
Abstract
Patients with schizophrenia (SCH) often demonstrate impairment in social-cognitive functions as well as disturbances in large-scale network connectivity. The ventromedial prefrontal cortex (vmPFC) is a core region of the default mode network, with projections to limbic structures. It plays an important role in social and emotional decision-making. We investigated whether resting-state functional connectivity (FC) relates to the cognitive and affective domains of theory of mind (ToM). Twenty-three SCH patients and 19 healthy controls (HCs) underwent resting-state functional magnetic resonance imaging scanning. vmPFC seed connectivity was correlated with behavioral measures assessing ToM domains. SCH performed less well than HCs in both ToM task domains. An analysis of the resting-state FC revealed that SCH had reduced connectivity from the vmPFC to the subcallosal cortex, right amygdala, and right hippocampus as a function of behavioral scores in both ToM domains. Within-group analyses indicated that in HCs, the performance in ToM was positively associated with frontoamygdalar resting-state connectivity, whereas in SCH, the performance in ToM was negatively associated with the frontosubcallosal connectivity. Differences in the pattern of the resting-state frontolimbic connectivity and its associations with performance in ToM tasks between the two study groups might represent a different setup for processing social information in patients with SCH.
Introduction
People with schizophrenia (SCH) frequently demonstrate impairment in social-cognitive functions (Pinkham et al., 2003). These abnormalities are linked to functional outcomes related to everyday social life (Couture et al., 2006; Irani et al., 2012) and seem to be a stronger predictor of optimal functioning than deficits in general cognition (Fett et al., 2011). Whereas social-cognitive abilities encompass a variety of processes, the most studied social cognition component in SCH is the theory of mind (ToM). ToM is defined as the ability to understand and interpret the intentions, emotions, and beliefs of other individuals to predict their behavior accurately and to act accordingly (e.g., Frith and Frith, 1999). There is general agreement that ToM consists of two domains, the first involved in understanding others' intentions and beliefs (cognitive domain), and the second implicated in understanding emotions and feelings (affective domain) (Abu-Akel and Shamay-Tsoory, 2011; Kalbe et al., 2010).
Only a few studies have explored both the cognitive and affective aspects of mental-state decoding in patients with SCH. Shamay-Tsoory and colleagues (2007b), for instance, report impairment in SCH patients that was present only during the attribution of affective mental states, without abnormalities in reasoning about others' cognitive mental states. In contrast, a study conducted by Lee and colleagues (2010) yielded the opposite results, whereby only cognitive but not affective mentalizing was impaired. Furthermore, Montag and colleagues (2011), using probably the most complex social-cognitive task of the studies conducted so far, found impairment in both domains.
Similarly, only two studies have explored brain activation during both modalities of ToM in SCH: Benedetti and colleagues (2009) and Lee and colleagues (2010). Weaker activation in superior temporal areas during cognitive perspective taking was found in SCH patients than in healthy controls (HCs) in both studies, whereas inferring emotional mental states resulted in greater activation in the left insula (Lee et al., 2010) or temporoparietal junction (Benedetti et al., 2009) in the patients with SCH than in the HCs.
Inconsistencies in the existing literature might reflect the heterogeneity of symptom profiles across different samples of SCH patients. The current clinical understanding of the disease is that a variety of symptoms, including cognitive or sociocognitive disturbances, are related to a failure to integrate the activity of local and widespread regions (Andreasen et al., 1996; Phillips and Silverstein, 2003). Thus, it is critical to study not only the deviations in regional activity but also, more importantly, the large-scale network connectivity.
A strong evidence of aberrant resting-state network connectivity in SCH supports this perspective (Garrity et al., 2007; Liang et al., 2006; Shinn et al., 2015). Similar findings have been reported in people at high risk of developing the disease (i.e., siblings of SCH patients; Collin et al., 2011). Furthermore, these disturbances in functional connectivity (FC) have been found to be related to several cognitive functions (Spreng et al., 2009), as well as to positive and negative SCH symptoms (Venkataraman et al., 2012).
The default mode network (DMN) is one of the most studied large-scale networks. Originally it was understood as a task-independent network, present at rest and deactivated during tasks. The most recent findings, however, indicate that the DMN is activated in certain types of tasks, and those of social-cognitive nature especially, given its large overlap with the social-cognitive neuronal network (Mars et al., 2012; Schilbach et al., 2008).
Various studies reported consistent findings that SCH patients have decreased resting-state FC between the midline structures of the medial prefrontal cortex (mPFC) and posterior cingulate cortex, the two core areas of the DMN (Fan et al., 2013; Holt et al., 2011; Zhou et al., 2007). This led to the hypothesis that such hypoconnectivity might contribute to SCH patients' social-cognitive impairment (Holt et al., 2011). The ventromedial prefrontal cortex (vmPFC) forms projections to the limbic regions (Ongür and Price, 2000; Young et al., 1994), which play an important role in social and emotional decision-making (Bechara, 2000; Grossman et al., 2010; Koenigs and Tranel, 2007; Moretti et al., 2009; van den Bos and Guroglu, 2009). Indeed, one study revealed an association between hypoconnectivity within the vmPFC and abnormal emotion regulation in chronic SCH patients (Fan et al., 2013). A strong association was found between gray matter abnormalities in the vmPFC and ToM skills in SCH, even after controlling for global cognition (Hooker et al., 2011). In addition, patients with lesions in the vmPFC have similar patterns of impaired affective ToM as compared with controls (Shamay-Tsoory et al., 2007a).
Given the scarcity of studies exploring both cognitive and affective domains in ToM, as well as the evidence that FC of the vmPFC seems to capture difficulties in social cognition, we aimed to investigate the extent to which DMN FC, assessed with vmPFC as the seed of interest, relates to performance in a ToM task pertaining to these domains, in a sample of SCH patients and HCs.
Materials and Methods
Participants
Twenty-three SCH patients (4 women) and 19 HCs (5 women) participated in the study (for demographic and clinical information, see Table 1). The control group was matched to the patients in terms of gender, age [t(40) = −0.485, p = 0.630], and completed education [t(40) = 1.194, p = 0.239].
Clinical and Demographic Information
CPZ, chlorpromazine; SD, standard deviation.
The patients were recruited from the Department of Psychiatry at the University Hospital Brno, Czech Republic; they were diagnosed with SCH according to ICD-10 criteria and the Mini-International Neuropsychiatric Interview (MINI), and treated with antipsychotic medication. Exclusion criteria for the patients were history of substance dependence or any other neurological or somatic condition potentially affecting brain structure and function. HCs were recruited among hospital staff, medical students, and at local community centers through information fliers. The inclusion criteria for controls were no presence of any Axis I psychiatric conditions (based on the MINI) in the participants themselves or in their family history, and no drug dependence/addiction or any neurological/somatic condition contraindicating the magnetic resonance imaging (MRI) examination. All the participants were right-handed except for one SCH.
The study was approved by the Institutional Review Board of the University Hospital Brno, and all the participants provided informed consent before taking part in the study.
Materials
Social cognition was assessed behaviorally by a ToM task called the faux pas recognition test (FPRT) (Baron-Cohen et al., 1999; Gregory et al., 2002; Stone et al., 1998) to examine cognitive and affective ToM components, and through self-reporting using questionnaires measuring empathy (Questionnaire of Cognitive and Affective Empathy [QCAE]) (Reniers et al., 2011) and emotion dysregulation (Difficulties in Emotion Regulation Scale [DERS]) (Gratz and Roemer, 2004). Both self-reporting instruments were translated into Czech using a standard procedure of independent forward and backward translation to ensure the conceptual equivalency of the items. Our team pretested the final versions of the translated questionnaires on a large sample of healthy participants (N = 255 [88 men]; mean age = 22.5 years; SD = 3.1), revealing excellent levels of internal consistency (QCAE cognitive: Cronbach's α = 0.87; QCAE affective: α = 0.77; DERS: α = 0.91; Czekóová et al., 2017).
To assess cognitive functions, we administered several neuropsychological tests. Selected subtests from the Wechsler Adult Intelligence Scale-III (WAIS-III) (Wechsler, 1997a) were employed to index participants' processing speed (digit symbol—coding), visual perception and attention to detail (picture completion), working memory, numeric reasoning (arithmetic), and abstract verbal reasoning (similarities). Verbal memory was assessed using the word list from the Wechsler Memory Scale-III (WMS-III) (Wechsler, 1997b). Cognitive control was evaluated with a Stroop interference score and calculated according to Chafetz and Matthews (2004); other executive functions were evaluated with the Wisconsin Card Sorting test (Heaton et al., 2003) and semantic and lexical (phonemic) verbal fluency tasks (Strauss et al., 2006).
Procedure
The subjects underwent functional MRI (fMRI) measurement first. They were instructed to lie still in the scanner in a relaxed state and with their eyes closed. After the scanning session, QCAE and DERS questionnaires were administered together with the FPRT in its full version. The FPRT consists of 20 short stories describing everyday social situations, with 10 stories involving faux pas (FP+) and the rest describing only minor misunderstandings (FP−). Participants listen to each story and at the end they are asked to determine whether a social FP occurred; that is, whether anyone said anything inappropriate or hurtful.
To perform well, participants have to understand two mental states simultaneously: first, that the person who commits the FP does so unintentionally (not knowing something important in a given context); and second, that their remark would be perceived as awkward/unpleasant/hurtful by the other character from the story. To control for the possible influence of cognitive functions (e.g., attention or working memory), the printed stories are available for reference for the entire testing time. General comprehension is tested by two control questions. FPRT responses were coded by two raters independently, with an excellent level of inter-rater reliability achieved (κ = 0.92). Cognitive measures were administered in a separate testing session.
Image acquisition parameters and data preprocessing
Resting-state fMRI measurements were obtained using a 1.5 T Siemens Symphony scanner equipped with Numaris 4 System (MRease). A total of 100 volumes were acquired (echo time [TE] = 40 ms, repetition time (TR) = 3000 ms, flip angle (FA) = 90°, 32 axial slices, slice thickness = 3.5 mm, in-plane resolution 220 × 220 mm, matrix size 64 × 64, voxel size 3.4375 × 3.4375 mm).
The FC data were analyzed using the MATLAB-based CONN-Toolbox (Whitfield-Gabrieli and Nieto-Castanon, 2012). The images were preprocessed. The preprocessing consisted of (1) correction for slice timing differences, (2) alignment to mean image to correct for head motions, (3) normalization to a common stereotactic space (Montreal Neurological Institute template) using an affine transformation, and (4) smoothing using an isotropic 8-mm Gaussian kernel. The denoising of the resting-state data consisted of regressing out the six movement parameters obtained during realignment, regressing out the white matter and ventricular signals, and scrubbing and filtering using a 0.008–0.9 Hz band-pass filter.
Behavioral and self-report data analyses
All experimental FPRT scores were employed in our analyses according to the test manual (Stone et al., 1998), namely (1) detection of FP (detection), (2) understanding of the inappropriateness of a remark mentioned in the story (inappropriateness), (3) understanding of the intentions of the characters in the story (intention), (4) belief that FP was not committed on purpose (belief), and (5) empathy for the story character affected by the inappropriate remark (empathy). Three of the five FP-related scores (inappropriateness, intention, and belief) represent cognitive ToM; detection ToM, and empathy index affective ToM (Poletti et al., 2012). Both cognitive empathy and affective empathy scores derived from QCAE and the sum score from DERS were used in the analyses.
Four participants (two SCH and two HCs) did not complete neuropsychological testing and were, therefore, excluded from the analysis involving cognitive functions. All the remaining analyses were performed on the complete sample.
Raw scores from Stroop test and the WAIS-III and WMS-III subtests were transformed into standard (scaled) scores. The rest of the variables are expressed as raw scores. Since the majority of the variables violated assumptions of normality, the performances of the two groups were compared using the Mann–Whitney U test. Relationships between the studied variables were calculated using the Spearman correlation coefficient. Statistical analyses were performed using SPSS 23 (IBM, Inc., Chicago, IL).
FC analysis
During the first-level (single-subject) analysis, Pearson correlation coefficients between the given seed regions and all other brain voxels were computed. The Pearson correlation coefficients were Fisher z-transformed. During the second-level (group) analysis, an ANCOVA model was used to assess the influence of the psychometric (FPRT) scores on the FC between the two groups. The DMN region of interest was centered in the vmPFC, the seed was a 12-mm-diameter sphere determined on previously published foci (x = −1, y = 47, z = −4) according to studies of task-related deactivations (Fox et al., 2005; Shulman et al., 1997). To further explore the differences between the groups, we performed a correlation analysis between ToM performance scores and vmPFC seed connectivity in each group separately.
Finally, to assess group differences in the association between ToM performance scores from the FPRT and the vmPFC connectivity measures with different clusters of interest, we conducted a regression analysis. Specifically, we used a hierarchical regression model in which FPRT scores represented the dependent variable and the connectivity measures; the group and interaction term (group × connectivity) constituted the independent factors that were entered into the model one by one, in this order. At each step, we tested the contribution of the newly added independent variable through an F-test assessing the change in the total variance explained by the model (R 2 change). Regression coefficients were estimated both using classical t-tests, as well as bootstrap tests based on 1000 samples, to account for the bias in estimation. Analyses were conducted using SPSS 24 (IBM, Inc.).
Results
Behavioral results
Social cognition
SCH reported lower cognitive empathy than HCs (U = 101.50; p = 0.003; r = 0.45), with the difference driven mainly by the perspective taking subscale (U = 100.00; p = 0.003; r = 0.46). No differences in affective empathy (p = 0.323) and emotion regulation (p = 0.800) were observed in total scores nor in any of the subscales.
SCH differed from HCs in overall performance in FPRT in both experimental (FP+; U = 63.50; p < 0.001; r = 0.60) and control stories (FP−; U = 105.50; p = 0.002; r = 0.48). In particular, the SCH group showed significantly lower scores than HCs in both cognitive (inappropriateness: U = 99.50; p = 0.002; r = 0.47; intention: U = 50.50; p < 0.001; r = 0.66, and belief: U = 64.50; p < 0.001; r = 0.60) and affective aspects of mentalizing (detection: U = 78.50; p < 0.001; r = 0.55; empathy: U = 114.50; p = 0.008; r = 0.40, see Fig. 1).
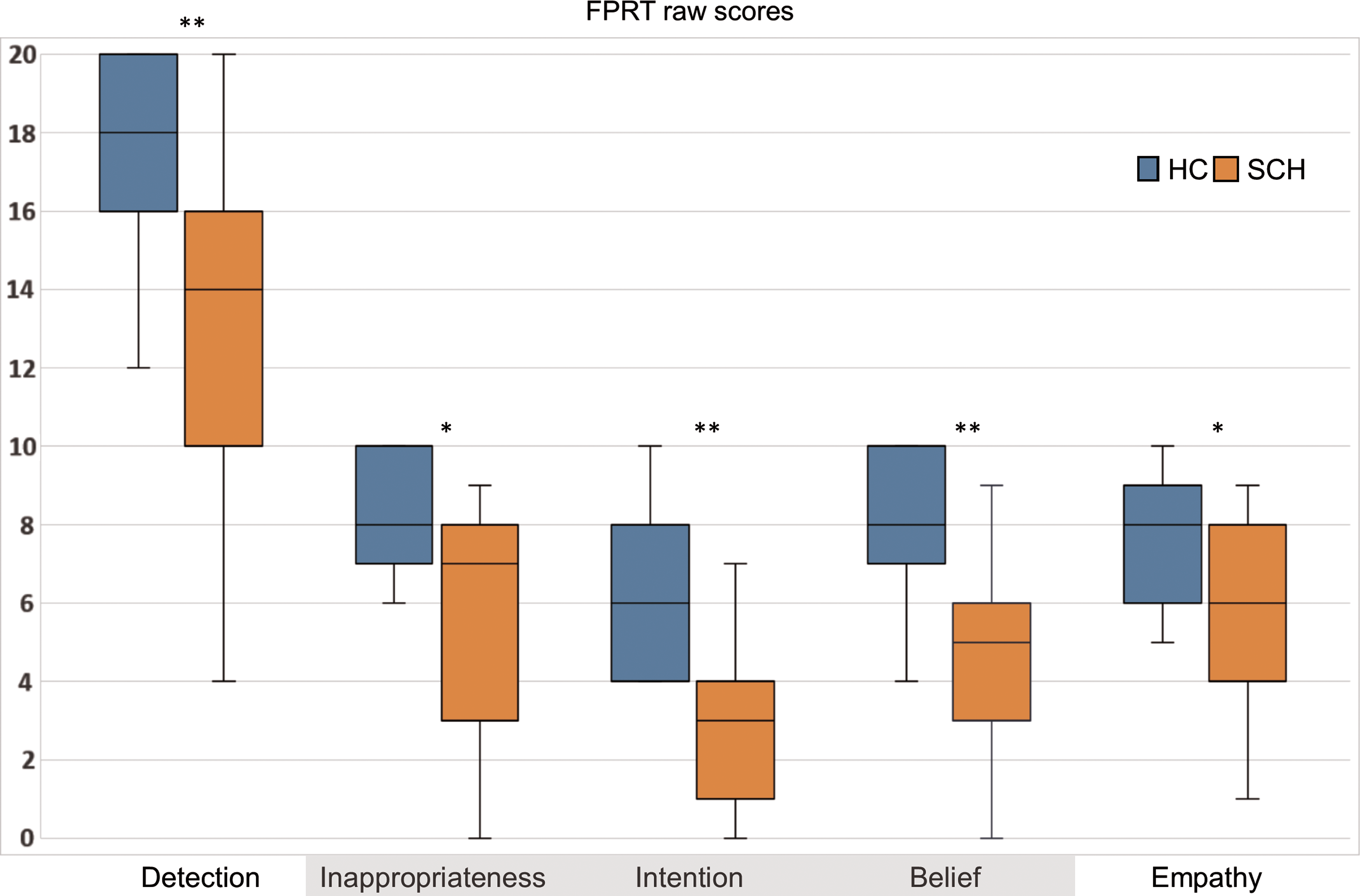
Comparison of cognitive (highlighted in gray) and affective components of FPRT between SCH patients and HCs; *p < 0.01; **p < 0.001. HCs, healthy controls; FPRT, faux pas recognition test; SCH, schizophrenia. Color images available online at
No relationship was found between social cognitive performance, antipsychotic medication dose, or disease duration.
Next, we analyzed relationships between self-reported measures of empathy and emotion regulation (as measured by QCAE and DERS) and FPRT scores. Neither of the questionnaire scores was related to FPRT performance in SCH. In contrast, a positive correlation between scores in affective empathy reported in QCAE and both affective and cognitive measures of ToM in FPRT (ρ = 0.38, p < 0.05; ρ = 0.60, p < 0.01) was found in HCs.
General cognition
No significant differences were revealed between SCH and HCs in abstract verbal reasoning (p = 0.144) and visual attention to detail (p = 0.096), but SCH performed significantly worse in the subtests measuring working memory (U = 63.50; p < 0.001; r = 0.55) and processing speed (U = 34.50; p < 0.001; r = 0.69). In addition, SCH performance was poorer than that of HCs in all three aspects of verbal memory, namely immediate (U = 34.00; p < 0.001; r = 0.69), delayed (U = 68.00; p < 0.001; r = 0.53), and recognition (U = 84.00; p = 0.003; r = 0.47).
Likewise, SCH performed considerably worse in categories completed within the Wisconsin Card Sorting test (U = 104.50; p = 0.022; r = 0.37) and the semantic verbal fluency subtest (U = 56.00, p < 0.001; r = 0.58). No significant differences were found between the two groups for Stroop interference score (p = 0.117) and lexical verbal fluency (p = 0.263). Table 2 presents an overview of descriptive statistic describing all cognitive measures.
Descriptive Statistic for Cognitive Measures
Although the data are not normally distributed, SDs are reported here to allow comparisons with other studies. Bold values indicate cognitive measures which differed significantly between patients and controls when nonparametric Mann–Whitney U tests were used.
p < 0.01; ** p < 0.001.
SD, standard deviation; WAIS-III, Wechsler Adult Intelligence Scale-III; WCST, Wisconsin Card Sorting Test; WMS-III, Wechsler Memory Scale-III.
As mentioned in the previous section, the largest differences between SCH and HCs based on effect size were shown in the ability to specify the intentionality of the story characters. Furthermore, FPRT intentionality score was associated strongly with cognitive performance in immediate verbal memory in SCH (ρ = 0.73, p < 0.001), with no such relationship observed in HCs.
Among SCH, there was no significant association between cognitive performance and antipsychotic medication dose; there was, however, a negative correlation between immediate verbal memory score and disease duration (ρ = −0.50, p = 0.018).
Functional connectivity
Resting-state vmPFC connectivity: group differences
SCH showed significantly stronger resting-state vmPFC connectivity to the right inferior frontal gyrus (IFG) region than HCs, whereas HCs exhibited stronger functional connections from vmPFC to the left inferior parietal lobule at rest. Connections to the limbic regions remained comparable in both groups (Fig. 2).
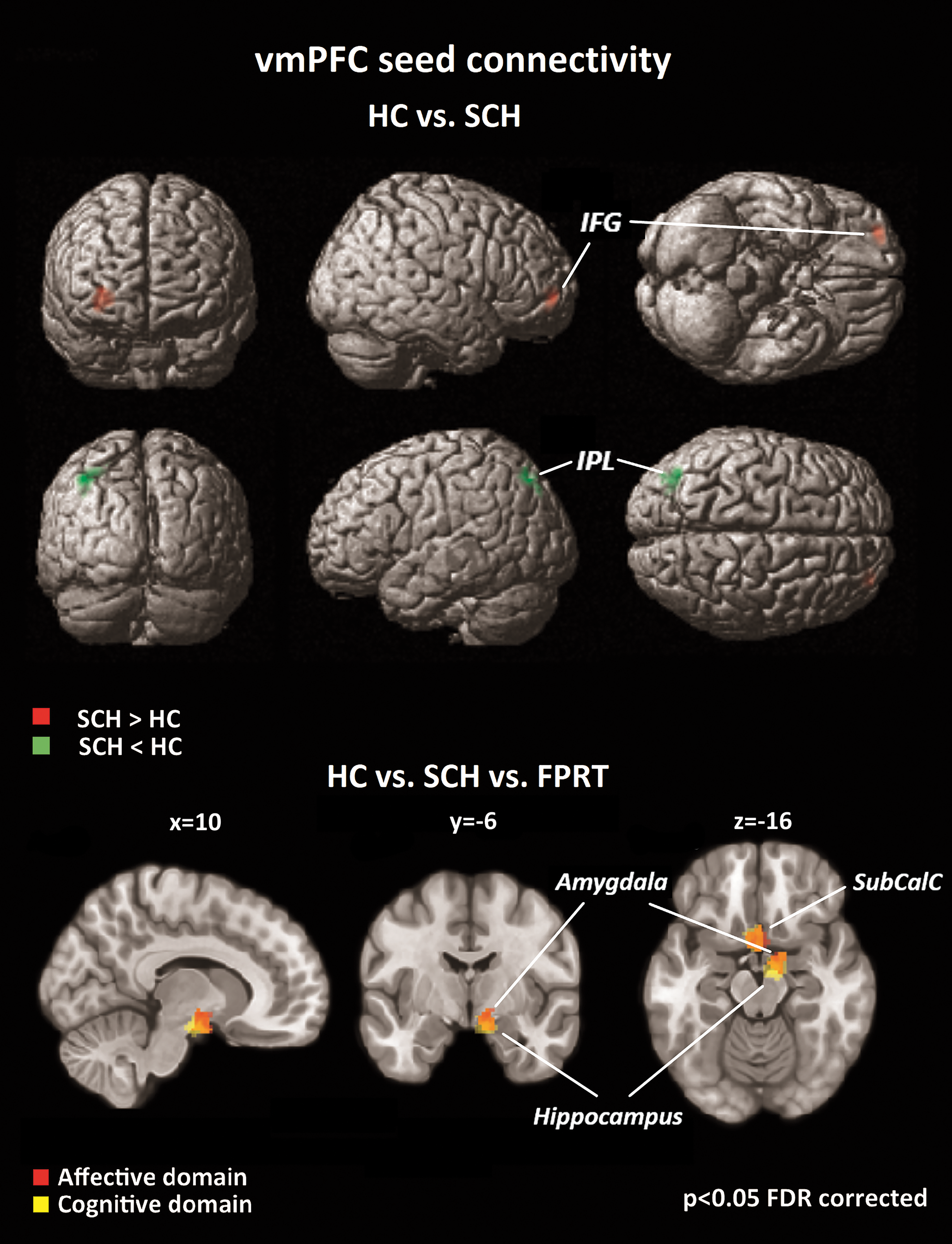
Resting-state vmPFC seed connectivity group comparison; SCH displayed stronger connectivity to the right IFG than HCs; HCs showed greater connectivity to the left IPL than SCH; when investigating group differences as a function of FPRT domains, weaker connectivity to limbic regions (SubCalC, amygdala, hippocampus) were revealed in SCH than in HCs. IFG, inferior frontal gyrus; IPL, inferior parietal lobule; SubCalC, subcallosal cortex; vmPFC, ventromedial prefrontal cortex. Color images available online at
Resting-state vmPFC connectivity related to social cognition: group differences
When regressed against affective and cognitive FPRT scores, ANCOVA revealed differences between the two groups in vmPFC seed connectivity, with similar results for both ToM domains. Specifically, SCH displayed weaker connectivity than HCs from the vmPFC (seed) to the subcallosal cortex, right amygdala, and right hippocampus as a function of FPRT scores (Fig. 2).
The hierarchical regression analysis revealed that there were significant group differences only for two clusters in the association between the connectivity measures with vmPFC and FPRT scores: the right amygdala (with inappropriateness) and the subcallosal region (with inappropriateness, detection, and empathy). The interaction term was significant (p < 0.05, based on a bootstrap test with 1000 samples) and negative in all cases, indicating that for SCH (compared to HCs), an increase in the connectivity measures was associated with a decrease in FPRT scores (see Table 3 for the regression coefficients and bootstrap test results).
Regression Models Estimating Faux Pas Recognition Task Scores Based on Connectivity and Group Membership and the Interaction Between These Two Variables
Coefficients highlighted in bold, p < 0.05 based on a bootstrap test with 1000 samples.
C, cortex; CI, confidence interval; HCs, healthy controls; R/L, right/left; SCH, schizophrenia patients; vmPFC, ventromedial prefrontal cortex.
A detailed analysis of the association of the two variables (connectivity and FPRT scores) within each group showed that the group difference for the right amygdala was driven by a significant positive relationship present in HCs, whereas group differences for the subcallosal connection were driven by a significant negative relationship seen in SCH (Fig. 3).
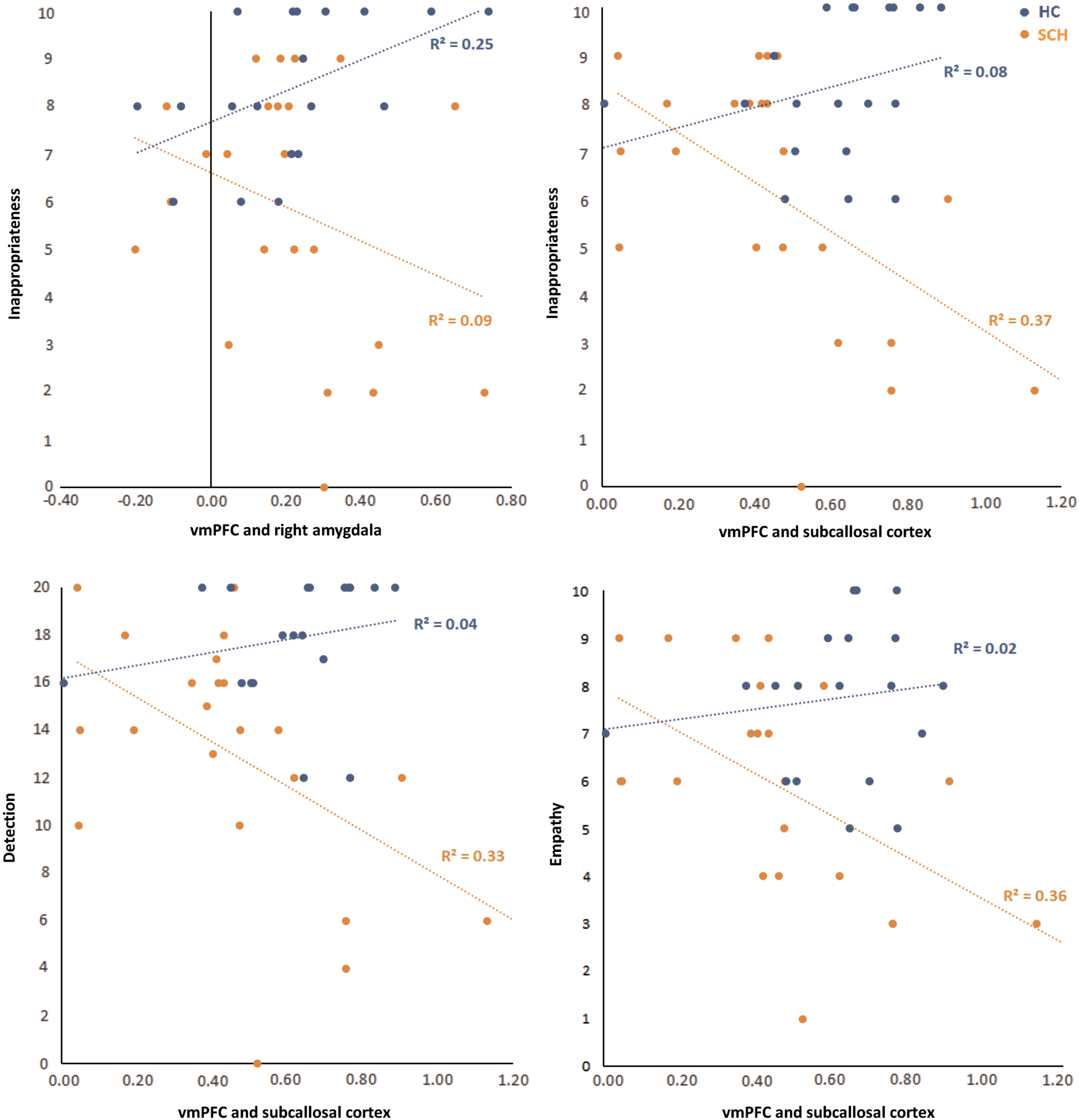
Performance in FPRT as a function of vmPFC seed connectivity strength between regions of interest. Color images available online at
Resting-state vmPFC connectivity related to social cognition: within-group results
HCs exhibited connectivity between the vmPFC and several limbic structures, which positively correlated with their performance in cognitive aspects of ToM (belief, inappropriateness, and intention). Specifically, these ToM scores were consistently related to neural projections to the right amygdala, and some of them were reflected by additional projections to the right nucleus accumbens (belief), right hippocampus (inappropriateness and intention), and right parahippocampus (Fig. 4 and Table 4). No statistically significant associations between affective scores and vmPFC connectivity at rest and no negative correlations were detected.
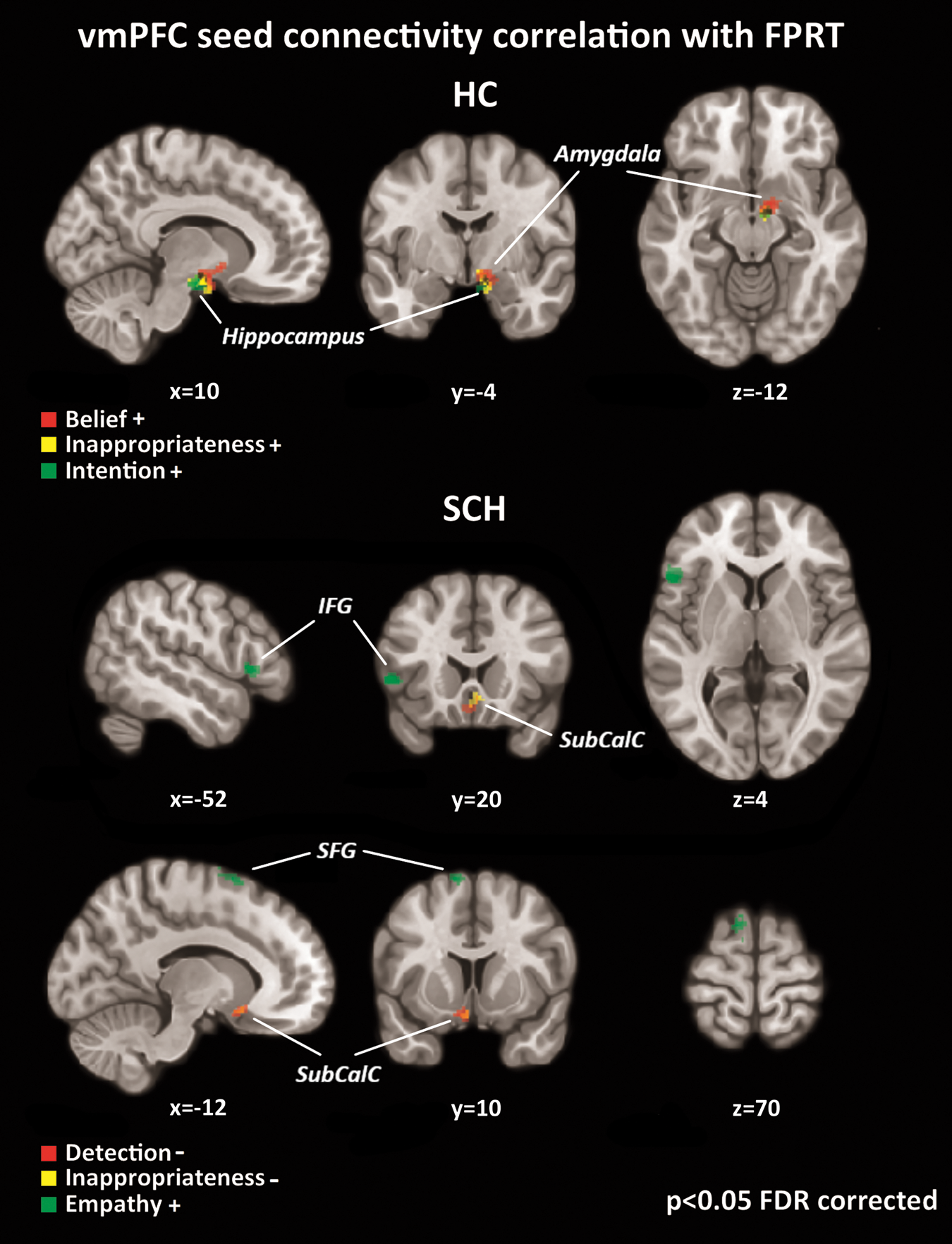
Correlations between resting-state vmPFC seed connectivity and FPRT scores; positive correlation (+) of FPRT scores (belief, inappropriateness, and intention) and vmPFC seed connectivity to limbic regions (right amygdala and right hippocampus) were revealed in HCs; SCH displayed negative correlations (−) of FPRT scores (detection and inappropriateness) and vmPFC seed connectivity to the SubCalC and orbitofrontal cortex, and positive correlation of the FPRT score for empathy and vmPFC connectivity to the left SFG/SMA and left IFG. SFG, superior frontal gyrus; SMA, supplementary motor area. Color images available online at
Resting-State Ventromedial Prefrontal Cortex Seed Functional Connectivity Related to Faux Pas Recognition Task Performance
FPRT, faux pas recognition task; IFG, inferior frontal gyrus; MNI, Montreal Neurological Institute; p-FDR, p-values after false discovery rate correction; R/L, right/left; SFG, superior frontal gyrus.
Different connectivity patterns were identified in the SCH group. Specifically, we observed negative correlations between detection and inappropriateness and connectivity between the vmPFC and the subcallosal and left orbitofrontal cortex (Fig. 4 and Table 4), respectively. In contrast, FPRT empathy scores were associated positively with projections to the left superior frontal gyrus (SFG) and left IFG (Fig. 4 and Table 4).
Discussion
In this study, we examined the resting-state FC network derived from vmPFC seed in SCH patients and HCs. We further investigated the relationship between ToM domains as assessed by the FPRT and these connectivity measures. In addition to the resting-state data and behavioral data from the ToM task, we collected self-reported data through questionnaires related to empathy and emotion regulation. The current cognitive status of all the participants was examined to ensure no clinically significant cognitive impairment was present.
Regarding the behavioral results on social-cognitive task, the SCH patients generally performed worse in FPRT. This applied for both experimental and control stories, indicating that SCH tended to overidentify social FP even in stories that described minor interpersonal conflicts. In line with existing literature on abnormal ToM functioning in SCH (Bora et al., 2009; Jáni and Kašpárek, 2017; Sprong et al., 2007), including studies that used the same task (de Achával et al., 2010), the largest differences were found in intention and belief subscales. Previous studies with SCH patients have reported deficits in empathy to be associated with poor functioning (Michaels et al., 2014), even when controlling for neurocognitive deficits (Smith et al., 2012).
In our study, SCH performed worse than HCs in the majority of cognitive domains. This result appears to provide support for the “hypofrontality” hypothesis in SCH that posits disruption of the frontal lobe functions (e.g., memory, executive functions) in this population (Beatty et al., 1993; Weinberger et al., 1994). Despite statistically significant differences in general cognition between the two groups, the SCH performance is still within the lower average range spectrum and, therefore, does not indicate significant cognitive deficit from a clinical perspective. Even though the FPRT performance should not rely on verbal memory, since the stories are available to participants for the entire task duration, we found that immediate verbal memory capacity was strongly associated with the performance in FPRT intentionality in SCH; no such relationship was detected in HCs. It seems that the verbal memory strongly interacts with the cognitive aspect of ToM skills in our SCH sample.
FPRT is thought to evoke personal experiences similar to those in the FP stories and people often bring their own experiences into the decision-making process. We found evidence of this process in both groups: the participants reported spontaneously how they felt or would have felt in a given situation, but the SCH answers were more frequently inadequate, indicating their limited ability to integrate the information correctly. Such inadequacy seems to be reflected in the association between short-term memory performance and cognitive ToM skills in the patients. This finding is consistent with reports suggesting that memory and ToM have converging networks (Spreng, 2013), and that, therefore, having intact cognitive functions, memory in particular, is essential to mentalize appropriately about other people.
Although SCH performed worse than HCs in both the cognitive and affective components of the FPRT, they reported significantly lower scores in cognitive empathy only. This is consistent with previous findings reported using the same QCAE questionnaire (Michaels et al., 2014). The difference in cognitive empathy in our sample was driven mostly by the perspective taking subscale, which reflects the tendency to see things from another's perspective. Our results indicate that patients with SCH might be over-responsive in some self-reporting methods assessing empathy. A similar discrepancy between self-reporting and behavioral performance in the affective empathy domain has been observed previously (Horan et al., 2015).
Furthermore, affective empathy reported by our HCs correlated significantly with the FPRT performance; there was no such relationship in SCH. No differences between the groups were found in any self-reported measure of emotion regulation in our sample, similarly to previously reported findings (Henry et al., 2008), despite existing evidence of difficulties in emotion management in SCH when assessed behaviorally (Kimhy et al., 2012). The discrepancy here might stem from targeting slightly different aspects of emotion regulation by each of the methods. In addition, the self-reported data indicate that patients do not have adequate insight into their socioaffective skills.
Indeed, it is common for patients with SCH to be unaware of various aspects of their illness that are linked to the impairment of meta-cognitive abilities (Bora, 2017). A lack of insight into SCH appears to be linked to impaired frontal cortical systems. Findings reported by various studies are inconclusive, with the discrepancies arising possibly from the complex nature of insight at various levels and different assessments employed (Ouzir et al., 2012).
Our resting-state connectivity results revealed that the HC group exhibited stronger functional connections between the vmPFC and left IPL than SCH did; SCH had stronger projections from the vmPFC to the right IFG than HCs. Although the majority of previous studies found altered resting-state connectivity of the vmPFC in SCH, specific results reported in the literature are inconsistent, with decreases as well as increases in connectivity (for review, see Karbasforoushan and Woodward, 2012). This abnormal connectivity has been linked to cognitive impairment and predicts the degree of risk of SCH development (Unschuld et al., 2014), supporting the neurodevelopmental theories of SCH as a functional brain network integration and segregation disruption that occurs during brain maturation (Karbasforoushan and Woodward, 2012).
The ToM-based connectivity analysis showed stronger associations in HCs than in SCH of FPRT variables and vmPFC connectivity with limbic regions at rest, namely with the subcallosal cortex, right amygdala, and right hippocampus. The results of hierarchical regression and correlation analysis of vmPFC connectivity and FPRT scores revealed that positive relationships with the limbic regions, most significantly with the right amygdala, were characteristic for HCs, whereas SCH exhibited the opposite connectivity pattern—that is, a negative relationship with the subcallosal region. The only positive relationship detected in SCH was between FPRT empathy score and corticocortical connectivity within the frontal lobe (left IFG and left SFG/supplementary motor area [SMA]).
The frontolimbic circuit serves a variety of functions, ranging from memory processing (Eryilmaz et al., 2016), mood regulation (Butler et al., 2012), and emotion-based decision-making (Morgane et al., 2005) to understanding the social behavior of others (Bickart et al., 2014). The circuit is involved in social cognition and requires integration and processing of two aspects: intrinsic information regarding self-awareness and extrinsic information about the outer social world. The amygdala in particular has been identified as an important structure in this multimodal information processing (Bickart et al., 2014). It has been shown, for example, that the connectivity between the amygdala and other brain regions was reduced in SCH when processing emotional and social information, compared with HCs. Specifically these regions were prefrontal (Bjorkquist et al., 2016), hippocampal, and parahippocampal (Mukherjee et al., 2014). These areas are known to link processing of social stimuli with previous experiences in episodic memory (Maguire et al., 2001). As such, it has been suggested that this connectivity impairment might contribute to social cognitive deficits in SCH (Mukherjee et al., 2014).
In contrast, no significant difference between SCH and HCs in resting-state connectivity between the vmPFC and limbic structures was found in our sample. In other words, there was no indication of frontolimbic hypoconnectivity in SCH per se. Different connectivity patterns were revealed when sociocognitive performance was added into the model, however, whereby HCs displayed projections to limbic regions such as the amygdala, hippocampus, and parahippocampus, but SCH did not.
Furthermore, the relationship between frontoamygdalar connectivity and performance in FPRT seems to support the notion that complex processing of social information requires understanding of emotions at multiple levels. Specifically, it indicates that not only detecting but also understanding the inappropriateness of a remark requires the ability to mentalize that another person would feel hurt.
In SCH, the detection and correct identification of inappropriate behavior in FPRT were negatively correlated with the connectivity between the vmPFC and subcallosal, as well as orbitofrontal cortex. Typically, the subcallosal and orbitofrontal cortex together with the mPFC are important in emotion-based decision-making (Bechara, 2000); however, our results suggest that the interaction between these regions is “counterproductive” in the context of social cognition in SCH.
Conversely, increased resting-state connectivity from the vmPFC to the left SFG/SMA and left IFG was associated with better performance in FPRT empathy scores in SCH. It has been shown that the activation of IFG, anterior cingulate cortex, and SMA is generally associated with affective empathy (Fan et al., 2011). This association is suggested also by our data—vmPFC connectivity with two of these core regions is associated with the empathy level in SCH. This is similar to previous findings showing that peak activation in the SMA during an empathy paradigm correlated with social competence and social attainment in SCH patients, but not in controls (Smith et al., 2014).
It seems that although the vmPFC connectivity to limbic regions at rest is comparable between SCH and HCs, the differences of vmPFC connectivity patterns that arise in relation to ToM processes might represent a different setup for social information processing. The activation or connectivity with the SMA might be a specific biomarker of sociocognitive-affective functions in SCH.
Interestingly, the addition of social-cognitive performance into the model revealed novel information regarding resting-state connectivity that might have otherwise remained undetected. This finding highlights the importance of studying resting-state data in connection with behavioral performance. Behavioral performance should be taken into account to observe important individual variations that likely contribute to the differences in connectivity and might be one reason for the high variability of resting-state results in SCH, as reported by Karbasforoushan and Woodward (2012).
As with any study, there are several limitations. First, this is a correlation study of seed-based resting-state data and behavioral data; thus, the result interpretation should account for this and for the fact that there could be latent or confounding variables accounting for the correlations. We found a relationship between verbal memory and FPRT score. Although the general consensus is that social and general cognition are separate constructs in nonclinical populations (Cosentino et al., 2014), this may not be the case in SCH, where encoding of new information is impaired. Finally, we did not find a relationship between antipsychotic medication dosage and scores in general and social cognition; however, there was a relationship between disease duration and memory function, suggesting that the chronic state may be linked with disrupted performance.
Conclusion
We observed similar vmPFC resting-state connectivity to limbic structures in patients and controls. In other words, no indication of frontolimbic hypoconnectivity in SCH per se was observed. The addition of sociocognitive performance into the model revealed differences in connectivity patterns. Specifically, the frontolimbic connectivity was positively correlated with cognitive aspects of ToM in the HC group, whereas the SCH group exhibited a negative relationship between FPRT scores and the subcallosal region. An increased corticocortical connectivity within the frontal lobe was linked with better empathy skills in SCH. We assume that the differences in the vmPFC connectivity pattern that arise from the relationship with ToM processes might represent a different setup for processing social information in the two groups. Our results thus stress the importance of studying resting-state connectivity in relation to behavioral performance.
Footnotes
Acknowledgment
This study was supported by Ministry of Health of the Czech Republic under the project Conceptual Development of Research Organizations (FNBr, 65269705).
Author Disclosure Statement
No competing financial interests exist.