
Abstract
Children with reading difficulties (RDs) often receive related accommodations in schools, such as additional time for examinations and reading aloud written material. Existing data suggest that these readers share challenges in executive functions (EFs). Our study was designed to determine whether children with RDs have specific challenges in EFs and define neurobiological signatures for such difficulties using magnetic resonance imaging (MRI) data. Reading and EFs abilities were assessed in 8–12-year-old children with RDs and age-matched typical readers. Functional MRI data were acquired during a Stroop task, and functional connectivity of the EFs defined network was calculated in both groups and related to reading ability. Children with RDs showed lower reading and EFs abilities and demonstrated greater functional connectivity between the EFs network and visual, language, and cognitive control regions during the Stroop task, compared to typical readers. Our results suggest that children with RDs utilize neural circuits supporting EFs more so than do typical readers to perform a cognitive task. These results also provide a neurobiological explanation for the challenges in EFs shared by children with RDs and explain challenges this group shares outside of the reading domain.
Introduction
Intact reading is fundamental for academic success. For this reason, providing appropriate accommodations and interventions for children with reading difficulties (RDs) is crucial. RDs are defined as ongoing challenges in reading accurately and fluently despite exposure to the written language and provision of appropriate intervention (IDA, 2011).
Many theories have been raised regarding the primary cause for RDs, including phonological deficit (Snowling et al., 1997), asynchrony (Breznitz, 2006), orthographic (Brunswick et al., 1999), and morphological (Nagy et al., 2006) theories. Although the main deficit in individuals with RDs is within the reading domain, studies conducted during the past decade have identified deficits in executive functions (EFs) in this group of readers, both behaviorally (Altemeier et al., 2008; Brosnan et al., 2002; Gooch et al., 2011) and with evidence of alterations in neural circuits involved in executive dysfunctions (Horowitz-Kraus, 2014, 2015; Horowitz-Kraus et al., 2015c, 2016).
Relationship between reading ability and EFs
The ability to read fluently involves intact subcomponents of reading, such as phonology, orthography, and semantics, but it also requires higher order cognitive ability, usually referred to as the umbrella term “executive functions” (EFs) (Breznitz, 2006). EFs are cognitive processes used to optimize performance through planning, organizing, and learning that include working memory, speed of processing, switching, and visual attention (Anderson, 2002). In the past decade, accumulated literature suggests that children with RDs demonstrate challenges in EFs in addition to their reading problems (Horowitz-Kraus, 2014, 2015; Horowitz-Kraus and Breznitz, 2008, Horowitz-Kraus and Holland, 2015, Horowitz-Kraus et al., 2015a, 2015c). However, there is still a gap in knowledge as to how general this deficit in EF is or if it is restricted to tasks involving reading, which obviously has clinical and educational implications.
Defining whether there is a specific underlying neurobiological deficit in EF has the potential to affect diagnosis of RD early in life (even before reading is acquired) and treatment in early childhood. Therefore, the aim of this study was to determine neurobiological markers for a specific EF deficit in children with RDs.
The neurobiological basis of the Stroop effect and EFs
An examination of EFs in children with RDs is challenging since finding one task that engages all EFs is itself challenging. In the past decade, the Stroop task was suggested as a paradigm that engages multiple EF domains, such as inhibition and switching (Stroop, 1935). For this task, participants are first presented with words representing colors (red, blue, green, etc.) that are each colored in the same ink as the presented word and then are asked to name the words. Next, words representing colors are presented in different-colored fonts, and participants are asked to identify the font color and ignore the word, which is referred to as the interference effect (Stroop, 1935). By inhibiting their automatic response and then determining the color of the font, subjects rely on their working memory and speed of processing (Stuss and Knight, 2002). This automatic timely response inhibition was coined “the Stroop phenomenon” and has been observed in both the colors and numerical Stroop tasks (Liua et al., 2014).
Neuroimaging studies have shown increased brain activation in several brain regions related to EF, such as the left lateral prefrontal cortex, left anterior cingulate, and left parietal and parieto-occipital cortices, in typical readers performing this task (Adleman et al., 2002). Previous studies identified different populations with an EF deficit who demonstrated an altered performance in this task [e.g., children with attention-deficit/hyperactive disorder (Homack and Riccio, 2004) and individuals with post-traumatic syndrome disorder (Moradi et al., 2000)]. Children with RDs also demonstrated decreased performance during this task compared with typical readers (Faccioli et al., 2008; Horowitz-Kraus, 2014, 2015; Protopapasa et al., 2007), even in the nonlinguistic version of the task (Liua et al., 2014). However, the involvement of neural circuits related to RDs during the Stroop task remains elusive, but are crucial for an objective early detection of such deficit.
The neurobiological basis of RDs and EFs
Several neuroimaging studies focusing on individuals with RDs suggest a controversy of inconsistent patterns of activation and connectivity of frontal regions and cognitive control networks, respectively, during verbal and nonverbal tasks. Some studies have reported an overactivation in anterior regions, usually related to EFs, during a written nonword rhyming task, such as in the inferior frontal gyrus, part of the cingulo-opercular cognitive control network (Shaywitz et al., 1998). Similar results were seen in a reading-aloud task (Brunswick et al., 1999). Others have reported increased functional connectivity between seeds in the occipital and frontal (inferior frontal gyrus) lobes in children with dyslexia during naming and reading tasks (Morken et al., 2017), as well as during functional magnetic resonance imaging (fMRI) sentence-reading comprehension, which poses a heavier load on cognitive processes (Yagle et al., 2017). And yet other studies suggest a decreased functional connectivity in cognitive networks involving these frontal regions during both reading and rest [cingulo-opercular (Horowitz-Kraus et al., 2015c) and frontoparietal networks (Horowitz-Kraus et al., 2015a)]. These networks are related to error monitoring and response evaluation (cingulo-opercular), as well as speeded response and working memory (frontoparietal) (Dosenbach et al., 2008). Other networks related to the involvement of cognitive control abilities during reading are the dorsal attention (Vogel et al., 2012) and salience (Li et al., 2012) networks, as well as the default-mode network (DMN) (Koyama et al., 2010), corresponding separately and in combination with cognitive flexibility, goal setting, information processing, and attentional control (Fig. 1) (Anderson, 2002). Owing to the existing controversy and the limited number of published studies examining specifically the functional connectivity patterns during an EF task rather than during a reading paradigm, and since only subnetworks related to EF have been investigated while those associated with EF required for the Stroop task were not, it remains unclear whether there is a neurobiological difference in the functional connections within these networks in children with RDs and the relationship with reading ability. This was the focus of this study.
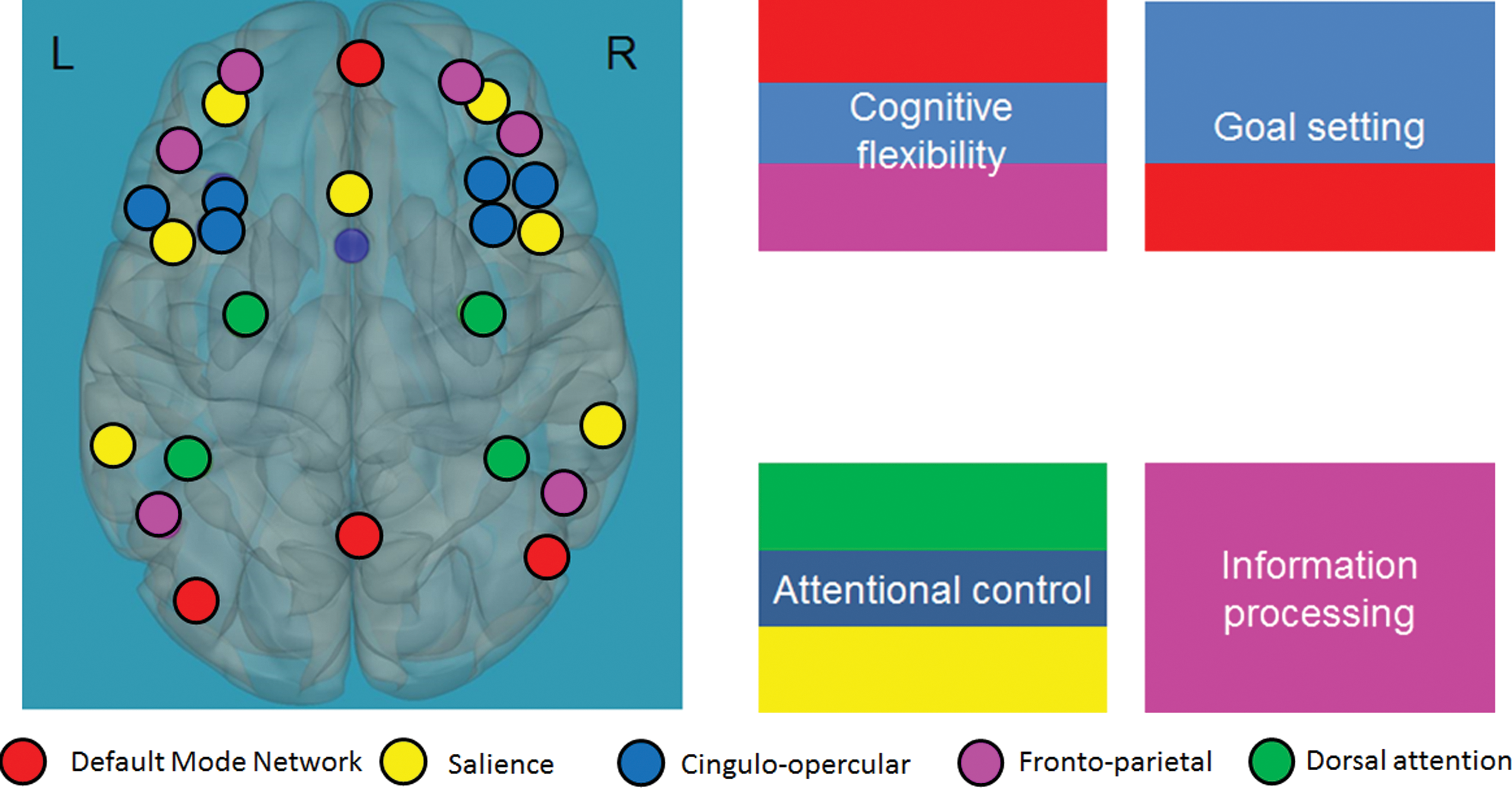
The EFs network. The EFs network is composed of the default-mode network (in red); the cingulo-opercular (in blue), frontoparietal (in pink), salience (in yellow), and dorsal attention (in green) networks; and the corresponding abilities (cognitive flexibility, goal setting, attentional control, and information processing). Adapted from the proposed developmental model for EFs (Anderson and Reidy, 2012). EFs, executive functions. Color images available online at
The study was designed to identify the neural circuits underlying the differences in EFs in children with RDs compared with typical readers during a Stroop task. To determine the differences in functional connections within the EF network and between the EF network and other regions related to the reading (i.e., cognitive, language, and visual processing regions) in children with RDs compared with typical readers, we defined the “EF network” as being composed of functional networks related to a variety of EFs and that were previously related to reading (i.e., frontoparietal, cingulo-opercular, dorsal attention, and salience networks and the DMN), and view these as a set of a priori functional networks involved in higher level cognitive abilities. Grouping all of the networks into one EF network enabled us to focus only on the role of the EF network as a whole and its engagement with other regions in the brain during an EF task, without referring to each network separately. Unlike previous work looking at the correspondence between cognitive control networks (Horowitz-Kraus et al., 2015c), the main aim of this study was to generate one network that includes all subnetworks of the executive system within the whole brain to understand the overall reliance on this network in children with RDs versus typical readers during an EF task. Since we were interested in within versus outside network connectivity for a set of a priori networks of interest, we explored outside network connectivity and also global efficiency exploring within-network differences. Global efficiency is a graph theory-driven measure that provides information on the average inverse shortest path length and can be computed on disconnected networks, assuming that the paths between disconnected nodes will have infinite length and, therefore, zero efficiency (Rubinov and Sporns, 2010).
We hypothesized that challenges in EFs in children with RDs would be accompanied by greater functional connections between the EF network and regions related to reading, as was previously suggested in other populations (Deslauriers et al., 2017). This hypothesis is based on the assumption that greater effort in performing the task will be accompanied by increased engagement and reliance on the EF network and increased synchronization within the reading and language regions. Owing to the inverse relationship between graph theory measures for network connectivity (Wong et al., 2007), we anticipated that greater global efficiency scores would be related to increased reading and EF scores. EFs develop before reading is acquired and support reading later in life. It is therefore important to define their involvement in reading in children with RDs. This will support detection of these challenges earlier in life to provide intervention before the evidence of reading failures.
Materials and Methods
Participants
Children between the ages of 8 and 12 years participated in this study: children with RDs (n = 28, mean age 9.86 years, SD = 1.46, 13 males, 28 right handed) and typical readers (n = 17, mean age 9.77 years, SD = 1.4, 9 males, 17 right handed) who were native English speakers participated in this study. Children attended the second to sixth grades without any significant difference in grade level between the two groups (children with RDs: mean grade level = 3.79, SD = 0.833; typical readers: mean grade level = 3.82, SD =1.058, t(45) = −0.123, p = 0.903). Written informed consents and assents were received from the parents and the participants, respectively. The appropriate Institutional Review Board approved the study. Participants had no history of psychiatric or neurological impairment, including attention difficulties.
Behavioral and neurocognitive measures
The nonverbal and verbal IQ of the participants was determined through administration of the Test of Nonverbal Intelligence (TONI-3) (Brown et al., 1997) and a vocabulary task (Peabody Picture Vocabulary Test [PPVT-4]) (Dunn and Dunn, 2007), respectively, to verify normal (>85) nonverbal and verbal IQ. Current RD literature suggests that children with RDs are defined as such if they demonstrate a score of −1 standard score and lower in at least two reading measures (after; Kovelman et al., 2012). These reading measures include several subdomains of reading, such as automatic and nonautomatic orthographical, decoding, reading comprehension, and fluency or phonemic awareness abilities. Therefore, in this study, children performed the following behavioral reading measures: (1) word and (2) nonword reading, as well as (3) reading fluency from the Test of Word Reading Efficiency second edition (TOWRE II) (Torgesen et al., 1999), (4) reading comprehension, and use of the (5) word-attack subtest, and (6) letter word subtest from the WJ-III (Woodcock and Johnson, 1989).
Neurocognitive testing
To evaluate differences between children with RDs and typical readers in EF abilities, the following EF tests were administered: (1) working memory (digit span; Wechsler, 1999), (2) speed of processing (symbol search/coding subtests; Wechsler, 1999), (3) naming ability (naming and letter naming from the Comprehensive Test of Phonological Processing [CTOPP]; Wagner et al., 1999), (4) switching and inhibition (Stroop, from Delis–Kaplan Executive Function System [D-KEFS]; Dellis et al., 2001), and (5) visual attention (SkySearch; Manly et al., 1999). Typical readers showed intact scores in all reading tests.
To determine the differences in reading and EF measures between the two groups of readers, independent t-test analysis was performed. To define the relationship between reading and EFs, a Pearson correlation was performed for both reading groups. Data were corrected for multiple comparisons using a Bonferroni correction.
Neuroimaging measures
Stroop task
In this task, participants were shown a series of color words in the center of the screen. Each word had a colored font, and participants were asked to identify the color of the font. Participants were given a 2-min practice session before beginning the scan. During the scan, they were asked to focus on the task and avoid sleeping and/or closing their eyes. The stimulation included three conditions: interference, control, and rest.
In the interference condition, the participants were presented with words colored in a different color than the actual word (i.e., the word “blue” was printed in a red ink). The participants were instructed to “push the button on the response box that matches the color of the ink and not the word.” The buttons on the response box corresponded by color and order to three colored circles on the screen (red, blue, and green) so that the participants would not be tempted to gaze toward their hands. Each stimulus was presented for 2 s, and the response screen (the three circles) was also presented on the screen for 2 s.
In the control condition, the participants were presented with a string of four “X” characters (“XXXX”) printed in different colors and asked to “push the button that matches the color of the string of X's”.
In the rest condition, the participants were presented with a cross on the screen and instructed to look at the cross.
There were five blocks of words for the interference condition (15 stimuli per block), five blocks of X's for the control condition (15 stimuli per block), and two blocks of rest (i.e., a cross), which alternated randomly. The duration for each block was 60 s. The task lasted for a total time of 12 min and 22 s.
MRI acquisition and data preparation
Participants were given time to acclimate to the MRI scanner and practice lying still. Head motion was controlled using elastic straps attached to the head-coil apparatus; along the child's forehead, a headband further reduced motion. Once comfortable inside the machine, the child began watching a movie through the MRI-compatible audiovisual system. At this point, image acquisition commenced. Headphones equipped with a built-in microphone were used to communicate with the child, to provide positive feedback throughout the scan. If the child did not wish to continue at any point, the process was stopped. All children were awake throughout the scan. All participants were scanned using a 3T Philips Achieva MRI scanner accompanied by the Avotec SS3150/SS7100 audio/visual system. A gradient echo planar sequence was used for T2-weighted blood oxygen level-dependent (BOLD) fMRI scans with the following parameters: TR/TE = 2000/38 ms, BW = 125 kHz, FOV = 25.6 × 25.6 cm, matrix 64 × 64, and slice thickness of 5 mm. Three hundred sixty image volumes were acquired during the fMRI experiment consisting of 60 s per condition for a total time of 300 s (150 volumes) for each stimulus (i.e., “word/color” interference and “XXXX” control). A T1-weighted inversion recovery gradient echo anatomical whole brain was acquired for each participant for an anatomical coregistration and used in spatial normalization of the functional data.
MRI data analysis
Data preprocessing and first-level analyses were performed using SPM12 implemented in the CONN toolbox (Whitfield-Gabrieli and Nieto-Castanon, 2012). Images were slice-time corrected and realigned. The data were normalized using a 3D anatomical whole-brain scan to match the Montreal Neurological Institute standard template, resampled (3-mm3 voxels), and smoothed with 8 mm full width at half maximum (FWHM). Voxels activated by the Stroop task were identified using a general linear model. Six motion parameters were determined through 3D affine transformation. Excessive motion led to removal from the postprocessing pipeline. All data met the criterion of median voxel displacement in the center of the brain (0.2 mm). Voxel-wise temporal denoising of the BOLD signal was applied through regression of zero- and first-order derivatives of the six motion parameters, regression of the five principle components of the white matter, and cerebrospinal fluid BOLD signal using a component-based noise correction approach (Behzadi et al., 2007). Images regarded as movement outliers were regressed out. Outliers detection was performed using the ART toolbox (
Functional connectivity analysis
Based on the relationship between the cingulo-opercular, frontoparietal, salience, dorsal attention networks, the DMN, and reading ability, these networks were included in the EF network (i.e., we chose only the edges of these networks to create the utilized “EF network”). This means that each network received the same “weight” within the large EF network (for additional information, see Rubinova and Spornsd, 2010). The networks, except for the cingulo-opercular, were defined by the networks atlas implemented in CONN (
Global efficiency calculation was performed for the entire EF network. Global efficiency was calculated in CONN using the formula (from Latora and Marchiori, 2001):
where Ei is the efficiency of node i, n is the number of network nodes, N is the set of all network nodes, and dij −1 is the inverse shortest path length between nodes i and j.
The shortest path length was calculated using a binarized connectivity matrix, as defined in CONN, where a value of 1 means a direct connection between two nodes, while a value of 0 would mean that a direct connection between the two nodes is absent. Adjacency matrix threshold of 0.15, two-sided was set in CONN. To measure the difference in global efficiency between the groups, two-sample t-test was performed and significance level was set at p < 0.05, false discovery rate (FDR) corrected for multiple comparisons.
Correlation of global efficiency with behavioral scores
To determine the associations between global efficiency of the EF network and reading and EF measures, a Pearson correlation between these measures was performed for both reading groups. Data were corrected for multiple comparisons using a Bonferroni correction.
Results
Behavioral results
Children with RDs demonstrated significantly lower reading ability, including orthographic ability (word reading), decoding (nonword reading), phonemic awareness, and comprehension, than typical readers. Children with RDs also demonstrated lower scores in all EF domains (switching, inhibition, working memory, speed of processing, attention, and naming). See Table 1 for these results.
Baseline Behavioral Reading and Executive Function Scores for Children with Reading Difficulties and Typical Readers
p < 0.05; ** p < 0.01; *** p < 0.001.
CTOPP, Comprehensive Test of Phonological Processing; D-KEFS, Delis–Kaplan Executive Function System; PPVT, Peabody Picture Vocabulary Test; RDs, reading difficulties; SD, standard deviation; TEA-Ch, Test of Everyday Attention for Children; TONI, Test of Nonverbal Intelligence; TOWRE, Test of Word Reading Efficiency; WISC, Wechsler Intelligence Scale for Children; WJ, Woodcock–Johnson Test.
Correlation between reading and EF measures
A Pearson correlation across both reading groups revealed significant positive correlations between measures related to orthographic ability (timed: TOWRE, nontimed: letter word) and EF measures for (1) memory (digit span) [r(45) = 0.343, p < 0.05; r(45) = 0.299, p < 0.05, respectively], (2) speed of processing (coding) [r(45) = 0.351, p < 0.05; r(45) = 0.341, p < 0.05, respectively], and (3) visual attention (Sky-search) [r(45) = 0.401, p < 0.01; r(45) = 0.39, p < 0.05, respectively]. Results suggest that increased reading ability is related to greater working-memory, speed-of-processing, and visual-attention scores.
Imaging data
Functional connectivity between the EF network and regions related to reading for typical readers: results suggest positive functional connectivity between the EF network and regions related to cognitive control (e.g., BAs 8, 9, 10, 24, 32), language (e.g., BAs 7, 21, 22, 41, 42), and visual processing (e.g., BAs 17, 18, 19); p < 0.05, FDR corrected. For the BA-related anatomical regions, see Supplementary Table S1 (Supplementary Data are available online at
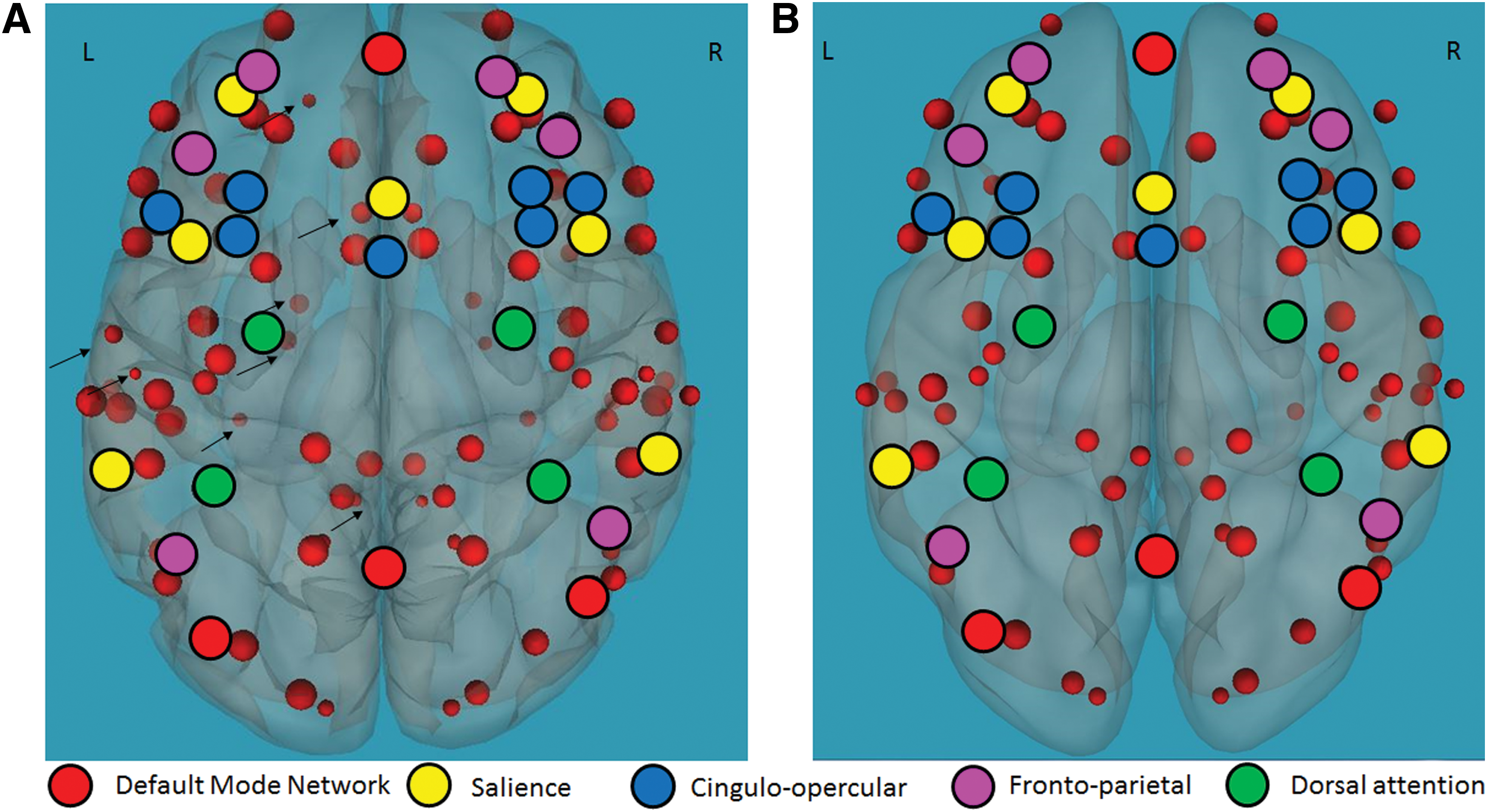
Functional connectivity of the EFs network and reading-related regions in typical readers and children with RDs. Positive correlation between functional connectivity of the EFs network (black circles) and reading-related language, visual, and cognitive control regions (red circles) during the Stroop task in
Functional connectivity between the EF network and regions related to reading for children with RDs: results suggest positive functional connectivity between the EF network and regions related to cognitive control (e.g., BAs 8, 9, 10, 24, 32), language (e.g., BAs 7, 21, 22, 41, 42), and visual processing (e.g., BAs 17, 18, 19); p < 0.05, FDR corrected. For the BA-related anatomical regions, see Supplementary Table S1 and Figure 2.
Differences in functional connectivity between children with RDs and typical readers: children with RDs demonstrated greater functional connectivity between the EF network and regions related to reading, p < 0.05 FDR corrected, including the left BAs 7, 39 and the right BA 21 in the language network, left BA 19 and right BAs 18, 19 in the visual network, and left BAs 31, 2, 8 and right BAs 9, 35, 43, 30, 31 in the cognitive control network. For the BA-related anatomical regions, see Supplementary Table S1 and Figure 3.
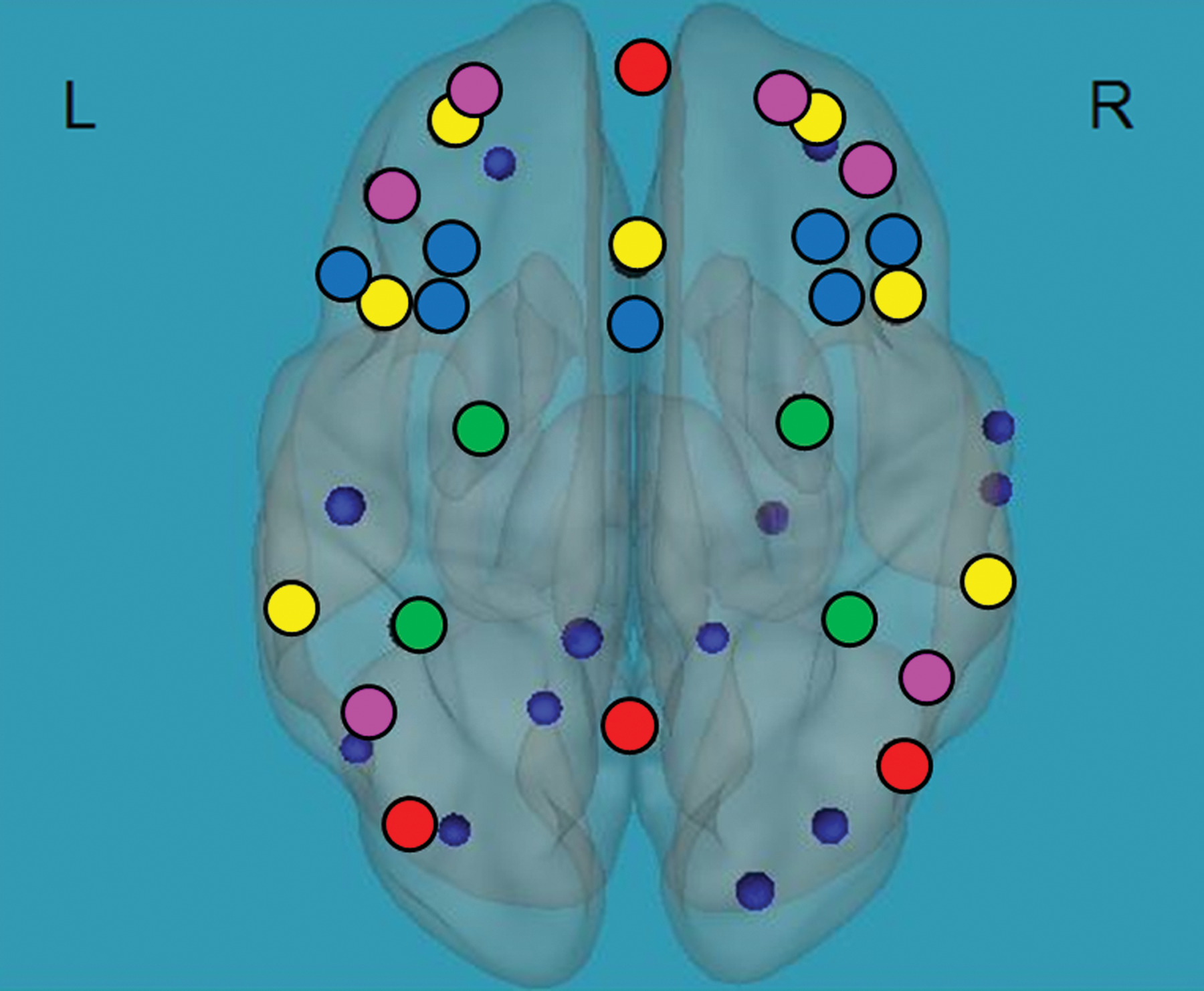
Difference in functional connectivity of the EFs network and reading-related regions in typical readers compared with children with RDs. Negative correlation between functional connectivity of the EFs network (black circles) and reading-related language and cognitive control regions (blue circles) during the Stroop task in children with RDs compared with typical readers. Orientation; L, left, R, right. p < 0.05, false discovery rate corrected for multiple comparisons. The blue circles represent regions that are more functionally connected with the EF network in children with RDs versus typical readers. Color images available online at
Graph theory measures for the EFs network as related to reading and EFs
Functional connectivity within the EF network was measured by the global efficiency measures and calculated for each participant separately, which was then fed into a two-way independent t-test analysis. Global efficiency values did not differ between the groups (mean global efficiency for typical readers: X = 0.667, SD = 0.018; children with RDs: X = 0.66, SD = 0.17, t(45) = −0.75, ns).
A Pearson correlation between global efficiency scores and reading measures among the two groups revealed a positive correlation between timed [TOWRE: r(45) = 0.387, p < 0.01] and nontimed[(letter word, WJ: r(45) = 0.404, p < 0.01] reading measures. Higher global efficiency scores were associated with greater reading ability.
Positive correlation between global efficiency measures and EF measures was also observed [working memory: digit span backwards r = (45) = 0.269, p < 0.05; speed of processing: coding r(45) = 0.355, p < 0.05; letter naming r(45) = 0.339, p < 0.05] and overall switching/inhibition ability [r(45) = 0.306, p < 0.05]. Greater global efficiency measures were related to overall better working memory, speed of processing, and EF measures among the entire study cohort.
Discussion
The aim of this study was to determine the different engagement of the EF network in children with RDs compared with typical readers, as related to their reading and EF abilities. We hypothesized that challenges in EFs in children with RDs would be accompanied by greater functional connections between the EF network and regions related to reading, and greater global efficiency scores would be related to increased reading and EF scores. Results did show that children with RDs performed significantly lower in all reading and EF measures tested in this study. The lower Stroop scores were accompanied by increased functional connectivity of the EF network and regions related to reading in children with RDs, as compared with typical readers. This may indicate a compensatory mechanism for individuals with RDs. Moreover, greater global efficiency of the EF network was related to higher reading and EF abilities in both groups, which highlights the role of the EF network in these academic abilities, as has been previously suggested (Horowitz-Kraus, 2016).
Increased functional connectivity between the EF network and regions related to reading: a possible compensatory mechanism?
Reading is a human invention that demanded an adaptation and a “recycling” of existing neural networks, as previously suggested by Dehaene and colleagues (2010). When examining the “highways,” our brains had to create to read, research shows the involvement of neural circuits related to EFs (Horowitz-Kraus and Holland, 2015; Vogel et al., 2012), language (Horowitz-Kraus et al., 2013, 2015b), and visual processing (Vogel et al., 2013, 2012, 2014). As reading develops, children rely on their language skills, visual and auditory attention, inhibition, memory, and processing speed to comprehend stories read by their parents (see Horowitz-Kraus and Hutton, 2015; Horowitz-Kraus et al., 2017; for review]. This activity of active stories listening was reported to facilitate future reading ability (Bus et al., 1995). These cognitive abilities have to synchronize in time to create the orchestra of reading, and in the case of a failure in synchronization, a reading failure may occur (see Horowitz-Kraus and Holland, 2015).
In addition to several theories to explain RDs, it seems that the overall slowness that characterizes readers with RDs may be related to a much more basic nonlinguistic challenge in EFs (Pennington, 2006; Welsh et al., 1991; Willcutt et al., 2001; and as also indicated by this study). In our study, this challenge in EFs was accompanied by an increased functional connectivity between the EF network and regions related to reading during the Stroop task in children with RDs that may serve as a compensatory mechanism for these readers, as has been observed in other pathologies and with an aging population (Deslauriers et al., 2017). Therefore, increased functional connectivity is often used as a compensation pathway to maintain the level of performance, which seems to still be impaired in children with RDs. However, since we included several EF networks in the current analyses, a future study should examine the relative contribution of each network included in the global EF network to the Stroop task. This contribution should then also be compared with the functional connectivity of each separate network during an fMRI reading task. Interestingly, greater involvement of regions in the left hemisphere was observed in typical readers during the Stroop task, which was not observed in children with RDs (Fig. 2) as was previously observed in the literature for reading tasks (Pugh et al., 2000). It may be that this differential involvement in the left hemisphere in children with RDs was not specific for reading, but generalized also to EF tasks. However, since the current version of the Stroop task did involve an exposure to words (even though the participants were requested to ignore them), an additional study using a word-free EF task specific to lateralization should focus on clarifying this point.
EFs as the infrastructure for reading ability
Previous studies suggest that reading and reading comprehension rely on intact language and EFs (Horowitz-Kraus, 2016). Others showed that an EF training (Horowitz-Kraus and Breznitz, 2009), video games (Franceschini et al., 2013), or EF-based reading training (Horowitz-Kraus, 2015; Horowitz-Kraus and Holland, 2015; Horowitz-Kraus et al., 2014a) improved reading ability in both individuals with RDs and typical readers. These intervention studies showing the positive effect of EF-based trainings on reading in typical readers as well emphasize that this effect is not attributed to the proposed joint impairment in EF and reading in individuals with RDs. It suggests a possible stronger linkage between these two abilities, even in the typically developing child, and future studies examining the relationship between an EF training before reading are acquired and future reading abilities may reveal whether there exists causality between training EFs and reading abilities.
The findings of this study provide the neurobiological evidence for the relationship between EFs and reading and may provide an explanation for previous intervention findings. The global efficiency of the EF network (i.e., the inverse average shortest path length of all pairs of nodes in the EF network; Watts and Strogatz, 1998) was found to be related to higher reading and EF scores. Greater global efficiency reflects the efficiency of the network and stems from improved learning abilities (Yang et al., 2015). These results support previous findings, suggesting that improved reading after an EF-based reading intervention was related to increased global efficiency in the cingulo-opercular network in 8–12-year-old children with RDs (Horowitz-Kraus et al., 2015c). However, the authors suggest that not only the cingulo-opercular network is related to higher reading scores, but also regions in the frontoparietal, salience, and visual-attention networks and the DMN are also critical for better reading and EF abilities. The relative contribution of each of these networks to the reading process warrants further research.
Another intriguing finding is that only orthographic abilities (i.e., timed and nontimed word-reading tests) were associated with the EF network global efficiency measures, but not comprehension. Recent findings regarding the critical relationship between planning abilities and reading comprehension (Georgiou and Das, 2016) and the role of the right hemisphere (as opposed to the left hemisphere for word reading) in reading comprehension both in adults and children (Horowitz-Kraus et al., accepted; Horowitz-Kraus et al., 2014b) indicate the challenge that reading comprehension has over word reading (see also the simple-view theory, Gough and Tunmer, 1986). It may be that selective networks that compose the EF network do not include specific network/regions related to this executive ability. A future study looking specifically into this point is warranted.
As previously suggested, speed-of-processing measures, working memory, and switching/inhibition were positively associated with higher global efficiency measures of the EF network. These findings reinforce previous findings highlighting the importance of fast and efficient words processing, which then decreases the working-memory bottleneck that often occurs in reading impairment (Breznitz and Share, 1992). Visual attention, which was previously reported to be related to reading ability (Facoetti and Molteni, 2001) and improvement after an EF-based reading intervention (Horowitz-Kraus et al., 2015a), was absent in this study. We suggest that the involvement of visual attention should include visual regions, and since these were absent in the EF network, global efficiency of this network did not show positive correlation with visual attention.
Study limitations
The results of this study should be considered with the following limitations. First, although the Stroop task is indeed an EF task, the condition we examined did involve reading. Despite the fact that the participants were explicitly required to ignore the word and respond to the color, reading is involved while performing the task (see Adleman et al., 2002). Therefore, to better demonstrate the neural engagement of EF networks during an EF task, a nonlinguistic Stroop task should be used in an MRI environment. Second, this study examined the functional connectivity and global efficiency of a network composed of several networks. Therefore, a future study should examine the relative contribution of each network to the overall network functional connectivity and task performance in children with RDs. Third, as noted, the definition of EFs is under debate. Although we chose to focus on the Andersen model for the development of EFs (Anderson, 2002) and the Dosenbach model for the corresponding neural circuits to EFs (Dosenbach et al., 2008), one must keep in mind that there are additional neuropsychological theories claiming that working memory and EFs are distinct constructs (Lezak et al., 2012) and that both are unrelated to processing speed (Salthouse, 1996). A future longitudinal neurobiological study examining the functional connections of each of the suggested networks during tasks specifically examining each of the mentioned EFs should be conducted in an attempt to separate the subcomponents of EFs. Another possible limitation is the definition of the EF network based on other networks defined using the CONN network atlas. A future study may be able to define this network using a functional approach (e.g., Cradock et al., 2004, functionally defined parcells that can be the basis for the EF network). Lastly, a controversy still exists between studies suggesting the decreased activation of the DMN and those suggesting an increased activation of this network during cognitive tasks (see Spreng et al., 2010, for further information). Sperg and colleagues suggested the coupled activation of the DMN with the frontoparietal network during a planning task, supporting the inclusion of this network as part of the EF network in this study. However, a future analysis focusing on the role of the DMN might demonstrate the exact involvement of this network in the EF network.
Conclusions
The results of this study suggest that children with RDs more heavily utilize the EF network to perform an EF task than do typical readers and that this may be an attempt to compensate for challenges performing the Stroop task. This connection between reading and EF suggests two important points: (1) A specific EF training even before reading is acquired may “set the stage” for future reading acquisition (e.g., see Rueda et al., 2005, 2012) and (2) functional connectivity of the EF network with the rest of the brain may serve as an early marker for RD. Both points should be examined in depth. Also, the relative contribution of each of the networks separately to the performance an EF task should be determined. The results of this study have several implications related to the educational and clinical fields: (1) The challenges children with RDs share in EFs should be taken into consideration in the classroom: teachers should try to assist children with RDs with avoiding visual and auditory distractions that may harm the child's attention. Since speed of processing is also slow, teachers may choose sharing the questions they are about to ask the student with RD before the formal lesson to allow an adequate processing time. (2) The results support the inclusion of neuropsychological tests related to EFs as part of the reading assessment battery to evaluate the strengths and weaknesses of students with RDs, which may influence their reading achievements as well. (3) Accommodations for children with RDs may not need to be specific only for topics involving reading, but may have to be extended to other topics learned in school that rely on EFs (e.g., mathematics).
Footnotes
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.