
Abstract
The cingulo-opercular (CO) and frontoparietal (FP) networks are part of the cognitive-control system of the brain. Evidence suggests that over the course of development, brain regions supporting cognitive-control functions become more integrated within their networks (i.e., have increased within-network connectivity), more separated from other networks, and, due to increased maturation along development, are more functionally connected between the networks. The focus of this study was to characterize the developmental trajectory of the CO and FP networks from early infancy (17 months) to 9 years of age in typically developing children while listening to stories, using functional connectivity analyses. Seventy-four children underwent a functional magnetic resonance imaging session while listening to stories inside the scanner. Within- and between-network functional connectivity and graph theory measures were compared during development. Developmental increase in functional connectivity within the CO network and between the CO and FP networks, as well as global efficiency of the CO network from 17 months to 9 years of age, was observed. These findings highlight the involvement of the CO and FP networks in story listening from early infancy, which increases along development. Future studies examining failures in language acquisition to further explore the role of these networks in story listening are warranted.
Introduction
How important is cognitive control?
While sitting in day care during a group activity, a 3-year old is asked to clap her hands three times. How challenging is this request for a 3-year-old child? She has to stop all of her other activities, focus her visual and auditory attention on the teacher, and ignore any distractors. She also needs to process the teacher's verbal request fast and efficiently, plan and execute a response in a timely manner, and use her working memory to count the number of claps so as to correctly complete the task. This set of abilities is referred to as executive functions (EF), an umbrella term for cognitive abilities, needed for learning and adapting our behavior (Horowitz-Kraus et al., 2016), that are used to optimize performance through planning, organizing, and learning (Horowitz-Kraus et al., 2016).
One of the most basic and innate linguistic abilities is the ability to listen to linguistic stimulation (Vannest et al., 2009). Although this ability is intuitive and basic, previous studies demonstrated the utilization of neural circuits related to cognitive control while listening to linguistic stimulation (Horowitz-Kraus et al., 2013).
Understanding the role of cognitive control in language perception during a story listening task is critical due to the inherent relationship between the two abilities. Story listening in children relies on brain regions supporting speech, language, and auditory abilities, along with the ability to attend to and process information included in EF (Horowitz-Kraus et al., 2017a). Verbal language perception relies on dynamic processes of inhibiting incorrect interpretations of spoken language (Gernsbacher and Faust, 1991), avoiding attending to irrelevant information (Lorsbach et al., 1996), and conflict monitoring when presented with confusing ambiguity (Ferreira and Patson, 2007), all of which are basic elements of cognitive control. Recent studies suggest a strong link between language-processing and EF abilities during story listening in preschool-age typically developing children (Horowitz-Kraus et al., 2017a) and in school-age children with reading comprehension difficulties (Roe et al., 2018).
The leading model for the development of EF was suggested by Anderson and Reidy (2012) and Anderson (2002) who demonstrated four main separable components: (1) cognitive flexibility, (2) goal setting, (3) attention abilities, and (4) information processing, all interacting with each other. Although assessment of these abilities is challenging in early childhood, it is possible and crucial to do so due to their central role in a child's cognitive development (Anderson and Reidy, 2012). Interestingly, although academic abilities seem to be dependent on intact EF, EF abilities actually are not fully developed until late adolescence (Casey et al., 2000; Lenroot and Giedd, 2006; Welsh et al., 1991). However, recent neuroimaging evidence suggests the presence and functionality of some EF networks as early as infancy (Gao et al., 2015b).
To understand the effects of the brain on behavior, it is crucial to characterize and trace the changes in neural circuits [i.e., networks of interconnected brain regions responsible for integrating and processing massive amounts of information and functions (Friston, 2011)] that are utilized in narrative comprehension, one of the first linguistic abilities to develop, from infancy through childhood.
Neurobiology and networks related to cognitive control
Cognitive-control research reveals that the main “supporter” for EF is the frontal lobe, more specifically the prefrontal cortex (Welsh et al., 1991). However, in the last decade, the neuroscientific community has identified a wider involvement of several brain regions active in synchrony and supporting EF. The leading networks echoing Anderson's developmental model are the cingulo-opercular (CO) and frontoparietal (FP) networks (Dosenbach et al., 2008; Fair et al., 2007). It has been suggested that the CO network is related to error monitoring and response evaluation, corresponding to attentional control in the Anderson developmental model (Anderson, 2002), and the FP network is related to rapid responding and working memory, corresponding to cognitive flexibility and information processing (Welsh et al., 1991), as represented by the dual-network cognitive-control model (Dosenbach et al., 2008). During story listening, fast processing of the external linguistic stimulus may be examined by looking at the functional connections within the FP network, while in parallel exploring the monitoring processes and the evaluation of the stimuli through changes in the CO network functional connectivity along development.
In a recent review, Grayson and Fair (2017) demonstrated changes in functional brain networks in several networks' properties, including community structure, hub organization, cross-module integration, and global efficiency. Network structure changes gradually from primitive and unorganized, into a denser, more spatially structured organization. Through childhood and adolescence, network refinement continues through stronger connections among functional hubs (Grayson and Fair, 2017). Despite an increasing number of studies examining the functional connections of these networks in adolescence, there remains a gap in knowledge as to the changes in functional connections within and between these networks in early childhood during story listening, which was the focus of this study.
Early childhood and the critical role of language development
The ability to understand language, which involves listening to and comprehending oral language, begins before birth (Dehaene, 2009). Children as young as 3 years of age and as old as 18 years of age demonstrate bilateral activation in the superior temporal gyri while listening to stories (Holland et al., 2007; Horowitz-Kraus, et al., 2017a; Szaflarski et al., 2012). In addition to this bilateral activation, involvement of key regions in the FP network was also evident as early as 3 years of age (Horowitz-Kraus et al., 2017a). Greater speed of processing scores is positively correlated with increased functional connections in key regions in the FP network (e.g., superior frontal gyrus) and increased white matter integrity in white matter tracts, as measured by fractional anisotropy connecting the frontal, temporal, and occipital lobes (inferior fronto-occipital fasciculus) (Horowitz-Kraus et al., 2017a). These results strengthen the involvement of cognitive-control-related regions in language processing, even at a young age (3 years). Due to the enhanced development of both language and EF abilities in infancy (Diamond and Lee, 2011), a question arises as to the change in functional connectivity of the CO and FP networks during story listening from early infancy to later childhood. Revealing the involvement of those networks in such basic abilities is further strengthened by the fact that EF are involved in academic achievements, such as reading and standardized tests like the American College Test (Horowitz-Kraus et al., 2015a).
The aim of the current study was to examine the changes within and between functional connectivity of the CO and FP networks during story listening in 17-month-old to 9-year-old children. Based on previous results on resting-state data (Gao et al., 2015a, b; Grayson and Fair, 2017), we hypothesized that increased within- and between-network functional connectivity and global efficiency would be observed along development, from early infancy to mid-childhood.
Materials and Methods
Participants
Seventy-four typically developing children between the ages of 17 months and 9 years (mean age: 69 months, standard deviation [SD] = 26, 46 females) participated in the current study (Fig. 1). All participants were native monolingual English speakers and passed the compatibility check for magnetic resonance imaging (MRI) participation. Participants had no history of neurological impairments or developmental disorders. Informed consent was provided by the parents of all participants. The study was reviewed and approved by Cincinnati Children's Hospital Medical Center Institutional Review Board.
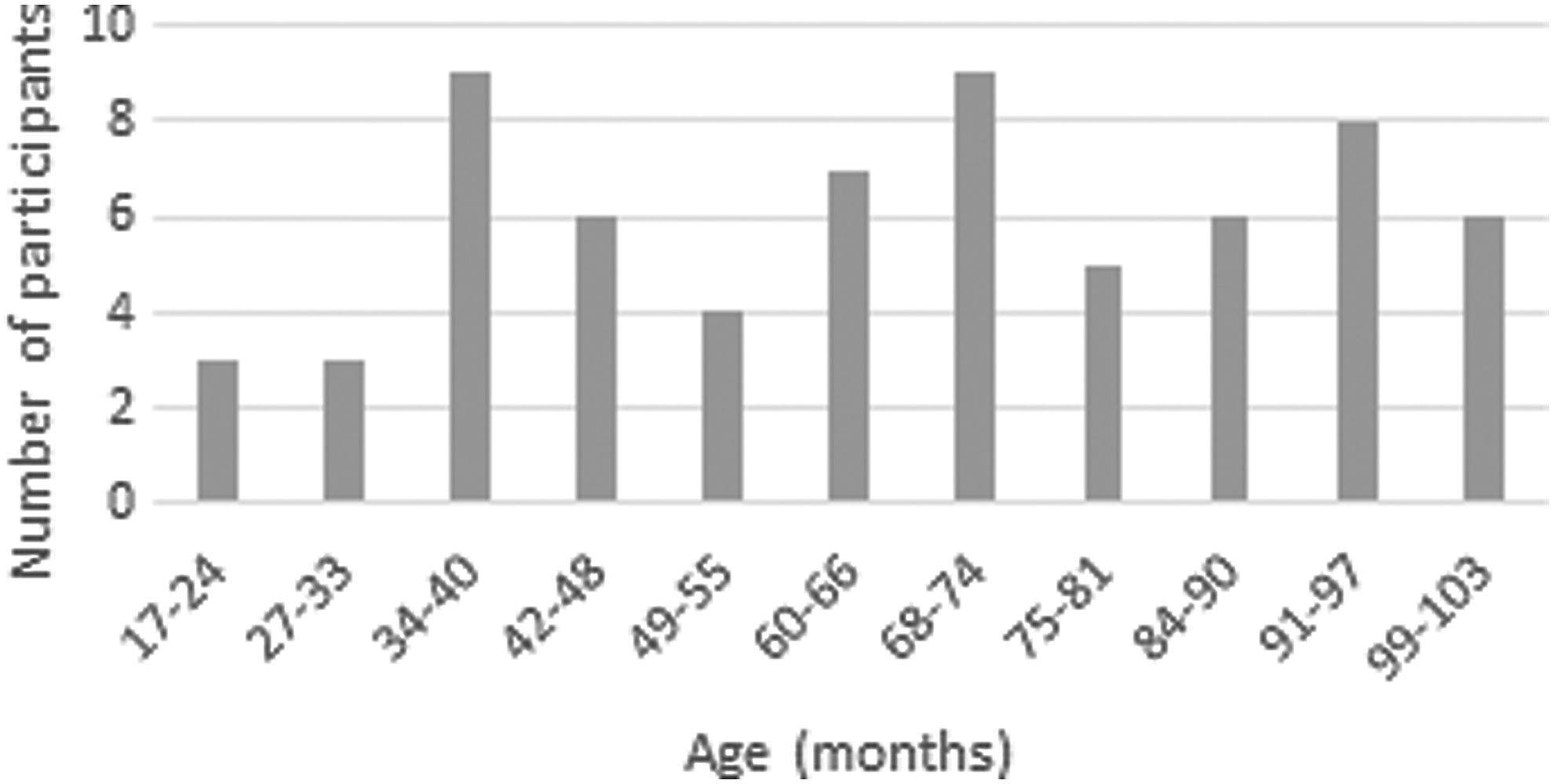
Participant age distribution. The distribution of age in months for study participants.
Behavioral measures
To verify that the nonverbal and verbal intelligence quotient was within the normal range, participants were administered standardized age-appropriate tests, including the Bayley Scales of Infant and Toddler Development (Bayley-III; Bayley, 2006), Wechsler Preschool and Primary Scale of Intelligence (WPPSI-III; Dunn and Dunn, 2007), and Wechsler Intelligence Scale for Children-Fourth Edition (WISC-IV; Wechsler, 2003). To verify that language ability was within the normal range, standardized age-appropriate tests were administered, including the Bayley-III and the Peabody Picture Vocabulary Test, Fourth Edition (PPVT™-4; Dunn and Dunn, 2007).
Neuroimaging data
Story listening task
The story listening task involved passive listening to natural speech stories presented in a female voice. A total of five stories, one for each 30-sec task period, were presented [transcripts of the stories used can be found in Schmithorst and colleagues (2006) and the audio of the stories can be downloaded from
MRI acquisition and data preparation
Participants were acclimated and desensitized to prepare them for comfortable testing inside the MRI scanner [see Byars and colleagues (2002); Vannest and associates (2014) for details]. In short, older participants explored the scanner environment, moved the scanner bed up and down, sat on the bed, and finally practiced being “as still as a statue.” The experimenter asked participants to pay attention to the stories so that participants could answer story-related questions postscan. Younger participants (17–48 months) were acclimated to the scanner using methods specific to nonsedated infants and toddlers. Younger participants and their families participated in a desensitization protocol for several days, which included a brief practice session (Vannest et al., 2014). Head motion was controlled using elastic straps attached to either side of the head-coil apparatus and a headband strap on the child's forehead. Once the child was comfortable, he or she was positioned within the scanner bore and began watching a movie via an MRI-compatible audiovisual system, while image acquisition began. For older children, communication with the study coordinator was established through headphones equipped with a built-in microphone. Verbal communication and positive reinforcement were maintained with the child throughout the scan. Scanning was terminated immediately if the child did not wish to continue. All participants were awake during the entire duration of the scan [see Vannest and colleagues (2014) for details]. Video monitoring and eye tracking were used to ensure that the child was awake and attentive and to assess the child's state and movement inside the scanner's bore. The children were reinforced to keep as still as a statue during the scan session.
All participants were scanned using a 3T Philips Achieva MRI scanner. An MRI-compatible audiovisual system (Avotec, SS3150/SS7100) was used for presentation of the stimuli, as well as for a movie during the preparation (e.g., shimming), and acquisition of the whole-brain anatomical scans. A gradient echo planar sequence was used for T2*-weighted blood oxygen level-dependent (BOLD) functional MRI scans with the following parameters: repitition time/echo time = 2000/38 ms; bandwidth = 125 kHz; field of view = 25.6 × 25.6 cm; matrix = 64 × 64; slice thickness = 5 mm. Thirty-five acquired slices covered the entire cerebrum. Seventy-eight image volumes were acquired during the functional MRI experiment, consisting of 30 sec per condition for a total acquisition time of 5 min and 30 sec. A three-dimensional (3D) T1-weighted inversion recovery gradient echo anatomical whole-brain scan was acquired from each participant for anatomical coregistration and use in spatial normalization of the functional MRI data.
Image quality ratings
Once the imaging session was completed, each set of images was rated on a 0–3 scale by a trained reviewer. Image quality was rated according to predefined criteria previously validated and published (Yuan et al., 2009) and guidelines established by the C-MIND consortium committee (
Quality Rating Criteria for Structural and Functional Images
A description of the rating scale for motion detection in the functional data (upper part) and the anatomical data (bottom part).
4D, four-dimensional; FOV, field of view.
MRI data analysis
Functional MRI data preprocessing
Data preprocessing and first-level analysis were performed using SPM12 (Wellcome Department of Cognitive Neurology, London;
Task-based functional connectivity analyses
Following spatial preprocessing, the task-based data were fed into CONN (Whitfield-Gabrieli and Nieto-Castanon, 2012), a functional connectivity toolbox for MATLAB (The MathWorks, Natick, MA;
Motion-related artifacts were controlled by (1) using images with motion quality rating of 0–1 (see Table 1 for details), (2) applying linear regression and band-pass filtering, and (3) removing different sources of possible confounders (BOLD signal from the white matter and cerebral spinal fluid [CSF] masks, within-subject covariate [realignment and scrubbing parameters], and the story condition effects) (Ciric et al., 2017).
Voxelwise temporal denoising of the BOLD signal was applied by regression of zero- and first-order derivatives of the six motion parameters, regression of the top five principal components of each of the average white matter and CSF BOLD signal, and applying 0.008 Hz high-pass filtering. As connectivity analyses are highly inclined to bias from head motion, especially in the pediatric population, following preprocessing, frames with composite motion >0.9 mm (Siegel et al., 2014) or global mean intensity z-score >±6 were marked as outliers and removed from the data.
Functional connectivity between and within the CO and FP networks was calculated as the mean of the sum of pairwise, Fisher-transformed, bivariate correlation coefficients for all regions of interest (ROIs) within or between the networks.
Defining the CO and FP networks
To determine the functional connectivity within and between the CO and FP networks, we first identified ROIs by defining the nodes of those networks based on coordinates reported by Dosenbach and associates (2008). MNI coordinates were transformed to the Talairach space using a best-fit transform (icbm_spm2tal;

The CO network (right pane—red) and the FP network (left pane—green). Network nodes defined following Dosenbach and associates (2008). The brain networks were visualized with the BrainNet Viewer (
Regions-of-Interest Nodes with Corresponding Coordinates and Brodmann Areas for the Cingulo-Opercular and Frontoparietal Networks
X, Y, Z, coordinates; BA, Brodmann area.
Functional connectivity analysis
Within- and between-network functional connectivity analyses were performed using in-house routines written in MATLAB for the CONN toolbox. Within-network functional connectivity constitutes average overall ROI connections (correlations between ROI time series) within each network, while the between-network functional connectivity averages the ROI-to-ROI connectivity values between all ROI pairs in both networks. Normalized bias-corrected T1 images were generated in SPM (
The global efficiency was calculated in CONN using the following formula (from Latora and Marchiori, 2001):
where Ei is the efficiency of node i, n is the number of network nodes, N is the set of all network nodes, and dij − 1 is the inverse shortest path length between nodes i and j.
Calculation of global efficiency was performed for the CO and FP networks separately for all participants. Correlation analyses of global efficiency with age were conducted in SPSS (IBM Corp., Armonk, NY). Pairwise correlations were corrected for multiple comparisons using an FDR correction [also reported by Benjamini and Hochberg (1995)], with the alpha set to 0.05. The data were normally distributed as indicated by the Kolmogorov–Smirnoff test (Corder and Foreman, 2009) with p > 0.05.
To demonstrate the relationship between global efficiency of the cognitive-control networks and attentiveness, global efficiency of the CO and FP networks was correlated with the full scale intelligence quotient (FSIQ) score from the WPPSI test (Wechsler, 2002). FSIQ score is derived from six subtests summarizing ability across a set of cognitive functions, including verbal comprehension, fluid reasoning, visual-spatial ability (used for imagery processes), processing speed, and working memory. Global efficiency of the brain's functional networks has been linked to attentional capacity and complex cognitive processing in young adults (Grayson and Fair, 2017).
Results
Behavioral results
Composite score from the Bayley Scales was used for younger participants (17–48 months). The cognitive composite score is derived from various sums of subtest scaled scores and is scaled to a metric with a mean of 100, SD of 15, and range from 40 to 160. For older participants, FSIQ composite score was assessed and used. The FSIQ composite score is derived from six subtests and summarizes ability across a diverse set of cognitive functions. All participants performed above average in these tests (Fig. 3).

Participant performance on tests of cognitive ability. Distribution of participant cognitive ability by age. Composite score from the Bayley Scales was used for younger participants, 17–42 months of age. For older participants, FSIQ composite score was assessed. FSIQ, full scale intelligence quotient.
Neuroimaging data
Within-network functional connectivity
CO network
A Pearson product–moment correlation coefficient was computed to assess the relationship between age and functional connectivity within the CO network. A significant positive correlation was determined, r = 0.222, n = 74, p = 0.029 (Figs. 4 and 5).
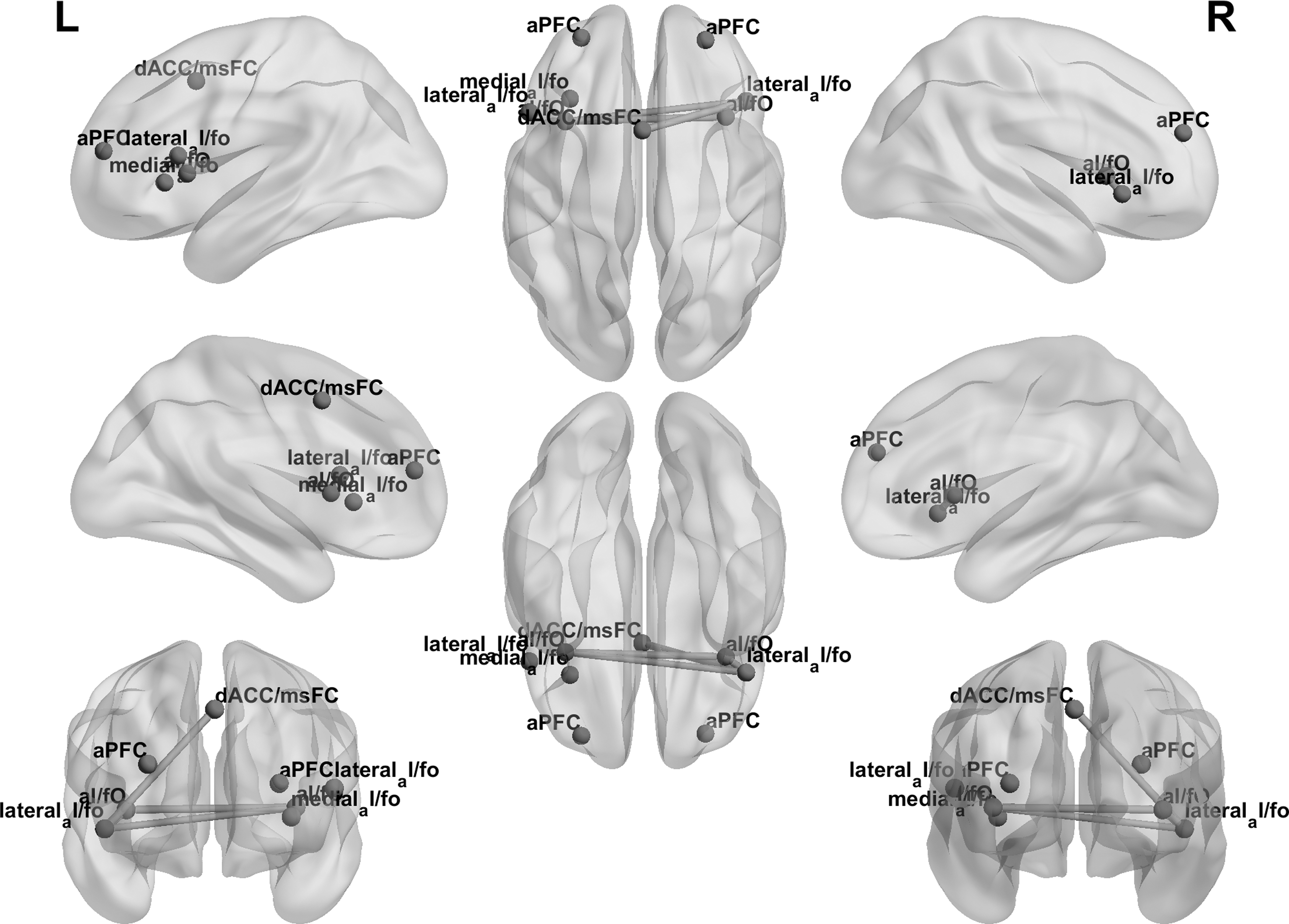
Functional connectivity within the CO network along development. Left and right full views of the CO network along development. Edge thickness represents the strength of functional connectivity between the nodes as a function of age.
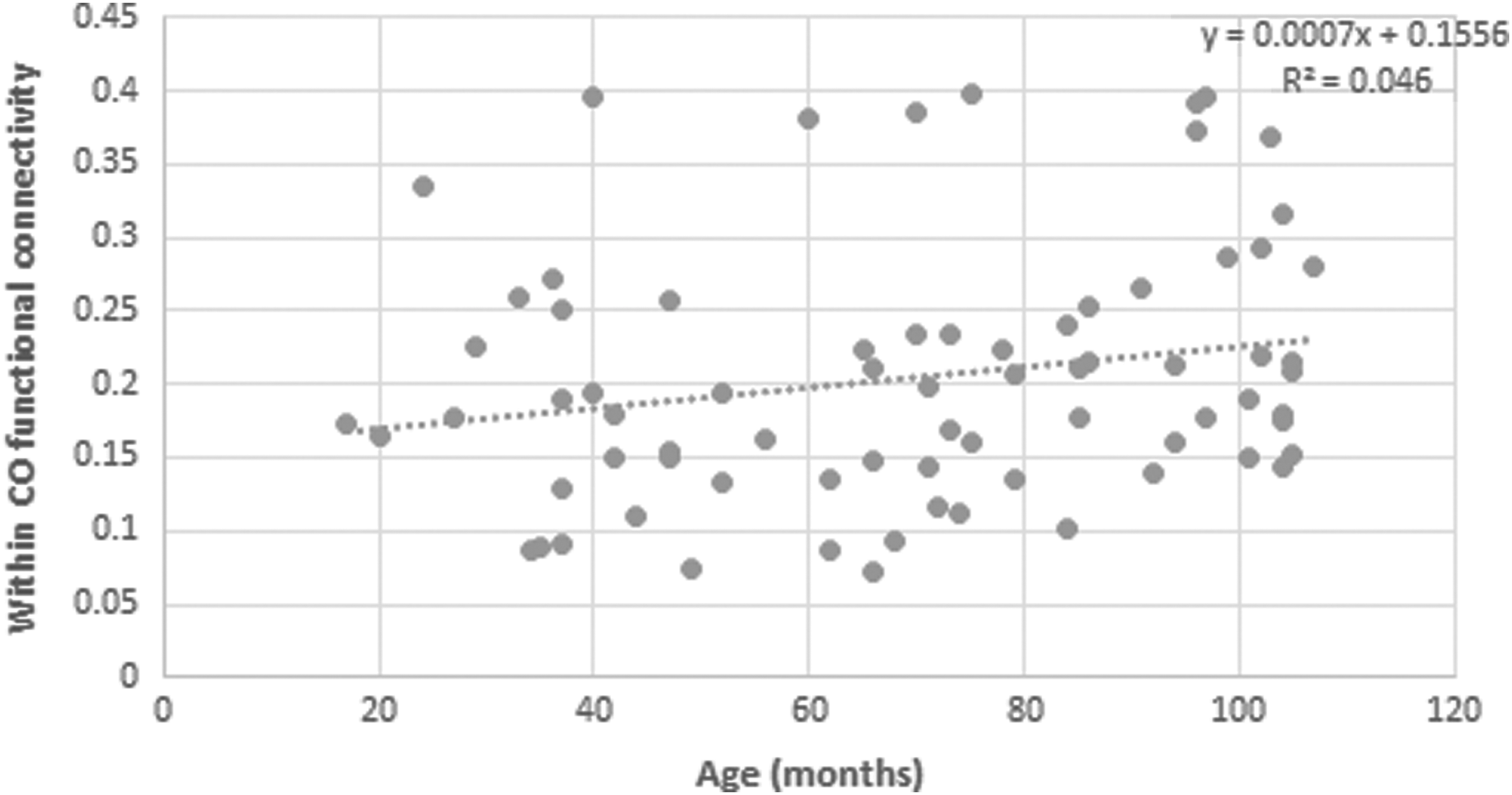
Functional connectivity within the CO network along development. A scatterplot demonstrating functional connectivity within CO network across age.
FP network
A Pearson product–moment correlation coefficient was computed to assess the relationship between age and functional connectivity within the FP network. No significant correlation was determined, r = 0.123, n = 74, p = 0.148 (Fig. 6).
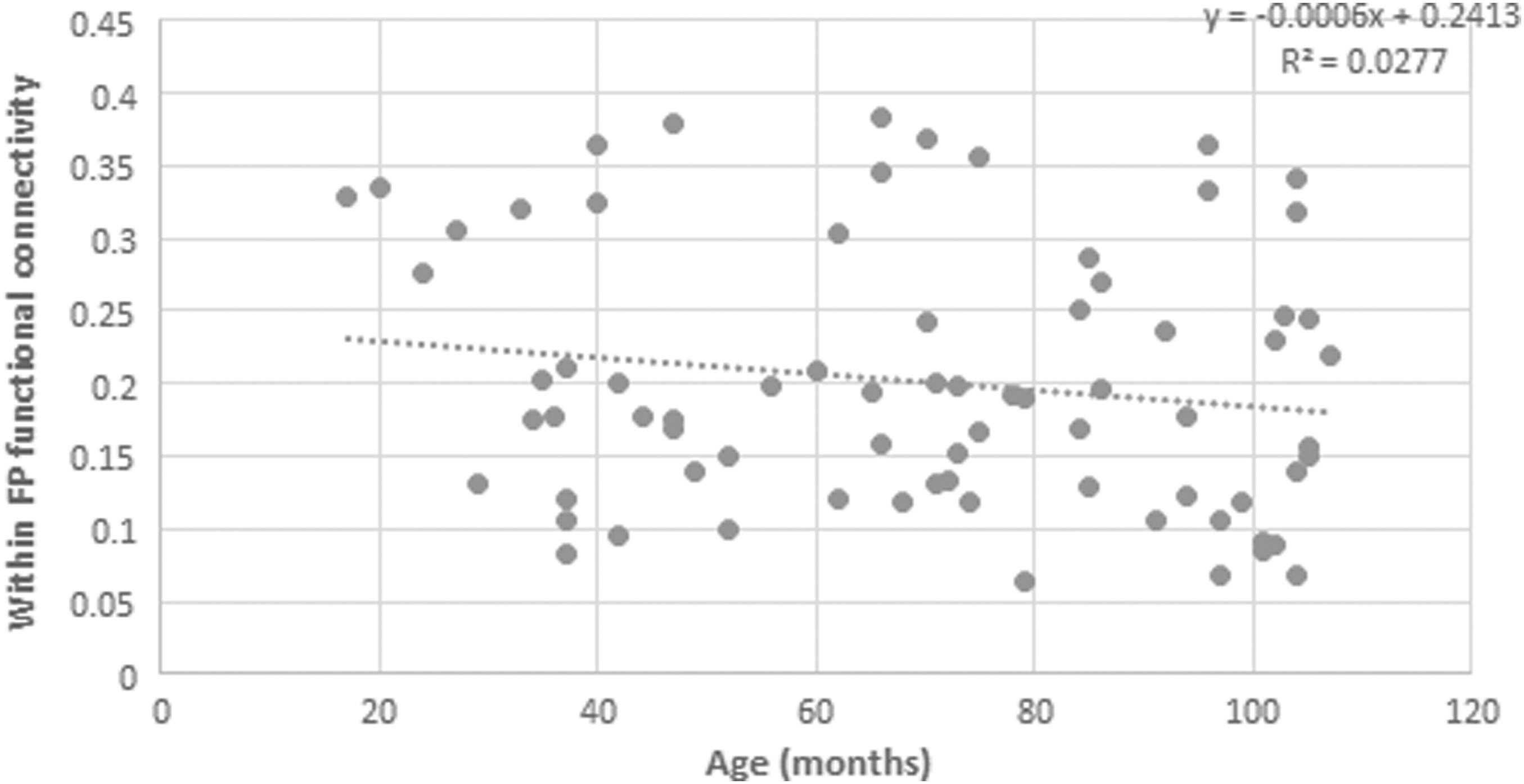
Functional connectivity within the FP network along development. A scatterplot demonstrating functional connectivity within FP network across age.
Between-network functional connectivity
A Pearson product–moment correlation coefficient was computed to assess the relationship between age and functional connectivity between the CO and the FP networks. A significant positive correlation was determined, r = 0.240, n = 74, p = 0.020 (Fig. 7).
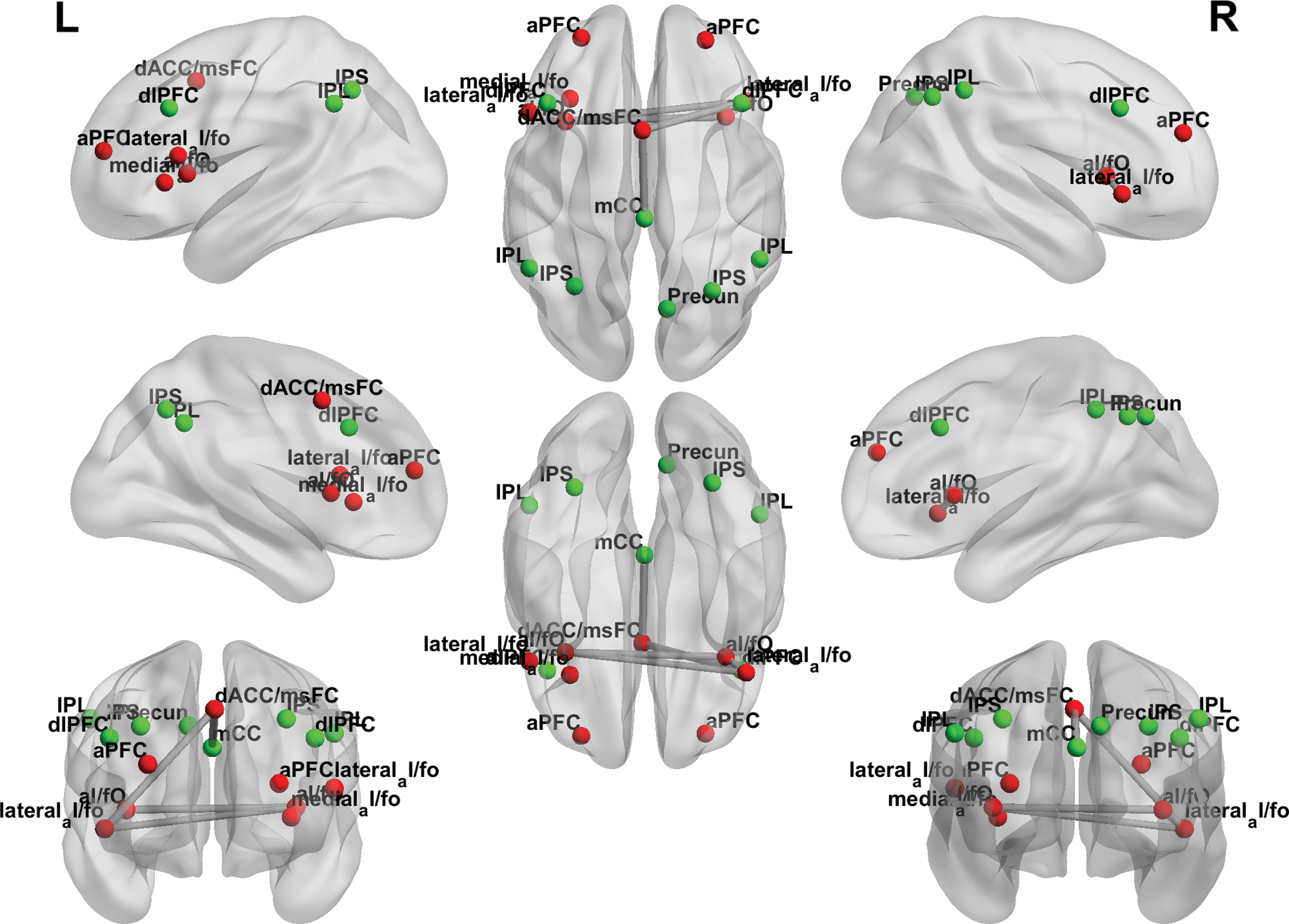
Functional connectivity between the CO and the FP networks along development. Left and right full views of functional connectivity between CO (red) and FP (green) networks along development. Color images are available online.
Global efficiency analyses
Global efficiency was calculated in the CO and the FP networks separately for all participants. Mean global efficiency in the CO network was 0.811 (SD = 0.08) and in the FP network was 0.809 (SD = 0.08). A Pearson product–moment correlation coefficient was computed to assess the relationship between age and global efficiency in the CO and the FP networks separately. Significant positive correlation with age was revealed in the CO network, r = 0.262, n = 74, p = 0.024, but not in the FP network, r = −0.142, n = 74, p = 0.227. Global efficiency of the CO and FP networks was calculated for all participants and correlated with the FSIQ score from the WPPSI test. Significant positive correlation between global efficiency of the CO network and FSIQ score was observed, r = 0.272, n = 74, p = 0.019. No significant correlation between global efficiency of the FP network and FSIQ score was observed, r = −0.084, n = 74, p = 0.478.
Discussion
The aim of the current study was to characterize the developmental trajectory within and between the CO and the FP networks from early infancy to 9 years of age in typically developing children while listening to stories. In support of our hypothesis, greater within- and between-network functional connectivity and global efficiency measures were observed across development.
Providing neurobiological evidence for the Anderson EF model
For over a decade, the developmental neurocognitive community has sought to pinpoint neurobiological evidence for the involvement of neural circuits related to cognitive control in very young children (Gao et al., 2009; Smyser et al., 2010). Several studies accentuated the crucial roles of attention (Posner et al., 2016), working memory (Fitch et al., 2016), and speed of processing (Horowitz-Kraus et al., 2017a) as early as in infancy. These studies provided foundational evidence for EF training in early childhood (Diamond and Lee, 2011; Rueda et al., 2005, 2012; Wardhana, 2016), which corresponds to the Anderson model for the development of EF (Anderson, 2002). As claimed, EF are challenging to assess in early childhood (Anderson and Reidy, 2012), especially since these abilities are supported by neural circuits that do not fully mature until later in life (Lenroot and Giedd, 2006). The results of the current study demonstrate the involvement of cognitive control related to working memory and speed of processing via the FP network and monitoring and attention via the CO network during story listening as early as in infancy. This evidence provides a possible explanation for the positive relationship between cognitive-control networks and language and reading development found in previous studies [see Horowitz-Kraus and colleagues (2017a, b) for a review]. The results of the current study may be used as an example for the typical involvement of the CO and FP networks during language processing to potentially identify alterations in functional connectivity of the cognitive-control networks as possible causes for some linguistic impairments and developmental disorders occurring in early childhood, as was previously suggested in older children (Horowitz-Kraus et al., 2015b, c). This could lead to the discovery of markers for early identification of such disorders (i.e., attention deficit, specific language impairment, dyslexia, others). However, due to the small effects found, these interpretations should be cautiously considered.
Never too young to start: stretching the development of cognitive-control networks to a younger age group
In partial support of the findings by Fair and colleagues (2007), who demonstrated a trend of both decreased short-range connections and increased long-range connections from childhood (mean age 8 years) to adulthood during resting state, our results demonstrate an increased functional connection within and between the two networks from 17 months to 9 years during story listening. An additional study by the same group showed that some of the short-range functional connections (in the same network) increased in strength over aging (Fair et al., 2009). Similarly, Marek and associates (2016) demonstrated both increased organization (within network connectivity) and integration (communication between networks) in older children. A recent study by Gao and colleagues (2015b) demonstrated consistent synchronization of cognitive-control networks in infants by the age of 2 years. Unlike the studies by Fair and colleagues and Gao and colleagues, which used a resting-state condition to demonstrate the functional connectivity between and within the networks, the current study used a linguistic stimulation that required attendance to the stories. It is possible that to attend to stories, greater engagement of each network (within-network functional connectivity) is required in addition to the synchronized engagement of both networks (between-network functional connectivity).
In other words, brain activity differences related to age might reflect both modified use of neural processing units and enhanced specialization in individual processing units along development. This interactive specialization involves both regional developmental changes in neural processing and inter-regional interactions leading to developmental changes (Johnson, 2000; Schlaggar and McCandliss, 2007). An additional study examining the relationship between functional connectivity and narrative comprehension scores should be conducted.
Study limitations
The results of the current study should be considered with the following limitations. First, due to the young age of the participants, no behavioral scores were obtained to assure narrative comprehension. Although this is challenging to compensate for, behavioral standardized testing for language comprehension ability should be obtained and conducted, exploring correlations within and between functional connectivity along development in future studies. Second, the study design was cross-sectional, potentially posing a limitation on inferring developmental changes across age, although previous research has indicated overlaps and discrepancies between cross-sectional and longitudinal designs in the adult brain (Pfefferbaum and Sullivan, 2015).
Conclusions
The aim of the current study was to determine the changes in functional connectivity within and between two cognitive-control networks from infancy to 9 years of age during one of the most basic linguistic abilities—story listening. Although previous studies examined the involvement of language regions and networks during narrative comprehension along development (Schmithorst et al., 2006; Szaflarski et al., 2006, 2012), the focus of the current study was to specifically determine the role of EF while processing language, and therefore, only the EF networks were examined. Based on our results, this approach has the potential to allow an early and objective identification of children who may suffer specifically from EF difficulties, such as children with attention difficulty disorders that may affect their ability to process language. This could enable establishing landmarks for an objective diagnosis of cognitive or language disruption before an actual gap in development occurs.
Footnotes
Acknowledgments
Data presented in this work were obtained from the database known as Cincinnati MR Imaging of Neurodevelopment (C-MIND), provided by the Pediatric Functional Neuroimaging Research Network (
Author Disclosure Statement
No competing financial interests exist.