
Abstract
Rapid advances in neuroimaging and network science have produced powerful tools and measures to appreciate human brain organization at multiple spatial and temporal scales. It is now possible to obtain increasingly meaningful representations of whole-brain structural and functional brain networks and to formally assess macroscale principles of network topology. In addition to its utility in characterizing healthy brain organization, individual variability, and life span-related changes, there is high promise of network neuroscience for the conceptualization and, ultimately, management of brain disorders. In the current review, we argue for a science of the human brain that, while strongly embracing macroscale connectomics, also recommends awareness of brain properties derived from meso- and microscale resolutions. Such features include MRI markers of tissue microstructure, local functional properties, as well as information from nonimaging domains, including cellular, genetic, or chemical data. Integrating these measures with connectome models promises to better define the individual elements that constitute large-scale networks, and clarify the notion of connection strength among them. By enriching the description of large-scale networks, this approach may improve our understanding of fundamental principles of healthy brain organization. Notably, it may also better define the substrate of prevalent brain disorders, including stroke, autism, as well as drug-resistant epilepsies that are each characterized by intriguing interactions between local anomalies and network-level perturbations.
Introduction
Neuroscience is moving beyond a modular apprehension of brain organization, which emphasizes the relevance of individual regions, toward an approach that prioritizes the wiring and interactions between areas (Schroter et al., 2017; van den Heuvel et al., 2015). In the study of the healthy human brain, the last decade has witnessed major advances in characterizing large-scale structural and functional networks, and in relating network descriptions to interindividual variability, brain development and aging, skill learning and plasticity, as well as cognition and affect (Misic and Sporns, 2016). While the shift from region-centric to network-based analysis is welcome and truly exciting, the increased focus toward high-level network models should not undermine relevant information embedded in regional structural and functional properties as well as local circuits.
This review argues for a description of the human brain that, while embracing network-level paradigms, should also take advantage of the increasingly detailed information on individual nodes and edges that can be gathered with modern neuroimaging. Such an approach offers a rich description of the graph's individual elements, including microstructural properties, local structural markup, and the embedding of a region within large-scale networks. Addressing interactions between these domains may offer utility in understanding fundamental principles of brain organization and structure/function correspondence. Moreover, fusing these different levels of analyses may serve the study of brain disorders, in which anomalies may vary in location, size, and which have been shown to differentially affect whole-brain networks.
In what follows, we overview contributions of network neuroscience to the understanding of macroscale brain organization, developmental and aging-related processes, and the study of individual variations. We briefly outline parallel advances in high-resolution neuroimaging of local morphology, tissue microstructure, and function. Bridging the gap between network-level descriptions and these increasingly detailed markers of local properties promises a comprehensive characterization of the brain [see Fig. 1; for additional reviews, see Bullmore and Sporns (2009); Stam (2014)]. Furthermore, recent years have witnessed increased efforts to enrich neuroimaging analysis with nonimaging data, including information on gene expression, protein synthesis, and cytoarchitecture. In addition to discussing the promise of such multiscale paradigms to study healthy brains, we illustrate the potential of microstructure-informed connectomics in the study of diseases. We specifically focus on three broad classes of disorders, namely acquired lesions (such as strokes), neurodevelopmental disorders mainly characterized by network-level imbalances (such as autism), and finally, drug-resistant epilepsies that show an intriguing interplay between focal lesions and network-level anomalies.
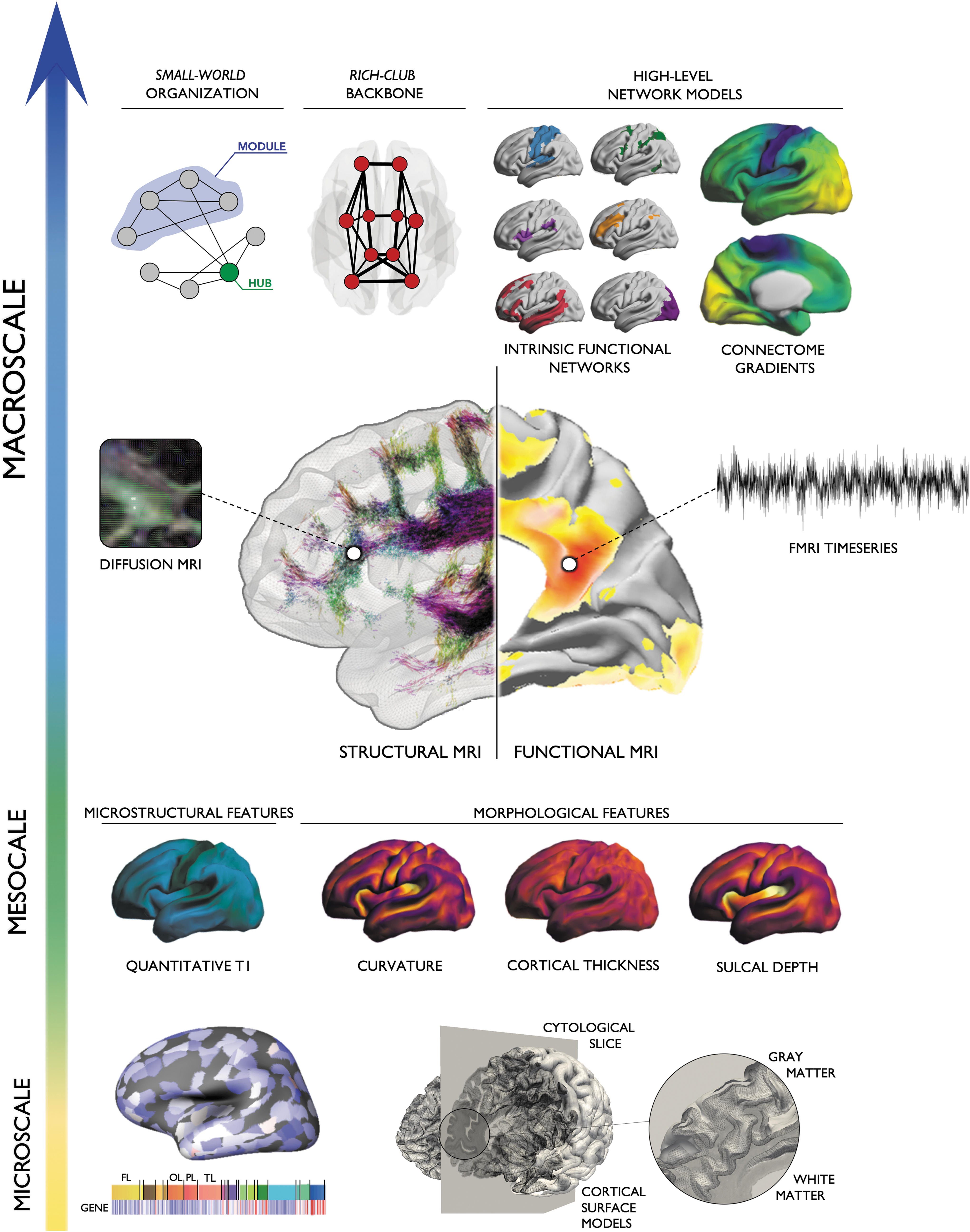
Enriched connectomics. Multiscale connectomics integrate various aspects of brain properties located along a continuum between microscale (gene expression, cytoarchitecture), mesoscale (MRI-derived markers of intracortical microstructure and morphology), and macroscale topology (small-worldness, core-periphery, modularity, connectome gradients). Leveraging parallel advances in high-resolution neuroimaging, detailed markers of local properties can be used to inform structural and functional network models, and vice versa. Fusing these different levels of analyses may shed light on fundamental principles of structure/function correspondence and unravel a more precise characterization of healthy and diseased brain organization.
Modeling Brain Networks
Over the past decade, the neuroimaging and neuroscience communities have witnessed a paradigm shift away from the focus on single regions and toward models that emphasize connectivity (Bassett and Bullmore, 2009; Bassett and Sporns, 2017; Bullmore and Sporns, 2009; Craddock et al., 2013). Increased availability of high-definition and multimodal neuroimaging, the maturation of data coregistration and integration processing pipelines, as well as advances in complex systems analytics have all contributed to an explosion of interest in the field. With respect to network-level approximations, diffusion MRI tractography has made it possible to approximate fiber tracts and derive structural connectomes in individual subjects (Jbabdi et al., 2015). In addition, analysis of statistical dependencies of functional MRI (fMRI) signals between different regions, especially in “resting” brains not engaged by any specific and controlled task, has been used to target multiple intrinsic functional networks using a single, preferably long acquisition (Biswal et al., 1995; Fox et al., 2005; Friston, 1994). Finally, structural MRI covariance analysis can tap into patterns of coordinated maturational trajectories in populations (Evans, 2013) and, more recently, in single individuals (Seidlitz et al., 2017).
The field of network science has offered formal analytics to parameterize whole-brain networks. Notable examples include decomposing the whole brain into a set of smaller communities (Girvan and Newman, 2002; Meunier et al., 2009; Newman and Girvan, 2004) and mapping network properties back to individual nodes, for example, in terms of overall connectivity degree or regional routes of communication efficiency (Sporns et al., 2007; van den Heuvel and Sporns, 2013). In parallel, graph-theoretical metrics can capture macroscale topology derived from fMRI (He et al., 2009; Honey et al., 2009; Salvador et al., 2005; Watts and Strogatz, 1998), diffusion MRI (Gong et al., 2009; Hagmann et al., 2008; Iturria-Medina et al., 2007), and MRI covariance analysis (Bassett et al., 2008; Chen et al., 2008; He et al., 2007), together with metabolic uptake data (Hu et al., 2015) as well as meta-analytical activations (Crossley et al., 2013). Collectively, these studies have shown that the human brain is neither organized purely randomly nor regularly, but rather according to principles that incorporate high clustering within segregated communities together with short paths between them. This small-world organization enables functional specialization and integration (Sporns et al., 2004), and closely relates to modularity, that is, network decomposability (Bullmore and Sporns, 2009). Modularity may offer adaptability and robustness to changing environmental conditions (Meunier et al., 2010), help to segregate externally oriented from internal processing (Vidaurre et al., 2017), and promote diverse functional dynamics (Gu et al., 2017; Kaiser and Hilgetag, 2010). Brain networks also seem to adhere to principles of hierarchy; connectomes have been subdivided into core and peripheral regions, a finding sometimes dubbed the rich-club principle, where a set of high-degree nodes are excessively interconnected (van den Heuvel and Sporns, 2011). Interestingly, the rich-club subnetworks seem to accumulate most long-range connections, indicating their role as a backbone for cross-module connectivity (van den Heuvel and Sporns, 2011) and functional diversity (Senden et al., 2014). By counting connections of a region (or alternatively the number of efficient paths passing through), network properties can be mapped back to individual nodes (Buckner et al., 2009; Hagmann et al., 2008; Zuo et al., 2012). Such “centrality mapping” can help to visualize connectome organization and accordingly identify hub regions that play key roles in network organization, communication, and dynamics. As an illustration of how different levels of network organization can be integrated, centrality mapping techniques can be enriched by modularity information, classifying hubs into those with provincial roles (mediating primarily connectivity within modules) versus connector hubs mediating between-module cross talk (Sporns et al., 2007). Conversely, it is also possible to simulate random failures as well as targeted attacks on specific nodes or edges to assess the relevance of specific network elements on global topology and community decomposition (He et al., 2008).
In addition to providing descriptors of human brain network organization, connectome properties may be useful markers of interindividual variability, life span- and experience-related changes, structure/function relationships, and ultimately behavior. Indeed, despite consistency of core network features across subjects (e.g., small-word organization, rich-club backbone, the layout of large-scale communities), there is also considerable variability across individuals (Smith et al., 2015; Tavor et al., 2016) and even within a given subject over time (Poldrack et al., 2015). Functional connectivity mapping has indeed demonstrated that some brain regions show more consistent connectivity profiles across individuals, while others have high variability (Mueller et al., 2013). Variable regions are often found in “multimodal association” cortices and, interestingly, correspond to those having undergone marked evolutionary expansion (Mueller et al., 2013). In the study of individual differences, recent work predicted subject identity using connectome “fingerprints” (Finn et al., 2015; Miranda-Dominguez et al., 2014; Xia and He, 2017), while others have used multivariate techniques to associate connectomes with cognitive/affective phenotypes (Smith et al., 2015; Tavor et al., 2016). These results illustrate significant promise in connectome-level studies to move beyond population-level inferences to inferences about single subjects, exploiting the uniqueness in network organization and the relationship to behavior. Accordingly, network neuroscience approaches have begun to address macroscopic plasticity as a result of experience and skill learning (Bassett and Mattar, 2017; Telesford et al., 2017). Finally, with ongoing efforts to make large-scale data sets available (Van Essen et al., 2012), often already in preprocessed and curated form, findings in young adults are increasingly complemented by targeted connectome analyses across the life span, covering developing (Satterthwaite et al., 2014) as well as aging cohorts (Zhou et al., 2012).
Enriching Connectomics with Local Information
In addition to the adoption of connectomics as one of the prevailing paradigms to study large-scale organization, there is also promise as well as need in refining the information from local and microstructural scales (Weiskopf et al., 2015). By guiding and enriching the definition of nodes and edges, the building blocks of connectomes, advances in microscale definition may improve biological validity of connectome models. Moreover, they lend novel opportunities in addressing interactions between microstructural and macroscale properties and ultimately increase the explanatory power of connectomics to address mechanistic as well as generative principles of brain organization.
At the scale of individual nodes, there is already considerable diversity in definition, a situation relating to the long-standing debate of what exactly constitutes a meaningful element or brain region. Postmortem anatomy has emphasized cytoarchitectural criteria, including lamination and cellular composition, for parcellations (Amunts et al., 2007). Alternative approaches build parcellations from in vivo measures directly. Features previously studied include anatomical landmarks and sulco-gyral patterns derived from cortical surface morphometry, and increasingly detailed markers of tissue microstructure obtained from quantitative MRI, including measurement of T1 relaxation times that may reflect intracortical myelin content (Waehnert et al., 2016). Structural markers are complemented with task-based functional activations as well as resting-state functional or structural connectivity (Eickhoff et al., 2015). Leveraging the Human Connectome Project data set, a recent study integrated spatial patterns in resting-state connectivity, task-based activations, image intensity, and cortical thickness measurements to parcellate the neocortex into 360 areas (Glasser et al., 2016). Although initial parcellations were generated at the group level, the authors notably devised an algorithm that parcellates an individual brain in native space, sidestepping potential problems relating to spatial normalization and between-subject averaging in a stereotaxic space (Braga and Buckner, 2017). Using spectral clustering of diffusion tractograms, another group identified 210 cortical/subcortical parcels (Fan et al., 2016), emphasizing that parcellations based on different input features may not necessarily converge toward equivalent solutions. Stability across algorithm choices and spatial scales will become increasingly relevant, as will be the definition of formal criteria to fuse methods. Recently, parcellations incorporated formal consensus procedures to maximize consistency across modalities, subjects, and data sets (Kelly et al., 2012). The continued move toward quantitative neuroimaging and increased access to nonimaging data in MRI-compatible reference frames provides new possibilities for parcellation enrichment and validation.
As for the nodes, the characterization of network edges has benefited from developments in MRI acquisition and modeling. Moving beyond early studies that were predominantly based on binary graphs, analytics is increasingly incorporating edge weighting when calculating network degrees, clustering, and efficiency (Rubinov and Sporns, 2010). Structural connectome studies based on diffusion MRI tractography have, for example, initially weighted edge strength through streamline counts, connectivity probabilities, and diffusion tensor metrics such as fractional anisotropy. In recent years, a microstructure-informed connectomic framework has become increasingly adopted, through which weights are derived from quantitative MRI markers of myelin, axon diameter (Assaf et al., 2013), and g-ratios (Mancini et al., 2017). In contrast to binary edge definitions or a weighting based on algorithm-dependent parameters, such as the number of streamlines, incorporating microstructural parameters into connectomics may provide more direct measures of conduction velocity (Mancini et al., 2017). Furthermore, the use of more sophisticated diffusion models resolving crossing fibers may reduce the number of false positive/negative connections, ultimately resulting in a better map of white matter connectivity (Jbabdi et al., 2015; Maier-Hein et al., 2017). Finally, enhancing the definition of structural connections will be of vital use in building predictive models of structure/function relationships (Daducci et al., 2016) and also in determining appropriate measures of functional connectivity itself (Buckner et al., 2013).
In addition to leveraging increasingly detailed data on individual nodes and edges based on advanced neuroimaging technique, there have been emerging efforts to enrich analyses with neurobiological information that is not neuroimaging derived. Examples include the Allen Institute transcriptional gene atlas (Hawrylycz et al., 2012; Sunkin et al., 2013), recently cross-referenced against morphological (Whitaker et al., 2016), functional connectivity (Krienen et al., 2016), and structural connectivity data (Romme et al., 2017). Other studies have related in vivo connectomics with atlas-based information on regional cytoarchitecture (Hilgetag et al., 2016) or chemoarchitecture (Turk et al., 2016). Interestingly, regions with similar microscale properties seem to preferentially interconnect, supporting homophily as an important generative principle of brain network formation (Betzel and Bassett, 2017; Betzel et al., 2016). By guiding and enriching the definition of nodes and edges, such multiscale advances may improve the biological validity of connectome representations and further increase the explanatory power of connectomics.
The Interplay of Local and Distributed Disturbances in Brain Disorders
In addition to its promise in typifying healthy brain organization, emerging work supports the potential of microstructure-enriched connectomics in the study of prevalent brain disorders. Three broad classes of disorders where local- and network-level anomalies interact in intricate ways are stroke, autism, and drug-resistant epilepsies. Stroke was chosen as an example of an acquired focal lesion that may lead to widespread network perturbations. Autism spectrum disorders (ASD) were selected as neurodevelopmental conditions primarily associated with connectome miswiring, but generally without consistently localizable substrate (i.e., lesional finding or brain abnormality typically detectable on an MRI). Finally, drug-resistant epilepsies often show a complex interplay between a focal lesion and widespread network anomalies, likely due to interacting developmental- and disease-related processes.
Stroke
Ischemic stroke is a leading cause of adult disability, with as many as 40% of stroke survivors classified as chronically disabled, being dependent on others for activities of daily living (Krueger et al., 2015). In addition to functional impairments, costs related to poststroke rehabilitation and treatment of morbidity and mortality further contribute to its economic burden worldwide (Mittmann et al., 2012; Rosamond et al., 2008).
Current models of stroke recovery based on functional and structural connectivity analyses continue to explain more and more of the variance in motor and cognitive recovery (Krueger et al., 2015; Stinear et al., 2012). Nonetheless, these have not been fully translated into clinical practice, that is, stroke survivors are not yet stratified into different subpopulations based on their residual network profile, and rehabilitation interventions continue to use a “one-size-fits-all” approach (Ward, 2017). A key impediment to the development of effective, individualized treatment may lie in the lack of a clear understanding of the dynamic interactions between local/regional circuit properties and macroscale functional reconfiguration that actively promote regeneration and recovery both at the lesion site and in remote areas after stroke. Previous studies have suggested that investigation of the underlying response that mediates the spread of pathology after an infarct is likely to yield a better understanding of the mechanisms that drive brain reorganization after an infarct (Carrera and Tononi, 2014; Rossini et al., 2003). This concept dates back to 1914 when Constantin von Monakow coined the term “diaschisis,” referring to a transient metabolic or functional dysfunction in areas that are remote to the primary site of injury (von Monakow, 1914). Although diaschisis is a well-established phenomenon after stroke, network analyses have become instrumental to further extend this concept. Resting-state fMRI experiments in patients with heterogeneous lesions, for instance, have shown that brain anomalies often extend beyond the primary lesion site but remain nevertheless constrained and predictable from connectivity patterns (Nomura et al., 2010). Likewise, computational models have shown that focal lesions have widespread dynamical consequences on system-wide integrative processes (Lariviere et al., 2018), which are influenced, to some extent, by the topological characteristics of the areas harboring the lesion (Honey and Sporns, 2008).
The connectivity between brain regions depends on axonal projections within white matter, a compartment highly susceptible to ischemic injury (Matute et al., 2013). Indeed, the white matter is less vascularized than gray matter (Fisher, 2011) and its anaerobic resistance declines with aging (Hamner et al., 2011). Furthermore, neuronal loss leads to Wallerian degeneration of axonal projections and contributes to disconnections of spared cortical areas, thus producing additional clinical deficits (Jones et al., 2013; Kuhn et al., 1989; Thomalla et al., 2005). The degree of disconnection is not always appreciable on MRI and brain damage is often underestimated, leading to less accurate models of poststroke impairments and potential for recovery.
Considering that a structural lesion resulting from a stroke may disturb the balance between local and large-scale connectivity, the concept of diaschisis can now be understood with respect to three categories: focal diaschisis (local changes in metabolism and neural activity), connectional diaschisis (changes between nodes of a same network), and connectome diaschisis (changes affecting the structural or functional connectome, including disconnections and reorganizations of subnetworks) (Carrera and Tononi, 2014). While the former often contributes predominantly to clinical deficits observed after subcortical lesions (e.g., thalamic strokes), network alterations seen in “connectional” and “connectomal” diaschisis appear to relate more consistently to clinical findings when lesions affect cortical areas.
Strikingly, investigations of multiscale, multimodal, or cross-network interactions in stroke research have so far been largely neglected. Influences of the microstructural state of the corticospinal tract on large-scale anomalies and motor performance have only been investigated sporadically (Borich et al., 2014; Carter et al., 2010; Schulz et al., 2015; Wu et al., 2015). One study, for instance, reported that interhemispheric functional connectivity significantly related to motor impairment only in patients with intact corticospinal tract, whereas those with greater damage did not show an association (Carter et al., 2012). While the interplay between structure and function at specific time points during stroke recovery has not yet been systematically studied, understanding cross-network interactions may be critical to move rehabilitation research forward. In fact, data based on microscale properties and macroscale organization may provide a rationale for patient stratification in addition to providing a neurobiological basis for rehabilitation interventions that target the optimal circuits for effective, individually tailored treatments. More specifically, network modeling could provide important information about the impact of focal lesions on overall network profiles. These models may extend standard connectomic frameworks, which generally focus on edge associations between two regions, by also incorporating indirect routes of information spread through a third region. In other words, information spread can be modeled as a cascading process through multiple paths and walks. A recent study leveraged these models based on structural connectome information in stroke, and demonstrated gains in predicting the degree of language impairments in individual patients [Fig. 2; Del Gaizo et al. (2017)]. Importantly, while many connections are lost in strokes, indirect pathways may still exist and may represent viable neurobiological targets to promote recovery.
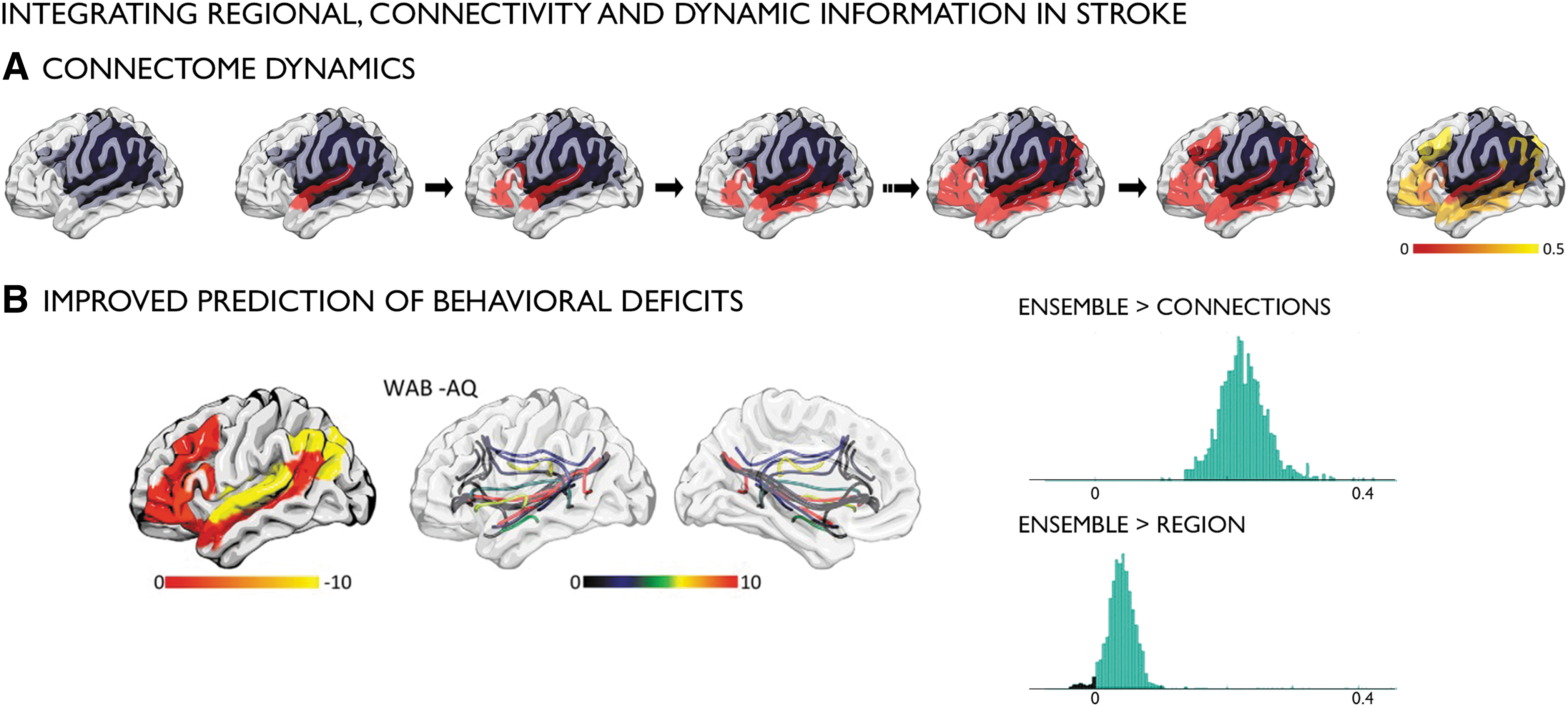
Dynamic network modeling in stroke (Del Gaizo et al., 2017).
Autism
ASD is among the most common developmental conditions, affecting currently around 1 in 70 children. With an onset in early childhood, autism spectrum conditions manifest with rather heterogeneous behavioral symptoms, but generally, deficits in communication and social cognition, often with repetitive behaviors/interests as well as sensory anomalies. While frequently assessed with neuroimaging, the literature has so far provided a rather complex pattern of alterations when comparing cohorts with autism with controls, dampening optimism that a single and consistent brain pattern can be identified.
At the level of brain structure, cortical thickness and voxel-based morphometric findings have indeed suggested both increases in gray matter, often in frontal and temporal regions (Ecker et al., 2013a; Valk et al., 2015; van Rooij et al., 2017), and also decreases (Wallace et al., 2010), null results (Haar et al., 2014), or mixed patterns (Ecker et al., 2012). Findings seem to vary with age of the cohorts studied, with increases in thickness being more consistently found in group-level analysis of children than in adults (Valk et al., 2015), suggesting an interaction between the disease and developmental trajectories (Raznahan et al., 2010; Smith et al., 2016). Earlier observations using MRI volumetry have indicated aberrant brain growth in early age and altered maturation in late childhood and adolescence (Courchesne et al., 2003). Likewise, histological reports have pointed to anomalies in prenatal stages, showing the occurrence of ectopic neurons in the white matter as well as cortical blurring, possibly reflecting incomplete neuronal migration (Avino and Hutsler, 2010). Recently, increased cortical blurring in autism has been confirmed in vivo using surface-based group-level analysis of MRI contrast (Andrews et al., 2017; Hong et al., 2017c). Furthermore, atypical cortical organization in autism was suggested using multidimensional cortical profiling, showing distorted surface area, curvature, and geodesic distance (Ecker et al., 2013a,b).
Challenges in the exact localization of structural anomalies across the spectrum could relate to a malformative process that variably affects network nodes and their communication patterns, ultimately contributing to the broader autism phenotype. In this scenario, anomalies in the formation and organization of large-scale networks may differentially contribute to impairments in higher order cognition or sensory integration. Accordingly, this may also be associated with some of the inconsistencies reported in the autism-related resting-state functional imaging literature, among which the majority has focused on mapping connectivity anomalies. Indeed, findings appear somewhat heterogeneous in location and even direction, with some studies reporting scattered functional connectivity increases in addition to prevailing and most frequently reported connectivity reductions (Deen and Pelphrey, 2012; Heinsfeld et al., 2018; Keown et al., 2013; Uddin, 2015; Yahata et al., 2016). Inconsistent differences at the group level may propagate down to the individual subject level, and classifiers designed to dissociate ASD from controls have provided rather variable accuracies, often below behavioral criteria (Heinsfeld et al., 2018; Plitt et al., 2015). Collectively, these results suggest that the ASD phenotype might arise from broader imbalances in functional architecture across individuals, which may not necessarily be localizable to specific regions but rather may reflect overarching principles of network-level organization, a finding also supported by higher variability in autism functional connectivity patterns than in controls (Nunes et al., 2018).
In addition to consolidating previous findings, for example, by providing more biologically plausible metrics of connectivity, enriched connectomics may be useful to assess the link between local anomalies and large-scale network perturbations in ASD (Fig. 3). Of interest will be to clarify the association between changes in local cortical organization (e.g., cortical thickness increases, cortical interface blurring, and laminar alterations) and structural and functional connectivity. Diffusion MRI group-level analyses have shown alterations in the microstructure and architecture of deep as well as superficial white matter tracts related to default-mode, auditory, and visual cortices (Barnea-Goraly et al., 2004; Fletcher et al., 2010; Sundaram et al., 2008). Analyzing diffusion MRI as well as cortical morphology, a recent study observed abnormalities in cortical folding together with white matter microstructure, suggesting coupled effects in both compartments (Ecker et al., 2016).
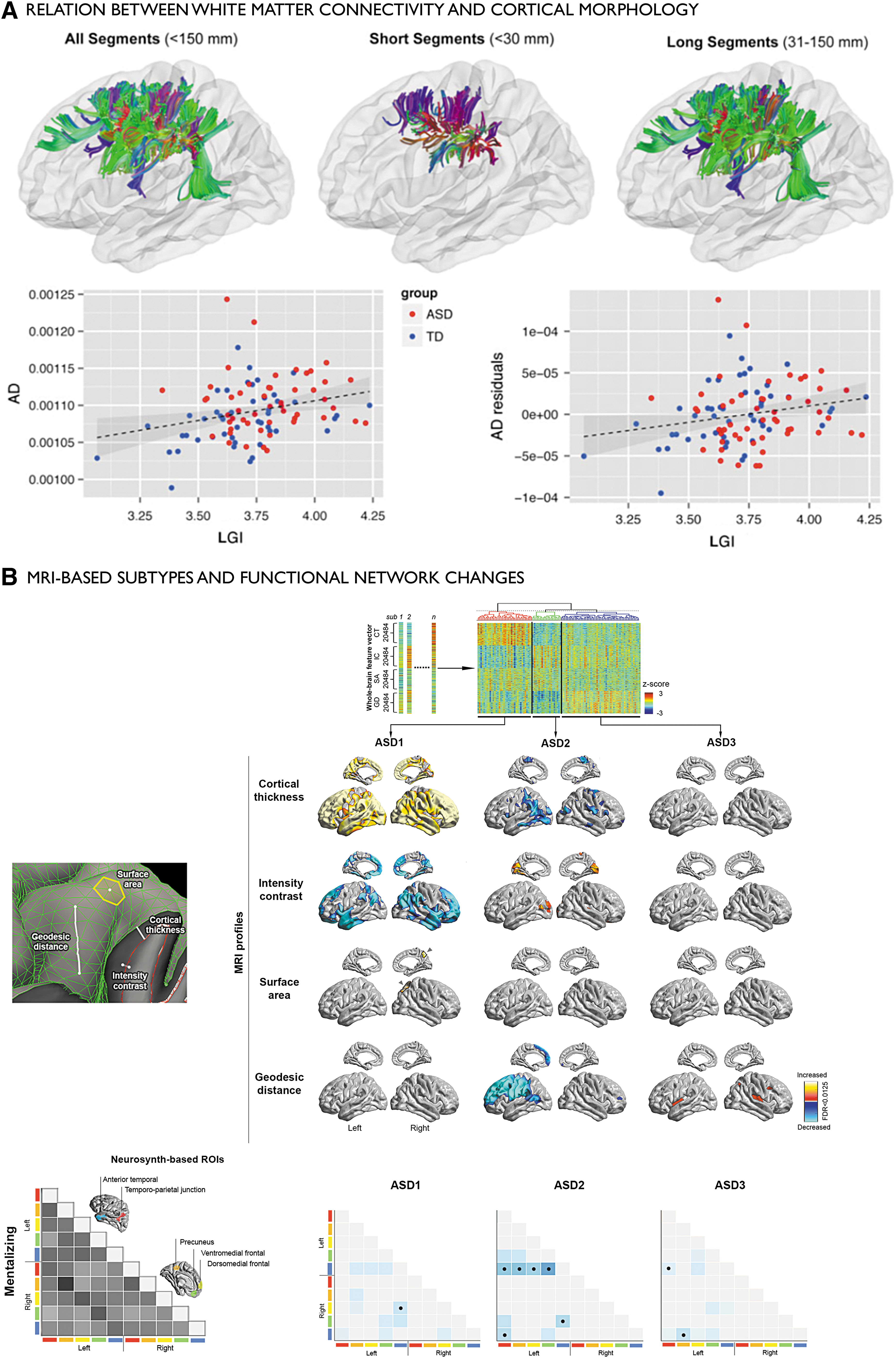
Findings in ASD showing interactions between local- and network-level anomalies.
Epilepsy
Around 30% of epileptic patients suffer from seizures that cannot be controlled by antiepileptic mediation and are therefore generally considered surgical candidates, given the established benefits of surgery for seizure control and quality life in epileptic patients (Engel et al., 2012; Wiebe et al., 2001). Neuroimaging of drug-resistant epilepsies has traditionally focused on the detection of focal lesions considering its relevance in surgical planning. Indeed, the MRI detection and subsequent resection of a lesion are currently the best predictors of postsurgical seizure freedom (Bernasconi et al., 2011). In temporal lobe epilepsy, the most prevalent syndrome, patients commonly present with mesiotemporal sclerosis, a lesion characterized by variable cell loss and gliosis in the hippocampal formation and adjacent structures (Blumcke et al., 2013; Thom et al., 2009). Frontal lobe epilepsy, another prevalent surgically remediable syndrome, often relates to cortical dysplasia, a malformation characterized by laminar and cytological anomalies. Surface-based structural MRI features of morphology as well as tissue contrast and intensity have provided an increasingly detailed in vivo characterization of these lesions and have been useful in guiding algorithms for automated lesion detection (Hong et al., 2014). Furthermore, these markers can capture interpatient variability and have been statistically related to histological subtypes (Bernhardt et al., 2015b, 2016; Hong et al., 2017b).
In addition to its utility for targeted local studies, MRI has also been instrumental in demonstrating distributed pathological networks in both syndromes [Fig. 4; Bernhardt et al. (2015a); Gleichgerrcht et al. (2015); Richardson (2012)]. In temporal lobe epilepsy, converging evidence from voxel-wise and surface-based morphometry (Bernhardt et al., 2010; Bonilha et al., 2004; Keller and Roberts, 2008; McDonald et al., 2008), quantitative longitudinal relaxation time mapping (Bernhardt et al., 2017), and diffusion imaging (Concha et al., 2012; Keller et al., 2017; Liu et al., 2016) has revealed anomalies beyond mesiotemporal regions and temporo-limbic tracts, often extending into a broad cortical/subcortical territory. In dysplasia-related frontal lobe epilepsy, cortical thickness and curvature analyses also emphasized widespread anomalies (Hong et al., 2016), complemented by diffusion MRI studies suggesting architectural and microstructural alterations of intrahemispheric fibers as well as the corpus callosum (Campos et al., 2015; Fonseca Vde et al., 2012). In both syndromes, structural data are increasingly complemented by resting-state fMRI studies suggesting an imbalance in connectivity between several intrinsic functional systems, predominantly temporo-limbic and default-mode networks (Bettus et al., 2010; Doucet et al., 2013; Maccotta et al., 2013; Morgan et al., 2011). In addition, graph theoretical network analyses consistently described large-scale alterations in global topology across several modalities, emphasizing system-level compromise in syndromes traditionally defined as focal (Bernhardt et al., 2011; Bonilha et al., 2012; Liu et al., 2014; Wang et al., 2014; Yasuda et al., 2015).
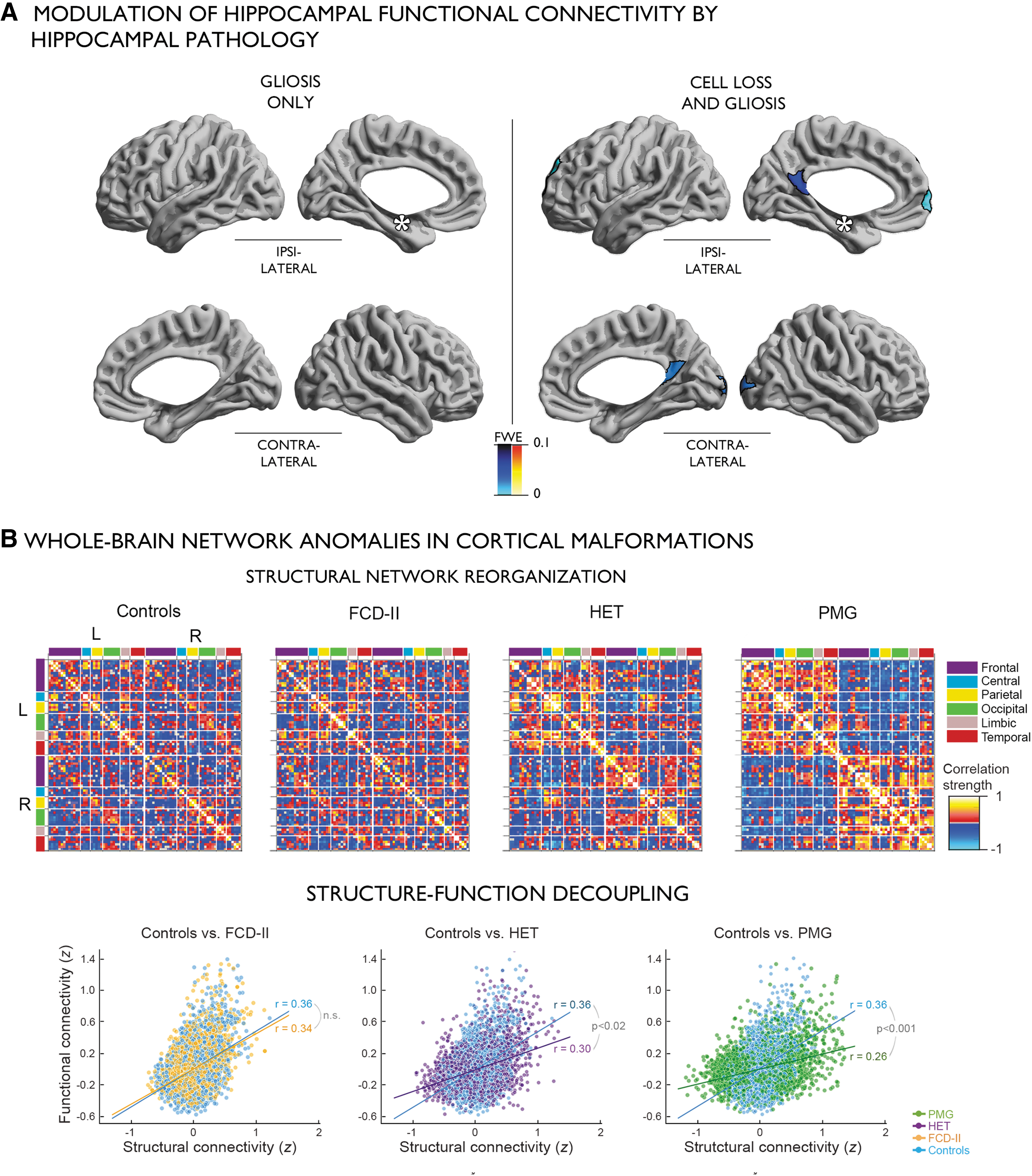
Findings in drug-resistant epilepsy suggesting an interplay between regional anomalies and large-scale network phenotypes.
To study interactions between whole-brain phenotypes and microscopic variations of specific regions, several studies addressed connectome anomalies in patients with postoperative histopathological lesion grading. In temporal lobe epilepsy, patients with marked hippocampal sclerosis have been shown to present with perturbed resting-state functional connectivity to default-mode hubs compared with those with isolated gliosis (Bernhardt et al., 2016). Using subfield-level volumetric and T2 intensity analysis of the hippocampal formation as a bridge, the study furthermore correlated degrees of functional connectivity decreases to structural alterations, supporting a parametric modulation of network anomalies by degrees of structural damage (Bernhardt et al., 2016). A separate analysis of the same group reported a similar modulation at the level of white matter microstructure, with patients presenting with severe hippocampal volume loss also showing more marked anomalies than patients with normal hippocampal volumes (Liu et al., 2016). By showing that microscopically different patterns of hippocampal pathology may result in rather variable perturbations of macroscopic functional and structural networks, these findings lend important evidence for multiscale interactions in pathological networks.
In extratemporal lobe epilepsy related to cortical malformations, a recent analysis showed a graded pattern of structural and functional network rearrangements across different lesion types, with severe alterations in patients with malformations believed to occur late during corticogenesis, while patients with malformation due to disruptions of early proliferative stages showed only modest disruptions (Hong et al., 2017a). Overall, these findings suggest that time of insult during corticogenesis impacts the severity of topological network reconfiguration.
As a surgically amenable disorder, epilepsy provides several opportunities to validate and inform neuroimaging and connectome approaches, for example, to develop and validate models for surgical outcome prediction. In temporal lobe epilepsy, several recent studies have combined machine learning techniques of different MRI and network features, and supported prognostic yield of limbic tract-wise diffusion tensor imaging assessment (Keller et al., 2017), extrahippocampal gray matter surface shape analysis (Bernhardt et al., 2015b), as well as large-scale diffusion connectomes in predicting outcome in individual patients (Bonilha et al., 2013, 2015). While independence of some of these features from hippocampal pathology (also a recognized predictor of outcome) still needs to be established, the literature is overall on a promising path toward predictive platforms, integrating local- and network-level information (Bonilha et al., 2013, 2015).
Conclusions
Neuroimagers and network neuroscientists live in exciting times, where a rich catalog of powerful measures of tissue microstructure and connectivity has become accessible and integrative analysis practical. Multivariate techniques, such as canonical correlation and partial least squares, provide promising formalisms to relate one multidimensional space to another. Accordingly, these models can capture complex relationships, including those between structural and functional domains, between macro- and microstructural properties, as well as those between brain and behavioral measures, with the latter characterizing observable phenotypic variability (Misic and Sporns, 2016). An emerging class of formalisms to integrate information in the structural and functional domain and between different scales is connectome-informed computational models, such as those predicting functional dynamics and interactions from structural connectome data (Bettinardi et al., 2017; Deco et al., 2013, 2017; Sanz Leon et al., 2013). In more clinically driven neuroimaging research settings, these in silico models have been used to assess network-level consequences of focal brain lesions, promising potential utility as prognostic tools (Alstott et al., 2009; Hutchings et al., 2015). Finally, the increasing dimensionality and complexity of multimodal and multiscale approaches justify the use of supervised as well as unsupervised statistical learning techniques that can be used to identify salient features, assess feature interdependencies, as well as to project the data into informative lower dimensional subspaces that facilitate integrative investigations (Bzdok et al., 2017).
While we have advocated for reconciling micro-, meso-, and macroscale information into a unified framework, challenges remain in fusing different levels of analysis. Indeed, despite continued efforts to integrate multiscale data into a common reference frame, technical and conceptual limitations remain to be addressed to improve interpretability. On the one hand, conducting multiscale analyses at high spatial resolutions (e.g., at the level of individual vertices or voxels) poses computational challenges. On the other hand, although aggregation of individual nodes into parcels or regions of interest has become a viable alternative (Fan et al., 2016; Glasser et al., 2016; Schaefer et al., 2017; Tzourio-Mazoyer et al., 2002), only few specific guidelines and formal procedures exist as to which parcellation scheme(s) should be chosen. Another challenge imminent to high-dimensional data and multivariate analysis techniques more generally lies in the increased risk of overfitting and false positive results, mandating the use of conservative cross-validation and reproducibility analyses aiming to control for false positive findings and to ensure replicability.
Despite these challenges, we are nevertheless confident that the near future brings promising new developments in enriched connectomics, a multidisciplinary and multiscale enterprise that will not only advance our understanding of fundamental aspects of healthy brain organization but also of the neurobiological underpinnings of prevalent and often detrimental diseases.
Footnotes
Acknowledgments
S.L. acknowledges funding from Fonds de la Recherche du Québec–Santé (FRQS). S.-J.H. was funded by the Canadian League Against Epilepsy and the Canadian Institutes of Health Research (CIHR). N.B. was funded by CIHR and received salary support from FRQS. A.B. acknowledges funding from CIHR, National Science and Engineering Research Council of Canada (NSERC), and the Quebec Bioimaging Network (QBIN). L.B. receives research support from the National Institutes of Health/National Institute on Deafness and Other Communication Disorders (DC014021, DC011739, and DC014664) and from the American Heart Association (SFDRN26030003). B.C.B. acknowledges research support from NSERC (Discovery-1304413), CIHR (FDN-154298), Azrieli Center for Autism Research (ACAR), SickKids Foundation (NI17-039), an MNI-Cambridge collaboration grant, and received salary support from FRQS (Chercheur Boursier Junior 1).
Author Disclosure Statement
No competing financial interests exist.