
Abstract
Since the discovery of electrical activity of the brain, electroencephalographic (EEG) recordings constitute one of the most popular techniques of brain research. However, EEG signals are highly nonstationary and one should expect that averages of the cross-correlation coefficient, which may take positive and negative values with equal probability, (almost) vanish when estimated over long data segments. Instead, we found that the average zero-lag cross-correlation matrix estimated with a running window over the whole night of sleep EEGs, or of resting state during eyes-open and eyes-closed conditions of healthy subjects shows a characteristic correlation pattern containing pronounced nonzero values. A similar correlation structure has already been encountered in scalp EEG signals containing focal onset seizures. Therefore, we conclude that this structure is independent of the physiological state. Because of its pronounced similarity across subjects, we believe that it depicts a generic feature of the brain dynamics. Namely, we interpret this pattern as a manifestation of a dynamical ground state of the brain activity, necessary to preserve an efficient operational mode, or, expressed in terms of dynamical system theory, we interpret it as a “shadow” of the evolution on (or close to) an attractor in phase space. Nonstationary dynamical aspects of higher cerebral processes should manifest in deviations from this stable pattern. We confirm this hypothesis through a correlation analysis of EEG recordings of 10 healthy subjects during night sleep, 20 recordings of 9 epilepsy patients, and 42 recordings of 21 healthy subjects in resting state during eyes-open and eyes-closed conditions. In particular, we show that the estimation of deviations from the stationary correlation structures provides a more significant differentiation of physiological states and more homogeneous results across subjects.
Introduction
Zero-lag synchronization and desynchronization of neural activity are considered as crucial communication mechanisms of the central nervous system (Engel and Singer, 2001; Fries, 2005; Singer, 1996, 1999; Varela et al., 2001). Such synchronization patterns may occur on various spatial scales, beginning from the integrated activity of several neuronal microcircuits up to the correlated dynamics of different cortical regions. The scalp electroencephalography (EEG) provides access to two spatial scales with excellent temporal resolution: the recording region of a single electrode and the largest spatial scale given by the coverage of the full set of scalp electrodes. Whereas in the first case synchronization manifests in the amplitude and shape of EEG oscillations of each individual EEG trace, the synchronization of the second scale, defining the functional network, is only accessible through the application of appropriate synchronization measures between multiple EEG traces.
In contrast, cerebral electrical activity is highly nonstationary because of the permanent influence of alternating external stimuli and a continuous feedback from internal control mechanisms under ever-changing conditions. Together with the high noise level of scalp recordings, this makes the quantitative analysis of EEGs a nontrivial task, and large fluctuations of the results obtained for different recordings, in particular when measured for different subjects, are not an exception but rather common.
For instance, even for clinically well-defined epileptic seizures one observes an ample panorama of apparently contradictory results (Bartolomei et al., 2002, 2004; Guye et al., 2006; Müller et al., 2011; Schindler et al., 2006, 2008; Wendling et al., 2003). Contradictory reports of increasing or decreasing spatial interrelations during epileptic seizures may originate from different implantation schemes of intracranial recordings, different EEG references, data preprocessing or the usage of different interrelation measures, which underlines the large diversity and pronounced dynamical changes of the interrelation pattern during such events. In particular, there are pronounced qualitative changes of the functional network during epileptic seizures (see e.g., Schindler et al., 2008).
Resting state with open and closed eyes is markedly different in its power spectra (Barry et al., 2007) and blood oxygenation level-dependent (BOLD) dynamics (Zou et al., 2015). However, the difference of eyes-open (EO) and eyes-closed (EC) conditions during rest is not only given by changes of the, for example, Alpha-power (Kaur et al., 2017) but, additionally, measures derived from graph theory indicate marked topological changes of the functional network (Tan et al., 2013). Also functional magnetic resonance imaging (fMRI) studies analyzing eyes-closed and eyes-open conditions reveal dynamical changes, expressed by the evolution of different correlation clusters (Allen et al., 2014, 2018; Damaraju et al., 2014).
Therefore, observations of temporal stable interrelation structures and uniform behavior across a group of subjects such as in the case of the default mode network are evidently important. Most of those findings have been reported for fMRI studies (Buckner et al., 2008; Fox et al., 2005; Greicius, 2008; Greicius et al., 2003; Honey et al., 2009; Jann et al., 2010; Nyberg et al., 1996; Raichle, 2006; Raichle and Mintun, 2006; Raichle et al., 2001; Shulman et al.,1997), a technique with a limited ability to capture dynamical features due to its poor temporal resolution, and only few indications are published on the basis of EEG recordings.
In an early study (Corsi-Cabrera et al., 1997), the EEGs have been recorded in a group of 9 young women in 11 sessions distributed over a period of 1 month to assess the within-subject and within-group stability of EEG correlations. The authors detected temporarily stable correlations of the EEG activity among electrodes, without significant differences between subjects. This result was confirmed by a second study (Corsi-Cabrera et al., 2007), where correlations have been estimated for a group of six young women over a period of 9 months. Within-subject reliability was very high (r > 0.89) and the results were homogeneous across the subject group.
In He et al. (2008), a stable correlation pattern of slow cortical potentials as accessed by electrocorticography across wakefulness, slow wave, and rapid eye movement (REM) sleep has been shown in five epilepsy patients. Surprisingly, the correlation structure has been found to be similar to that of spontaneous fluctuations of BOLD signals, which suggests a direct link between measurements of blood oxygen consumption and electrical brain activity. Also Kramer et al. (2011) reported that stable network templates emerge after as little as 100 sec. The authors claim topological stability, whereas several graph theoretical measures show dependency on physiological states and frequency bands.
In Müller et al. (2014), stable correlation structures have been observed in 20 scalp EEG recordings of 9 patients containing focal onset seizures. The authors show explicitly that this observation is not due to volume conduction, but reflect genuine interrelations stable in time. This skeleton of large-scale interrelations, produced by permanent oscillatory neural background activity, assures simultaneously an efficient coordination of local functional networks as well as large-scale integration and provides an adequate framework capable to coordinate bottom-up and top-down processes.
Although the mentioned early results have been obtained under comparable physiological conditions, the recent results have been found in epileptic brains. It is, therefore, impossible to determine if the stable interrelation pattern is a generic feature of the human brain, just an intrinsic expression of the pathology or the consequence of pharmacological treatment. Therefore, the question of the existence of a stationary pattern in nonpathological brains under normal but physiologically varying conditions persists.
Similar to epileptic seizures also during wakefulness, during sleep and transitions between both states constitute pronounced variations of brain function with profound electrophysiological changes and, hence, present an ideal testing ground for probing such hypothesis. For instance, REM sleep is considered as a state of brain activation where EEG signals show low-voltage fast activity including Beta and Gamma oscillations (Achermann et al., 2016; Llinás and Ribary, 1993; Rechtschaffen and Kales, 1968), whereas non-REM (NREM) sleep is characterized by high-voltage, Delta and Theta oscillations containing special signatures such as sleep spindles and K-complexes (Rechtschaffen and Kales, 1968; Steriade, 1997; Steriade and McCarley, 1990).
In addition to changes of the power of ongoing electrical activity, the temporal coupling among brain regions is modified during sleep stages relative to wakefulness in a state-dependent manner (Achermann and Borbély, 1998; Cantero et al., 2000; Corsi-Cabrera et al., 1987, 2003; De Gennaro et al., 2001; Duckrow and Zaveri, 2005; Guevara et al., 1995; Kaminski et al.,1997; Llinás and Ribary, 1993; Nielsen et al., 1990; Pérez-Garci et al., 2001; Voss et al., 2009). In addition, we consider resting state EEG recordings of healthy subjects while having eyes open and closed, two conditions with remarkable changes of the spectral contents of the electrical brain activity (Allen et al., 2018), that is, with qualitatively different dynamical features.
Hence, due to the fact that sleep recordings as well as the comparison of the eyes-closed and eyes-open state show pronounced changes of the morphology of EEG signals, their spectral contents and the interrelation pattern of different brain regions makes them an appropriate testing ground for probing the existence of a stationary correlation structure.
However, if there is a stationary pattern of functional interrelations among cortical areas in nonpathological conditions, it should be observable independently of the physiological state of a subject, whereas nonstationary dynamical aspects of interrelations should occur as deviations from the stationary pattern.
The first objective of this study is to investigate if a stationary pattern of functional connectivity exists under nonpathological conditions. This has been done by a quantitative comparison of average correlation matrices taken separately for different sleep stages and eyes-open/eyes-closed condition. In addition, we performed a quantitative comparison with the results presented in Müller et al. (2014) to provide further evidence for the independence of the observed pattern from the physiological brain state, even though derived from signals with severe pathological signatures.
Second, we evaluated in which manner the distance to the hypothesized stationary pattern may serve as an improved quantifier of dynamical changes in comparison with an established scheme of analysis.
We hypothesize that (1) if a stable pattern of functional connectivity exists, it will be present independently of the physiological state and show a high similarity to those interrelation structures encountered in epilepsy patients; and (2) that transient dynamical features of the brain activity manifest themselves as deviations from the stable pattern.
Materials and Methods
Experimental design
In total, we consider 72 recordings of 41 subjects, which are divided into 20 recordings of 9 epilepsy patients, 10 sleep recordings of 10 healthy subjects, and 42 recordings of 21 healthy subjects in resting state during eyes-open and eyes-closed conditions. Data have been recorded in four different laboratories, using different EEG equipment and data preprocessing routines.
Sleep data
Sleep of 10 right-handed neurologically healthy subjects recorded in the Sleep Laboratory of the Faculty of Psychology of the National Autonomous University of Mexico participated in this study, after giving written informed consent. Table 1 provides information about the participants and the percentage of time they stayed in each sleep stage.
Information About the 10 Male Subjects of the Sleep Recordings
The age and the percentage of the recording each subject stayed in the unambiguously identified awake or a particular sleep stage.
REM, rapid eye movement; NREM, non-REM.
Before the study, all subjects had a structured clinical interview and kept a 15-day sleep log. Only those with regular sleeping habits and no symptoms of sleep disorders, history of medical, psychiatric or neurological disorders, drug or medication were included. The protocol was approved by the Ethical Committee of the Faculty of Medicine of the National Autonomous University of Mexico and followed the ethical standards of the Declaration of Helsinki (1964).
All subjects slept two nights at the laboratory, the first for adaptation to recording procedures and the second for EEG analysis. Standard polysomnography (PSG) and a standard scalp EEG were recorded at Fp1, Fp2, F3, F4, F7, F8, C3, C4, T3, T4, T5, T6, P3, P4, O1, O2, Fz, Cz, and Pz of the 10–20 International System (Lesser, 1986) referenced to A1 with a Grass 8–20 polygraph with filters set at 0.1 and 70 Hz for EEG, at 10 and 70 for EMG, and 0.3 and 70 Hz for EOG. All night PSG data were digitized and stored with 1024 Hz sampling rate and using a 12-bit A–D converter of the GRASS-GAMMA acquisition program.
Wakefulness and sleep stages were identified by standard procedures using 30-sec epochs (Rechtschaffen and Kales, 1968). Percentages were calculated over the total recording time of the whole night. Sleep stage percentages as shown in Table 1 correspond in general terms with the values expected for young adults (Williams et al., 1974).
Epilepsy patients
EEG data were recorded from nine patients (five men and four women; age range 21–45 years) suffering from pharmacoresistant temporal lobe epilepsy. Subjects have been under presurgical evaluation at the department of neurology of the Inselspital of the University of Bern. The ethics committee of the Kanton of Bern approved this retrospective study. Further, all patients gave written informed consent that their EEG data might be used for research and teaching purposes.
For the EEG recordings standard 10–20 montage positions (Lesser, 1986) were used. After passing an anti-aliasing filter with a cutoff frequency of 70 Hz and an attenuation of 24 dB/oct, the EEG signals were sampled at 200 Hz (seizure 1–12) and 256 Hz (seizure 13–20) using the earlobe reference. A/D conversion had a resolution of 16 bit. EEG seizure onset and seizure offset were visually determined by an experienced electroencephalographer (K.S.) in bipolar montage. Table 2 provides information about patients and seizure durations.
Information About Gender, Age, and Seizure Duration of 20 Peri-Ictal Recordings from Nine Epilepsy Patients
Further information about the EEG recordings from epilepsy patients can be found in Müller et al. (2014).
Healthy subjects in resting state during eyes-open and eyes-closed conditions
The EEG recordings during resting state were provided from two different laboratories and contain in total 21 subjects (Table 3). In all cases, subjects were in resting state for 2 min with closed eyes and open eyes, respectively. When eyes open, they were instructed to look at a mark in the center of a computer screen. All the subjects gave written informed consent.
Information About Gender and Age from 21 Healthy Subjects in Resting State
Eleven elderly healthy subjects (five men and six women; age range 67–94 years) have been measured in the Laboratory of the Clinical Neurophysiology of the Institute National of Medical Science and Nutrition Salvador Zubirán INCMNSZ by using the 10–20 International System (Lesser, 1986) standard scalp EEG at Fp1, Fp2, F3, F4, F7, F8, C3, C4, T3, T4, T5, T6, P3, P4, O1, O2, Fz, Cz, and Pz referenced to earlobe with filter set at 0.1–70 Hz, and the signal were sampled at 200 Hz.
The other 20 EEGs data in resting state (closed eyes and open eyes) of healthy subjects (seven women and three men; age range 23–38 years) were recorded in the Laboratory of Psychophysiology of Cognitive and Emotional Processes of the Institute of Neuroscience of the University of Guadalajara, Mexico by using an Electro-Cap with the same 19 electrodes positions of the 10–20 International System as mentioned earlier. The signals were referenced to earlobe with filter set at 0.1–70 Hz and sampled at 200 Hz and using neuronic acquisition program.
EEG data of all recordings (sleep data, resting state, and recordings from epilepsy patients) were transformed to median reference (Müller et al., 2011, 2014). All EEG signals were filtered to obtain a broadband ranging between 0.5 and 25 Hz to diminish the influence of muscle artifacts by using a fourth-order Butterworth filter. Fp1, Fp2, O1, and O2 were excluded from the analysis because usually these electrodes are mostly contaminated by blink and eye movements and muscular artifacts, respectively.
Statistical analysis
Nowadays there exists a broad palette of different interrelation measures to construct the functional brain network (Boccaletti et al., 2006), usually divided into linear and nonlinear estimators (Galka, 2000; Kantz and Schreiber, 2004; Pereda et al., 2005). Owing to the fact that single neurons, the elementary building blocks of the brain, show pronounced nonlinear dynamical properties (Keener and Sneyd, 1998) one might expect that nonlinear estimators are better suited for the extraction of relevant features from empirical time series. However, for interrelation properties it was proven that even for the favorable situation of low-dimensional stationary nonlinear systems, the performance of linear cross-correlations is highly competitive with several commonly used nonlinear measures stemming from different areas such as information theory, phase space reconstruction, or synchronization measures (Kreuz et al., 2007; Mormann et al., 2005). Also in Ansari-Asl et al. (2006) it was shown that numerically robust but simple linear measures may perform better than sophisticated algorithms, which aim to extract linear as well as nonlinear interrelations.
Based on these findings, we mainly focus in this study on the estimation of zero-lag cross-correlations. Maximum lag correlations are computed additionally to provide evidence that the observed effects are not trivial artifacts of volume conduction.
In all practical circumstances, cross-correlations are estimated over finite data segments, whose length is adjusted to balance between stationarity requirements and statistical accuracy. For a multivariate data set of M EEG channels, the zero-lag cross-correlation matrix is estimated by
Here T denotes the number of samples of the data segment and
Equation (1) can be understood as an average over the product of real values. If the two sets
In fact the situation is even worse. The magnitude of nonzero estimates depends not only on the length of the data window but also on the spectral content of the data, which may change drastically during the time course of an EEG recording (Müller et al., 2011). The larger the contribution of slow frequency components of a signal, the larger is the amount of random correlations (Rummel et al., 2010). This phenomenon, caused by the fact that cross-correlations are estimated over finite data segments (and in many practical applications over quite short segments to improve time resolution), is called “random correlation” (Laloux et al., 1999; Plerou et al., 1999).
Marín-García et al. (2013) proposed a method, which aims to obtain reliable estimates for genuine correlations with a well-defined significance level, based on the use of appropriate surrogate data. It turned out that the so-called significant average correlation (SAC) matrix performs best in terms accuracy of the results, the sensitivity to detect correlations, and the robustness against noise, in comparison with several other proposals. The matrix elements of the SAC matrix are defined as
The sum is just the average over matrix elements of the cross-correlation matrix estimated over NT
data windows, numbered by the index k, each of length T, whereas
To probe the hypothesis, of the existence of a pronounced correlation pattern that is stable in time as in Müller et al. (2014) and universal in the sense of a high similarity across subjects, we studied similarity between average genuine correlation matrices. In the case of the sleep data, we used 30 sec segments that coincided with the so-called sleep epochs used by experienced electroencephalographers (M.C.-C. and I.Y.R.-P.) for sleep scoring. These were divided into
Note that the large length of a segment is due to the fact that in this contribution exclusively broadband signals are considered. For the analysis of, for example, alpha-activity much shorter segments of about 1–2 sec are already appropriate.
To quantify the statistical similarity of two average correlation matrices
Note that the Pearson coefficient estimated for different SAC matrices quantify structural similarity between these matrices, but due to normalization it is insensitive for differences in the magnitude by which a certain correlation pattern is expressed. To test statistical equivalence also for the overall strength of the stationary pattern, we applied additionally the MWW rank test.
If no significant differences between averaged SAC matrices estimated separately for different physiological stages can be found, we concluded that the average over the whole recordings is justified. The result of this overall average is termed “stationary pattern” in the sequel and will be denoted by
To probe stability in time and further to test the hypothesis that dynamical features are imprinted in deviations from
We estimated the SAC matrix using a running window approach along the whole recordings, with a step width of 30 sec for sleep data and 10 sec epilepsy data and recordings of eyes-open and eyes-closed conditions during rest. For each window, we then computed the correlation between the SAC matrix and the stationary pattern
Finally, to probe our second hypothesis, we determined in which manner deviations from the stationary pattern were capable to distinguish different physiological states and compared the results with a classical strategy, namely cross-correlation values itself. For this purpose, we calculated the average of the absolute value of the nondiagonal elements of the SAC matrices
obtained for a particular sleep stage, for the eyes-open and eyes-closed conditions, or, respectively, the different phases of the peri-ictal transition, across all subjects. Thereafter, we computed from this set of real numbers the median and the 95% confidence interval separately for each physiological state.
Finally, we also considered deviations from the average matrix (Equation 2):
Note that to evaluate characteristic deviations of a particular physiological stage, this difference matrix can be used in Equation (3) instead of
Model calculations
To substantiate the interpretation of the observed phenomena, which we provide in the Discussion section, we performed additionally some model calculations of explicit nonstationary dynamical systems. In a first step, we estimated the correlation matrix of two strongly coupled Rössler systems, where each of the variables of the six-dimensional phase space were erratically perturbed by independent Gaussian white noise. Because the random perturbations have been frequently applied (every 250 time steps), the system is almost permanently located on a transient around the attractor and is thus nonstationary by definition. Then we estimated the average of the six-dimensional correlation matrix and compared the outcome with the equivalent nonperturbed system.
In the second case, we deformed the attractor (actually we considered a mixed system) showing that in this case the correlation matrix strongly depended on such deformations. Here time series are derived as a mixture of the strongly coupled Rössler system and a system of two anticorrelated Lorenz oscillators. The mixture has been varied gradually along the time course such that at the first time step the recording consisted of the pure Rössler systems and at the last time step the anticorrelated Lorenz systems remain. In this way, we permanently deform the invariant set of the whole system.
A detailed description of the differential equations, chosen parameters as well as the numerical results can be found in the Supplementary Data (Supplementary Data are available online at
Results
To illustrate certain details of our analysis, we present results of one healthy subject during sleep (subject 9 of Table 1) before we provide summary figures merging the results of all recordings. We start with the search of a pronounced temporarily stable correlation structure.
Existence of a pronounced stationary correlation pattern
In a first step, we searched for stationary correlation pattern. Based on the findings published in Müller et al. (2014), such pattern is supposed to be independent of the physiological state, namely, a particular sleep stage. In Figure 1, we show mean correlation matrices from the recording of subject 9, where averages have been taken over all 30 sec segments of the whole night, but separately for each sleep stage. A similar figure of a young and an old adult with eyes open and eyes closed is presented in the Supplementary Data (Supplementary Fig. S1).
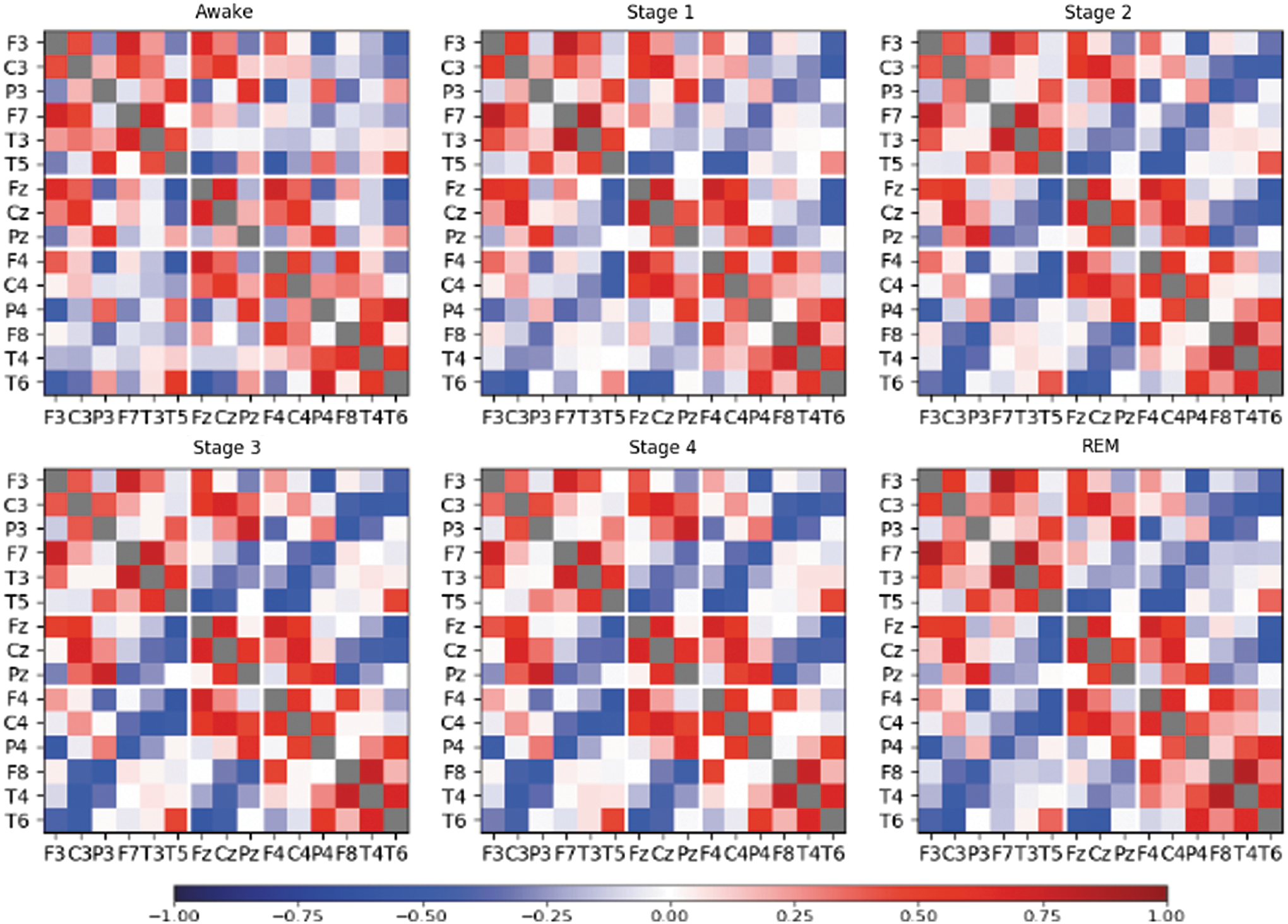
Average correlation matrices of subject 9 of Table 1. Each average is taken over all data segments belonging to a particular sleep stage. Color images available online at
In close similarity with findings derived from the scalp EEG recordings of epilepsy patients (Müller et al., 2014), we here observe a pronounced correlation structure in all sleep stages (as well as for eyes-open and eyes-closed conditions), despite the marked electrophysiological and neurophysiological differences between REM, non-REM, and the awake state. Similar to Müller et al. (2014), positive correlations are more prominent for connections within the same hemisphere, whereas diagonal contralateral electrode pairs tend to be anticorrelated. Considering furthermore the high noise level of scalp EEGs and the nonstationarity of the brain activity, the appearance of high magnitude average cross-correlations is a counterintuitive result.
The intriguing similarity of the pronounced correlation patterns shown in Figure 1 leads to the question whether an average over the whole recording without distinguishing sleep stages is justified. To substantiate such procedure, we estimated for all subjects the correlation between all pairs of the average correlation matrices obtained separately for the sleep stages. We found that average SAC matrices of different sleep stages are indeed highly correlated with Pearson coefficients >0.9. Furthermore, we compared quantitatively the equivalence of the median off-diagonal matrix elements by using the MWW rank test, without finding significant differences.
In consequence, we found that despite drastic physiological differences, marked morphological changes of the EEG recordings between sleep stages as well as alternations of the functional network, we observed a similar pronounced average correlation pattern. The same is true for the comparison of the eyes-open and eyes-closed conditions. Pearson coefficients encountered surprisingly high estimates. Consequently, we conclude according to these findings that the average over the whole recording, termed as “stationary pattern” in the sequel, is justified. Corresponding results and a detailed discussion can be found in the Supplementary Data (see Supplementary Figs. S2 and S3 in the Supplementary Data).
Stability over time and temporal fluctuations
In the following step, we study the temporal stability of the correlation coefficients to further probe for the stationarity of the average correlation pattern. For this purpose, we estimated the correlation between the stationary pattern and SAC matrices estimated with a running window approach and visualized the time course of all 105 matrix elements of the SAC matrix. Results for subject 9 are provided in Figure 2.
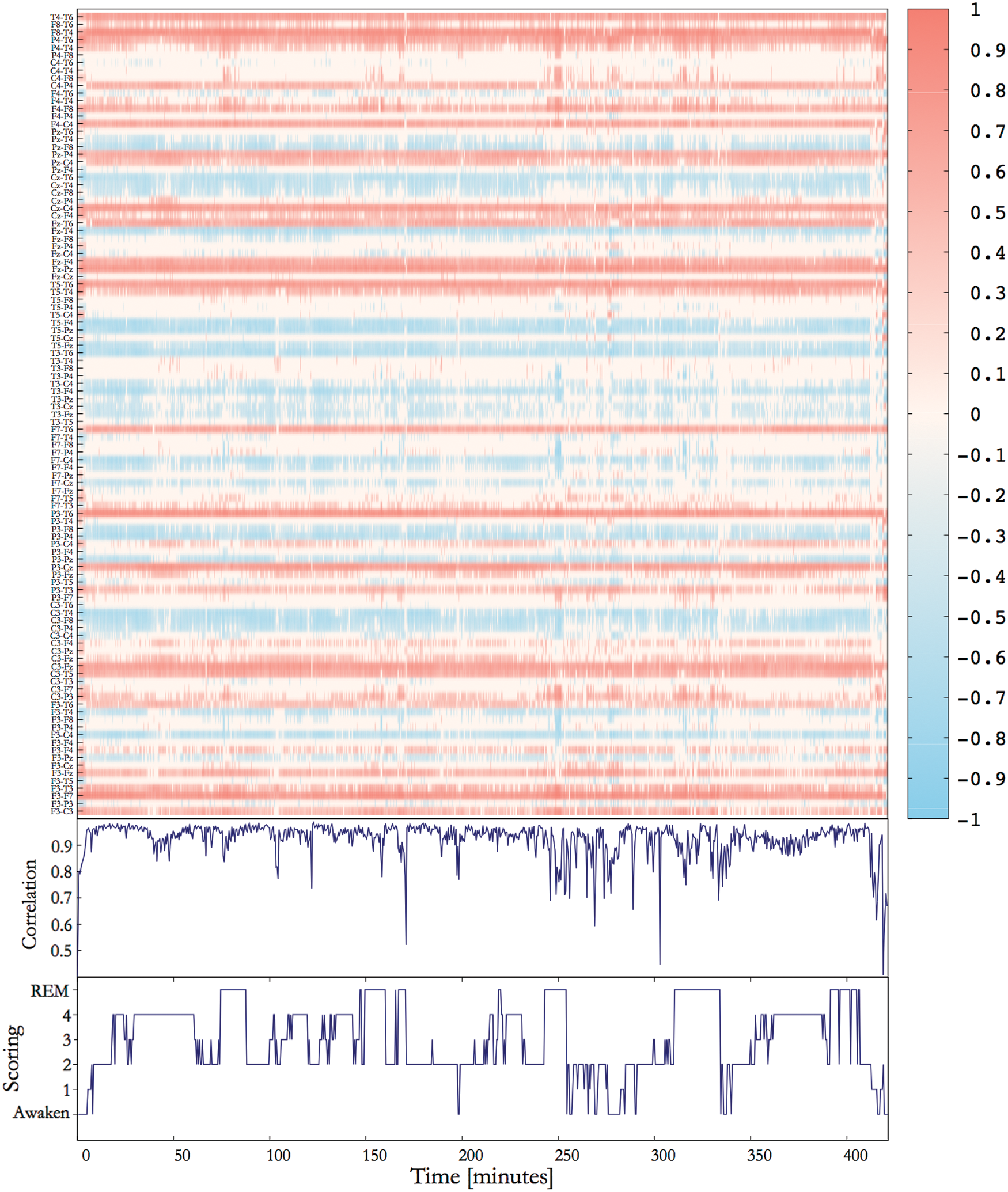
Case study of subject 9. Upper panel: Time evolution of all 105 correlation coefficients of the SAC matrix. Middle panel: Correlation between the time-dependent matrix vectors formed from the SAC matrix and the stationary pattern. Lower panel: Scoring of the corresponding polysomnography. SAC, significant average correlation. Color images available online at
In Figure 2, we observe that the correlation between the SAC matrix and the stationary pattern is almost always >0.85, which constitutes an extremely high value. Only at the beginning of the recording, one notices a gradual increase of correlations up to values of about 0.95. After this initial phase, the correlation appears to be quite stable and even outliers (e.g., during short waking stages between minute 250 and 300) never drop <0.6, which still implies a pronounced similarity. Furthermore, such minimal values are encountered only for short moments and appear as sharp spikes in Figure 2. Besides such short episodes, the correlation of the time-dependent SAC matrix with the stationary pattern is considerably high and fluctuates around 0.9.
The visual inspection of the time course of the SAC matrix elements (upper panel of Fig. 2) confirms these results. Despite pronounced changes of the spectral composition as well as significant topological alternations of the functional network during sleep cycles (Achermann and Borbély, 1998; Cantero et al., 2000; Corsi-Cabrera et al., 1987, 2003; De Gennaro et al., 2001; Duckrow and Zaveri, 2005; Gast et al., 2014; Guevara et al., 1995; Kaminski et al.,1997; Llinás and Ribary, 1993; Nielsen et al., 1990; Pérez-Garci et al., 2001; Voss et al., 2009) the majority of the matrix elements never change their sign and fluctuations around some nonzero constant value seem negligible. Matrix elements with a low average magnitude show noteworthy fluctuations, whereas the overall impression of Figure 2 resembles a remarkably stable correlation pattern. Even transitions between sleep stages are hardly visible.
Note that this observation does not imply a lack of interesting dynamical features imprinted in the EEG signals, and we do not challenge the highly nonstationary character of such recordings. On the contrary, we believe, that the extraction of such nonstationary features provides important and novel progresses for the understanding of the brain dynamics. On this ground, the observation of a stable scaffold of functional connectivity is even more surprising.
The results obtained for all subjects (Fig. 3) corroborate these observations. For all sleep stages, the medians are about 0.85 and fluctuations are remarkably small. Only for the waking state (and partly for the transitional stage 1) one observes a considerable amount of fluctuations. However, the median of the correlation coefficient during waking state is about 0.87 and the lower value of the 95% confidence interval of the cross-correlations is still >0.5. Furthermore, one observes that the fluctuations during sleep stages are not generated by a large discrepancy between subjects. Median values estimated for each subjects separately are almost always >0.9. Only a few subjects and only during sleep stages 3 and 4 encounter values between 0.85 and 0.9 and also the comparably large fluctuations during the awake state cannot be explained by variations across subjects, given that, besides the exception of two subjects, all medians are clearly >0.85. However, 90% confidence intervals are considerably larger for the wake state than for all sleep stages.
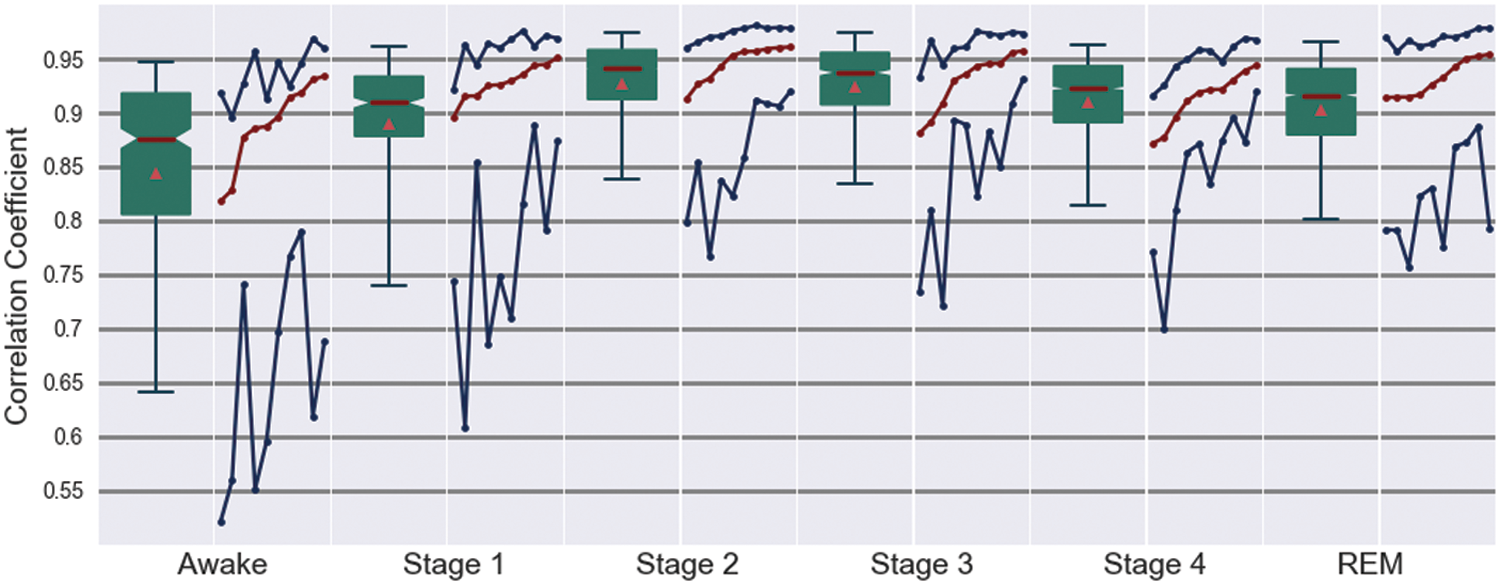
Summary statistics over all 10 subjects of Table 1. For each sleep stage two columns are drawn. The box plot on the left shows median value and the borders of the 50% and 90% confidence interval of the Pearson correlations estimated between the SAC matrix of a given data window and the stationary pattern. Furthermore, as a red triangle, the average value is shown. The right column shows median (red color) and the borders of the 90% confidence interval (blue color) separately for the 10 healthy subjects, ordered by ascending median correlation. Solid lines are just for guidance of the eye. Color images available online at
Based on these results, one may conclude that there exists a kind of rigid skeleton of oscillatory interrelations between brain regions covering the whole scalp. This spatially extended but stable structure could be advantageous for an efficient communication between different, probably distant brain sites. On the one hand, it might coordinate global dynamical features of widely interconnected neuronal populations and additionally it may admit that local deformations of this structure quickly propagate to cover larger spatial scales. Hence, such a skeleton could balance between segregation of local activity and global information integration (Singer, 1999; Tononi, 2004, 2010; Tononi and Edelman, 1998). Consequently, this picture suggests that transient dynamical features should manifest themselves through deviations from this structure. Accordingly, one should expect that the waking state, with a major variety of different kind of activity, is characterized by larger fluctuations around the stationary pattern as shown in Figure 3.
Subject-specific or generic feature
The question remains whether the pronounced stationary correlation pattern represents individual signatures of the brain dynamics of a single person, such as a stationary dynamical fingerprint of neural activity, or, whether it is a generic phenomenon, which reflects general principles of brain functioning. To answer this question, we compare in Figure 4 the stationary pattern obtained for each EEG recording, namely, sleep and resting state EEGs as well as the epilepsy recordings.
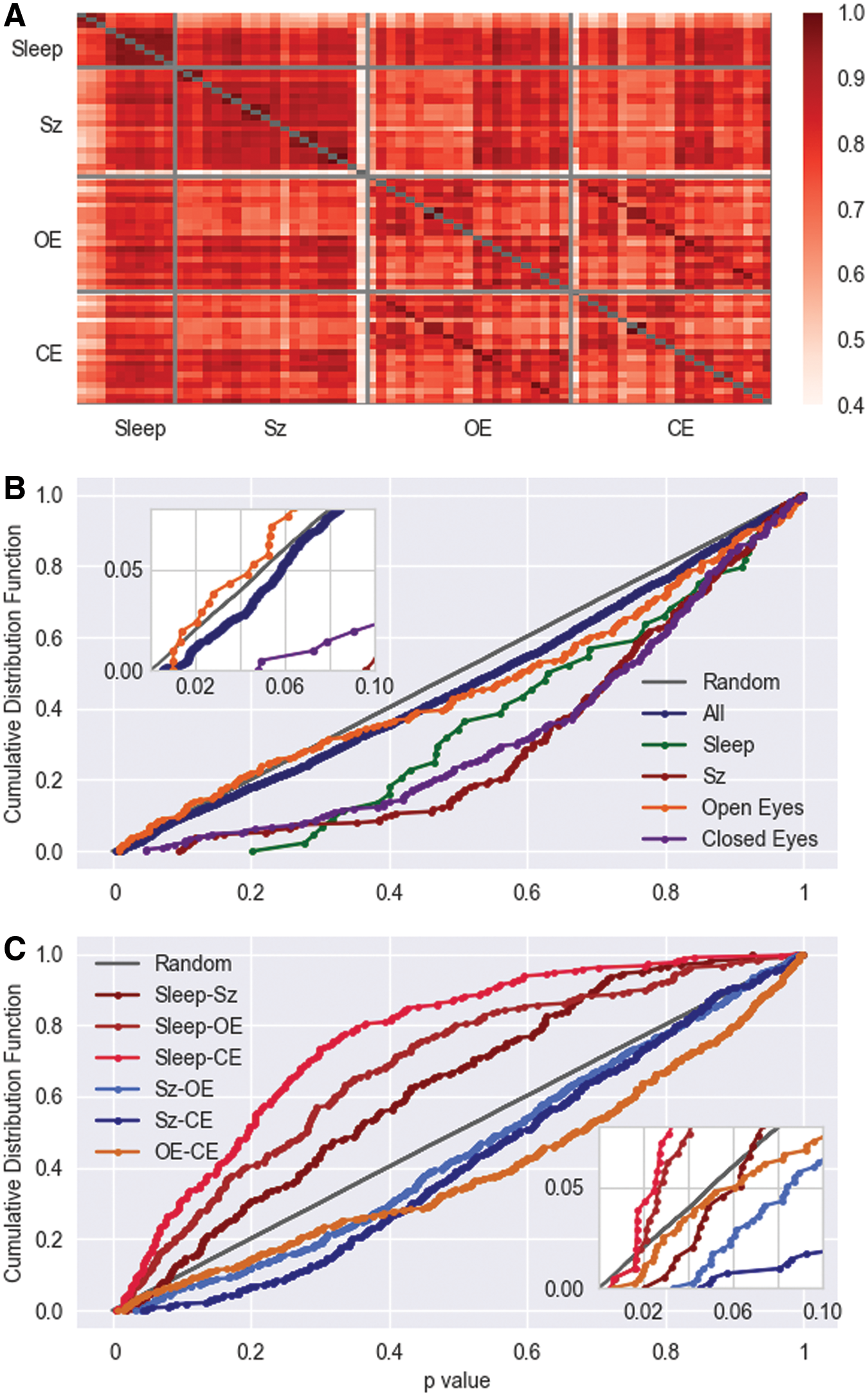
In Müller et al. (2014), it was already found that mean correlation matrices, estimated over periods of several minutes, show also a marked correlation pattern, which, as in this case, seem to be independent of the physiological state of the subject. Namely, average matrices derived from the preseizure, postseizure, or the seizure period show a remarkable similarity [Pearson correlations are always higher than 0.6; Fig. 3 of Müller et al. (2014)] and also peri-ictal averages of different subjects are surprisingly similar. Pearson coefficients are >0.5 [Fig. 4 of Müller et al. (2014)]. Smallest values are obtained exclusively for the comparison with recording 20 of the epilepsy patients, which already has been identified as an outlier (Müller et al., 2014). Furthermore, by visual inspection one gains the impression that the stationary pattern obtained from this pathological activity shares many features with the structures observed for the sleep data.
The Pearson coefficient quantifies topological similarity between the average correlation pattern, which turns out to be considerably high for all cases (Fig. 4A). Only for the pairwise comparisons with the mentioned recording 20 of the epilepsy patients and one recording of the young adults during resting state (EO and EC-condition) the correlation values drop slightly >0.5, whereas, in general, Pearson coefficients are considerably >0.6. Furthermore, a certain structure of the matrix shown in Figure 4A can be identified. First one can observe that the sleep EEG segregates in two clusters (subject 1–3 and 4–10), where the stationary pattern are extremely similar within these clusters, given that the Pearson correlations take values about 0.9, whereas intercluster correlations are, in general, smaller. Furthermore, one identifies a high similarity between the second cluster and almost all remaining recordings, independently whether the pathological case of epilepsy or healthy subjects during rest are considered. The peri-ictal average of the seizure EEGs, in contrast, correlates stronger with the resting state EEGs of older subjects than those obtained for young adults, independently where eyes are open or closed during rest.
But most importantly one observes nonambiguously that also resting state EEGs fall into two similarity clusters, which separate young and old adults. Whereas almost no difference between EO and EC conditions can be noticed for both groups, Pearson coefficients between these groups are systematically lower, which may indicate that although the stationary pattern seemingly does not depend on the physiological state, it may evolve with the age of the subjects.
Although the Pearson coefficient is sensitive for the topological differences of the correlation pattern, the application of the MWW rank test probes similarity of the correlation strength. For comparison and, hence, as a kind of null hypothesis the cumulative distribution of p values for the comparison of white noise samples is drawn. Note that the cumulative distribution of the white noise data indicate the probability that p values occur solely by chance. Hence, a necessary condition for significance is that the smallest p values should lie above the gray straight line.
In Figure 4B, we compare average correlation matrices obtained for the same condition as, for example, all 45 pairwise comparisons of the sleep EEGs. As a result, within the same condition no significant differences can be found. However, when EEGs obtained for different conditions are compared, the sleep EEGs stand out (Fig. 4C). Independently whether they are compared with epileptic seizures, or with the eyes-closed and eyes-open conditions, the cumulative distributions are above the null hypothesis of white noise samples. However, for small p values all three curves drop below the white noise model such that only a few comparisons encounter sufficiently small p values (inset of Fig. 4C). Thus, although the three curves for the comparison with sleep EEGs are notable above the null hypothesis for a large range of p values, no significant differences between physiological conditions can be found. In addition, the results shown in Figure 4 involve multiple testing and require a Bonferroni correction, which lessens the critical p-value by a factor of 200 for the comparison of the sleep and epilepsy data. Thereby none of the obtained p values mark significant differences, a result which is corroborated by the estimation of Pearson coefficients (Fig. 4A). Further comparison of correlation matrices of different physiological stages are shown in Supplementary Figure S4.
In summary, one observes that the similarity between the stationary pattern of the sleep or resting state EEGs and the peri-ictal averages of epileptic seizures is high. Furthermore, one notices that the resting state EEGs fall into two correlation clusters, which do not distinguish between eyes-open and eyes-closed conditions but separate subjects with a notable age difference. Consequently, the high similarity between the average correlation matrices and the apparent statistical equivalence of the set of matrices lead us to the conclusion that the observed pattern is a generic feature and may reflect universal principles of the brain dynamics, which may evolve with the age of the subjects, although we do not provide sufficient quantitative support for this last suspect.
Discrimination between different physiological states
To test the hypothesis that nonstationary dynamical features are imprinted in the deviations from the stationary pattern, we compared the different sleep stages, resting state for open and closed eyes, as well as the preictal, ictal, and postictal phase by estimating the Pearson coefficient of pairs of the difference matrices (Equation 4), averaged separately for different physiological states for each of the subjects. These results should be compared with those obtained for the SAC matrix shown in Supplementary Figures S2 and S3 of the Supplementary Data as well as those presented in Figure 3 in Müller et al. (2014).
The Pearson coefficients for the comparison of the average difference matrix obtained for sleep and wakefulness are shown in Figure 5. In contrast to Supplementary Figures S1 and S2, the color scales of Figure 5 are much larger and thus the differences matrices are distributed over a much wider range, which implies that deviations from the stationary pattern constitute an improved discrimination of sleep stages. Results shown in Figure 5A and B fit to neurophysiological findings about sleep–wake cycles. NREM sleep stages 3 and 4, and to a lesser degree stage 2, are highly correlated, which is consistent with the fact that they belong to the same Thalamo-cortical oscillatory mode (Steriade and McCarley, 1990). Furthermore, one observes that pairs of average difference matrices may be anticorrelated, which indicates that deviations from the average correlation pattern are toward opposite directions in matrix space. For example, anticorrelations are observed for the comparison between the three NREM sleep stages and activated states W and REM as expected from the different physiology of REM, W, and NREM sleep (Steriade and McCarley, 1990). Also the mean difference matrix obtained for stage 1 is anticorrelated to those of stages 3 and 4. If dynamical aspects are imprinted in the deviations from the stationary pattern, the dynamics of anticorrelated sleep stages are qualitatively different. In contrast, REM sleep and stage 1, as well as REM sleep and W share rather similar features, which is consistent with the activated electrophysiological pattern of REM sleep, which hinders the differentiation of W and stage 1 sleep. Until the REMs and the muscle atony allows to recognize them as different brain states with its own physiological characteristics (Steriade and McCarley, 1990).
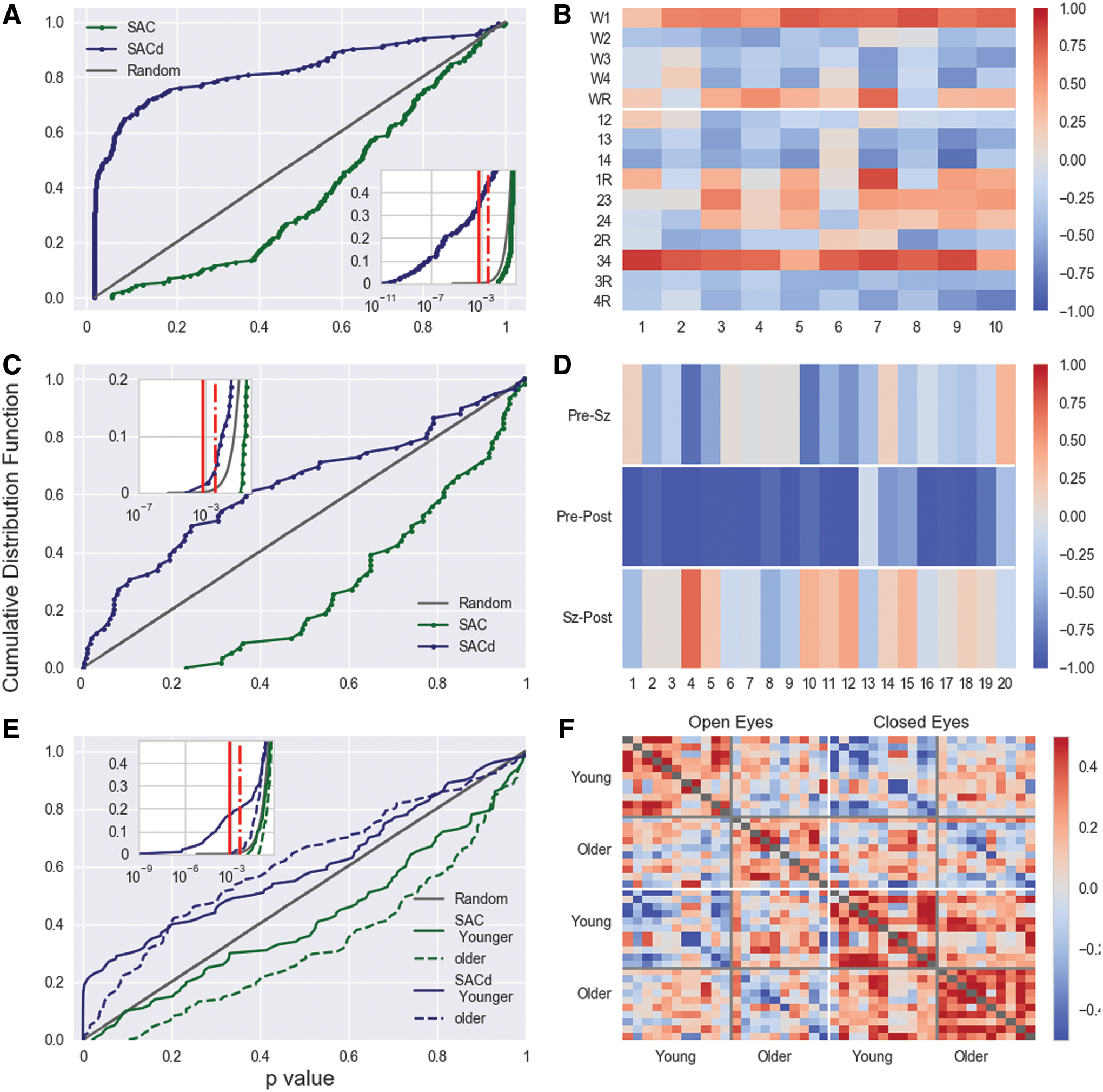
It is conspicuous that stage 2 sleep is identified as qualitatively different from stages 3 and 4 in some subjects. The same is true for the comparison between stage 1 and the awake state. However, these results are not unexpected given that sleep stages 1 and 2 are transitional stages toward stages 2 and 4 as well as REM, respectively.
The p values derived from the MWW rank test confirmed the results obtained for the Pearson coefficients (Fig. 5A). We observed a high discriminative power when deviations from the stationary pattern were considered. About 60% of the results derived for the difference matrix encounter p values <5% significance level. Even when a Bonferroni correction is employed about 40% of the results are still significant on a 5% level and about 33% when a significance level of 1% is desired (inset of Fig. 5A). At the same time, p values of the SAC matrix are all >5% and only six of them encounter values <0.1.
The significant results for the difference matrix are primarily observed for the comparison of sleep stage REM with stages 3 and 4, for the comparisons of stage 1 with sleep stages 2, 3, and 4 and also the comparisons of stages 2 and 4 provide mostly p values <0.05. In principle, striking similarities of the structure of the average difference matrices as displayed in Figure 5 lead also to notable high p values for the MWW rank test. For instance, the comparison of the wake state and stage 1, or the comparison of sleep stages 3 and 4, leads also to high p values when magnitudes of the correlations are under consideration.
In the case of the EEG recordings of the epileptic patients (Fig. 5C, D), one observes largest discrepancies for the comparison of difference matrices of the pre- and postseizure periods. All Pearson coefficients of this comparison are negative, namely, the average deviations from the stationary pattern are diametrically different, a clear indication of the effect the epileptic activity on the electrical brain dynamics even after seizure termination (Gast et al., 2014; Müller et al., 2011). For the direct comparison of the preseizure with the seizure period, no consistent result could be derived. In some cases average deviations are positive, whereas in others negative correlated, which could be due to different durations of immediate preseizure states in different recordings. For the comparison of the postseizure with the seizure period, Pearson coefficients are dominantly anticorrelated, although also in this case some estimates lead to positive values.
For the comparison of the overall correlations strengths (Fig. 5C), one observes qualitatively the same results as for the sleep recordings. Average difference matrices lead to more significant differences between phases than correlation matrices themselves. However, now the outcomes are much less significant than for the sleep data such that almost none of the comparisons (of the pre with the postseizure state) lead to sufficiently small p values.
Finally, the comparison of resting states with open and closed eyes leads to some interesting observations (Fig. 5E, F). First, average distance matrices allow the distinction between the two resting state conditions. In particular for the young adults, Pearson correlations between the eyes-open and eyes-closed conditions are dominantly negative. About 15% of the p values derived from MWW rank test are below the 5% significance level (Bonferroni corrected). For elderly adults, the difference is less obvious and p values are not significant, although also in this case the outcomes turn out to be strikingly worse when average correlation matrices are used.
However, besides the distinction of different physiological conditions, also deviations from the stationary pattern seem to be age dependent. As in the case of Figure 4, correlations between the two groups for either condition take notably lower values. If, as suspected earlier, deviations from the stationary pattern indicate nonstationary dynamical aspects, it could be that not only the stationary pattern itself but also transient electrical brain activity might evolve with the age. However, given that both groups are measured in different laboratories, these remarks are rather speculative and require confirmation through further studies.
In conclusion, when analyzing deviations from the stationary pattern one obtains a more differentiated picture for dynamical changes of different physiological states than by considering cross-correlations itself. We find remarkable quantitative differences between the results derived for the SAC matrix compared with those obtained by the difference matrix. Deviations from the stationary pattern provide a highly improved discriminative power for physiologically distinct brain stages. Thus, the results presented in this article further indicate that the deviation from the temporarily stable average correlation pattern is a more appropriate measure than correlation pattern itself.
Nonstationary dynamical aspects of brain dynamics
In Figure 2, we depict the complete time course of all 105 cross-correlation coefficients of subject 9. This figure underpins the quantitative results concerning the stability in time of the stationary pattern. Apparently, most of the correlation coefficients are extremely stable and specifically do not change their signs. Transitions between sleep stages are hardly visible and the time course of the correlation between the SAC matrix and the stationary pattern encounters almost always high values. As the average cross-correlations prove to be virtually time-independent, we in the following investigate whether nonstationary aspects of brain dynamics are imprinted in the fluctuations of the cross-correlations and, specifically, which cross-correlations reflect those aspects. To answer this question, one may investigate if there is some relation between the mean value of the correlation coefficients and the size of their fluctuations. To this end, we depict in Figure 6 separately for each sleep stage the size of its 95% confidence interval versus the mean value of each correlation coefficient.
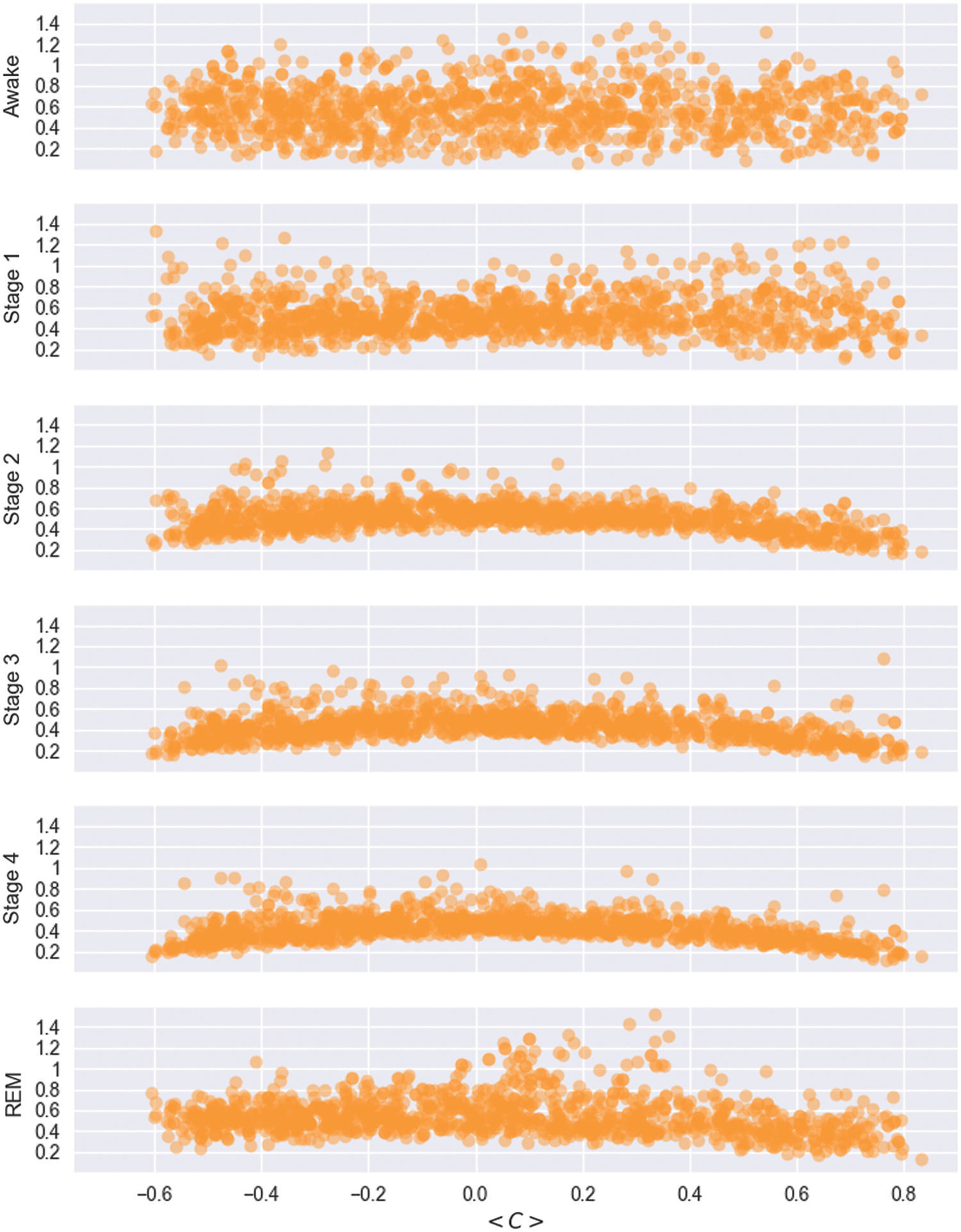
Size of the 90% confidence interval of cross-correlation coefficients in each sleep stage (ordinate) versus their mean value (abscissa) for all 10 sleep recordings. Color images available online at
One observes largest fluctuations and a tendency of lower absolute mean values during wakefulness (see also Fig. 3). The estimates of the widths of the 95% confidence intervals vary between 0.2 and 1.4 almost over the whole range of the obtained mean values. Note that correlation estimates are restricted within the range
This picture changes qualitatively when turning to sleep stage 2. Now one observes a clear tendency of the 95% confidence intervals: correlations coefficients with large dynamical means show smaller fluctuations and vice versa. For estimates of
This behavior is even more pronounced for the distributions obtained for deep sleep, namely, sleep stages 3 and 4. Similar to sleep stage 2, the size of the average value of correlation coefficients is related to the size of its fluctuations. By trend this is also true for REM sleep.
Correlation coefficients with large average values tend to fluctuate less than those with small averages. In particular, states with higher activation levels (i.e. wakefulness, REM-sleep, and sleep stage 1), or those which are more susceptable to external stimulation (i.e. wakefulness and stage 1), or to endogenous and dream-like activity during sleep (REM sleep), show a trend toward higher fluctuations of the correlation coefficients. According to the picture depicted earlier, this result implies that those correlation coefficients with small average values are involved stronger in nonstationary dynamical aspects of the brain activity than large ones.
Discussion
In this article, we reported two unexpected observations. First, analyzing EEG recordings we discovered a pronounced correlation pattern of the oscillatory brain activity, which is stable in time. This pattern is generic in the sense that it does not depend on the physiological state, irrespective of whether subjects are awake, during light, deep, or REM sleep, if they are in different conditions of resting state, or if they encounter a pathological state such as an epileptic seizure. The correlation of the temporally resolved SAC matrix (estimated through a running window approach along the whole night) with the stationary pattern (temporal average over the whole recording) is almost always >0.85 and only within short moments it drops to values of about 0.6, which still implies high similarity. Furthermore, this correlation pattern is generic in the sense that it shows an extraordinary similarity across subjects. This affinity between subjects represents a strong indication that this stationary pattern is due to an intrinsic design of a functional network organization, similar to the default mode network found in fMRI studies, but here expressed by the much faster oscillatory electrical brain activity.
Second, we could provide numerical evidence that nonstationary dynamical aspects of the electrical brain activity are more conveniently described by deviations from this stable pattern than by the conventionally used cross-correlations itself. The discriminative power of the difference matrix in comparison with the SAC matrix is notably higher, such that significance levels may improve by orders of magnitude when deviations are under consideration (Fig. 5A).
We are confident that these observations are not due to technical details of the recording process, particular specifications of the recording equipment, or details of the data preprocessing. In total, we considered 72 recordings of 41 subjects in completely different physiological states. Furthermore, the data was acquired by four different laboratories in Mexico and Switzerland. Also the chosen EEG reference is not responsible for the observations as could be proven in Müller et al. (2014). Also volume conduction can be excluded as a possible explanation. Besides, it is hard to believe that volume conduction causes the same effect with the same magnitude in physiological completely different conditions and, furthermore, that volume conduction generates pronounced anticorrelations. In Müller et al. (2014), it was shown explicitly by estimating maximum lag correlations as well as the weighted phase lag index that the stationary pattern is certainly not due to volume conduction. We repeated such computations for the sleep recordings obtaining similar results (not shown in the figures). Instead of volume conduction, we propose another interpretation of the observed phenomena.
Systems that dissipate energy and interact with its environment such as the brain usually do not exploit the whole variety of theoretically possible dynamical states. They prefer to move in phase space within a restricted region, the invariant set, or the attractor of the system.
The phase space is the set of all (theoretically) possible dynamical states that the corresponding system may encounter. The dimension of this space is given by the minimal number of variables, which are necessary and sufficient for the complete description of the system, namely, for the determination of all possible observables. For instance, in case of a simple pendulum with friction (energy dissipation) and an external force (interaction with its environment) the minimal number of such variables is two, for example, the position and the velocity of the pendulum.
Knowing these quantities, at a given time, one is able to calculate all physical observables such as kinetic and potential energy or momentum. Furthermore, the dimension of the phase space equals the number of degrees of freedom of the system. The pendulum, for instance, describes a closed trajectory in a two-dimensional phase space called limit cycle. If the external force is switched off, the friction causes that the attractor will be just one point in phase space, a stable fix-point, which resembles the resting state of the pendulum. Evidently, more complicated systems may have a much higher dimensional phase space, and in the case of brain dynamics this dimension should be an astronomically high number, given that the brain consists of a highly nontrivial network of about 1011 dynamical units (the neurons) each of them with about 103–104 connections (synapses).
The attractor of a system is invariant in the sense that independent from the starting point of the dynamics in phase space, or, equivalently, independent from the kind of external perturbation the system suffers, the dynamics always approaches the same submanifold of the phase space (provided that the system stays within a basin of attraction).
For the brain dynamics, it is essential that the attractor is not a fix-point. Otherwise, it would have the permanent tendency to switch off and would require permanent energy supply to avoid the definite shutdown. For the brain it is much more convenient that this invariant set contains all those processes, which are responsible for the maintenance of all vitally indispensable activities, such as breathing, control of heart rhythm, temperature control, and digestion. However, neuronal circuits controlling such processes are located in evolutionary older brain regions and the spinal cord, the vegetative nerve system, whose electrical activity is not likely to be measured by a scalp recording. Instead, surface EEG captures mainly electrical activity of cortical neurons.
The cortical brain activity shows electric field oscillations on all temporal and spatial scales. This permanent oscillatory behavior is generated by the intrinsic activity of single neurons and extends to the self-organized motion of large neuronal populations (Nunez, 1995, 2000). Continuously synchronized neuronal ensembles emerge, persist for a while before they decay, making room for the appearance of other synchronization patterns. In this manner, synchronized oscillations of ever-changing neuronal populations weave a network of functional relations even between distant cortical regions, such that information transfer and information processing is facilitated. Thus, the hierarchical arrangements of varieties of parallel loops of neural circuits (Buzsaki, 2006) get bound and local information processing integrated (Nunez, 2000; Tononi, 2004, 2010; Tononi and Edelman, 1998).
In fact, to maintain the brain in an efficient operational mode, a finely tuned balance between activation and rest should be adjusted, such that the brain dynamics approximate a critical state close to a second-order phase transition (Beggs and Plenz, 2003; Chialvo, 2010; Hesse and Gross, 2014; Poil et al., 2008). This balance is expressed by power laws of the, for example, distribution of spatial sizes of synchronization pattern as well as the distribution of live times of synchronized neuronal populations (Eguiluz et al., 2005; Linkenkaer-Hansen et al., 2001; Miller et al., 2009; Poil et al., 2008). In this regime the brain produces a maximal variety of different synchronization patterns, whereas simultaneously spatial correlation encounters the largest length. In this sense, the attractor dynamics would be a kind of dynamical ground state.
Our results show that this variety of synchronized space–time structures are not randomly distributed over the scalp but obey some global order principles such that a temporarily stable scaffold of pronounced cross-correlations emerges. In this way, a specific task-related synchronization pattern does not require to be created within a tissue of almost silent neurons. Rather, already existing space–time interrelations merely have to be modulated. Hence, if the enduring neural background activity can be related to the attractor of the brain dynamics, task-related actions or activities provoked by external stimuli are expressed by a specific adjustment of a permanently existent oscillatory mode. Thus, higher order processes, such as cognitive functions, motor control, or sensory processing, are expressed by deviations from the attractor mode. Taking into account that we are almost permanently subject to external stimuli, this implies that in fact the brain is almost permanently located on transient states, such that the system fluctuates continuously around the attractor in phase space.
This picture is further supported by the brains energy expenditure. Considering that the human brain represents about 2% of the whole body weight its energy consumption with about 20% of the total energy budget is notably high (Raichle and Mintun, 2006). However, this number is independent of the brain's activity and stays rather stable. It was estimated that the local increase of the energy consumption related to the processing of external stimuli varies between 0.5% and 2% (Raichle, 2006; Raichle and Mintun, 2006), a surprisingly low value. Even the fact that only 10% of the synapses carry information of the external world (Peters et al., 1994) does not fully explain such small variations of the energy consumption. However, the scenario described earlier may provide a consistent explanation. If the resting state activity is characterized by cortical standing waves of the electric field (Müller et al., 2014; Nunez, 2000), generated by a continuous alternation of the number of active synapses per unit volume (Nunez, 2000), and the response to demands of the external world merely consists in the modulation of this oscillating mode, one does not expect a huge increase of the energy consumption. Instead, it is plausible that the majority of the energy is used by the attractor activity itself, namely, the maintenance of the oscillatory neural background activity.
If this scenario resembles a realistic picture, the attractor should imprint a characteristic correlation pattern on the average cross-correlation matrix as observed in this contribution, which should be stable in time because fluctuations around the attractor cancel out when averages are taken over long time intervals.
We proved the last statement through some model calculations (Supplementary Data), where we derived correlation matrices from different types of nonstationary dynamical systems. In a first case we investigate a system, which moves almost permanently on transients around a stable attractor (Supplementary Fig. S5). For this system it turns out that deviations from the attractor mode, which manifest as deviations from the unperturbed correlation matrix, cancel out by the time average, such that both correlation matrices are also quantitatively extremely similar (C = 0.98, p-value = 0.346, derived from MWW rank test). This observation is in line with our findings of the average correlation matrices derived from the EEG being extremely stable.
For the second system, we continuously deformed the attractor geometry along the time series (Supplementary Fig. S6). Then we compared the average correlation matrices from the first and second half of the derived time series obtaining striking differences between the average correlation patterns (C = 0.26, p-value = 0.038). By deforming the attractor geometry the cross-correlation structure changes qualitatively in time, a situation that definitely does not reflect the observations reported earlier.
Note that if in both model calculations the information about the deterministic structure is destroyed, for example, through Iterative Amplitude Adjusted Fourier Transform Surrogates (Schreiber Schmitz, 2000), all cross-correlations are destroyed and the distribution of p values follow closely the null hypothesis generated by white noise. Key point of these surrogates is that amplitude distribution as well as the power spectrum of the signals are conserved, whereas Fourier phases are substituted by random numbers, uniformly distributed between zero and
Hence, the first scenario is consistent with what one observes analyzing empirical real-world data. The result of such averages leads to the observation of a pronounced stable correlation pattern, which seems to be independent of the physiological state of the brain. Independently if one considers the sleeping brain in different sleep stages, the brain in the awake state, different conditions of resting state, or drastic pathological activity such as an epileptic seizure, the topology of spatial correlations as well as the overall average correlations result to be extremely stable and statistically indistinguishable.
Finally, such rigid skeleton of large-scale spatial correlations may result beneficial for the orchestration of segregated local functional networks and could provide an efficient mechanism for the integration of local information processing. Waves are entities that implement spatial–temporal structures and large-scale correlation, as observed in the stationary pattern, which allow a fast communication even between distant areas. This oscillatory mode provides a convenient framework for the generation of bottom-up and top-down processes, where local excitations may rapidly propagate due to large-scale correlations provoking global modulations of the standing waves, and, in contrast, local perturbations might be suppressed by a globally collective dynamics.
Note that these findings do not imply that brain activity is a static monolithic process (as proven by the model calculations presented in the Supplementary Data) and this study does not challenge the highly nonstationary character of EEG signals. But it provides a strong evidence for the existence of an attractor of the brain dynamics, a picture that is consistent with abundant literature about the energy consumption and that portrays the brain as a complex system close to a second-order phase transition, a situation most favorable to maintain the brain in an efficient operational mode and transient dynamics provoke excursions in phase space around the attractor.
Hence, if nonstationary dynamical aspects of the brain dynamics are imprinted in transient behavior in phase space, the quantification of deviations from the stationary correlation structure constitutes a more appropriate strategy of analysis than conventional concepts, providing a higher significance level of numerical estimates and a more consistent picture across subjects. So far, we reported solely absolute deviations of broadband signals, averaged over the whole scalp. However, to get an improved description of nonstationary dynamical aspects one needs to look into more details to the spatial distribution of the deviations from the stationary pattern in a time-resolved manner separately for different frequency bands. As shown in this work, the pattern of deviations are qualitatively different for, for example, different sleep stages, that is, corresponding difference matrices are located toward different directions in matrix space, which implies that a qualitative different behavior should be expected for different sleep stages.
Figure 6 documents that preferentially those matrix elements with low average values show dominant fluctuations and, in line with the interpretation of a transient movement around the attractor activity, in particular these matrix elements carry relevant information about the dynamics of nonstationary dynamical features such as higher order cognitive processes or reaction to external stimuli. Although it remains to study which brain areas are dominantly affected by such changes and under which conditions, the present findings already imply that any kind of thresholding, where matrix elements with small cross-correlation values are principally excluded from the analysis, constitute an unfavorable strategy. If, as in many studies, matrix elements less than, for example, 0.3 or 0.4 are systematically discarded, essential dynamical features might be missed, in general.
Another approach that could be used to characterize deviations from the stationary pattern follows a philosophy similar to tradional principle component analysis. If the EEG recordings are projected to the eigenbasis of the stationary pattern, the transformed data sets do no longer contain the observed temporarily stable correlation pattern. Hence, nonzero elements of the SAC matrices constructed from signals transformed this way also indicate deviations from the stationary pattern.
Conclusions
The stationary correlation pattern identified in this study is consistent with the attractor dynamics of the brain activity, which contains the minimal set of vitally indispensable activities and maintains the cerebral cortex in an efficient dynamical mode. In this picture, higher order processes, such as cognitive acts or different physiological states (e.g., different sleep stages), are conveniently expressed by deviations from the stationary pattern. We provided numerical evidence that in comparison with traditional concepts discriminations becomes higher when derivations from the stable correlation pattern are studied.
The observations presented in this article lead to a consistent picture in terms of dynamical system theory and could open new avenues for brain signal analysis, which allow to illuminate novel aspects of the brain dynamics.
Footnotes
Acknowledgments
This work was supported by CONACyT Mexico, Project No. CB-156667. C.R. received support from the Swiss League Against Epilepsy. Recordings of the elderly subjects during rest have been obtained by Mtra. Josefina Montes Rojas and Dr. Estañol Vidal from the Instituto Nacional de Ciencias Médicas y Nutrición Salvador Zubirán.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.