
Abstract
To investigate resting-state functional connectivity of lupus patients and associated subgroups according to the ACR NPSLE case definitions (ACR ad hoc). In addition, we investigated whether or not the observed alterations correlated with disease duration, the systemic lupus erythematosus (SLE)-Disease Activity Index-2000 (SLEDAI-2k), and Systemic Lupus International Collaborating Clinical/ACR organ damage index (SDI)-scores. Anatomical 3T magnetic resonance imaging (MRI) and resting-state functional MRI were performed in 61 female lupus patients (mean age = 37.0 years, range = 18.2–52.0 years) and 20 gender- and age-matched controls (mean age = 36.2 years, range = 23.3–52.2 years) in conjunction with clinical examination and laboratory testing. Whole-brain voxelwise functional connectivity analysis with permutation testing was performed to extract network components that differed in lupus patients relative to healthy controls (HCs). Lupus patients exhibited both inter- and intranetwork hypo- and hyperconnectivity involving several crucial networks. We found reduced connectivity within the default mode network (DMN), the central executive network (CEN), and in-between the DMN and CEN in lupus patients. Increased connectivity was primarily observed within and between the sensory motor network in lupus patients when compared to HCs. Comparing lupus patients with and without neuropsychiatric symptoms, hypoconnectivity was more pronounced in the group with neuropsychiatric complaints. The functional connectivity of SLE patients was both positively and negatively correlated to duration of disease. We conclude that SLE patients in general and neuropsychiatric SLE patients in particular experience altered brain connectivity. These patterns may be due both to direct neuronal damage and compensatory mechanisms through neuronal rewiring and recruitment and may partly explain neuropsychiatric symptoms in SLE patients.
Introduction
S
Neuropsychiatric SLE (NPSLE) has been classified as a complex neurological disorder and is categorized by 19 diagnostic criteria set forth by the American College of Rheumatology (ACR) (American College of Rheumatology Ad Hoc Committee on Neuropsychiatric Lupus Nomenclature, 1999), based on the presence of SLE and neuropsychiatric conditions. Manifestations of NPSLE range from mild cognitive impairment and mood disorders to cerebral microangiopathy, psychosis, epilepsy, and stroke (Unterman et al., 2011).
The pathophysiology of NPSLE is probably diverse and may include production of autoantibodies, intrathecal productions of proinflammatory cytokines that disintegrate the blood–brain barrier (Gulati et al., 2016), vascular compromise by microangiopathy, chronic diffuse ischemia, thromboembolism, and atherosclerosis. There is also evidence that hormonal, hereditary, and environmental factors contribute to the clinical manifestations of the disease (Bertsias and Boumpas, 2010; Lisnevskaia et al.; Rahman and Isenberg, 2008; Tsokos, 2011). There is still no gold standard clinical or laboratory procedure for predicting who is at risk of developing neuropsychiatric symptoms of the disease, and diagnosing NPSLE is still a matter of exclusion of all other explanation models.
Magnetic resonance imaging (MRI) is the standard tool for evaluating suspected brain lesions in patients with SLE. Brain atrophy and white matter (WM) hyperintensities are the most frequent findings in SLE patients, but no obvious relationship between disease and WM lesion load has been established (Ainiala et al., 2005; Sarbu et al., 2015; Sibbitt et al., 2010; Toledano et al., 2013). Large vessel infarcts are one of the most debilitating complications of NPSLE and are found in up to 15% of NPSLE patients.
Previous studies utilizing MR spectroscopy (Cagnoli et al., 2013; Sundgren et al., 2005) and diffusion tensor imaging have demonstrated metabolic alterations and microstructural changes in the WM, as well as reduced WM tract integrity in SLE and NPSLE patients compared to healthy subjects (Schmidt-Wilcke et al., 2014; Shastri et al., 2016; Hughes et al., 2007).
In general, the findings are nonspecific and are not proven effective markers neither of disease activity nor in terms of explaining cognitive dysfunction.
Functional MRI (fMRI) provides an objective method for studying large-scale neuronal networks of the brain. The method records blood–oxygen-level-dependent (BOLD) signals—a measure of neuronal metabolism and generally considered a proxy to neuronal activity (He and Liu, 2012). Studies of BOLD response in NPSLE patients have shown extended areas of activation and increased connectivity in the frontoparietal cortex (Fitzgibbon et al., 2008; Mikdashi, 2016). These changes have also been shown to correlate with the SLE-Disease Activity Index-2000 (SLEDAI-2k score) (Hou et al., 2013).
A relationship between task-based activation (i.e., subjects performing an explicit task) of neuronal networks and disease duration has been exposed, in which increased neuronal activation was observed in areas associated with working memory. In addition, it was shown that increased whole-brain activation was associated with short-term disease, whereas long-term disease was associated with increased neuronal damage and decreased activation (Mackay et al., 2011).
fMRI studies of spontaneous BOLD patterns in the absence of external stimuli have led to the identification of several so-called resting-state networks, which exhibit sustained, high intraregional functional connectivity (synchronous BOLD oscillations). Some of these networks that have gained particular interest due to their particular stability and reproducibility are briefly outlined below. The default mode network (DMN) is a large-scale intrinsic connectivity network, known to be involved in retrieval of self-referential memory, introspective activities, and mental processing (Menon, 2011). By contrast, the DMN tends to deactivate in stimuli-driven cognitive settings. The central executive network (CEN) fulfils a crucial role with its executive function, working memory, and in decision-making (Menon, 2011). The ventral attention network (VAN, also referred to in the literature as the salience network) is pivotal in task switching between the DMN and CEN and is believed to be involved in attention and switching between internal and external events (Bressler and Menon, 2010; Menon, 2011). In the context of SLE, it is worth noting that the VAN encompasses the anterior insula (AI). AI dysfunction is believed to be a core feature of anxiety and pain (Bressler and Menon, 2010; Menon, 2011).
Some evidence of early changes in resting-state functional connectivity (i.e., studying the spontaneous fluctuation of BOLD signals when the subject is not performing an explicit task) has also been exposed, highlighting regional attenuation of activity in DMN and the cerebellum in non-NPSLE, which was found to be positively correlated to the SLEDAI-2k score (Lin et al., 2011).
This reorganization of intracerebral functional connectivity could be a potential coping mechanism triggered by neuronal damage, attempting to maintain function by engaging extracortical pathways.
Overall, only few resting-state (rs)-fMRI studies have been performed and the vast majority of these focused exclusively on NPSLE patients.
We hypothesize that rs-fMRI may exhibit significant alterations in brain connectivity, which precede clinically overt neuropsychiatric symptoms. Although longitudinal studies are needed to provide more conclusive evidence, the size and broad spectrum of our cohorts allow for some interesting and relevant interpretations.
The aim of our study was to investigate group-level changes in intracerebral networks (as gauged by rs-fMRI) in SLE patients with and without neuropsychiatric symptoms. Furthermore, we investigated the association of connectivity changes with Systemic Lupus International Collaborating Clinical/ACR organ damage index (SDI), SLEDAI-2k score, and disease duration.
Materials and Methods
Cohort demographics
In this cross-sectional study, 71 consecutive female SLE patients with and without NP symptoms (mean age = 36.6 years, range = 18.2–52.0 years) and 26 age-matched female healthy controls (HCs) (mean age = 38.1 years, range = 23.3–52.2 years) were initially recruited. The local ethics committee approved the study and informed consent was obtained from all subjects before inclusion. All SLE patients fulfilled at least 4 of the American college of Rheumatology classification criteria for SLE (Tan et al., 1982).
Inclusion criteria for all studied subjects were female gender and age between 18 and 55 years. Second, wave exclusion criteria were left-handedness, abnormal cognitive test scores, and major depression (defined as Montgomery Asberg Depression Rating Scale [MADRS]-score >34).
Exclusion criteria for HCs were male gender, age <18 or >55, major depression (defined as MADRS-score >34), left-handedness, autoimmune disease, or any previously diagnosed neurological condition.
From the original cohort, 10 SLE patients (1 due to previous temporal lobe resection, 3 due to not being able to fulfill the MRI examination, 2 due to failure to meet rs-fMRI quality assurance criteria [see heading Resting-state fMRI in the Materials and Methods section], 3 due to being predominantly left-handed, and 1 due to severe depression) and 6 HC subjects (3 due to abnormal cognitive test scores, 1 due to other autoimmune diseases, 1 due to dyslexia, and 1 due to MRI susceptibility artifacts) were excluded.
Given these exclusions, the final cohort consisted of 61 female SLE patients (mean age = 37.0 years; range = 18.2–52.0 years) and 20 HCs (mean age = 36.2 years; range = 23.3–52.2 years).
Patients were classified as non-NPSLE (n = 25) or NPSLE (n = 36) according to the ACR NPSLE case definitions. Cohort demographics and results are presented in Table 1.
Demographic information is annotated for HCs and SLE patients as well as for subgroups of SLE patients analyzed in the current study (i.e., non-NPSLE and NPSLE). p Values are described for each subset of group comparisons when possible. Level of significance was p < 0.05. There was a significant difference in FSS-score and MADRS score between SLE and HC subjects. Only FSS was significantly increased in the NPSLE subgroup when compared to non-NPSLE patients.
Age when entering the study (years).
Disease duration (time since diagnosis upon time when included in the study in years).
Disease activity index (SLEDAI-2k).
Chronic organ damage index (SDI).
Daily dose of glucocorticoids (recalculated into equivalent doses of prednisolone).
Previous treatment.
Ongoing treatment.
Fatigue index (FSS).
Depression score (MADRS).
DMARDs, disease-modifying antirheumatic drugs; FSS, fatigue severity scale; HC, healthy control; MADRS, Montgomery Asberg Depression Rating Scale; NPSLE, neuropsychiatric SLE; SLE, systemic lupus erythematosus; SLEDAI-2k, SLE-Disease Activity Index-2000.
After providing informed consent, all subjects completed self-assessment forms, including the Fatigue Severity Scale (FSS) (Krupp et al., 1989) and the MADRS (Montgomery and Asberg, 1979). Subjects who were classified as suffering from severe depression were excluded from further analysis. Present and previous glucocorticoid (GC) treatment as well as daily doses of prednisolone when entering the study was registered. The same was true for ongoing and previous treatment with antimalarial medication, disease-modifying drugs (DMARDs), and chemotherapy.
All patients underwent rheumatologic and standardized neurologic clinical examination, including assessment of disease activity measured with the SLE-Disease Activity Index-2k (SLE-DAI-2k) (Bombardier et al., 1992) and accumulated organ damage measured with the Systemic Lupus International Collaborating Clinical/ACR organ damage index (SDI) (Gladman et al., 1996, 2002).
Magnetic resonance imaging
All subjects underwent pre- and postcontrast-enhanced structural MRI and rs-fMRI on a 3T MR scanner (Siemens MAGNETOM Skyra, Erlangen, Germany). The imaging protocol included among others the following sequences: T2W-TSE (33 transversal slices TE/TR = 100/6870 ms), T2W-FLAIR (33 transversal slices, TE/TR/TI = 81/9000/2500 ms), and 3D T1W-MPRAGE (1 mm3 isotropic voxels, TE/TR/TI = 2.54/1900/900 ms). Intravenous contrast administration of 0.2 mL/kg of gadolinium-DOTA (Dotarem®; Gothia Medical/Guerbet) was given to all subjects.
Scans were visually evaluated, by one specialist in neuroradiology and one PhD student in neuroradiology independently, and scanned for morphological abnormalities. Described findings were presented after consensus by the reviewers. Global lesion burden, evaluated in the form of WM T2/FLAIR hyperintense lesions, was defined as mild (1–5 discrete WM lesions), moderate (5–10 discrete WM lesions), severe (>10 discrete WM lesions), or very severe (confluent WM lesions).
Resting-state fMRI
Spontaneous BOLD oscillations in the absence of external stimuli were imaged with a gradient-echo echo-planar pulse sequence (eyes closed, in-plane resolution = 3 × 3 mm2, slice thickness = 3 mm, TE/TR = 30/1850 ms, flip-angle = 90°, 256 dynamic scans).
rs-fMRI data preprocessing
Data preprocessing was performed using software packages FMRIB software (FSL) (Jenkinson et al., 2012), Analysis of Functional NeuroImage (AFNI) (Cox, 1996), and Advanced Normalization Tools (ANTs) (Avants et al., 2014). Anatomical processing involved skull stripping, segmentation of WM/gray matter (GM)/cerebrospinal fluid (CSF), and normalization to MNI152-template space (Grabner et al., 2006). Dropping the first five frames in anticipation of steady state, functional data were bulk motion and slice timing corrected, furthermore nuisance regressed using the WM/CSF average signal, 6 components of physiological noise (Behzadi et al., 2007), 24 motion parameters (Friston et al., 1996), and linear/quadratic trends. Finally, the functional data were transformed to MNI space using the nonlinear normalization warp calculated for the anatomical scan. Frames causing outliers (75th percentile +1.5 times interquartile range) in total signal frame-to-frame variation were censored (Power et al., 2012) (to avoid unwanted aliasing effects, outlier frames were interpolated over before band-pass filtering, after which they were deleted). The signal was band-pass filtered to 0.01–0.1 Hz, further discriminating against scanner drift and physiological noise.
The impact of subject motion was assessed excluding subjects with mean/maximum framewise displacement (Power et al., 2014) exceeding 0.5/3.0 mm (no subjects exceeded these thresholds). As an extra precautionary step, the voxel-to-voxel BOLD-signal correlations across the whole brain (including GM, WM, and CSF) were calculated and summed. Outliers (identified using Thompsons' tau with p < 0.01) in this measure likely originate in a motion-induced global signal confound capable of eluding conventional motion detection (He and Liu, 2012). Two subjects were strong outliers in this measure and subsequently removed.
Statistical analysis
Analysis of variance (ANOVA) was used to compare all groups for differences in age, FSS-index, MADRS, disease duration, and current dose of GC. Previous and current use of GCs, antimalarial treatment, and DMARDs were all evaluated as dichotomous variables using the chi-square test for group comparisons. All statistical computations of demographic data were performed utilizing SPSS (SPSS for windows, IBM, version 24.0, Armonk NY).
Network components of differing connectivity in 5 × 5 × 5 mm3 voxelwise, whole-brain connectivity networks (correlation between voxel BOLD time series) were calculated with algorithms similar to the network-based statistic (NBS) algorithm (Zalesky et al., 2010), as were the components correlating with patient data (see Supplementary Data for a detailed description of these algorithms; Supplementary Data available online at
Results
There was no significant difference in age between patients and HCs.
According to the ACR NPSLE case definitions, two groups were identified: SLE without NP symptoms (non-NPSLE, n = 25) and SLE with NP symptoms (NPSLE, n = 36).
There was no difference in age, disease duration, or previous/ongoing treatment between the non-NPSLE and NPSLE subgroups. SDI-score and SLEDAI-2k-score were mild or moderate in most patients and no differences were identified between subgroups.
Morphological changes in anatomical MR images
No significant differences in ischemic lesions, microbleeds, or contrast-enhanced lesions could be found between any of the groups (HC, non-NPSLE, NPSLE) and no lesions were identified among HCs (Table 1).
Resting-state fMRI
Group-level network components of altered connectivity
Significant network components containing both hypo- and hyperconnectivity parts were found when comparing HCs to non-NPSLE and NPSLE, but no component was found between non-NPSLE and NPSLE. Similarly, the SLE group as a whole had a significantly different network component relative to HC with many similarities with the HC-NPSLE/non-NPSLE contrasts. Narrowing the scope to study effects more closely related to neuropathology, we focused our attention on the latter (see Supplementary Figure S2 for HC-SLE contrast).
The significantly different network components comprise a connected set of links with both intra- and internetwork connectivities as summarized in Figure 1. The so-called connectograms provide an overview of the significant network components by showing the number of affected links within and between the large-scale resting-state networks. The NPSLE group exhibited a much greater degree of hypoconnectivity, particularly within the DMN, and also within the CEN. Moreover, there was crucial internetwork hypoconnectivity, such as DMN-CEN/VAN, VAN-visual network (VIN), and multiple connections, with the frontotemporal network (FTN).
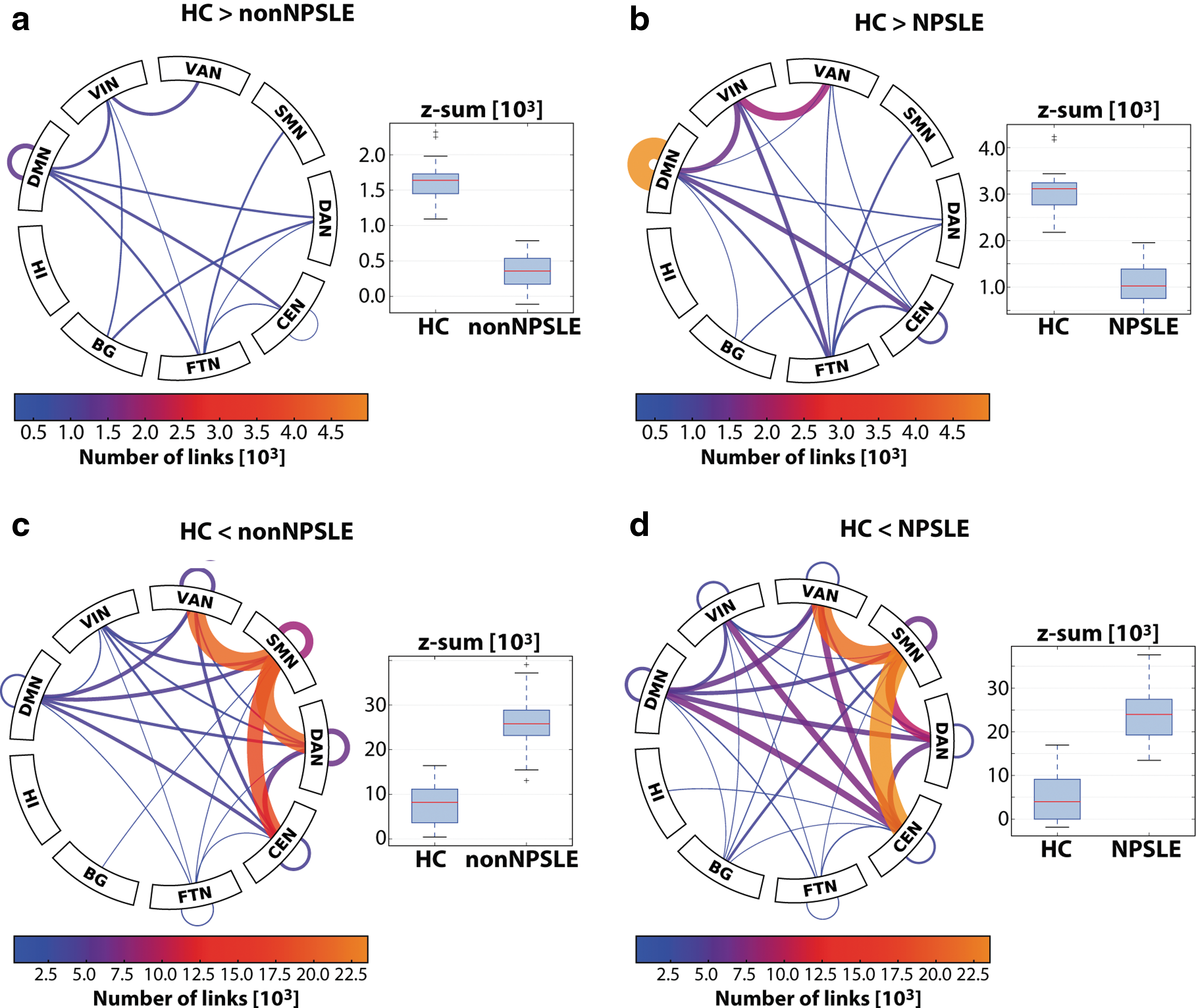
Connectograms showing the number of links between and within resting-state networks and subcortical structures (BG, HI) for hypo- and hyperpart of significantly differing network components in non-NPSLE and NPSLE relative to HC. The accompanying boxplots show the distributions of the sum of functional connectivity (Fisher-z values of constituent links) on the displayed connectogram for the groups in question. The range of the connectogram scales has been matched for hyper- and hypoparts to facilitate comparison between the two different group tests. Both contrasts were dominated by hyperconnectivity converging on SMN
The hyperconnectivity contrasts comprised a much larger set of links than their hypoconnectivity counterparts (Fig. 1c, d), heavily focused on the sensory motor network (SMN). We found many internetwork links with increased connectivity, most notably the SMN-VAN/dorsal attention network (DAN)/CEN with concomitant intranetwork hyperconnectivity. DMN was hyperconnected with all of these networks in both contrasts. In general, hyperconnectivity was more widespread in the HC-NPSLE contrast, although the HC-non-NPSLE contrast had a greater SMN-DAN contribution.
Note that hyperconnectivity may contain elements of decreased anticorrelations, accounting for some of the increased connectivity between DMN and the heavily affected SMN, VAN, DAN, and CEN (Fig. 1c, d), overlapping with task-positive regions. The box plots of Figure 1a and b seem to indicate that loss of correlation is the dominant contribution, in particular for NPSLE.
In what follows, we will perform a more in-depth analysis of hypo- and hyperconnectivity components in terms of the resting-state networks, during the course of which the connectograms of Figure 1 will serve as a useful road map. Furthermore, quantitative information in the form of the z value sums on the whole component and relevant subcomponents is provided for the groups in question (see above definition of summary statistic). For the group tests, the box plots show the subjects' z value sums on the pooled HC-NPSLE and HC-non-NPSLE contrast. Below, the detailed analysis of selected subcomponents showing the most relevant parts of the components is based on their dominance in the contrast and their neuropathological relevance.
Hypoconnectivity
The default mode network
Both non-NPSLE and NPSLE had reduced intra-DMN connectivity relative to HC (Figs. 1a, b, and 2a) with qualitative similarities. In NPSLE, however, the DMN part of the significantly reduced connectivity component comprised a substantially larger set of links. Figure 2a shows the detailed hypoconnectivity within the DMN for the HC-NPSLE and HC-non-NPSLE components.
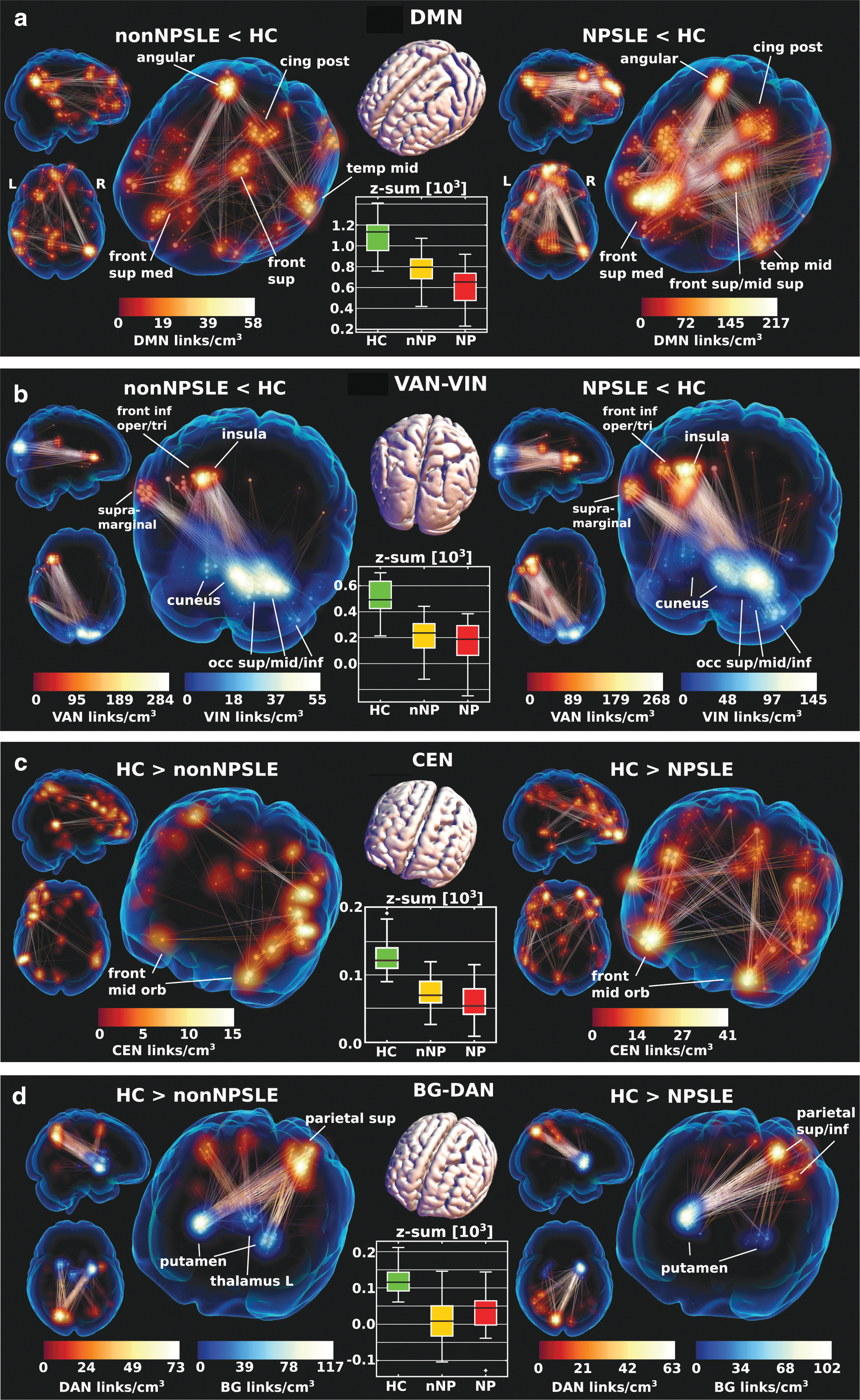
Link plots of dominant hypoconnectivity in non-NPSLE (left) and NPSLE (right) relative to HC within DMN
The hypoconnectivity was most severe between the posterior DMN nodes: right angular gyrus and posterior cingulum (with some spillover into precuneus), and the anterior parts of DMN: primarily frontal medial superior gyrus, but also covering superior frontal and superior frontal middle gyrus. Moreover, a cluster in the left middle temporal gyrus had hypoconnectivity with the anterior and posterior DMN in both contrasts.
The DMN-CEN hypoconnectivity consisted largely of parts adjacent to the regions already highlighted in intra-DMN, in large part, owing to spillover across network borders, which cannot be precisely defined (partly an effect of the severe dimensionality reduction entailed in network identification). Notably, a right-hand counterpart to the frontal superior left cluster in Figure 2 a and b (cross-connecting to left temporal middle gyrus) appears in DMN-CEN. Likewise, the DMN-FTN consisted of FTN close to inferior frontal DMN parts with hypoconnection to the prominent posterior intra-DMN nodes (angular gyrus and posterior cingulum).
There was also reduced DMN-VIN/DAN connectivity involving DMN areas adjacent to insula in VAN, a pattern very similarly but much more strongly expressed in the VAN-VIN contrast (See The ventral attention network).
The ventral attention network
Both contrasts exhibited a sharply focused reduction in connectivity (Fig. 2b) between the left insula (extending laterally over frontal inferior operculum/triangularis) and right VIN (primarily cuneus, but also parts of occipital middle and inferior gyrus). Although the HC-non-NPSLE/NPSLE patterns were qualitatively similar, the number of links with reduced VAN-VIN connectivity was greater in HC-NPSLE expanding over a greater volume.
The central executive network
The relatively weak intra-CEN hypoconnectivity part of the contrast was clearly stronger in HC-NPSLE (Fig. 2c) with bilateral reductions in connectivity involving primarily frontal middle orbitus with scattered clusters supero-anteriorly and the temporal lobe.
Dorsal attention and interconnected networks
Hypoconnectivity was observed in both SLE subgroups between DAN and BG, specifically linking left posterior precuneus with right posterior putamen in HC-NPSLE and to a greater extent both left and right putamen in HC-non-NPSLE (Fig. 2d).
Hyperconnectivity
The sensory motor and interconnected networks
The connectograms (Fig. 1c, d) revealed striking hyperconnectivity in both groups converging on the SMN and projecting to CEN, VAN, and DAN, primarily on the right-hand side (Fig. 3a–c), but SMN-DAN was not as pronounced for NPSLE. There was a sharp maximum in SMN left postcentral gyrus connecting primarily to supplementary motor area/cingulum mid in VAN, right frontal inferior operculum in CEN, and right frontal middle gyrus in DAN.
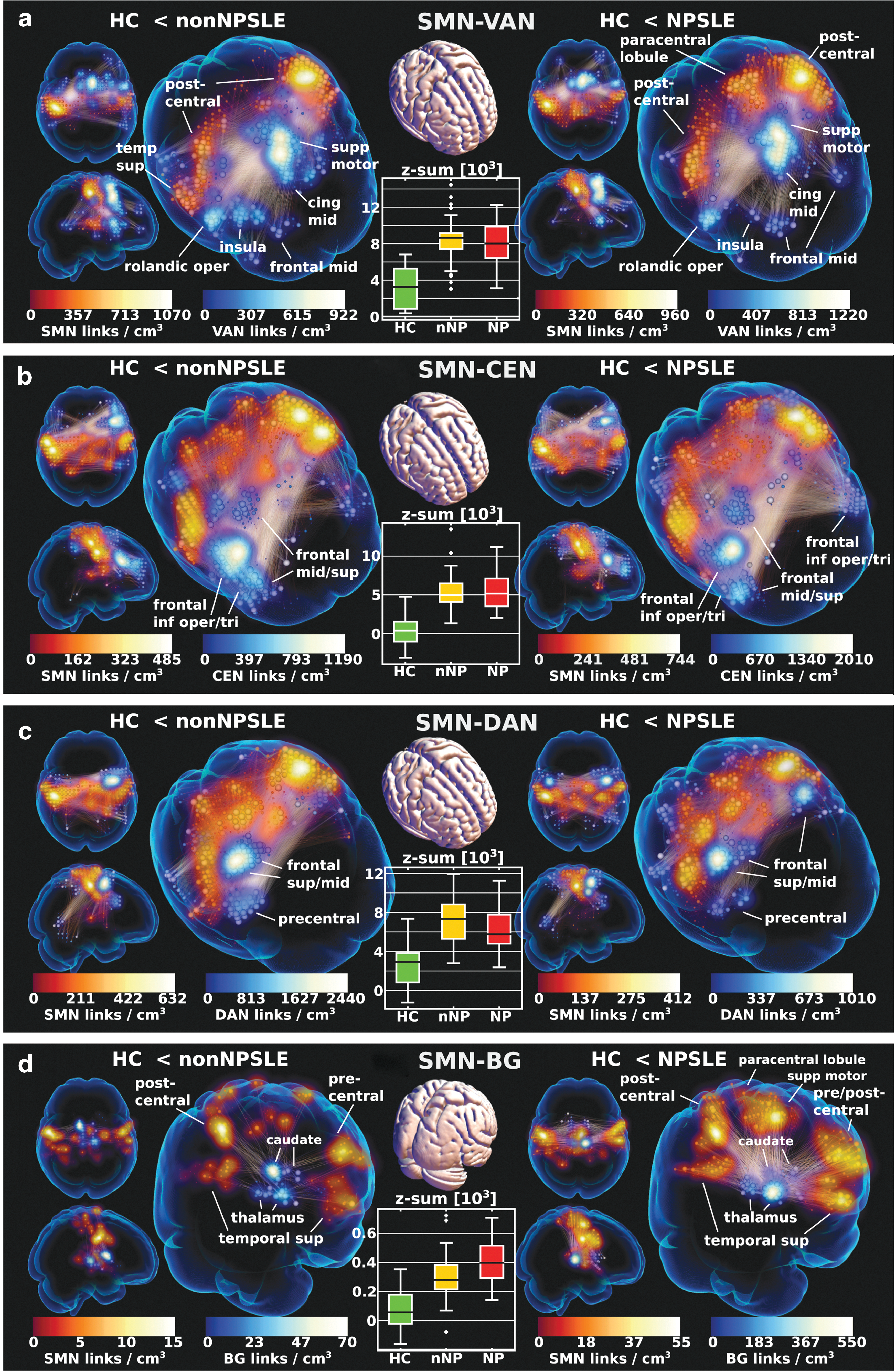
Link plots of hyperconnectivity in nonNPSLE and NPSLE relative healthy controls (HC) in between
The SMN-VAN/CEN hyperconnectivity was present in both subgroups and although there was variation in the sharpness of convergence (link density) on various substructures, this part of the hyperphenomenon did not seem strongly related to disease severity in terms of neuropsychiatric complaints (see z-sum box plots in Fig. 3a, b).
In addition, the same SM regions converged on subcortical structures in BG (Fig. 3d). Although generally more strongly expressed in HC-NPSLE, right thalamus had pronounced hyperconnectivity with high connectivity area in left SMN.
Ventral/dorsal attention and interconnected networks
There was hyperconnectivity within the VAN (also referred to as the salience network) and DAN in both contrasts (although somewhat stronger in HC-non-NPSLE), and involved the same VAN and DAN parts as in the SMN-VAN and SMN-DAN parts, respectively (Fig. 3a, c). Also, the increased VAN-CEN connectivity involved the same structures as in SMN-VAN/CEN, that is, primarily medial anterior VAN and right frontal CEN, with a similar correspondence for DAN.
Hippocampus
The dominant hippocampal hyperconnectivity with the frontal parts of CEN was present in both contrasts, but was markedly more pronounced in HC-NPSLE (Fig. 4a). The pattern involved mainly anterior hippocampus and was emphasized on the right side, although with less apparent laterality in NPSLE, where both hippocampi (HI) were engaged. The HI-CEN pattern was accompanied by concomitant intra-HI connectivity with the same left-right balance as HI-CEN. In the box plot of Figure 4a, we see that HC tends toward resting-state anticorrelations on this subcomponent.
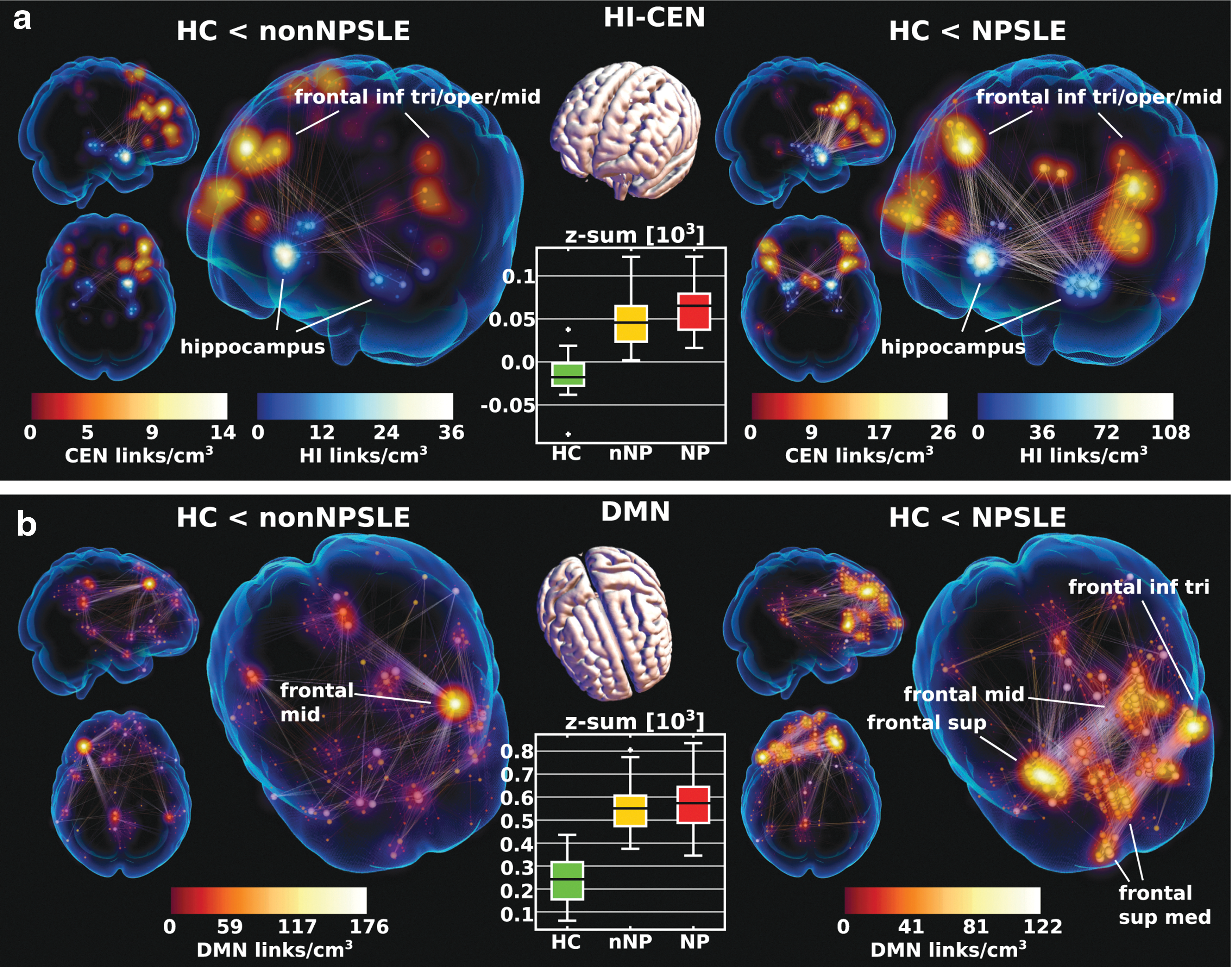
Dominant hippocampal (HI-CEN,
DMN and interconnected networks
The DMN exhibited hyperconnectivity in the anterior parts (Fig. 4b), especially in NPSLE. Similarly, intra-CEN and DMN-CEN also involved mainly frontal hyperconnectivity.
Correlation components
SLEDAI-2k, SDI, and disease duration
Using all SLE patients, we looked for network correlation components with three disease variables of interest: SLEDAI-2k, SDI, and disease duration.
Only significant negative network correlation components were found with SLEDAI-2k and SDI (Fig. 5a, b) with focus on inter-DMN and FTN connectivity. As most patients had the lowest index, the component was essentially powered by a higher index in less than half of all the SLE patients.
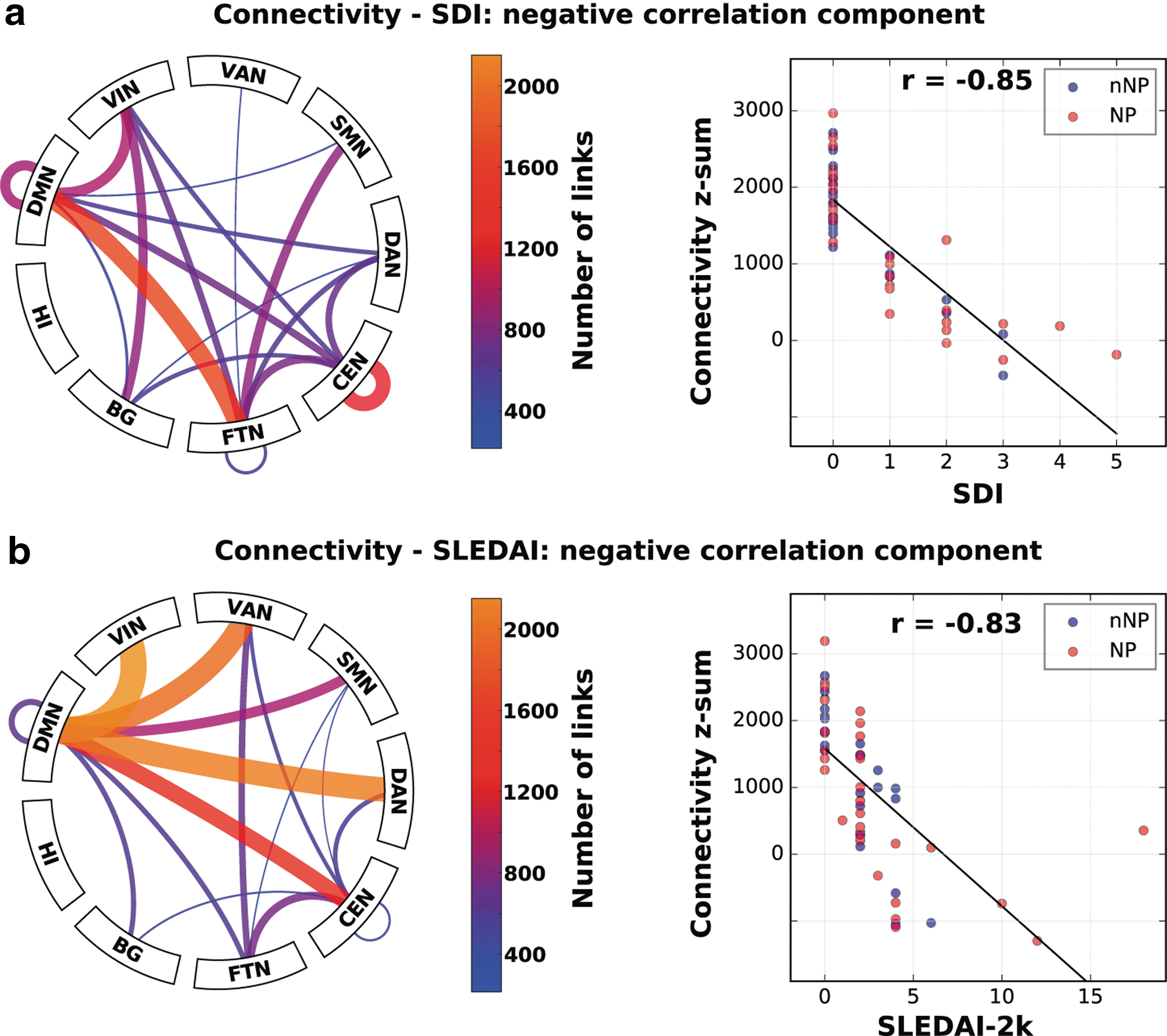
Correlation network components on which there was a significant negative correlation with SDI
In addition, network components correlating negatively and positively with disease duration were found, signifying reduced and increased functional connectivity with disease duration, respectively. The negative correlation component was much smaller than the positive and involved FT-DMN/VIN/SMN/VAN/DAN (focused on right parahippocampal gyrus) and right CEN with some spillover into DAN (Fig. 6a–c). Sum of connectivity on the negative correlation component yielded a Spearman's r = 0.76 with disease duration. This figure dropped slightly when restricting the sum to the subsets depicted in Figure 6b and c, giving an indication of the variations in the subpatterns across subjects.
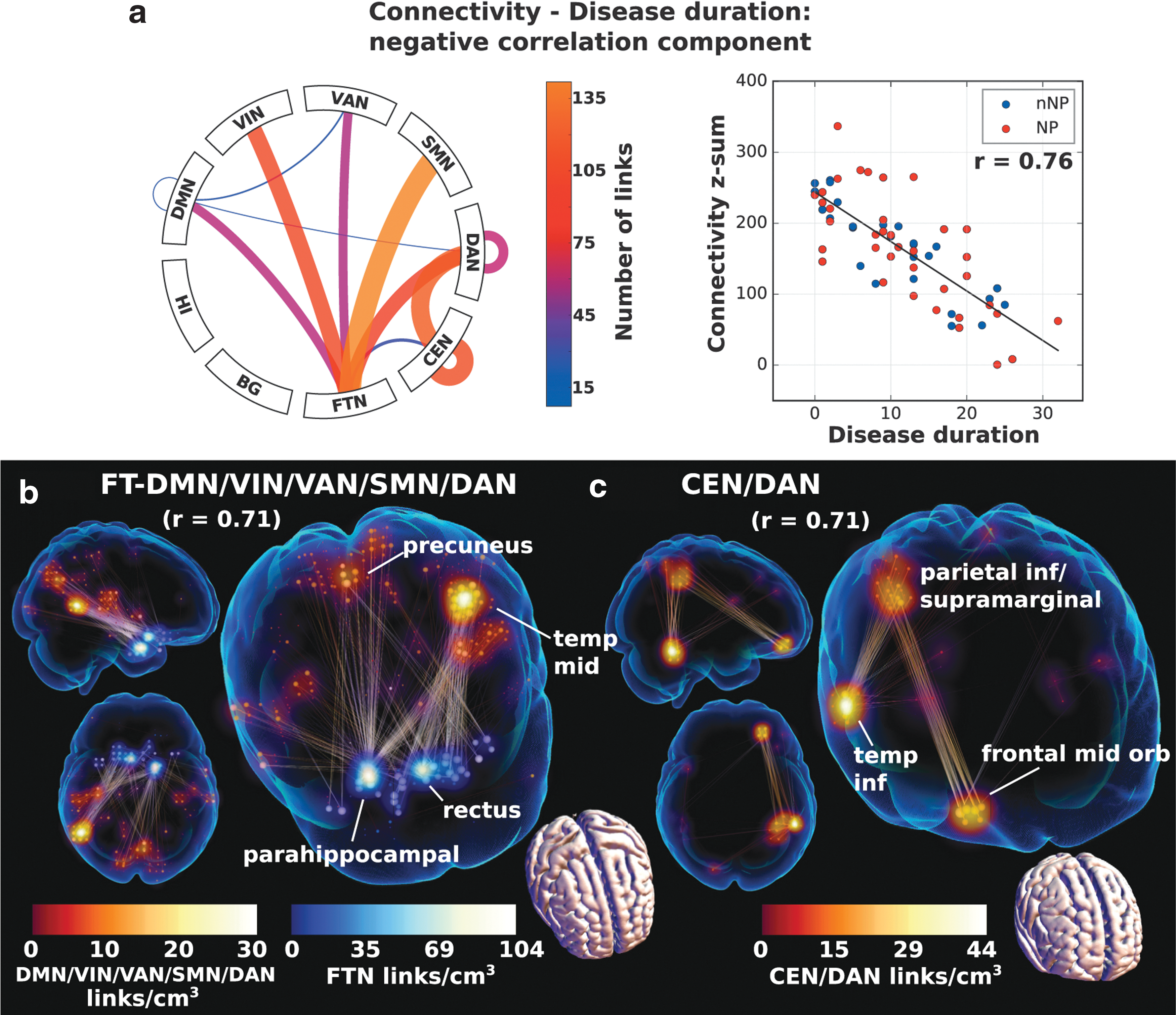
Correlation network components on which there was a significant negative correlation with disease duration and the link plots
The positive correlation component was much larger by comparison (Fig. 7a, b) and involved increased connectivity with duration in the anterior DMN (Fig. 7b) and similarly anteriorly weighted DMN-CEN. The SMN connectivity increased with primarily anterior parts (Fig. 7c–e). Spearman's coefficient was r = 0.76 between the sum of connectivity and duration on the whole component, a figure that persisted on the DMN part, but reduced slightly on the other subparts. Neuropsychiatric status had no association with disease duration and both subgroups spanned the whole spectrum of duration.
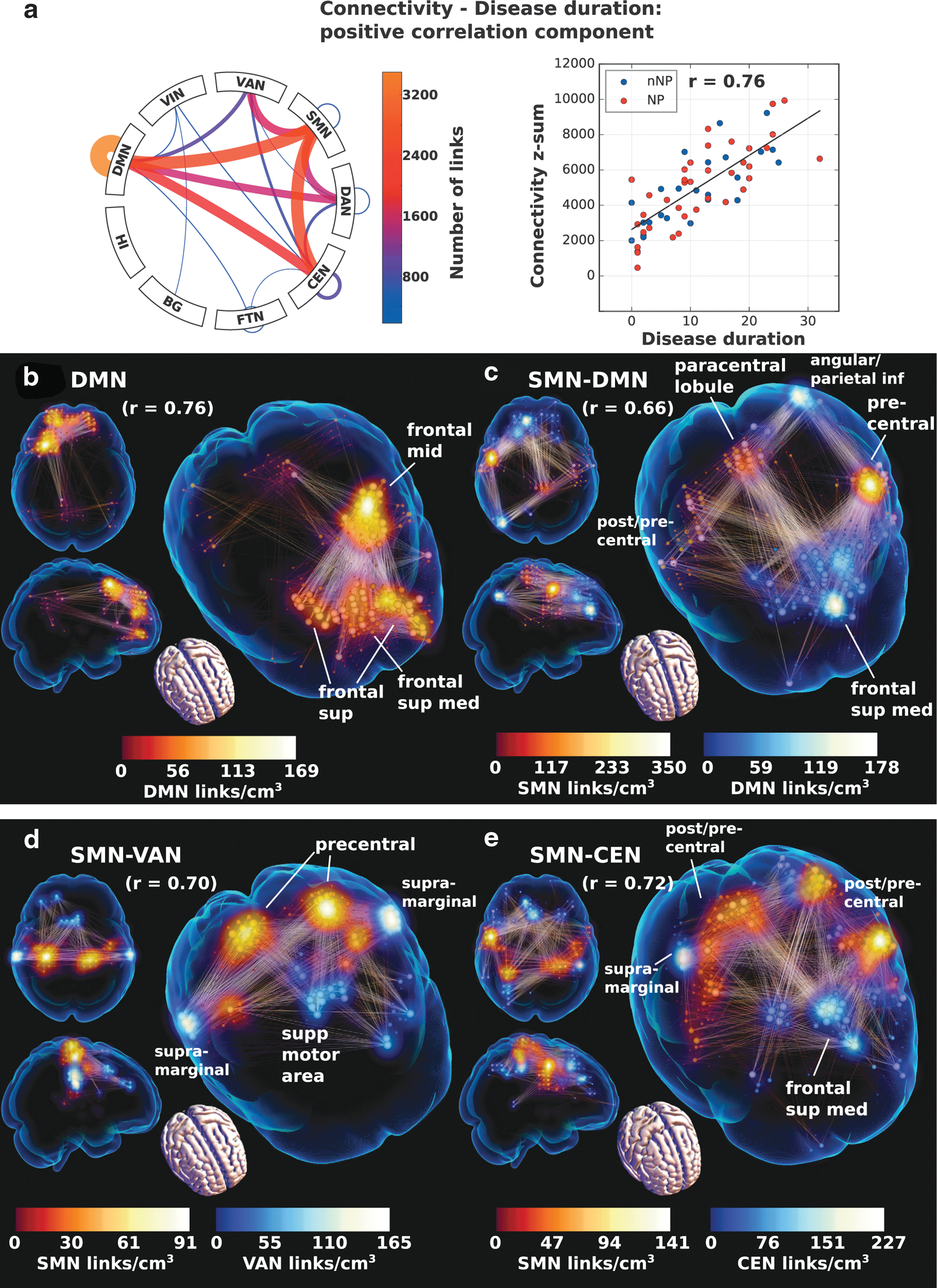
Correlation network component on which there is a significant positive correlation with disease duration and the link plot of the dominant parts of the full component in the connectogram
Discussion
We have, in this rs-fMRI study, demonstrated significant alterations in whole-brain connectivity within crucial networks in SLE patients when compared to HCs. Intriguingly, connectivity changes are present even in benign disease (Fig. 1), indicating that SLE per se is a disorder that already in an early stage affects the central nervous system. Moreover, we identified network components correlating with disease duration and severity (SLEDAI-2k and SDI), further elucidating the link between functional connectivity and standard disease characterization variables.
When comparing the connectivity of NPSLE to HC in our cohort, the hypoconnectivity within the DMN (Figs. 1 and 2) together with the hyperconnectivity of intra- and inter-SMN projecting to CEN and VAN was the most striking finding. Although qualitatively similar to non-NPSLE, the NPSLE hyperconnectivity comprised a larger set of links with the exception of SMN-DAN, which was more pronounced in non-NPSLE (Figs. 1 and 3).
We also found hyperconnectivity in the anterior parts of DMN at a non-NPSLE/NPSLE balance matching that of the DMN hypoconnectivity.
The hyperconnectivity profiles relative to HC were rather similar for non-NPSLE and NPSLE, perhaps containing an element related to pain levels and mobility, manifested in the heavy focus on SMN. It is, however, also probable that part of the hyperconnectivity represents some compensatory action and we note that there is clearly more involvement of DAN in the HC-non-NPSLE contrast.
The pronounced hypoconnectivity in DMN and CEN observed in non-NPSLE as well as in NPSLE when compared to HC, together with the left-hand side hypoconnectivity of VAN-VIN (involving insula connecting to areas of higher order associative and not primary visual cortex, see Fig. 2), is highly interesting.
VAN is a large-scale network known to be crucial in task switching and switching between internal and external events, such as engaging CEN and disengaging DMN (Menon et al., 2011; Yang et al., 2016), and there is some evidence that VAN modulates the relationship between DMN and CEN (Di and Biswal, 2014). The interaction between VAN, CEN, and DMN has been related to higher level functions (Chen et al., 2013).
Previous studies have suggested that the altered connectivity of VAN could be compensatory to the decreased function of DMN and CEN (Bressler and Menon, 2010; Menon, 2011; Seeley et al., 2007; Sridharan et al., 2008). Given the laterality of the VAN-VIN hypoconnectivity, which is focused on the left insula, this connectivity reduction may also be related to the hyperconnectivity converging on the right-hand side of VAN.
Although spatial similarities of CEN, DMN-CEN, and DMN connectivity may be associated with the inherent difficulty of defining networks (high individual variability and dependence on a number of networks sought), the internetwork connectivity cannot be entirely explained by spillover. Rather, it is likely a pathological feature of disrupted task-positive and task-negative switching.
Internetwork hypoconnectivity may very well manifest itself between these networks as a critical failure to integrate the modularized intranetwork processing.
It is tempting to regard the hyperconnectivity as a purely beneficial compensatory function and the hypoconnectivity as a detrimental functional failure, but we hypothesize that this simple interpretation may not always apply; the hypo- and hyperphenomena are linked in a complex way, ultimately reflecting their intimate relationship in the remodeling of functional networks. We can envision two not mutually exclusive mechanisms for the relative excess of both hypo- and hyperconnectivities in the HC-NPSLE contrast: (1) a greater insult to network functionality caused by neuronal dysfunction (manifest in the hypoconnectivity) leads to a greater, but still suboptimal, compensatory action (hyperconnectivity), and (2) a greater functional remodeling in NPSLE manifests in dominant hyperconnectivity, which is not necessarily all compensatory or beneficial, causing some hypoconnectivity by way of functional remodeling (thereby a consequence of the powerful hyperpart draining resources). For our cohorts, the HC-NPSLE hyperconnectivity, presumed to, in part, represent compensatory remodeling, is generally greater in HC-NPSLE than for HC-non-NPSLE. According to mechanism 2 above, it is feasible that the relative excess of hyperconnectivity in NPSLE leaves a greater mark of concomitantly reduced connectivity related to this remodeling. We find it likely that this reasoning applies to the smaller excesses of hypoconnectivity in HC-NPSLE relative to HC-non-NPSLE, but does not necessarily extend to the dominating DMN reduction (for which mechanism 1 above is perhaps more reasonable). This hypothesis is supported by the relative similarities of the hyperparts versus the imbalance in hypoparts dominated by HC-NPSLE DMN (Fig. 1).
Hypo-DMN is a major feature separating HC-non-NPSLE from HC-NPSLE and an especially sensitive marker for network disruption and failure to regulate internal/external attention switching. As such, the larger set of DMN hypoconnectivity links in HC-NPSLE is consistent with reduced functionality and the cognitive symptoms of NPSLE. In this context, we note that a compensatory hyperconnectivity may produce a more easily detectable signature as it represents a coordinated response mechanism, whereas underlying hypoconnectivity could have a greater variance, rendering it more elusive in group comparisons.
Close examination of network components reveals that mechanism 2 (concomitant hypotriggered by dominant hyperconnectivity) is consistent with the seemingly contradictory result of HC-non-NPSLE having a small excess of hypoconnectivity in DAN-BG relative to HC-NPSLE (Fig. 3b). Carefully comparing DAN-BG hypo- and SMN-DAN hyperpatterns (Fig. 3c), we find them to be spatially related (with a larger set of SMN-DAN links in HC-non-NPSLE converging on the left-hand side dorsally, and a concomitant larger left-hand, dorsal hypoconnectivity in DAN-BG). In this study, the complete dominance of hyperconnectivity highlights compensatory activity as the main effect and the small DAN-BG hypoconnectivity as concomitant functional remodeling, ultimately related to the finite resources of the human brain.
Overall, our findings of increased fronto-temporal-parietal and reduced DMN connectivity suggest that patients without any clinical or subjective signs of neuropsychiatric symptoms might still have a compromised network architecture. Since NPSLE and non-NPSLE in general displayed similar patterns, the results may indicate that connectivity changes are not so dependent on the status of clinically overt NPSLE, whose main distinguishing feature was DMN dysfunction.
Anterior DMN hyperconnectivity appears proportional to the amount of DMN hypoconnectivity and a spatial correspondence can be seen comparing the two patterns (Figs. 2a and 5b). A partitioning with anterior/posterior hyper/hypoactivity is been associated with age and is very pronounced in Alzheimer's patients, indicating a remodeling of DMN in response to neural dysfunction (Jones et al., 2011). It could be hypothesized that SLE patients, even in mild disease, alter their functional connectivity in a compensatory manner, to maintain their premorbid intellectual functions. Of course, there is always a balance between insult and compensation, where the former should by definition not be dominant in the non-NPSLE group.
This reorganization of functional connectivity may, in part, be a coping mechanism enlisted in response to neuronal damage, recruiting auxiliary pathways to maintain cognitive and motor function. Increased levels of pain and disrupted mobility can also have an impact on functional connectivity. In fact, to isolate changes originating in direct neurological damage, an alternative modality such as glial PET imaging could be of use, providing a map of neuroinflammatory responses (Kannan et al., 2009). It is beyond the scope of the present article, and a detailed analysis of the relationship between connectivity and cognitive test scores in a forthcoming article will hopefully provide further insight into the role of compensatory mechanisms.
Whereas it is tempting to regard hyperconnectivity as beneficial and connected with improved performance, this is not necessarily the case, clearly illustrated by the HI-CEN hyperconnectivity dominant in NPSLE. Presumably, this could represent suboptimal attempts at compensatory action to counter the NPSLE symptoms, where detrimental effects have overtaken existing cognitive reservoirs.
Similar findings of recruitment of additional pathways have previously been illustrated in task-positive fMRI studies in lupus patients (Barraclough et al., 2015; Bokde et al., 2006; Fitzgibbon et al., 2008; Mikdashi, 2016).
It could be speculated that the decreased connectivity of DMN and hyperconnectivity in the SMN network might be due to difficulties relaxing during the MRI scan, in turn due to corporal pain, arthritis, or anxiety. However, these findings could also be related to the actual neuronal dysfunction and need of recruiting additional pathways. No matter the cause, there is a hyperconnectivity signature associated with SLE and a more severe hypoconnectivity primarily affecting DMN in NPSLE. These signatures are distinctive enough that they could serve as biomarkers for detecting and perhaps even staging the disease.
We also looked for patterns correlating with some disease variables of paramount interest: disease duration, SLEDAI-2k, and SDI. Note that the comparison of these with the hypo- and hyperconnectivity network components is not straightforward, as the correlation components are simply a set of links that correlate with the given variable and may as such overlap with both hypo- and hyperparts of the group components. To avoid confusion, we reserve the hypo- and hyperconnectivty concepts for comparison with HCs.
Disease duration had a small negative (decreased connectivity with time) and a large positive component (increased connectivity with time). It is noteworthy that the small negative component cannot simply be interpreted as a general absence of large reductions in connectivity with time, as the impact of reduced significance could also be a manifestation of the heterogeneous nature of mixed reduced connectivity profiles. However, since short disease duration subjects are closer to HCs, there are similarities between the hypoconnectivities of NPSLE/non-NPSLE>HC and the disease duration component, primarily in the connections with FTN and also intra-CEN (c.f. Figs. 1 and 7). Likewise, similarities between the hyperconnectivities of the group contrast and the larger positive duration component, where we recover the anterior DMN hyperconnectivity hypothesized to be compensatory. The DMN hypoconnectivity does not appear to have a negative duration component counterpart, but this is not strange since hypo-DMN is an SLE hallmark and there is no difference in disease duration between the two SLE groups.
These results are consistent with previous studies, indicating increased neuronal activation in areas associated with working memory and increased full brain activation being related to short-term disease, whereas long-term disease was associated with increased neuronal damage and decreased activation (Mackay et al., 2011).
Elevated SLEDAI-2k score previously has been postulated as a possible marker for cerebral involvement in SLE (Hou et al., 2013; Lin et al., 2011). Although we found no strong support for this hypothesis as many of the functional connectivity changes occur before the two indices increase (less than half of all SLE subjects have higher index powering the SDI/SLEDAI-2k correlation in Fig. 6), we did find significant negative correlation components (reduced connectivity with increased index). The fact that we could only find negative components goes against the notion that corporeal ailments are a major cause of the strong hyperconnectivity phenomenon and could be interpreted as support for the presence of compensatory mechanisms. The SDI appeared more similar than the SLEDAI-2k component when comparing with the negative disease duration component (c.f. Figs. 5 and 6) and the group contrast (Fig. 1a, b). Although there were similarities also with SLEDAI-2k (please note the differing scales when comparing), there was clearly a heavier focus on inter-DMN connectivity in SLEDAI-2k.
Our results are in line with Mackay and colleagues (2011), neither SDI nor SLEDAI-2k seems to strongly relate to altered brain connectivity on a group level, as no significant differences in these parameters were noted in our study between the SLE subgroups. However, it should be emphasized that patients in our cohort had a generally low SLEDAI-2k score when entering the study.
We recognize that this study has some limitations. First and foremost, NPSLE is a highly heterogeneous disease with a wide range of symptoms and unknown pathology. Our analyzed groups were relatively small and not entirely comparable size-wise.
However, we consider that our group sizes measure up to the number of subjects in previous studies (Mackay et al., 2011; Mikdashi, 2016; Mosca et al., 2011) and are at least indicative, as several previous studies with fewer patients have demonstrated sufficient sensitivity (Mikdashi, 2016).
Second, we did not distinguish between active or chronic NPSLE (Klein et al., 2010) or between the different symptoms the patients suffered from, resulting in a heterogeneous group of NPSLE patients.
Furthermore, it could be argued that the pronounced fatigue in the nonNPSLE and NPSLE subgroups in comparison to HCs directly could affect the patterns of intrinsic connectivity (Gobbi et al., 2014; Pardini et al., 2010; Wortinger et al., 2016). There was, however, no significant difference in fatigue score between the non-NPSLE and NPSLE, and why the fatigue itself cannot explain the altered connectivity. On the contrary, it could suggest that the altered connectivity might be a cause of fatigue. This hypothesis would be in concordance with previous studies on adolescent chronic fatigue syndrome and fatigue in multiple sclerosis (Pardini et al., 2010; Wortinger et al., 2016).
Major depression has been described to alter hubs in the DMN and cause increased interaction between the DMN and the VAN. Furthermore, decreased interaction in the CEN in patients with major depression has been suggested (Yang et al., 2016). Hypothetically, depression could somewhat explain the altered connectivity seen in SLE patients in general. To minimize possible bias from mood disorder in the current study, we excluded patients with severe depression defined as MADRS-score >34.
No significant differences in ischemic lesions, microbleeds, or contrast-enhanced lesions could be found between any of the groups (HC, non-NPSLE, NPSLE) and no lesions were identified among HCs (Table 1).
Our results are, however, in line with previous resting-state and task-positive fMRI studies on lupus patients that altogether coherently conclude that SLE per se causes alterations in intrinsic connectivity even before other NP symptoms are detectable (DiFrancesco et al., 2013; Hou et al., 2013; Lin et al., 2011) and that these changes might have a progressive nature.
It seems that, regardless of cause, the SLE patients in general have network alterations and supposed microscopic brain damage although appearing normal on standard MRI examinations.
We believe that to better understand the underlying pathophysiological mechanism, further studies are necessary. Ideally, these should involve longitudinal studies and combine modalities that allow more careful delineation of neuroinflammation, such as glial PET (Kannan et al., 2009). This would also aid in further separating functional changes due to direct neurological damage from those generated by bodily symptoms.
In summary, this study demonstrates that SLE patients have significant alterations in their resting-state network integrity, although with a different pattern of intrinsic connectivity for patients with and without neuropsychiatric complaints. These findings might be due to both compensatory mechanisms through recruiting additional networks and due to neuronal damage and may partly explain neuropsychiatric symptoms in SLE patients. Findings indicate that SLE per se is a disorder that affects the central nervous system and that some of these alterations show a significant correlation to disease duration.
We conclude that SLE patients in general and NPSLE patients in particular suffer from altered brain connectivity, suggesting compensatory effects, demonstrated as hypo- and hyperconnectivity, in several crucial resting-state networks.
Footnotes
Acknowledgments
This study has been supported by the Skåne University Research Funding, the Swedish Rheumatism Association, the Alfred Österlunds Research Foundation, the King Gustav V 80-years Foundation, and Greta and Johan Kocks Foundation.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.