
Abstract
This study examines the resting-state functional-connectivity (RsFc) in young adults with high-functioning autism spectrum disorder (HF-ASD) using state-of-the-art fMRI data acquisition and analysis techniques. High temporal resolution fMRI using simultaneous multi-slice acquisition aided unbiased whole-brain connectome-wide multivariate pattern analysis (MVPA) techniques for assessing RsFc. MVPA revealed two clusters (Crus I/II and lobule IX) of abnormal connectivity in the cerebellum that are consistent with the notion of a triple representation of nonmotor processing in the cerebellum. Whole-brain seed-based RsFc analyses informed by these clusters showed significant under connectivity between the cerebellar and social, emotional, and language brain regions in the HF-ASD group compared to healthy controls. The results we report are coherent with existing structural, functional, and RsFc literature in autism, extend previous literature reporting cerebellar abnormalities in the neuropathology of autism, and highlight the cerebellum as a potential target for therapeutic, diagnostic, predictive, and prognostic developments in HF-ASD. The description of functional connectivity abnormalities reported in this study using whole-brain, data-driven analyses has the potential to crucially advance the development of ASD biomarkers, targets for therapeutic interventions, and neural predictors for measuring treatment response.
Introduction
Autism spectrum disorder (ASD) is a highly morbid neurodevelopmental disorder characterized by deficits in social–emotional functioning along with restricted repetitive behaviors (APA, 2013). It is estimated to affect up to 2% of children and adolescents in the general population (Blumberg et al., 2013). Previous network-wide analyses, predominantly conducted in children and adolescent populations, have reported significant functional connectivity differences in whole-brain resting-state networks in ASD. Default mode network (DMN) is the most studied resting-state functional connectivity (RsFc) network in ASD, predominantly revealing intra-DMN underconnectivity in ASD (Padmanabhan et al., 2017).
A recent review of ASD RsFc MRI studies reported findings of both over- and underconnectivity in ASD (Hull et al., 2016), concluding that ASD resting-state functional connectivity alterations are inconsistent across studies or have not been robustly characterized. This largely nonconclusive state in the ASD literature is likely related to the inherent complexities of the disorder, as well as the methodological approaches adopted in some investigations of ASD. In addition to a wide range of variability in symptom profiles and severity, ASD frequently presents with comorbid symptoms characteristic of ADHD, anxiety, and mood disorders (Doshi-Velez et al., 2014; Joshi et al., 2010, 2013, 2014; Lenroot and Yeung, 2013).
Some previous studies (Uddin, et al., 2013) have provided hypothetic models regarding RsFc development patterns in HF-ASD, especially hyperconnectivity in children compared with healthy controls (HCs) followed by hypoconnectivity in adolescents and adults. Although early detection is key to developing early interventions for ASD (Emerson et al., 2017), one advantage of studying adult ASD populations is that the patterns of functional connectivity in adults are not susceptible to developmental changes, offering an opportunity to examine an established profile of functional connectivity. Limited studies on functional connectivity are conducted in adult populations with ASD. In a recent publication, while simultaneously examining positive and negative correlations, our research group has documented significant differences in the functional integration (within the major DMN nodes) and segregation (right temporoparietal junction) of the DMN in ASD (Joshi et al., 2017). Furthermore, our neural characterization using pattern recognition analyses has revealed marked heterogeneity in the ASD group. Etiology of ASD is notoriously diverse (Chaste and Leboyer, 2012)—examples range from monogenic diseases to neonatal cerebellar injury, and also include a large proportion of idiopathic cases likely involving numerous complex polygenetic and environmental factors. Taken together, the combination of phenotypic variability, frequent comorbidity, and diverse etiology is a fundamental challenge underlying the study of brain differences in ASD (Hoppenbrouwers et al., 2008).
Despite the task-free nature for data acquisition in RsFc mapping, it is now fairly established that the human functional connectome is highly reliable and replicable (Kelly et al., 2012; Shehzad et al., 2009). Enhanced/optimized acquisition methods and data analysis techniques to tap in and characterize the connectome can result in a wide range of clinical applications. Specifically, challenges inherent in imaging ASD population call for unbiased methodological approaches to ensure that inconsistencies in results are not due to biased data analyses techniques. Methodological approaches adopted in RsFc studies of ASD frequently include a priori seed regions and region of interest (ROI) analyses, which contribute to the generation of inconsistent findings, while amplifying the heterogeneity of findings in an already massively diverse psychiatric entity. Such anatomically constrained analyses may also miss important neural signatures that exist outside of canonical networks, and reveal topographical distributions of abnormality that are a result of methodological impositions. Therefore, the current state of the literature highlights the need for data-driven approaches in the context of this greatly diverse disorder. Unbiased methodological strategies are of capital importance to robustly delineate brain differences, and in this way guide therapeutic, diagnostic, predictive, and prognostic developments in ASD.
Coherent with this vision, the primary objective of this study was to make use of an unbiased whole-brain data-driven approach by using multivariate pattern analysis (MVPA) to investigate RsFc differences between typically developing HCs and adults with high-functioning ASD (HF-ASD). The utility of MVPA-informed RsFc analysis has already been demonstrated in various clinical contexts such as traumatic brain injury (Thompson et al., 2016) and in the context of predicting treatment responses in social anxiety disorder (Whitfield-Gabrieli et al., 2016). A reduction in the number of impositions to statistical methods increases the requirements of data quantity and quality—the data that we used in this study (Autism Brain Imaging Data Exchange [ABIDE], Di Martino et al., 2014) provided an unprecedented opportunity to overcome this barrier. An exceptional subsecond temporal resolution, emerging from simultaneous multislice (SMS) acquisition, offered an additional level of advantage by curbing the detrimental effects of physiological noise-related contamination of low-frequency fluctuations, thereby providing enhancements in statistical power. In addition, whole-brain coverage and overall improved temporal signal-to-noise ratio (which is directly proportional to BOLD contrast-to-noise ratio) capabilities of multichannel array coils were crucial for our implementation of an analysis that conforms with the data-driven methods necessary to successfully advance toward a robust characterization of the neural basis of ASD subtypes.
Methods
We carried out a whole-brain data-driven MVPA on the ABIDE data set (N = 59) contributed by principal investigator Michal Assaf, MD (Olin Neuropsychiatry Research Center [ONRC], Institute of Living, Hartford Hospital and Yale School of Medicine, Department of Psychiatry). All ASD participants met criteria for autism on the Autism Diagnostic Observation Schedule-Generic (ADOS-G). History of ASD and psychopathology was ruled out in HCs by administering the ADOS-G and the Structured Clinical Interview for DSM-IV Axis I Disorders-Research Version (SCID-I RV), respectively. Detailed information on the assessment and recruitment procedures is provided in the release website (
Data acquisition
Resting scan images were collected on 3T Siemens Skyra (TR/TE/flip angle were 475 ms/30 ms/60°, with 3 mm voxels, multiband factor of 8, 947 time points). These high-temporal resolution parameters provided unprecedented statistical analysis power and denoising capabilities, facilitating the implementation of a fully unbiased data-driven analysis. T1-weighted structural images were acquired using 3D-MPRAGE (TR/TE/TI/flip angle were 2200 ms/2.88 ms/794 ms/13°, with 0.8 mm voxels).
Data analysis
Before RsFc analysis, data were realigned, normalized (to MNI space with respect to the echo planar imaging template), and spatially smoothed with a 6-mm kernel using SPM12 (Wellcome Department of Imaging Neuroscience, London, United Kingdom;
Multivariate pattern analysis
For unbiased identification of seed regions for standard seed-to-voxel analysis, we employed whole-brain connectome-wide group MVPA, as implemented in the CONN Toolbox (Whitfield-Gabrieli and Nieto-Castanon, 2012). For each voxel, a low-dimensional representation of the entire pattern of seed-based correlations between this voxel and the rest of the brain is derived as explained in (
For dimensionality reduction, group MVPA estimates a multivariate representation of the connectivity pattern by computing the pairwise connectivity pattern between each voxel and the rest of the brain, and using principal component analysis (PCA) to characterize those patterns using a small number of components in a two-step process. In the first PCA step, separately for each subject, 64 PCA components are retained while characterizing each subject voxel-to-voxel correlation structure. This is a form of subject-level dimensionality reduction typically used in independent component analysis applications [e.g., (Calhoun et al., 2011)]. In the second PCA step, jointly across all subjects but separately for each voxel, the three strongest components were retained from a principal component decomposition of the between-subjects variability in seed-to-voxel connectivity maps between this voxel and the rest of the brain. An F-test was performed on all three MVPA components (which explain the maximum intersubject variability) simultaneously in a single second-level analysis to identify the voxels that show significant differences in connectivity patterns between the two groups. This is an omnibus test (equivalent to seed-level F-test in ROI-to-ROI analyses) to identify abstract multivariate representations [e.g., (Shehzad et al., 2014)]. The resulting clusters from a height-level statistical threshold of p < 0.001 and cluster threshold of p < 0.05 family-wise error (FWE)-corrected and k > 50 were retained for post hoc characterization using seed-to-voxel bivariate correlation to explore what particular aspects of functional connectivity differ across groups. This was done by first producing the time course from the MVPA-derived cluster of interest, and computing Pearson's correlation coefficients between that time course and the time course of all other voxels. Correlation coefficients were then converted to normally distributed z-scores using the Fisher transformation to improve the validity of second-level general linear model analyses. A height threshold of whole-brain p < 0.001 (T = 3.24) and false discover rate (FDR)-corrected cluster threshold of p < 0.05 (nonparametric statistics) were used for between-group characterization. Finally, as an exploratory analysis within the HF-ASD group, Fisher-transformed r-maps from the MVPA-derived clusters of interest were subjected to a second-level analysis regressing the whole-brain functional connectivity correlations with total ADOS scores (height threshold of whole-brain p < 0.005 and FDR-corrected cluster threshold of p < 0.05).
Results
Our study included 24 ASD participants (20 males, mean age = 21.1 ± 2.9; 4 females, mean age = 24.4 ± 1; range = 18–31 years) and 35 HCs (20 males, mean age = 24.2 ± 3.5; 15 females, mean age = 23.9 ± 2.7; range = 19–30 years). Among the ASD participants, 25% (6/24) had a psychiatric comorbidity. Subject demographics, quality control metrics, and psychiatric comorbidity with ASD are summarized in Table 1.
Demographics and Psychiatric Comorbidity with Autism Spectrum Disorder
Values expressed as n (%) or mean ± standard deviation.
ASD, autism spectrum disorder; ADOS-G, autism diagnostic observation schedule-generic; HCs, healthy controls; IQ, intelligence quotient; n.s., not significant; N/A, not applicable.
The psychotropic treatment status of the 24 ASD participants at the time of scan was as follows: 17 ASD participants were medication naive; 7 participants were receiving combination therapy for the management of associated psychopathologies with serotonin reuptake inhibitors (N = 4), stimulant class of anti-ADHD medication (N = 3), atypical antipsychotics (N = 3), and benzodiazepines (N = 2).
MVPA results
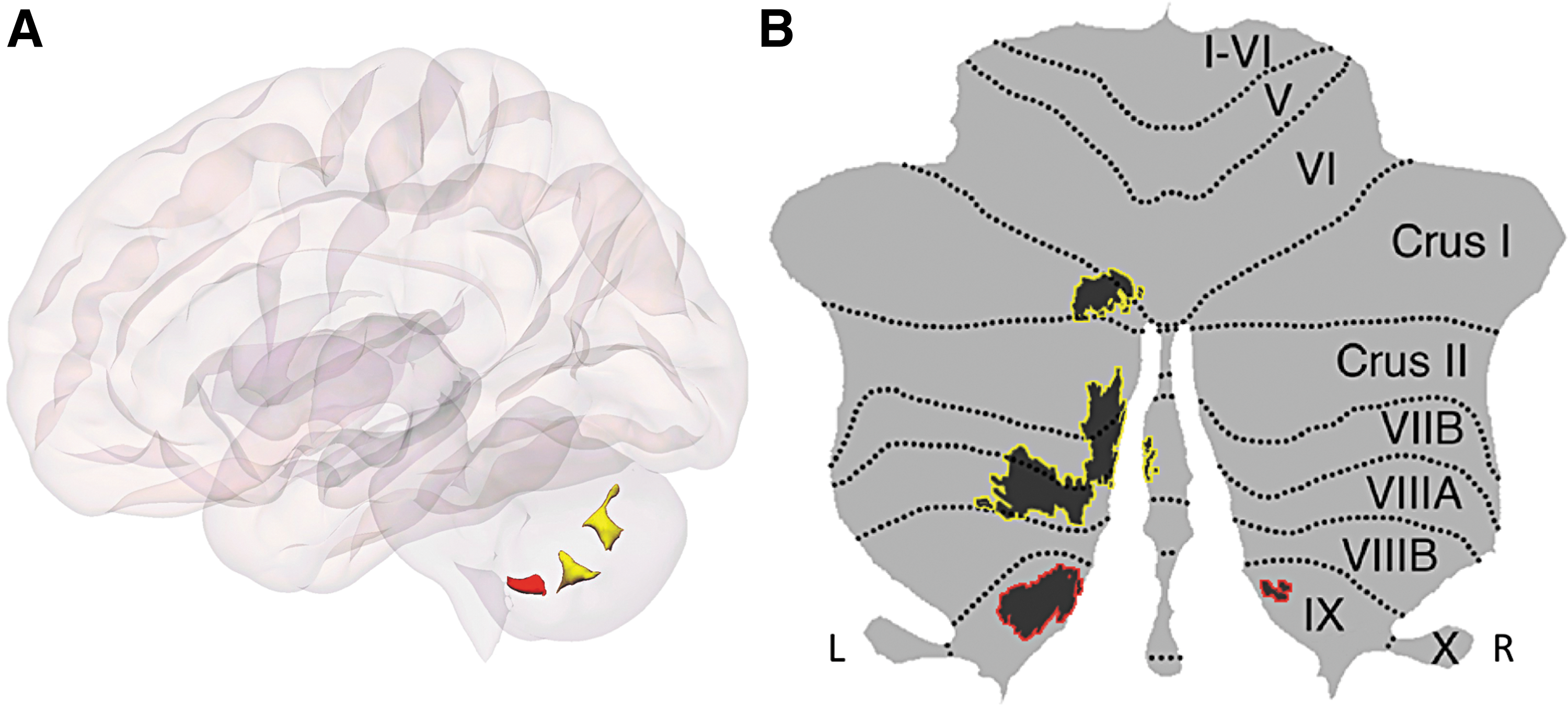
Statistically significant clusters resulting from the whole-brain connectome-wide MVPA are shown in Fig. 1A. Whole-brain MVPA results with significant differences between individuals with ASD and HCs in two cerebellar clusters are visualized on a flat map representation in Fig. 1B. Cluster 1 (shown in yellow) is located in the left cerebellum (Crus I and II, extending to lobules VIIB and VIIIA as well as lobule VI) and cluster 2 (shown in red) included bilateral lobule IX.
Post hoc seed-to-voxel analysis of MVPA-derived clusters of interest
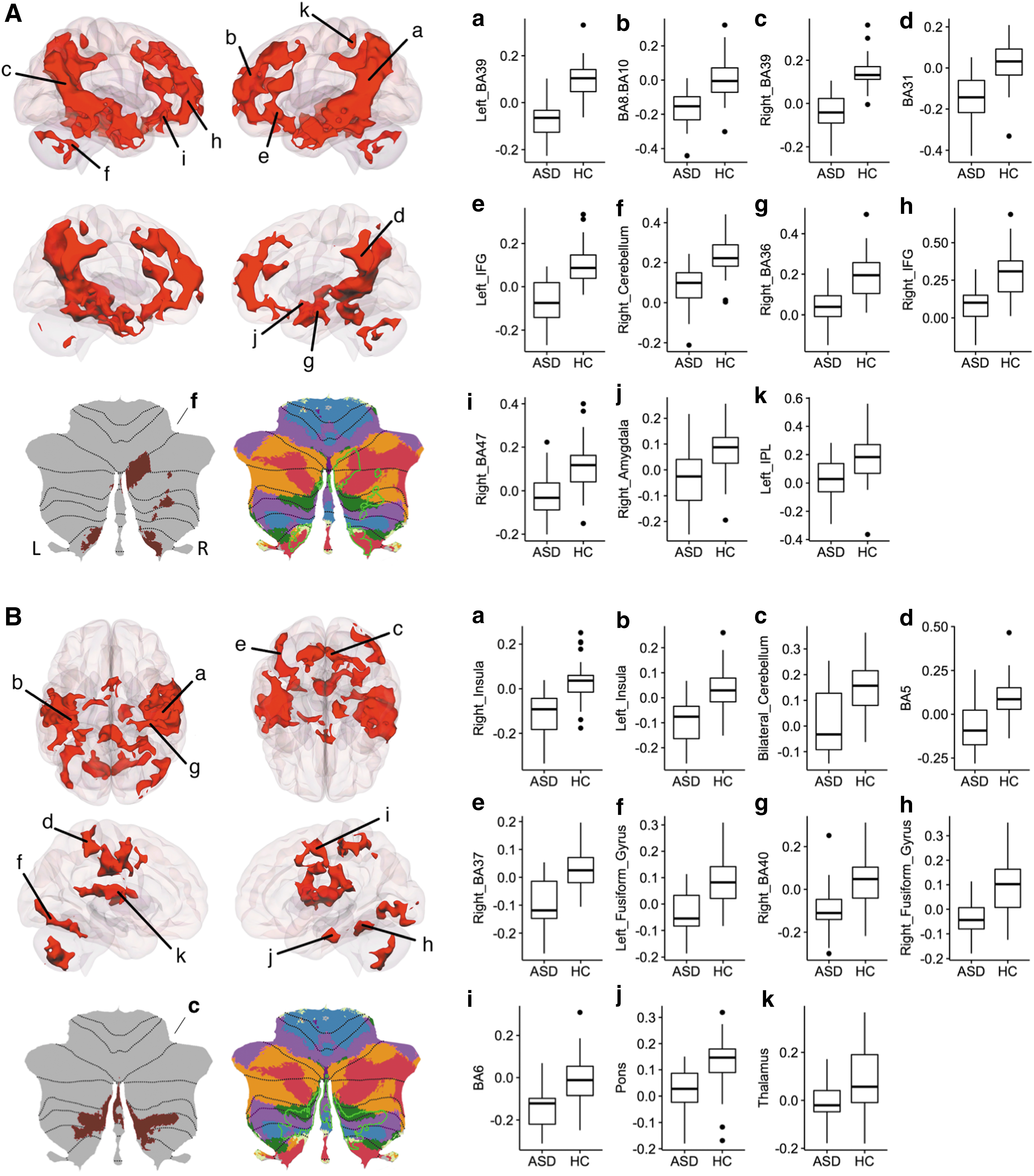
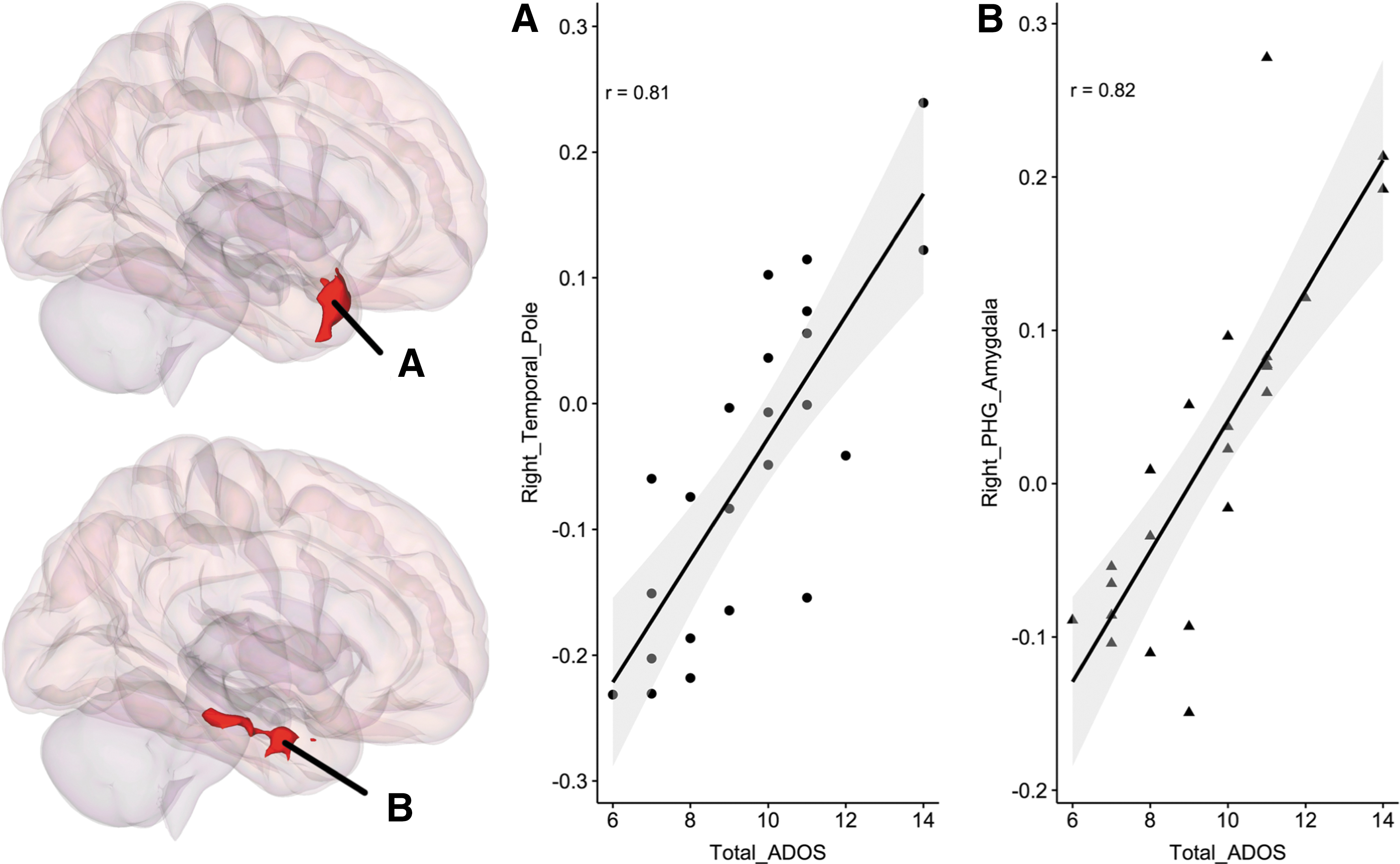
Results using MVPA-derived clusters are shown in Fig. 2A and B. Cerebellar clusters from Fig. 2A (labeled as f) and B (labeled as c) are also represented on a flat map for better visualization. Results are summarized in Table 2. This analysis revealed underconnectivity both within the cerebellum and between the cerebellum and cerebral cortical regions in individuals with HF-ASD. Although there was marked individual variability in both ASD and HC groups, RsFc strength using the cerebellar clusters as seed regions correlated with total ADOS scores within the ASD group, specifically within right temporal pole and right amygdala with cluster 1 and cluster 2, respectively (Fig. 3).
Results from second-level seed-to-voxel RsFc analysis for HCs>ASD contrast (opposite contrast was nonsignificant) for MVPA-derived cluster 1
Whole-brain functional connectivity correlations with total ADOS scores for MVPA clusters 1 and 2 are shown in
Results from Second-Level Seed-to-Voxel RsFc Analysis of MVPA-Derived Clusters for HCs>ASD Contrast (Opposite Contrast Was Nonsignificant)
A height threshold of whole-brain p < 0.001 (T = 3.24) and FDR-corrected cluster threshold of p < 0.05 (nonparametric statistics) were used for characterization.
MVPA, multivariate pattern analysis; RsFc, resting-state functional connectivity.
Discussion
In this study, we adopted a completely data-driven and unbiased, connectome-wide MVPA approach to examine whole-brain RsFc in ASD. Using a synergistic combination of 32 channel head coil and high-temporal resolution SMS imaging, we provide robust evidence of cerebellocerebral disruptions in HF-ASD. Our unbiased MVPA revealed two clusters in the cerebellum: (1) a left cerebellar cluster (Crus I/II, extending to lobules VIIB/VIIIA as well as lobule VI, and including vermal aspects of these structures) and (2) bilateral lobule IX. Post hoc investigations revealed disrupted connectivity between cerebellum and social, emotion, and language processing regions in the cerebral cortex. In addition, strength of cerebrocerebellar functional connectivity correlated with ADOS scores in the ASD group. The results of our unbiased MVPA are in alignment with existing structural, functional, and RsFc literature in autism, and extend previous literature reporting cerebellar abnormalities in the neuropathology of autism (Bauman and Kemper, 2003).
Multivariate pattern analysis
Evidence from multiple fields of study has established that the cerebellum is involved not only in motor but also in cognitive and affective processing (Koziol et al., 2014). The cerebellum is anatomically connected to motor and nonmotor regions of the extracerebellar cortical and subcortical structures (Bostan and Strick, 2010; Kelly and Strick, 2003; Schmahmann, 1996; Schmahmann and Pandya, 1997), and cerebellar lesions lead to a cerebellar motor syndrome as well as to a cerebellar cognitive affective syndrome (Guell et al., 2015; Hoche et al., 2018; Schmahmann and Sherman, 1998), including deficits in social processing (Hoche et al., 2016). Neuroimaging investigations reveal cerebellar involvement in nonmotor RsFc and task processing (Buckner et al., 2011; Guell et al., 2018b; Stoodley et al., 2012), and numerous studies have described cerebellar structural and functional abnormalities in neurological and psychiatric conditions that degrade cognition and affect [e.g., (Guo et al., 2016; Phillips et al., 2015)]. Not surprisingly, multiple studies signal that the cerebellum may play a crucial role in the pathophysiology of ASD (Fatemi et al., 2012; Rogers et al., 2013; Stoodley et al., 2017; Wang et al., 2014). Specifically, structure and function of left and right Crus I/II have been associated with ASD symptom severity across subtypes of ASD (D'Mello and Stoodley, 2015; D'Mello et al., 2016; Riva et al., 2013; Stoodley et al., 2017). Also coherent with the distribution of our cerebellar results, vermal aspects of the cerebellum have been classically regarded as the “limbic cerebellum” (Schmahmann, 1991), and multiple neuroimaging studies have identified emotion processing task activation at the cerebellar vermis (Baumann and Mattingley, 2012; E et al., 2014;Stoodley and Schmahmann, 2009). In many task-based functional magnetic resonance imaging (fMRI) studies, aberrant cerebellar activation corresponds with structural abnormalities in ASD [see (Becker and Stoodley, 2013) for review]. In addition, cerebellar midline manipulation results in emotion modulation alterations in animals and humans (Berman et al., 1978; Heath, 1977; Levisohn et al., 2000), and clinical studies have revealed cerebellar vermal involvement in patients with pronounced affective symptomatology (Gudrunardottir et al., 2016; Levisohn et al., 2000; Schmahmann and Sherman, 1998). Furthermore, reduced volume of the posterior vermis was one of the first reported cerebellar differences in ASD (Courchesne and Magasanik, 1988).
Our MVPA also revealed abnormal functional connectivity in bilateral lobule IX. In addition to Crus I/II, abnormal function and structure in lobule IX are consistently reported in neuroimaging studies of ASD (D'Mello and Stoodley, 2015; Stoodley, 2014). Although lobule IX is considered important for visual guidance of movement (Stoodley and Schmahmann, 2010; Voogd and Glickstein, 1998), multiple RsFc analyses have reported lobule IX connectivity with cerebral cortical regions corresponding to the DMN (Buckner et al., 2011; Habas et al., 2009; Krienen and Buckner, 2009; O'Reilly et al., 2010). Coherent with these observations, viral tract tracing work revealed that lobule IX is anatomically connected to the prefrontal cortex (Kelly and Strick, 2003), and neuroimaging investigations have reported lobule IX engagement in emotion task processing (E et al., 2014; Schraa-Tam et al., 2012) as well as other nonmotor processes such as working memory (Desmond et al., 1997) and timing perception (Liu et al., 2008).
Our Crus I/II and lobule IX results are consistent with the notion of a triple representation of nonmotor processing in the cerebellum. Motor processing engages lobules I–VI as well as lobule VIII (Stoodley and Schmahmann, 2010). This organization has been classically described as a cerebellar “double motor representation” (first motor representation, lobules I–VI; second motor representation, lobule VIII) (Woolsey, 1952). Recent studies have described an analogous pattern of three nonmotor representations (VI/Crus I, first nonmotor representation; Crus II/VIIB, second nonmotor representation; lobules IX/X, third nonmotor representation). Specifically, this pattern of triple nonmotor representation has been observed in studies of cerebellar functional connectivity with cerebral cortical resting-state networks (Buckner et al., 2011), cerebellar task activation (Guell et al., 2018a), and cerebrocerebellar seed-based resting-state functional connectivity (Guell et al., 2018a). Of note, first and second nonmotor representations are sometimes contiguous [e.g., language task processing in Guell et al., 2018a, or DMN in Buckner et al. (2011)] and sometimes separate [e.g., working memory task processing in Guell et al., 2018a, or frontoparietal network in Buckner et al. (2011)]. Under this view, our Crus I/II MVPA cluster corresponds to the area of first and contiguous second nonmotor representation, and our lobule IX MVPA cluster corresponds to the area of third nonmotor representation.
This is the first study to discuss cerebellar pathological findings in relation to the triple nonmotor representation hypothesis. This approach is coherent with previous studies describing similarities between pathological and physiological topographical distributions in neuroimaging (Seeley et al., 2009; Zhou et al., 2012). In line with this concept, our description expands the notion of a triple nonmotor organization in the cerebellum by showing that it is observable not only in healthy brain processes such as task activation and functional connectivity but perhaps also in pathological processes such as cerebellar regions with abnormal whole-brain connectivity in an ASD cohort. The functional significance of the triple nonmotor organization is considered to be one of the principal outstanding enigmas in recent reviews of cerebellar neuroscience (Sokolov et al., 2017), and preprints in the literature have very recently started to hint at potential functional properties of the areas of first, second, and third nonmotor representation (Guell, 2018). This study contributes to this line of inquiry by noting that the triple nonmotor organization may also be observed in pathological functional changes in psychiatry, and highlights the need to investigate the functional significance of this organization in health and disease.
Taken together, cerebellar abnormalities revealed in our unbiased data-driven approach strongly suggest a crucial cerebellar role in the pathophysiology of ASD, supporting and extending previous literature. Of particular note, no other brain regions survived the impositions of our analysis, suggesting that cerebellar differences are especially robust. Correlations with ADOS clinical scores within our HF-ASD cohort further support the validity and relevance of our observation and hint at the possibility that cerebellar functional connectivity disruptions may not only constitute a crucial aspect of ASD brain abnormalities but also a dynamic feature related to variability of symptom profile and severity. ASD-like symptoms after cerebellar injury or degeneration in neurology have been classified as overshoot (stereotypical behaviors and self-stimulation behaviors) and undershoot phenomena (avoidant behaviors, tactile defensiveness, and easy sensory overload) (Schmahmann et al., 2007). The results of this study support cerebellar involvement in ASD, and thus suggest a potentially relevant position of “Dysmetria of Thought” as a conceptual framework for future studies investigating the nature of ASD symptomatology in psychiatry. This theory holds that motor, cognitive, and affective symptoms that arise from cerebellar abnormalities are a reflection of a singular neurological dysfunction. At a physiological level, Dysmetria of Thought is predicated on the concept of a Universal Cerebellar Transform, which hypothesizes that one single neurological process subserves cerebellar modulation of movement, thought, and emotion (Schmahmann, 1991, 1996; Schmahmann, 2010); see also a recent review in Guell et al. (2018a). This physiological theory emerges from two contrasting anatomical realities of the cerebellum—on one hand, cerebellar cortical cytoarchitecture is largely uniform (Ito, 1993; Voogd and Glickstein, 1998); on the other hand, distinct cerebellar regions are anatomically connected with distinct motor and nonmotor extracerebellar regions (Dum and Strick, 2003; Schmahmann and Pandya, 1997, 2008). A common computation thereby emerges from a homogeneous cytoarchitecture, accesses distinct streams of information processing as a result of heterogeneity of anatomical connectivity, and in this way manifests as different functions. Within the framework of the Universal Cerebellar Transform and Dysmetria of Thought hypotheses, nonmotor symptoms observed after cerebellar injury or degeneration in neurological patients are conceptualized as nonmotor analogues of cerebellar motor ataxia. Thus, at a larger scale, the observation of robust cerebellar functional disruptions in this study is coherent with a large volume of literature connecting cerebellum and psychiatry, and resonates with the broader unifying concept of Dysmetria of Thought.
RsFc characterization from MVPA-derived cluster 1
Post hoc seed-to-voxel analyses of MVPA cluster 1 (left Crus I/II and lobules VIIB–VIIIA) yielded underconnectivity with social brain regions in the HF-ASD group including DMN (bilateral AG, ITG, precuneus, and mPFC), language regions including bilateral STG and IFG, and regions important in emotion (amygdala). Importantly, social interaction deficits are a core diagnostic criterion in ASD (American Psychological Association, 2013). Regions that emerged as significantly different in the HCs>ASD contrast overlap with the recent meta-analyses on social brain regions as revealed by both task and resting-state fMRI (defined as “social brain atlas,” including: right IFG, bilateral hippocampus, rostral ACC, ventromedial and dorsomedial PFC, bilateral amygdala, bilateral nucleus accumbens, bilateral MTG, precuneus/PCC, bilateral TPJ, bilateral temporal pole, posterior-mid cingulate cortex, bilateral MT/V5, bilateral fusiform gyrus (FG), bilateral STG, left SMA, bilateral anterior insula (AI), bilateral SMG, and bilateral cerebellum, (Alcalá-López et al., 2017). In ASD, specifically, abnormal pSTG activation and connectivity have been associated with social deficits (Redcay et al., 2013) and deficits in emotion recognition (Alaerts et al., 2014). Underconnectivity of cerebellar regions with DMN (specifically, left Crus I/II) is also consistent with our recent publication in young adults with ASD (Joshi et al., 2017).
Post hoc analysis from our MVPA-derived left cerebellar cluster also identified disrupted functional connectivity within the cerebellum. The ASD group showed decreased connectivity with right cerebellar regions (right Crus I/II, VIIB, and bilateral lobule IX). These results support previous literature finding decreased Purkinje cells, abnormal structure and function, and aberrant resting-state connectivity from right Crus I/II in ASD (Bauman and Kemper, 2005; D'Mello et al., 2015; Fatemi et al., 2002; Khan et al., 2015; Stoodley et al., 2017). Right Crus I/II has been implicated in social processing (Van Overwalle et al., 2014) and previous examinations of cerebellocerebral resting state in ASD have reported reduced connectivity between Crus I/II and cerebral regions implicated in social cognition and language (Khan et al., 2015). The localization and specificity of our left cerebellar results from MVPA and post hoc characterization of whole-brain group differences are especially striking, as our cerebellar cluster was derived from unbiased methods, and seeding this region revealed group differences within the social/language network in cerebellar and cerebral cortical structures. In particular, both the left cerebellar cluster identified by MVPA and post hoc identification of right cerebellar homologues are significant in the context of the functional topography proposed for the cerebellum, whereby posterior cerebellar regions are associated with social and language behaviors (Stoodley et al., 2012) and functionally connected to social and language networks (Buckner et al., 2011; Van Overwalle et al., 2014). Previous studies have found that connectivity between left Crus I/II and bilateral posterior superior temporal regions is associated with animacy attributions (Jack & Pelphrey 2015), providing further support that this network may be involved in imitation and other behaviors that may be important for the development of proper social interaction. Neuromodulation of right posterior cerebellar regions by transcranial magnetic stimulation has been used to demonstrate connectivity changes with other cerebral regions important in imitation, and can rescue social impairments in a mouse model of ASD (Stoodley et al., 2017).
RsFc characterization from MVPA-derived cluster 2
Post hoc seed-to-voxel analyses of MVPA cluster 2 (bilateral lobule IX) yielded significant underconnectivity with a network of brain regions including the bilateral AI, bilateral fusiform gyrii (BA 37), somatosensory regions (BA 5), and thalamus in the HF-ASD group. The ventral and dorsal aspects of the AI have been associated with empathy (Singer et al., 2004) and cognitive control (Nelson et al., 2010), respectively. Similar to connectivity results from cluster 1, the insula is one of the most consistently identified social processing brain regions, and hypoactivity in the insula has been reported in task-related fMRI studies in autism (Uddin and Menon, 2009). Network analyses indicate that the AI acts as a hub, mediating interactions between large-scale networks involved in externally and internally oriented cognitive processing (Uddin et al., 2013). Furthermore, a meta-analysis of neuroimaging studies of social cognition revealed consistent hypoactivation of the AI in ASD (Uddin and Menon, 2009). Previous studies have reported that RsFc of insula is reduced in ASD (Paakki et al., 2010) and negatively correlated with autistic symptoms (Lai, et al., 2015). Hypoactivity in FG is known in the fMRI literature of ASD, and there is postmortem evidence that FG neurons are fewer and smaller in autism (van Kooten et al., 2008). The cerebellocerebral differences in RsFc we report hone in on the mounting evidence of cerebellar deficits in ASD (Hampson and Blatt, 2015). As with cluster 1, post hoc analysis of MVPA-derived cluster 2 identified significantly decreased RsFc within the cerebellum for the ASD group, with cerebellar clusters spanning bilateral VIIB–VIIIB.
Behavioral correlations
Given the large heterogeneity in ASD (Doshi-Velez, et al., 2014; Joshi et al., 2010, 2013, 2014; Lenroot and Yeung, 2013), distinct neural substrates may be associated with distinct ASD symptoms and/or severity. In this study, we report that increased functional connectivity between the left cerebellum and right-lateralized language/social regions such as the temporal pole and amygdala is associated with increased total ADOS scores, reflecting both the range and severity of symptomatology. Reduced emotional awareness in ASD patients has been shown to be associated with a dysfunctional limbic system, in particular amygdala (Silani et al., 2008). Previous research has also related language delay phenotypes (early language delay vs. no early language delay) in ASD with lateralization of cerebellar differences (D'Mello et al., 2016; Lai et al., 2015), and shown that decreased gray matter in specific cerebellar lobules is associated with differing core ASD symptoms (D'Mello et al., 2015). However, in this study, a full behavioral characterization was not available and, therefore, links between functional connectivity and specific ASD symptoms cannot be made. Future research should examine how specific behavioral phenotypes are associated with cerebellar functional connectivity differences.
Limitations and future work
This study used a combination of behavioral and neuroimaging techniques to assess neural substrates of ASD. We report significant correlations between cerebrocerebellar connectivity strength and ADOS total score. ADOS total score is a composite score of core symptoms of ASD, including social communication impairments and the presence of repetitive or restricted behaviors. Unfortunately, individual subscales of the ADOS (social communication; repetitive behaviors) were not available and, therefore, no hypotheses regarding the association of connectivity with specific behaviors could be tested. However, the ADOS total score is able to capture the complete ASD phenotype, and is relevant to clinical diagnosis. Future research should examine associations between functional connectivity in specific circuits, and scores on individual ADOS subscales.
This study was performed in adults. Many previous studies finding cerebellar contributions to ASD have been performed in children and adolescents (e.g., (D'Mello et al., 2015, 2016; Khan et al., 2015). Previous studies have reported that cerebellar structure–behavior relationships change as a function of age (Moore et al., 2017), and that cerebellar subregions reach peak development at different times (Taki et al., 2013; Tiemeier et al., 2010). In this study, cerebellar differences in ASD are similar to those found in child and adolescent studies, suggesting that cerebellar dysfunction in ASD may occur independently of age. However, to our knowledge, no studies have examined the effects of age on cerebellar contributions to ASD symptoms. Future studies should recruit longitudinal samples to examine this question.
Previous studies have reported both increased and decreased cerebrocerebellar functional connectivity in ASD (e.g., Khan et al., 2015; Verly et al., 2014), and future work should investigate how increased versus decreased cerebrocerebellar connectivity relates to ASD symptoms. Importantly, the clusters identified in the present analyses (Crus I/II, lobule VIII, and lobule IX) have all been identified as the most consistently abnormal regions in meta-analyses of ASD (Stoodley, 2014). Given that altered structure, function, and connectivity from bilateral Crus I/II are some of the most consistently reported differences in ASD [see Crippa et al. (2016); D'Mello and Stoodley (2015); Sokolov et al. (2017) for review], these regions may be promising candidates for neuromodulation or targeted therapies. Indeed, recent research suggests that cerebellar neuromodulation of these specific subregions could be effective in altering ASD symptoms (Stoodley et al., 2017). Our left lateralized finding of significantly different Crus I/II RsFc is strikingly consistent with recent reports of underconnectivity between left Crus II and rTPJ's dorsal subdivision of anterior AG, a region attributed for mediating social cognition (D'Mello and Stoodley, 2015; Doyle-Thomas et al., 2015; Igelstrom et al., 2016).
Future research may test the generalizability of our findings to other ASD cohorts. Such studies may investigate differences in connectivity in independent data sets seeding from clusters resulting from our MVPA, and in addition aim to predict ASD versus HC group membership using cross-validation approaches (Takagi et al., 2017). In addition, future studies may explore differences between cerebellar abnormalities in lobules IX/X (area of third nonmotor representation) compared with cerebellar abnormalities in lobule VI/Crus I (area of first nonmotor representation) and Crus II/VIIB (area of second nonmotor representation) in ASD. The functional significance of the cerebellar triple nonmotor organization has not been explored in depth yet [(Buckner et al., 2011; Guell et al., 2018b; Sokolov et al., 2017); see a preprint in (Guell, 2018) for initial hypotheses], and advances in this line of inquiry might improve our understanding of cerebellar functional abnormalities in ASD.
Conclusions
Although the cerebellum is one of the most consistent regions of abnormality in ASD, it is an under-reported territory in RsFc studies compared with the neocortex. Using connectome-wide MVPA, we report robust cerebellar functional connectivity disruptions in a cohort of HF-ASD subjects, thereby highlighting the cerebellum as a potential target for therapeutic, diagnostic, predictive, and prognostic developments in ASD. Over all, the findings we report advance our understanding of the neurobiology of HF-ASD and support the utility of whole-brain unbiased data-driven approaches in the development of biomarkers and therapeutic approaches in psychiatry.
Footnotes
Acknowledgments
This work is funded, in part, by the Athinoula A. Martinos Imaging Center, MIT, the Alan and Lorraine Bressler Clinical and Research Program for Autism Spectrum Disorder, and the MGH Pediatric Psychopharmacology Council Fund. The authors would like to thank the contributors of ABIDE, and Alfonso Nieto Castañón and Kaustubh Patil for helpful comments.
Author Disclosure Statement
No competing financial interests exist.