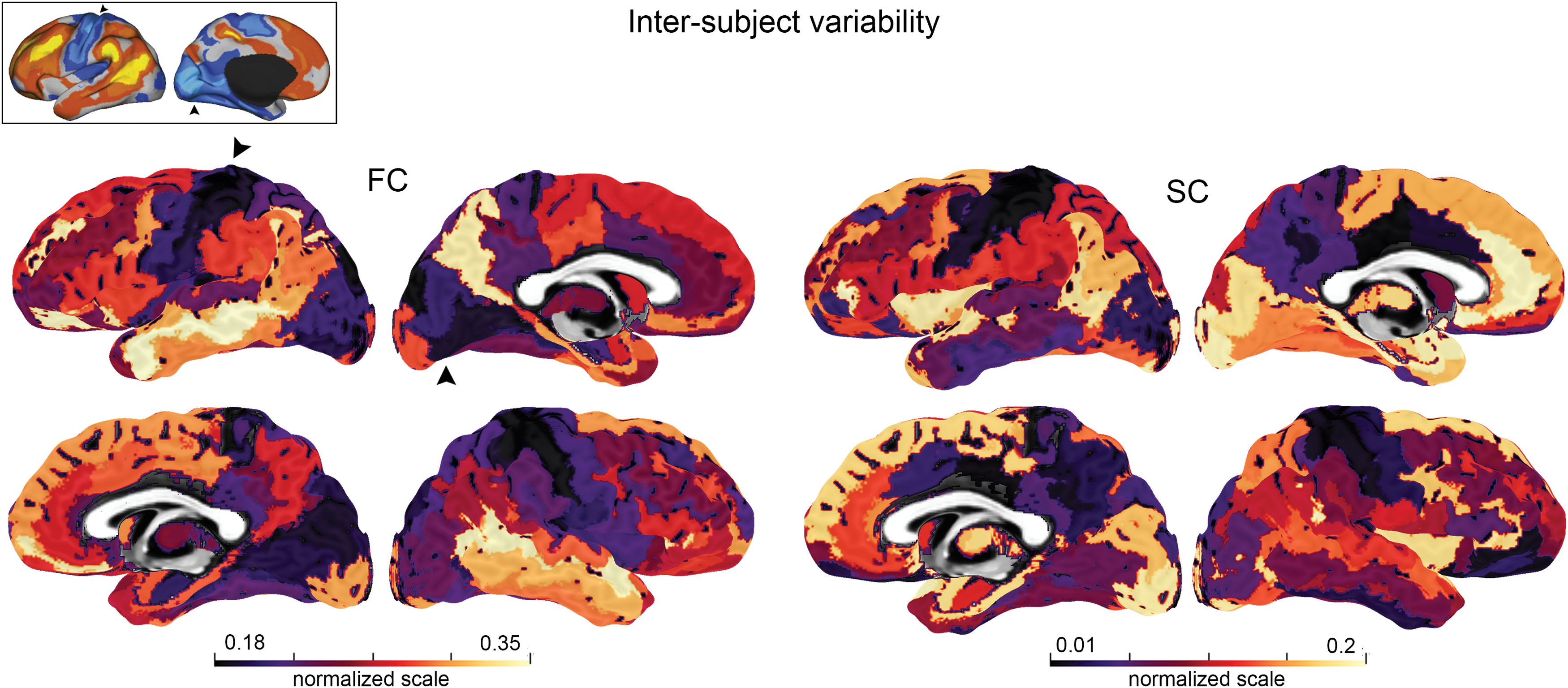
Normalized functional and structural inter-subject variability maps back-projected to a brain surface for visualization. Functional variability (left) is in line with previous work from Mueller et al. (2013) (black arrowheads). Top-left: inter-subject variability mapping (blue: low, yellow: high) reprinted from Mueller and associates (2013), with permission from Elsevier. Regions of low variability (purple–black) are identified in the motor network for both functional and structural analysis. Normalized scale visualization was done by using the Fiber Navigator (Chamberland et al., 2015). Color images available online at
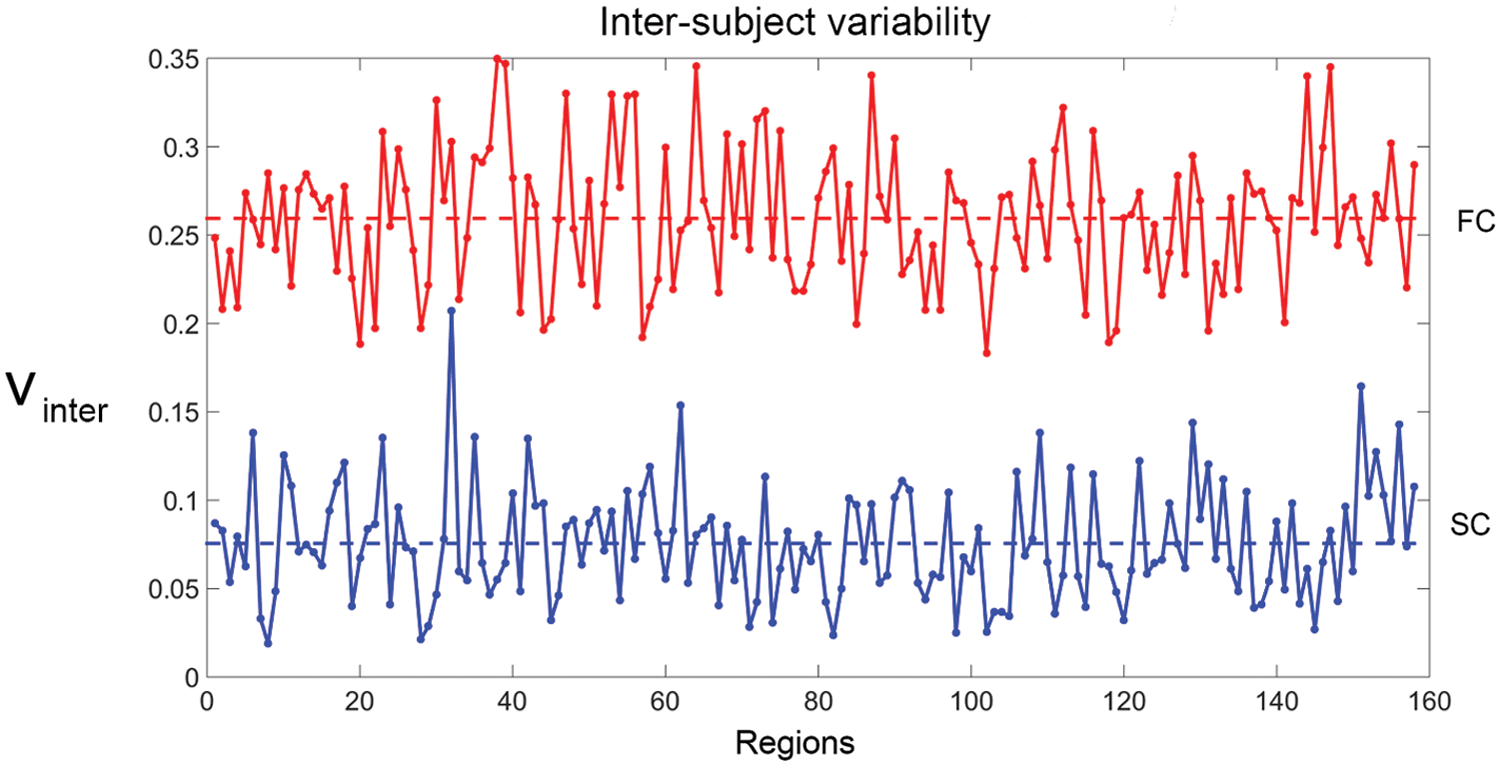
Inter-subject variability indices. Top: functional variability scores associated with each of the 159 regions (mean = 0.25, dotted line). Bottom: structural variability associated with each of the 159 regions (mean = 0.07, dotted line). Note that functional variability is globally higher than structural variability. Supplementary Data can be referred for label indexing. Color images available online at
Structural variability
To ensure the validity of the linear model used in this study (Fig. 2, step 4), the root mean square error (RMSE) was computed at all three time points. A value closer to 0 is preferred and indicates a good estimation of the structural residual maps. Figure 5 shows a validation of the structural residual maps (

Linear relationship between group variability (1-
The resulting
Complementary to the previous results, Figure 4 (bottom) shows a quantitative indexing of structural variability indices (
Structure–function relationship
To assess the potential link between functional and structural variability, variability indices between the two modalities were compared by using two separate approaches. First, region-to-region variations in
Low inter-subject variability
To qualitatively illustrate SC in a region of low variability, Figure 6 presents a group mosaic of the left central sulcus tractography (anterior view). In this example, streamlines show high consistency with multiple inter-hemispheric connections sites across different subjects (e.g., CC, yellow arrowheads). The CST and its projection fibers is also represented across all subjects. Red arrowheads indicate intra-hemispheric similarities. Streamlines were visualized by using Orientation-Dependent Transparency Rendering (Tax et al., 2015) to improve the rendering of hidden structures (e.g., central sulcus region, green isosurface).
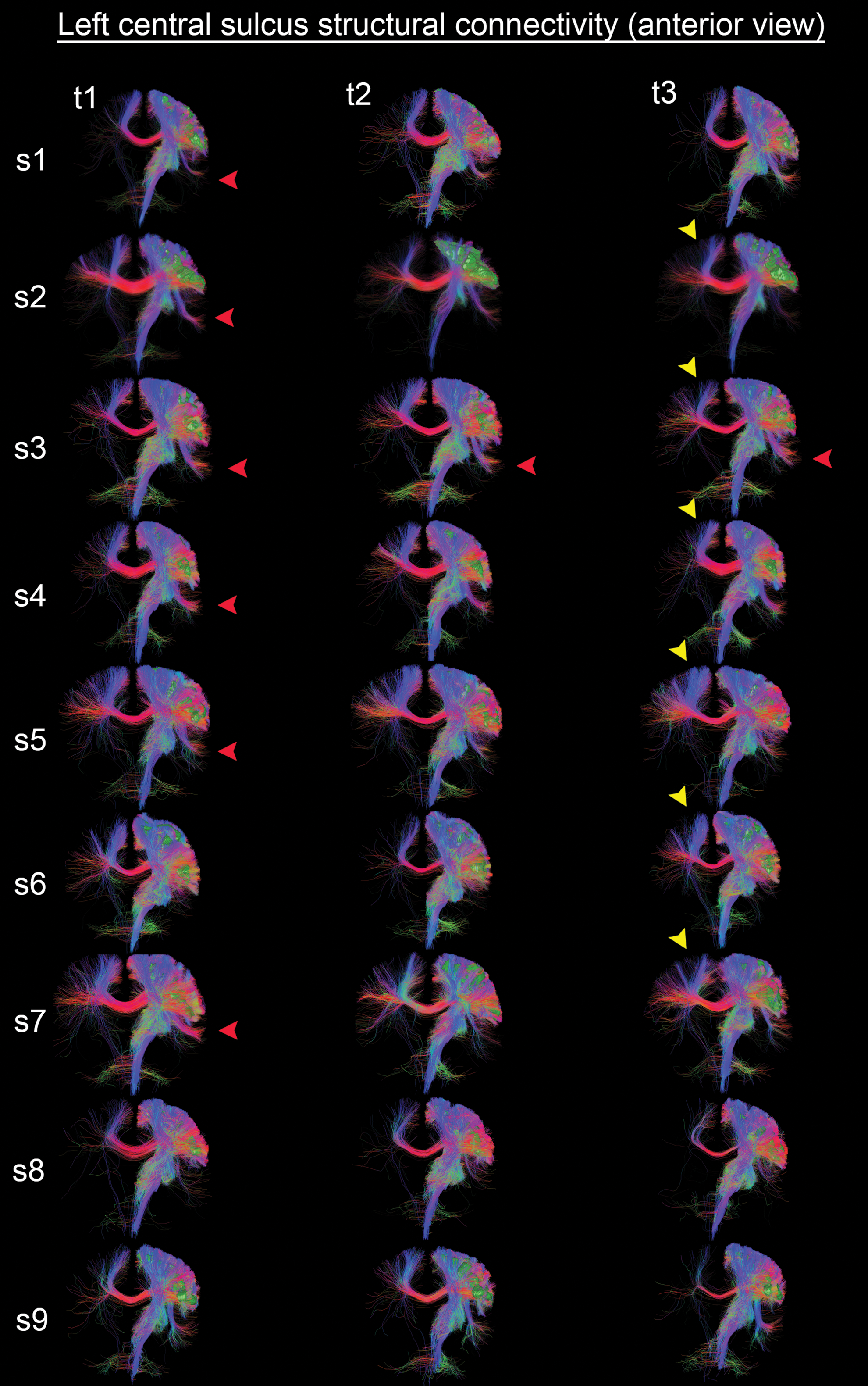
Left central sulcus tractography (anterior view) shows high consistency with multiple commissural connections sites (e.g., corpus callosum, yellow arrowheads). Red arrowheads indicate intra-hemispheric similarities. Streamlines were downsampled by a factor 10 for visualization. Visualization was done by using Orientation-Dependent Transparency Rendering (Tax et al., 2015). Color images available online at
High inter-subject variability
In contrast, Figure 7 shows tractography in a region of higher variability (i.e., left parahippocampal gyrus, lateral view). In this case, streamlines show a larger variability with multiple intra-hemispheric connections sites (e.g., cingulum). One can also notice differences in the orbitofrontal projection of the uncinate fasciculus for multiple subjects. Intra-subject differences are indicated with yellow arrowheads, and inter-subject differences are pointed out with paired red arrowheads. Streamlines were also rendered transparent for a better appreciation of the parahippocampal gyrus isosurface (orange).
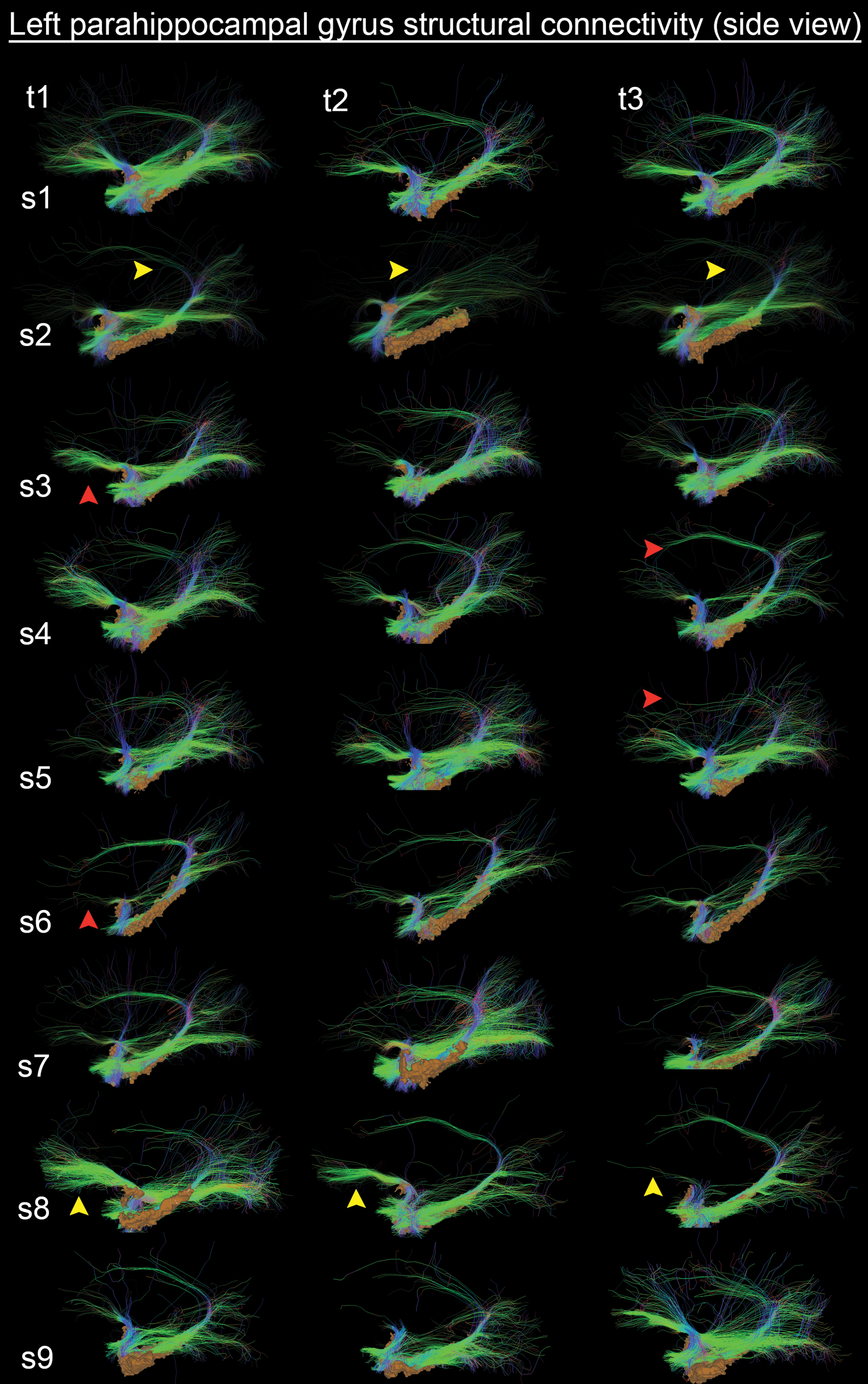
Left parahippocampal gyrus tractography (lateral view) shows a large variability with multiple association connections sites (e.g., cingulum and uncinate fasciculus). Yellow arrowheads indicate intra-subject differences, and red arrowheads indicate inter-subject differences. Streamlines were downsampled by a factor 10 for visualization. Visualization was done by using Orientation-Dependent Transparency Rendering (Tax et al., 2015). Color images available online at
Confounding factors correction
We further explored the effect of region size on structural variability. Figure 8 shows the effect of seeding surface area (left) and number of streamlines (right, normalized) on inter-subject variability scores for all subjects. The results indicated no correlation between
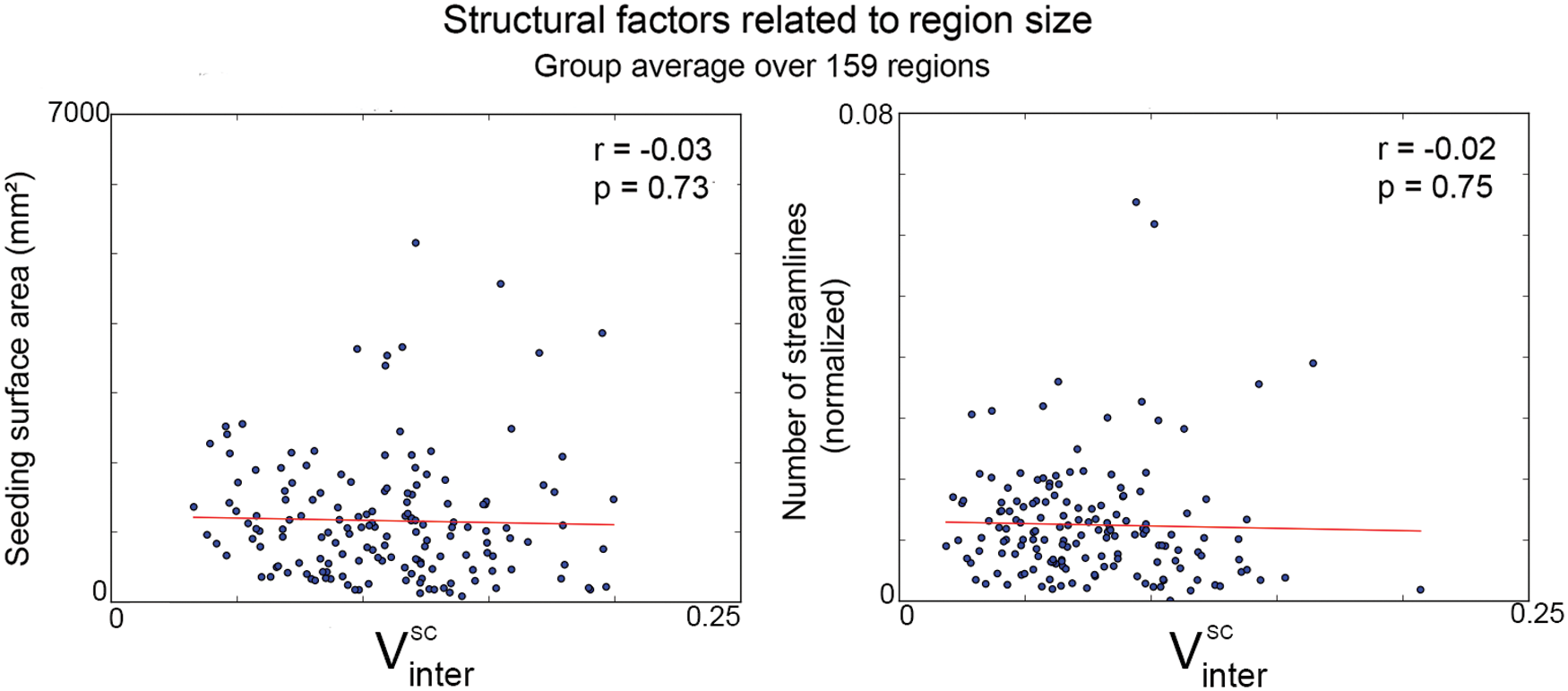
Structural factors related to region size averaged across subjects. Left: Seeding surface area of each region shows no correlation with inter-subject variability (r = −0.03, p = 0.73). Right: Number of streamlines within each region also revealed no link with inter-subject variability (r = −0.02, p = 0.75). Color images available online at
Discussion
A major hurdle in comparing FC with SC is that it is nearly impossible to accurately quantify their respective strengths. To resolve this, we adopted a recently described repeated-measures approach for FC and applied it to tractography-based SC. Rather than correlating the “strength” of FC to that of SC, we computed inter-individual variability in FC of major cortical and sub-cortical areas and compared it with corresponding inter-individual variability in SC. Some areas exhibited mutually low (e.g., primary motor cortex) and high (e.g., language area)
Low inter-subject variability
Inter-subject variability mapping revealed key regions of the brain with low structural and functional variability (Figs. 3 and 4). Specifically, the left and right post-central gyri, left central and post-central sulci were the most reproducible in terms of connectivity overall (all below 0.04 for the SC analysis, and all below 0.22 for the FC analysis). This result indicates that, at least in these sensorimotor regions, inference of brain connectivity using tractography and resting-state functional magnetic resonance imaging (rs-fMRI) is increasingly reliable compared with everywhere else in the brain. These results are also in line with those of Mueller and associates (2013), Gonzalez-Castillo and colleagues (2014), Chen and associates (2015), and Finn and colleagues (2015), who reported that connectivity patterns of motor regions were more consistent than the ones associated to higher order cognitive processes (e.g., memory and language).
When qualitatively looking at the underlying WM motor connections (Fig. 6), tractography of the left sulcus central showed considerable reproducibility in terms of connectivity patterns (e.g., commissural CC connections and cortical fanning of the CST). This low variability is closely in line with previous studies that compared functional variability with evolutionary cortex expansion (Mueller et al., 2013), which also reveals lower surface expansion indices within the motor network. Another group also reported low structural variability in brain gyrification of the motor cortex, which may explain its consistent connectivity profile (Hill et al., 2010).
Indeed, with streamline tractography seeded from the GM/WM interface, the first few voxels in streamline propagation are crucial to determine the final shape and course of the bundle. Likewise, other studies recently demonstrated that long-range connections are impeded by the superficial layers of GM (Burgel et al., 2006; Reveley et al., 2015). This degree of gyrification and layering of the GM may explain why some of the WM fascicles are consistently reproduced across subjects with tractography. Our findings also revealed lower variability within the posterior end of the Sylvian fissure, on the right side of the brain. This lower variability may be due to the fact that this sulcus is one of the earliest developing sulci in the human brain (Chi et al., 1977). Interestingly, the human secondary somatosensory cortex is also located at the ceiling of the Sylvian fissure (Penfield and Jasper, 1954), in line with our aforementioned motor cortex findings.
Low variance was also identified in the right superior occipital gyrus, where multiple WM pathways connect. Visual inspection of this area revealed the following underlying major WM bundles: the inferior longitudinal fasciculus, optic tracts, and splenium of the CC. For consideration, changes of cortical thickness and corresponding WM microstructure during brain development can also play an important role in defining individual brain regions, as these regions evolve at a different rate (Jeon et al., 2015). For example, the optic radiation (OR) is a well-understood fascicle with the primary function of transmitting visual information to the visual cortex. It is also among the few primary WM fiber pathways to attain complete myelination in the early developmental stages (Kinney et al., 1988). It is generally believed that the OR projects to the calcarine fissure (in the primary visual cortex, V1), though it has been shown recently that it may reach V2 and V3 as well (Alvarez et al., 2015; Renauld et al., 2016; Chamberland et al., 2017). This may indicate that after a certain age, this fascicle is already fully developed and its connectivity profile is less likely to vary across healthy subjects. In this study, potential confounding factors related to age difference (Mattay et al., 2002; Nenonen et al., 2015) were reduced by the inclusion of a homogeneous subject sample (i.e., nine young adults, mean age 25 years ±3.1; range 21–30).
High inter-subject variability
Conversely, higher variability profiles are noticeable on the medial aspect of the temporal lobe. Specifically, the parahippocampal gyrus and the subcallosal gyrus (BA25) (two regions that are part of the limbic system) were among the highest varying regions of this study, both functionally and structurally. The parahippocampal gyrus plays an important role in memory retrieval (Whittingstall et al., 2014) and is composed of multiple interdigitating association fascicles (Schmahmann and Pandya, 2006; Yogarajah et al., 2008) such as the uncinate fasciculus, the temporal projections of the cingulum and of the fornix, the medial and inferior longitudinal fasciculi, and the inferior fronto-occipital fasciculus (Catani et al., 2003; Catani and Thiebaut De Schotten, 2012). This highly complex configuration can potentially lead to a widespread connectivity spectrum, as denoted by our variability mapping results. For instance, the orbitofrontal projections of the uncinate fasciculus were either absent from this region or partially represented across multiple subjects.
Interestingly, variability indices were also the largest in the temporo-parietal cortex for both FC and SC analysis. Notably, the sulcus intermedius primus of Jensen (bilateral) revealed signs of higher inter-subject variability for both structural (left: 0.11, right: 0.14) and functional (left: 0.33, right: 0.30) analysis. This region marks the boundary between the supramarginal and angular gyri (Kiriyama et al., 2009) and closely overlaps with Wernicke's area. In addition, the right superior temporal gyrus (planum temporale) is a cortical region of the temporal lobe also overlapping Wernicke's area. Studies also revealed a preferential leftward development of this cortical area (Geschwind and Levitsky, 1968; Steinmetz, 1996). Even though this area is most often found on the left side of the brain, a lack of functional specificity on the contra-lateral hemisphere may also lead to a higher inter-subject variability, indicating potential variable WM connectivity. Our results also indicated higher variability within the left superior temporal sulcus. This unilateral profile is closely related with the brain lateralization of higher cognitive functions such as language [e.g., Broca's area Catani and Mesulam (2008); Lurito and Dzemidzic (2001); Vassal et al. (2016)]. Other structure–function relationships studies have previously reported a lateralization of FA values and number of streamlines within the arcuate fasciculus (underlying WM pathway involved with language) (Powell et al., 2006; Vernooij et al., 2007). Such structural asymmetries may also be related to a highly specific functional lateralization. The group of Vassal and colleagues (2016) also reported leftward lateralization of the arcuate fasciculus using diffusion tensor imaging, and major inter-hemispheric differences in streamlines connecting Broca's area and Wernicke's area were noticed in more than half of their subjects.
SC confounding factors
Our choice of local modeling of the diffusion data worked toward reducing potential connectivity bias by being robust to crossing and kissing regions. Indeed, it is generally accepted by the community that HARDI-based techniques can overcome the limitations of diffusion tensor imaging (DTI) tractography by resolving the many crossing fiber regions in the brain (Descoteaux, 2015; Farquharson et al., 2013; Neher et al., 2015; Nimsky et al., 2016; Seunarine and Alexander, 2009; Tournier et al., 2011). Possible structural confounding factors of this study can be related to the construction of SC matrices (Jones et al., 2013; Smith et al., 2015). For instance, a region expressing a higher volume (mm3) may also show higher reproducibility due to an over-representation in the number of underlying streamlines. Here, potential bias toward seed density was mitigated by initiating tractography from the WM/GM interface (Girard et al., 2014) instead of using the whole WM volume. In addition, SC matrices were also normalized across regions and subjects to account for various streamline counts and inter-subject variability values were also corrected for seeding surface area (mm2) to remove any remaining bias toward brain region size. This step was performed independently of the variability pipeline to ensure matching processing between rs-fMRI and dMRI connectivity matrices. As a result, no significant correlation between inter-subject variability indices and region surface area (r = −0.03, p = 0.73) was observed. Further, the number of streamlines in each region showed no significant correlation with variance measures (r = −0.02, p = 0.75, Supplementary Fig. S3). For these reasons, the heterogeneous size of the parcellated brain regions cannot explain our
Weak link between functional and structural inter-subject variability
As described earlier, certain brain regions showing the lowest
One possible explanation for this result is that the bulk of the reconstructed streamlines emanating from the motor cortex are made up of either the CC or the CST, with little to no terminations observed in other cortical areas. FC, on the other hand, may be visible in these areas due to indirect anatomical connections, thus explaining its higher inter-individual variability as shown in Supplementary Figure S4. Even though FC of the left central sulcus showed similar patterns between different subjects, individual differences are identifiable in the cerebellum and are supported by direct anatomical pathways. However, SC revealed no direct link between parahippocampal connectivity and the left central sulcus. Indirect structural pathways may, therefore, play a key role in explaining the mismatch between
Another possible explanation is that diffusion- and BOLD-based measures of brain structure and function, respectively, are sensitive to different sources of noise. For instance, BOLD signal correlations in areas that are highly vascularized may be artificially inflated compared with other regions (Curtis et al., 2014; Kazan et al., 2016; Vigneau-Roy et al., 2014). On the other hand, blood vessels may interfere with diffusion measures, potentially leading to an incorrect estimation of SC due to intravoxel incoherent motion (Le Bihan, 2008). Taken together, the sum of all those sources of variability effectively contributes toward establishing inter-subject variability.
Clinical implications
Fingerprint patterns of FC can be used to identify subjects across sessions and individuals (Finn et al., 2015), indicating that the topology of the brain substantially differs between individuals. Assuming that patterns of spatial heterogeneity are also present in SC, our finding is also of clinical importance with the recent advances in surgical approaches based on connectomics (Griffa et al., 2013; Meola et al., 2016; Yu et al., 2016). For instance, a region of inter-individual variability can have potential influence on preoperative planning as the spatial localization of this area may vary from one subject to another. As of today, rs-fMRI imaging protocols are often used in clinical studies to guide surgical gesture. In addition, tractography has been shown to be valuable for surgical planning, and neurosurgeons and radiologists are mostly convinced of the importance of achieving an accurate delineation of WM pathways using tractography (Duffau, 2005; Leclercq et al., 2010; Nimsky et al., 2016). Defining accurate boundaries is critical in surgery targeting frontal and temporal regions (e.g., space-occupying lesion, temporal lobe epilepsy), as these areas exhibit higher variability between subjects. With the advent of personalized medicine (Wang et al., 2015), we believe that such a tailored mapping may improve decision making and is likely to impact pre-operative planning approaches, where capturing the idiosyncrasies of individuals is paramount.
Conclusion
In this study, we generated brain functional and structural inter-subject variability maps by taking into account the intra-subject variability using a test–retest approach. With this approach, we showed that structural inter-subject variability was globally lower than functional variability. Our approach also revealed that variability indices were smallest in motor areas for both functional and structural analysis, indicating proper reproducibility within this region. Our results also indicate that functional and structural variability profiles were less consistent near the memory and language areas of the brain. These findings suggest that, at least in these regions, functional inter-subject variability can be explained by the structural organization of the brain. In other words, stable FC does not always imply stable SC for the rest of the brain. Altogether, this study highlights a different aspect of brain connectivity analysis, by not only exploring the link between FC and SC but also investigating the variability between them.
Footnotes
Acknowledgments
The authors would like to acknowledge the funding agencies that have supported this research: Natural Sciences and Engineering Research Council of Canada (NSERC) Discovery Grants, QBIN (Quebec Bio-Imaging Network), and the FMSS graduate scholarship program. Main author M.C. was supported by the Alexander Graham Bell Canada Graduate Scholarships-Doctoral Program (CGS-D3) from the NSERC at the time.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.