
Abstract
Alcohol misuse is associated with thalamic dysfunction. The thalamus comprises subnuclei that relay and integrate information between cortical and subcortical structures. However, it is unclear how the subnuclei contribute to thalamic dysfunctions in problem drinking. We investigated resting-state functional connectivity (rsFC) of thalamic subregions in 107 nondependent drinkers (57 women), using masks delineated by white matter tractography. Thalamus was parceled into motor, somatosensory, visual, premotor, frontal association, parietal association, and temporal association subregions. Whole-brain linear regression, each against Alcohol Use Disorders Identification Test (AUDIT) and positive alcohol expectancy (AE) score with age as a covariate, was performed for each seed, for men and women combined, and separately. Overall, problem drinking was associated with increased thalamic connectivities, whereas AE was associated with a mixed pattern of increased and decreased connectivities. Motor, premotor, somatosensory, and frontal association thalamic connectivity with bilateral caudate head was positively correlated with AUDIT score in men and women combined. Connectivity of the right caudate head with frontal association and premotor thalamus was also positively correlated with AE score in men and women combined. In contrast, motor and premotor thalamic connectivity with a number of cortical and subcortical structures showed sex differences in the correlation each with AUDIT and AE score. In mediation analyses, AE score completely mediated the correlation between thalamic caudate connectivity and AUDIT score, whereas the model where AE contributed to problem drinking and, in turn, altered thalamic caudate connectivity was not supported. To conclude, thalamic subregional rsFCs showed both shared and distinct changes and sex differences in association with problem drinking and AE. Increased thalamic caudate connectivity may contribute to problem drinking via enhanced AE. The findings suggest the importance of examining thalamic subdivisions and sex in investigating the functional roles of thalamus in problem drinking.
Introduction
Chronic alcohol use is known to alter cerebral structures and functions. Functional magnetic resonance imaging (fMRI) studies have implicated the thalamus in altered response inhibition (Hu et al., 2015, 2016b; Schmaal et al., 2013b), risk taking (Gilman et al., 2012), reward anticipation (Wrase et al., 2007), and cue reactivity (George et al., 2001) in alcohol users. These psychological processes are central to the etiologies of alcohol misuse and the thalamus appears to be particularly vulnerable to the influence of alcohol (Pitel et al., 2015), as shown in studies of thalamus development and thalamus-specific neurological defects in fetal alcohol spectrum disorder (Donald et al., 2015, 2016; Nardelli et al., 2011; Rodriguez et al., 2016). Thus, there is a need to better understand the role of thalamic dysfunction in problem drinking.
The thalamus comprises subnuclei with distinct anatomical connections that relay and integrate information between cortical and subcortical structures (Behrens et al., 2003). For example, the medial dorsal nucleus responds to reward anticipation and mediates working memory and executive control, which are often compromised following excessive alcohol consumption (Chanraud et al., 2007; Pitel et al., 2015). The anterior thalamic nucleus is part of the Papez circuit and supports episodic memory and emotional expression (Bubb et al., 2017). Deficits in episodic and emotional memory are key manifestations of alcohol dependence (Kim et al., 2009; Pitel et al., 2015). Connected directly with the motor cortex, the ventral lateral thalamus gates motor command in action control. Alcoholics demonstrated cognitive motor deficits in relation to cortical pontine cerebellar circuit dysfunction that involves the ventrolateral thalamus (Ide and Li, 2011; Pitel et al., 2015). Other thalamic nuclei, including the pulvinar that supports attention and cross-modal integration of information (Sherman, 2016), are all likely to be influenced by chronic alcohol use. On the contrary, no studies to our knowledge have systematically examined how specific thalamic nuclei may be impacted by problem drinking.
Resting-state functional connectivity (rsFC) characterizes the intrinsic functional organization of the brain. Studies have elucidated thalamic functional organization and revealed connectivity patterns that conform to anatomy (Ansari et al., 2011; Ji et al., 2016; Zhang et al., 2010). Numerous rsFC studies have described how thalamic circuits are compromised in neuropsychiatric conditions, including substance use disorders (Fein et al., 2017; Kim et al., 2009). However, rsFC findings are mixed as to the effects of alcohol misuse, with studies reporting both increased (Müller-Oehring et al., 2015; Shokri-Kojori et al., 2017; Zhu et al., 2017) and decreased (Camchong et al., 2013; Kim et al., 2009; Wang et al., 2016) thalamic connectivity in drinkers relative to nondrinkers. The various methods used to quantify rsFC such as independent component analysis and seed-based correlation may underlie some of the discrepancies. The majority of studies investigated the whole thalamus as a seed region and none to our knowledge parcellated the thalamus into more than two subregions (Camchong et al., 2013; Kim et al., 2009, 2017). Furthermore, previous work largely did not consider the effects of sex, known to play a major role in determining the neurobehavioral consequences of alcohol misuse (Nixon et al., 2014; Padula et al., 2015; Petit et al., 2017; Seo et al., 2011; Squeglia et al., 2011). For instance, women appeared to show more rapid progression to problem drinking and severe structural brain changes, despite consuming less alcohol, as compared to men (Flannery et al., 2007; Hommer et al., 2001; Pfefferbaum et al., 2009). Thus, considering subregional variation and sex may help clarify changes in thalamic rsFC in alcohol drinkers.
The great majority of imaging studies are conducted in alcohol-addicted individuals and the literature of thalamic function and dysfunction in relation to social drinking is relatively scanty. Alcohol ingestion compromised inhibitory control and altered corticostriatal thalamic cortical activities in social drinkers performing the stop signal task (Nikolaou et al., 2013). In social drinkers, alcohol compared to placebo increased risk-taking behavior, particularly among participants who experienced more stimulating effects of alcohol, and diminished caudate, thalamus, and insula response to feedbacks in a risk-taking game (Gilman et al., 2012). Hippocampal and thalamic activations during working memory were positively correlated with obsessive/compulsive drinking measures in both dependent and nondependent drinkers (Vollstädt-Klein et al., 2010). In a multimodal imaging study of social drinkers, baseline metabotropic glutamate receptor 5 (mGluR5) availability in the thalamus, caudate, and prefrontal cortices was positively correlated with experienced “high” during acute alcohol intake (Leurquin-Sterk et al., 2018). As alcohol inhibits N-methyl-d-aspartate (NMDA) signaling via intracellular mGluR5 pathways (Holmes et al., 2013), the study suggested a role of the thalamic cortical circuits in modulating the reinforcing effects of alcohol (King et al., 2011) and positive alcohol experiences in social drinkers.
Positive alcohol expectancy (positive AE, abbreviated as AE) increased alcohol use (Brown et al., 1985; Sebold, 2017). AE facilitated attentional bias to alcohol-related cues and predicted alcohol consumption and hazardous drinking in adolescents and adults (Cranford et al., 2010; Field et al., 2011; Grotmol et al., 2010; Lee et al., 1999; Palfai and Wood, 2001; Pastor and Evans, 2003; Urban et al., 2008; Wardell et al., 2012). Problem and binge drinkers demonstrated significantly higher AE than nonproblem drinkers (Townshend et al., 2014; Vilenne and Quertemont, 2015). These studies support AE as a psychological risk factor and predictor of alcohol misuse. AE is a complex construct and involves affective motor processes, such as attention, arousal, as well as an urge to seek alcohol, all likely to engage the thalamus. We posited that problem drinking and AE are associated with both shared and distinct patterns of thalamic connectivities.
In the present study, we examined thalamic rsFC of 107 nondependent social drinkers. As previous rsFC work has focused on dependent (Camchong et al., 2013; Jansen et al., 2015; Kim et al., 2017; Morris et al., 2018; Schmaal et al., 2013a; Sjoerds et al., 2017; Wang et al., 2016; Zhu et al., 2017) or heavy drinkers (Shokri-Kojori et al., 2017; Vergara et al., 2017), studying nondependent social drinkers may provide critical information on thalamic dysfunction in early habitual alcohol use. We segmented the thalamus into seven subregions according to tractography-derived templates (Behrens et al., 2003) to examine the roles of individual thalamic divisions. To examine sex differences, we performed the analysis on all participants as well as on men and women separately. Sex differences in the relationship between thalamic rsFC and problem drinking or AE were tested directly by comparing the slopes of regressions. In particular, we identified thalamic connectivities in relation to both problem drinking and AE and used mediation analyses to examine whether thalamic connectivities contributed to AE and led to problem drinking or, alternatively, AE increased problem drinking and resulted in altered thalamic connectivities.
Methods
Participants and clinical assessments
Participants were recruited by advertisements in local newspapers and radio stations and by flyers posted in the greater New Haven area. One hundred seven (57 women, mean age = 30.9 ± 12.3 years) nondependent adult drinkers participated in the study. All were without major medical illness, past or present neurological or Axis I psychiatric illnesses including substance use disorders (Structural Clinical Interview for Diagnostic and Statistical Manual IV Axis I Disorders), denied current use of illicit substances, and showed negative urine toxicology tests for stimulants, opioids, marijuana, and benzodiazepines at the time of initial screening and fMRI. Candidates currently using any psychotropic medications were excluded. Pregnant or lactating women were also excluded. Participants were required to be free of MRI contraindications based on the Yale Magnetic Resonance Research Center's safety guidelines. All subjects signed a written informed consent, in accordance with a protocol approved by the Yale Human Investigation Committee.
The Alcohol Use Disorders Identification Test (AUDIT) was used to assess drinking behavior (Babor et al., 2001). Twenty-five of the 107 participants showed an AUDIT score ≥8, a score considered a positive screen for alcohol use disorders (Bradley et al., 1998; Chung et al., 2000; Fiellin et al., 2000). Participants were also assessed for years of drinking as well as average frequency of drinking and number of drinks consumed in a month during the past year. Participants were assessed with the Alcohol Expectancy Questionnaire (AEQ-3; George et al., 1995). The AEQ-3 consisted of 40 items to address both positive (six subscales: global positive, social and physical pleasure, social expressiveness, sexual enhancement, power and aggression, tension reduction and relaxation) and negative (two subscales: cognitive and physical impairment, careless unconcern) expectancy, confirmed by factor analysis and invariant across sex and race. Each subscale contained four to six statements to be endorsed on a six-point scale, from “disagree strongly (1)” to “agree strongly (6).” Despite the eight factors identified and confirmed with factor analysis, ratings on the eight factors were highly correlated, with a Pearson correlation coefficient of 0.42 to 0.92 (mean = 0.78) pairwise among all subscales (George et al., 1995). Thus, the authors acknowledged that the discriminant validity among the eight subscores was at best moderate. We used the total score of the six positive expectancy subscales to index AE. A summary of demographic and clinical measures is presented in Table 1.
Demographic and Clinical Data
All values are mean (SD).
p < 0.05.
AUDIT, Alcohol Use Disorders Identification Test; AE, alcohol expectancy: sum of all 6 positive subscores.
Imaging protocol and data analysis
Conventional T1-weighted spin-echo sagittal anatomical images were acquired for slice localization using a 3-Tesla scanner (Siemens Trio). Anatomical images of the functional slice locations were next obtained with spin-echo imaging in the axial plane parallel to the anterior commissure/posterior commissure (AC-PC) line with repetition time (TR) = 300 msec, echo time (TE) = 2.5 msec, bandwidth = 300 Hz/pixel, flip angle = 60°, field of view = 220 × 220 mm, matrix = 256 × 256, 32 slices with slice thickness = 4 mm, and no gap. Functional blood oxygenation-level dependent (BOLD) signals were then acquired with a single-shot gradient echo echo-planar imaging sequence. Thirty-two axial slices parallel to the AC-PC line covering the whole brain were acquired with TR = 2000 msec, TE = 25 msec, bandwidth = 2004 Hz/pixel, flip angle = 85°, field of view = 220 × 220 mm, matrix = 64 × 64, 32 slices with slice thickness = 4 mm, and no gap. Three hundred images were acquired in the resting state during which participants were instructed to close their eyes but stay awake for a period of 10 min (Farr et al., 2014).
Brain imaging data were preprocessed using the same routine as described in our previous work (Kline et al., 2016; Li et al., 2014; Zhang et al., 2012; Zhang et al., 2017; Zhang et al., 2018), using SPM12 (
As extensively investigated earlier microhead motion (>0.1 mm) may represent an important source of spurious correlations in rsFC analysis (Van Dijk et al., 2012). We applied a “scrubbing” method to remove time points affected by head motions (Power et al., 2012). Briefly, for every time point t, we computed the framewise displacement given by
Seed regions for connectivity analyses
We used the templates of seven thalamic subregions as seeds, each with distinct cortical and subcortical projections based on diffusion tensor imaging (Behrens et al., 2003; Fig. 1).
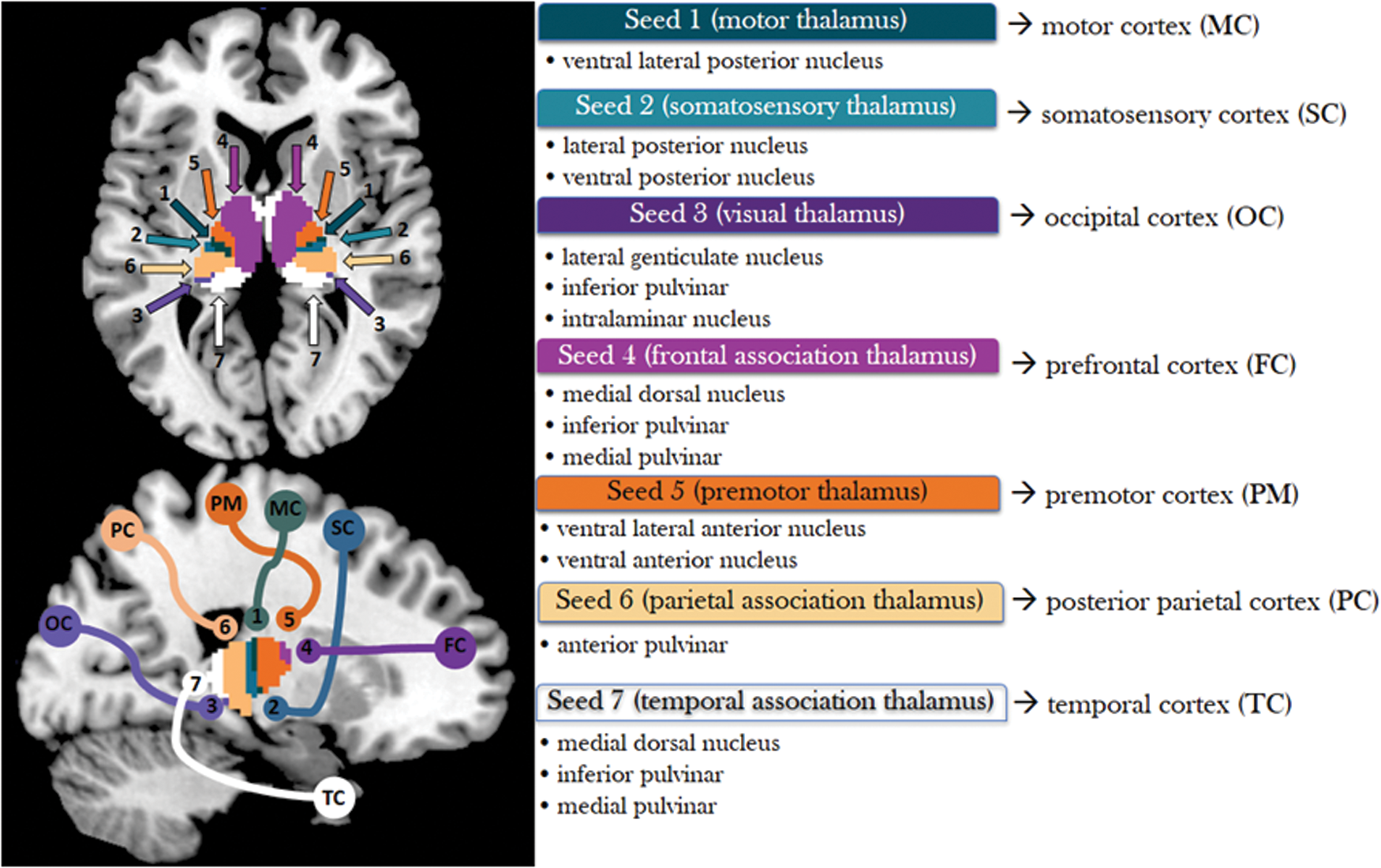
Seven thalamic seeds shown in an axial (z = 5) and sagittal (x = 20) slice with cortical projections (Behrens et al., 2003). Color images available online at
We named each seed region according to its major cortical projections. Seed 1 (motor thalamus) includes the ventral lateral posterior nucleus, which projects to the primary motor cortex. Seed 2 (somatosensory thalamus) includes the lateral posterior and ventral posterior nuclei, which project to the somatosensory cortex. Seed 3 (visual thalamus) includes the lateral geniculate nucleus, parts of the inferior pulvinar, and intralaminar nuclei. Seed 4 (frontal association thalamus) includes part of medial dorsal nuclei, ventral anterior nuclei, and parts of the anterior complex (anterior medial and anterior dorsal nuclei) that project to the prefrontal cortex. Seed 5 (premotor thalamus) includes the ventral lateral anterior and ventral anterior nuclei, which project to premotor areas. Seed 6 (parietal association thalamus) includes anterior parts of the pulvinar projecting to the posterior parietal cortex. Seed 7 (temporal association thalamus) includes some of medial dorsal nuclei, receiving inputs from the temporal lobe, and parts of the anterior complex projecting to limbic areas in the medial temporal region.
Statistical analyses of connectivity
In analysis of rsFC of each thalamic seed, we performed whole-brain multiple regression, each against AUDIT and AE score, with age as a covariate for men and women combined and separately. We did not include both AUDIT and AE scores in the same regression model because the two variables were highly correlated (see the Results section). All results were examined at peak voxel p < 0.001 uncorrected in combination with cluster p < 0.05 family-wise error corrected, according to current reporting standards. Clusters that met this statistical threshold were identified as regions of interest (ROIs), and the effect size (z value) of rsFC was extracted for each of the ROIs for individual subjects. A slope test was performed to examine whether men and women were significantly different in the correlation between the effect size of thalamic rsFC and regressor of interest (Zar, 1999). To anticipate, a total of 13 and 12 ROIs were identified each from the regression against AUDIT and AE score, and a p < 0.0038 (0.05/13) and <0.0042 (0.05/12) was each used to examine the statistical significance of slope test.
Mediation analysis
The right caudate nucleus showed frontal association and premotor thalamic connectivity in correlation with both AUDIT and AE scores (see the Results section). We used mediation analyses to examine whether AE mediated the correlation between thalamic caudate connectivity and problem drinking, as indexed by AUDIT score, or whether AE led to problem drinking and, in turn, resulted in changes in thalamic caudate connectivity, both with age as a covariate. We performed mediation analyses (MacKinnon et al., 2007), using the toolbox M3, developed by Tor Wager and Martin A. Lindquist (
In a mediation analysis, the relationship between the independent variable X and dependent variable Y, that is, X→Y, is tested to see if it is significantly mediated by a variable M. The mediation test is performed by using three regression equations (MacKinnon et al., 2007):
where a represents X→M, b represents M→Y (controlling for X), c′ represents X→Y (controlling for M), and c represents X→Y. The constants i1, i2, i3 are the intercepts, and e1, e2, e3 are the residual errors. In the literature, a, b, c, and c′ were referred as path coefficients or simply paths (MacKinnon et al., 2007; Wager et al., 2008), and we followed this notation. Variable M is said to be a mediator of the correlation X→Y if (c–c′), which is mathematically equivalent to the product of the paths a × b, is significantly different from zero (MacKinnon et al., 2007). If the product a × b and the paths a and b are significant, one concludes that X→Y is mediated by M. In addition, if path c′ is not significant, there is no direct connection from X to Y and that X→Y is completely mediated by M. Note that path b is the relationship between Y and M, controlling for X, and should not be confused with the correlation coefficient between Y and M.
Results
Demographic and clinical variables
Sociodemographic variables are shown in Table 1. Compared to women, men showed a significantly higher AUDIT score and average monthly frequency of drinking. AE was highly correlated with AUDIT score in men and women combined (r = 0.568, p = 1.82e-10), as well as in men (r = 0.538, p = 5.67e-05) and in women (r = 0.623, p = 2.29e-07) separately.
Thalamic rsFC and problem drinking
Supplementary Figure S1 shows the whole-brain rsFC of each thalamic seed (Supplementary Data are available online at
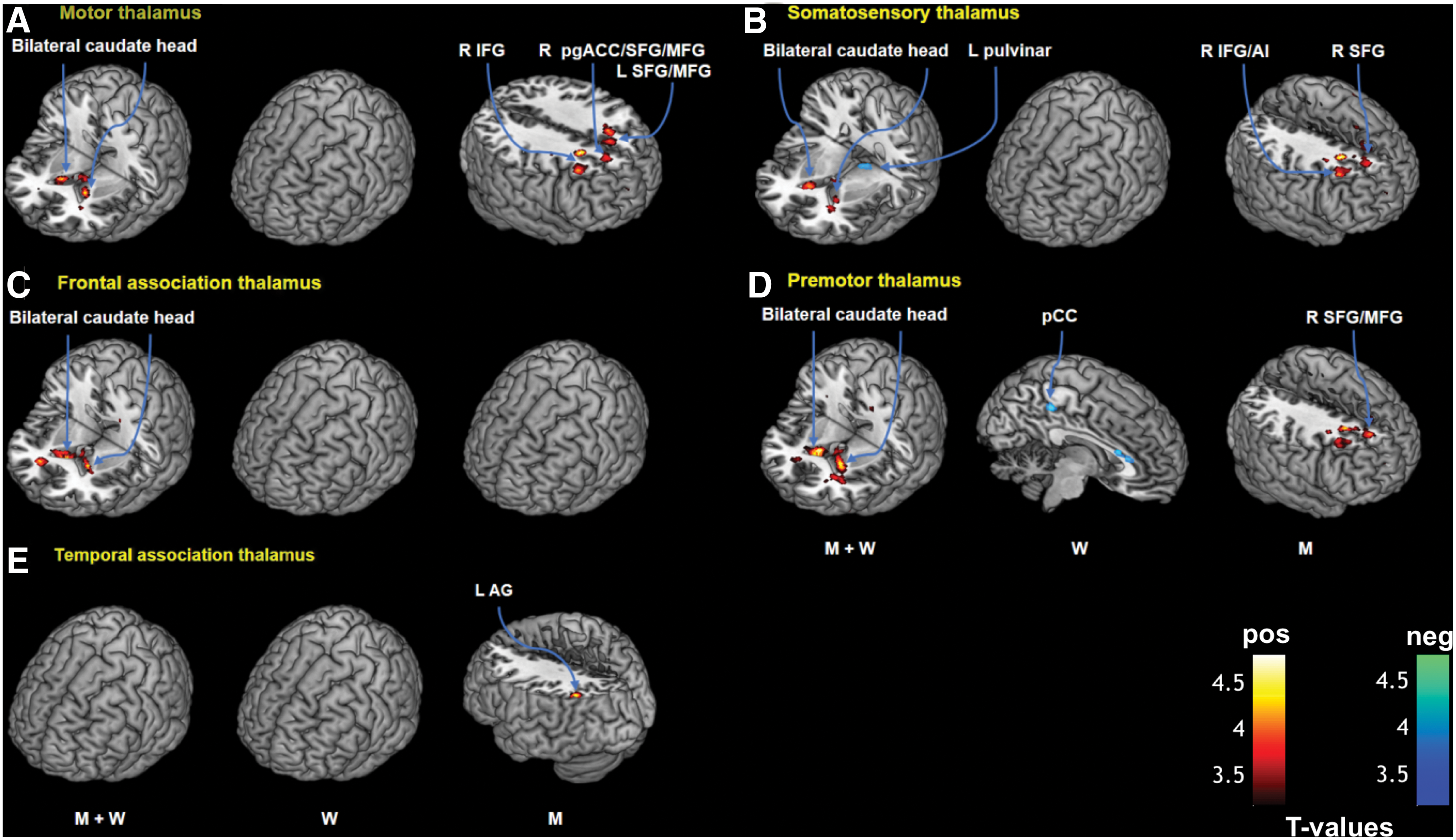
Three-dimensional rendering of clusters showing significant AUDIT-related connectivity with a thalamic subregion
Brain Regions Showing Thalamic Connectivity in Correlation with Alcohol Use Disorders Identification Test Score
All peak voxels 8 mm apart are identified. Only seed regions with significant findings are shown.
—, no clusters showing significant correlation; AI, anterior insula; IFG, inferior frontal gyrus; L, left; MFG, middle frontal gyrus; MNI, Montreal Neurological Institute; PCC, posterior cingulate cortex; pgACC, pregenual anterior cingulate cortex; R, right; SFG, superior frontal gyrus.
Seed 1: motor thalamus
Multiple regression in men and women combined showed positive correlation with AUDIT score in motor thalamic connectivity with bilateral caudate head. In men only, motor thalamus showed connectivity with a cluster encompassing the right perigenual anterior cingulate cortex, superior frontal and middle frontal gyri (pgACC/SFG/MFG), left SFG, and right inferior frontal gyrus (IFG) in positive correlation with AUDIT score (Fig. 2A). In a slope test, the connectivity with the right pgACC/SFG/MFG and right IFG was significantly different between men and women (Fig. 3A, B). There were no significant correlations between AUDIT score and motor thalamus rsFC in women alone at the same threshold.
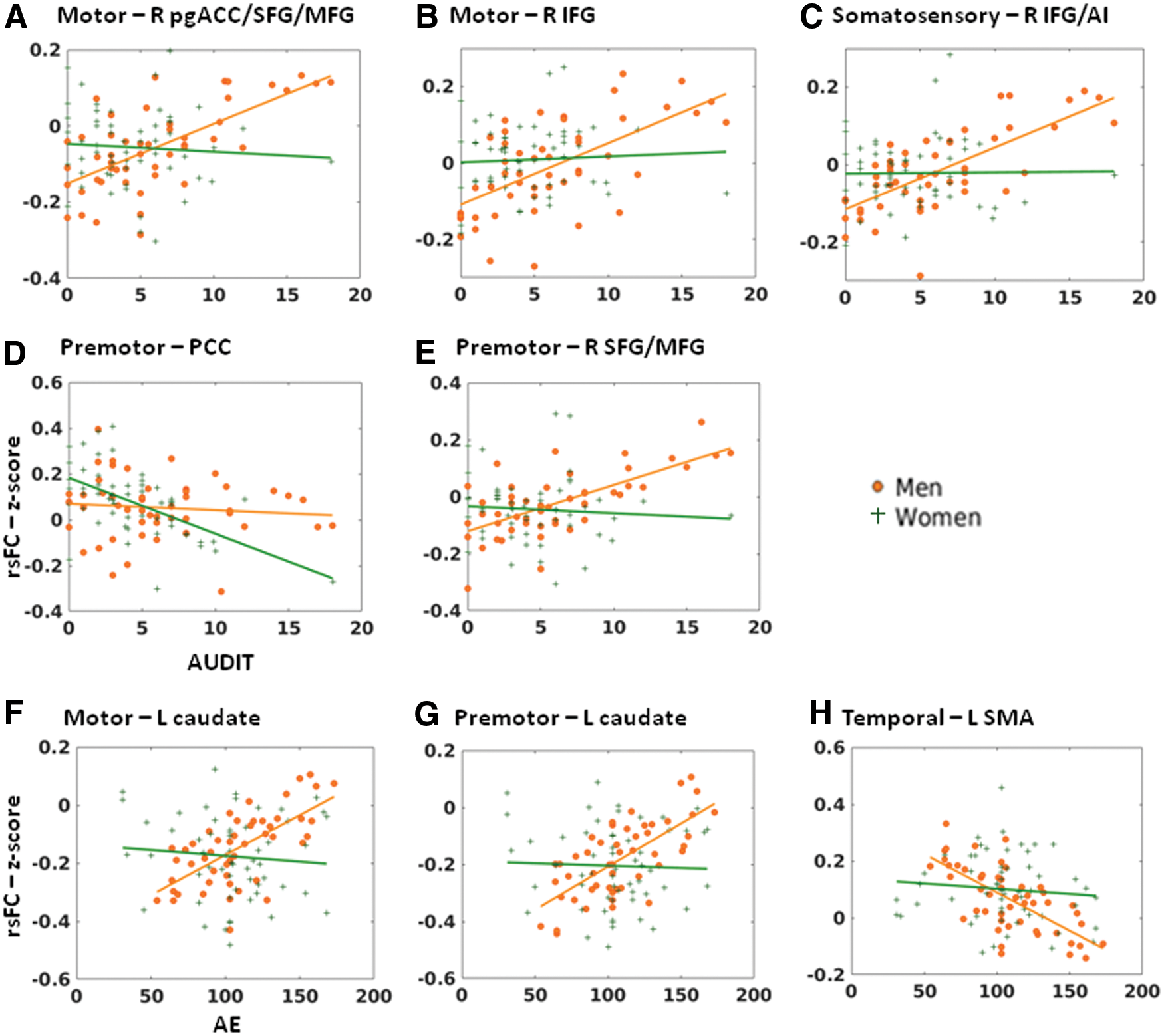
Sex differences in thalamic connectivity in relation to AUDIT
Seed 2: somatosensory thalamus
In men and women together, AUDIT score was positively correlated with somatosensory thalamic connectivity with bilateral caudate head and negatively with the left pulvinar (Fig. 2B). In men, AUDIT score was positively correlated with somatosensory thalamic connectivity with the right IFG/anterior insula (IFG/AI) and right SFG in positive correlation with AUDIT score. The correlation of right IFG/AI connectivity with AUDIT was significantly different between men and women; Fig. 3C). There were no significant correlations between AUDIT and somatosensory thalamic rsFC among women.
Seed 3: visual thalamus
There were no significant findings for visual thalamus connectivity in men only, women only, or in men and women combined when regressed against AUDIT.
Seed 4: frontal association thalamus
In men and women combined, there was a significant positive correlation between AUDIT score and frontal association thalamic connectivity with bilateral caudate head (Fig. 2C). There were no significant findings for men or women examined separately.
Seed 5: premotor thalamus
AUDIT score was positively correlated with premotor thalamic connectivity with bilateral caudate head in men and women combined (Fig. 2D). In women and men examined separately, AUDIT score was negatively correlated with premotor thalamic connectivity with the posterior cingulate cortex (PCC) in women and positively correlated with the right SFG/MFG in men, respectively. The latter two correlations showed a significant sex difference in a slope test (Fig. 3D, E).
Seed 6: parietal association thalamus
There were no significant findings in men only, women only, or in men and women combined.
Seed 7: temporal association thalamus
Temporal association thalamic connectivity with the angular gyrus (AG) was positively correlated with AUDIT score among men; however, this correlation was not significantly different between men and women in a slope test. There were no significant findings when considering women alone or men and women combined.
Thalamic rsFC and AE
The results of linear regression against AE score are shown in Figure 4 and summarized in Table 3.
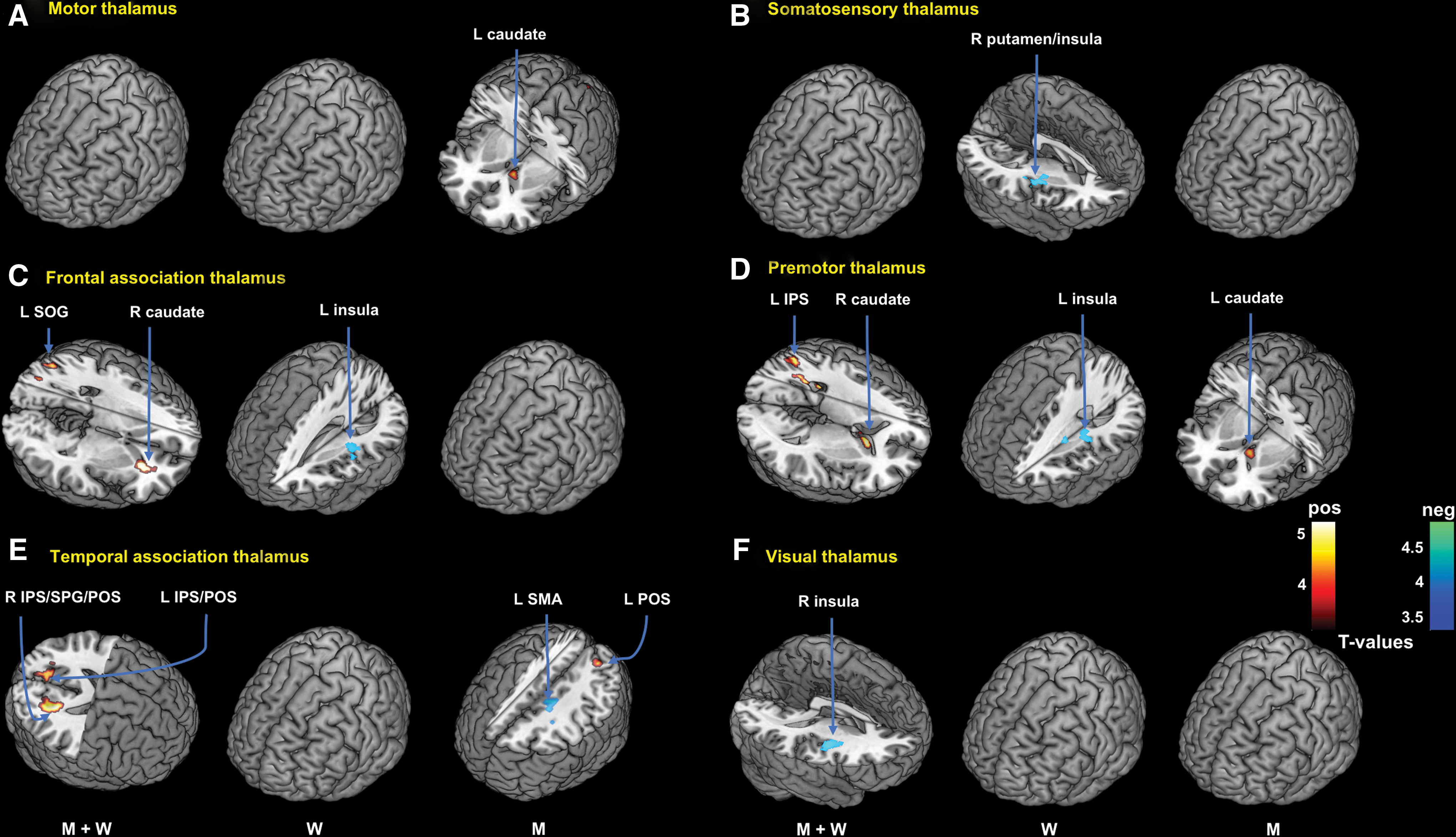
Brain Regions Showing Thalamic Connectivity in Correlation with Alcohol Expectancy Score
All peak voxels 8 mm apart are identified. Only seed regions with significant findings are shown (voxel p < 0.001, uncorrected, in combination with cluster p < 0.05, FWE corrected).
IPS, intraparietal sulcus; POS, parieto-occipital sulcus; SMA, supplementary motor area; SOG, superior occipital gyrus; SPG, superior parietal gyrus; FWE, family-wise error.
Seed 1: motor thalamus
Multiple regression in men showed positive correlation with AE score in motor thalamic connectivity with left caudate head (Fig. 4A). In women only or men and women combined, there were no significant correlations between AE score and motor thalamus rsFC at the same threshold. Men and women differed significantly in motor thalamic connectivity with the left caudate in a slope test (Fig. 3F).
Seed 2: somatosensory thalamus
In women, AE score was negatively correlated with somatosensory thalamic connectivity with right putamen and insula (Fig. 4B). At the same threshold, there were no significant correlations between AE score and somatosensory thalamic rsFC in men or in men and women combined.
Seed 3: visual thalamus
In men and women combined, AE score was negatively correlated with visual thalamic connectivity with the right insula and putamen (Fig. 4F). In men or women examined separately, no clusters showed significant visual thalamic rsFC in correlation with AE score.
Seed 4: frontal association thalamus
In men and women combined, AE score was positively correlated with frontal association thalamic connectivity with the right caudate and left superior occipital gyrus. In women only, AE score was negatively correlated with connectivity to the left insula (Fig. 4C). There were no significant correlations in men alone at the same threshold.
Seed 5: premotor thalamus
In men and women combined, AE score was positively correlated with premotor thalamic connectivity with the right caudate head and left intraparietal sulcus (IPS). AE score was positively correlated with premotor thalamic connectivity with the left caudate in men alone and negatively correlated with the left insula in women alone (Fig. 4D). Men and women differed significantly in premotor thalamic connectivity with the left caudate (Fig. 3G).
Seed 6: parietal association thalamus
There were no significant findings in men only, women only, or in men and women combined.
Seed 7: temporal association thalamus
In men, AE score was negatively correlated with temporal association thalamic connectivity with the left supplementary motor area (SMA) and positively correlated with the left parieto-occipital sulcus (POS) (Fig. 4E). In men and women combined, AE score was positively correlated with the right IPS/superior parietal lobule/POS and left IPS/POS. There were no significant findings in women examined separately. Men and women differed significantly in temporal association thalamic connectivity with the left SMA (Fig. 3H).
Mediation analyses
Frontal association and premotor thalamus showed higher connectivity with the right caudate head in correlation with both AUDIT and AE scores. We thus computed the averaged effect size of thalamic caudate connectivity and along with AUDIT and AE scores conducted mediation analyses. Specifically, we distinguished two hypotheses: (1) whether the relationship between thalamic caudate connectivity and problem drinking was mediated by AE and (2) whether AE contributed to problem drinking and in turn resulted in altered thalamic caudate connectivity. We conducted the mediation analyses for men and women combined and for men and women separately, and considered a corrected p-value of 0.05/6 = 0.0083 in examining the results (Fig. 5). In men and women combined and, nearly significantly, in women alone, AE score completely mediated the correlation between connectivity effect size and AUDIT score (Fig. 5D, E). In contrast, the model where AUDIT score mediated the correlation between AE score and connectivity effect size was not supported.
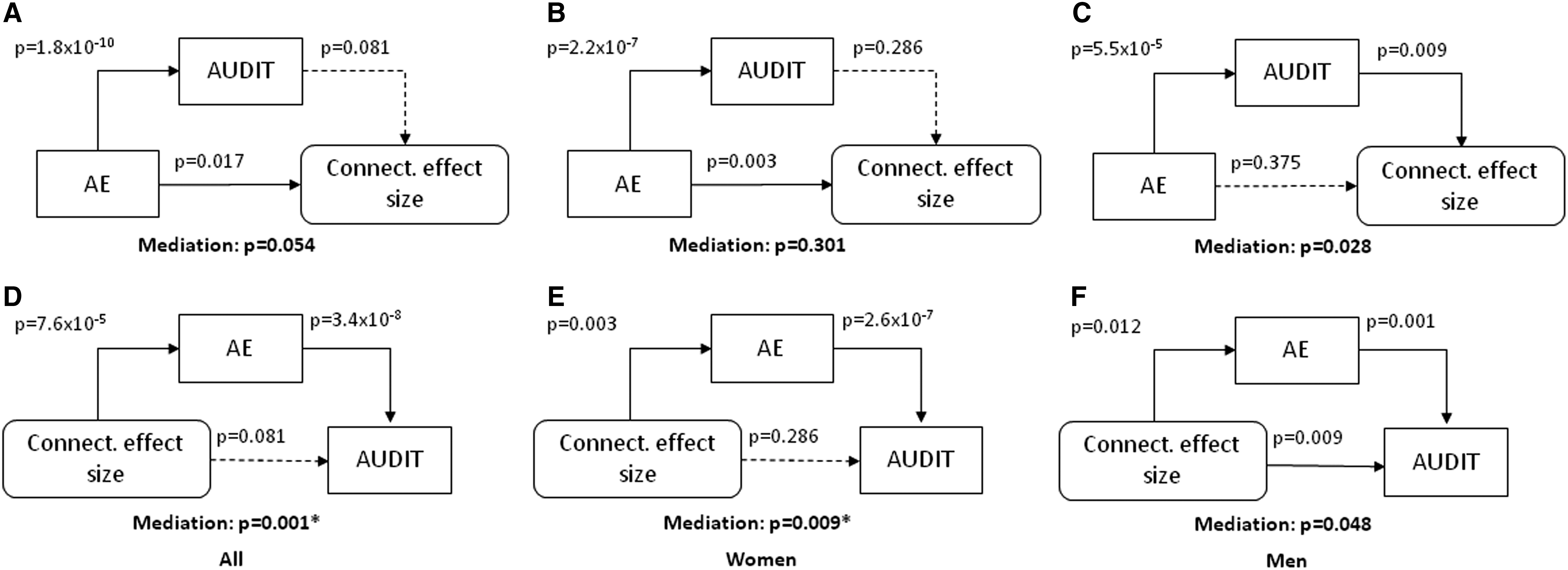
Analyses to test two models of mediation:
Discussion
The current findings characterized correlations of thalamic rsFC with problem drinking and AE. Although AUDIT and AE scores were highly correlated, the two variables were associated with different patterns of thalamic connectivities. Overall, problem drinking was associated with increased thalamic cortical and decreased “within-thalamus” connectivity. The findings appeared to be consistent with a recent report of alcohol administration significantly increasing functional connectivity density of the thalamus in heavy drinkers (Shokri-Kojori et al., 2017). These findings were also broadly consistent with an earlier work of expanded network connectivity as a neural mechanism for functional compensation in heavy drinkers (Müller-Oehring et al., 2015). In contrast, AE was associated with a mixed pattern of increased and decreased thalamic cortical connectivities. The caudate head showed increased thalamic connectivity in association with both AUDIT and AE scores. We computed the connectivity effect size and showed in mediation analyses that thalamic caudate connectivity contributes to problem drinking via enhanced AE. We highlight and discuss the main findings below.
Problem drinking
Motor, premotor, somatosensory, and frontal association thalamic connectivity with bilateral caudate head was positively correlated with AUDIT score. The head, body, and tail of the caudate nucleus partake in functionally distinct corticostriatal loops, with caudate head connected with prefrontal cortex to support cognitive functions, including impulse control (Zhang et al., 2012). Alcohol cue-evoked activations of the caudate head were positively correlated with negative urgency (the tendency to act rashly when experiencing negative emotions) in nondependent drinkers (Chester et al., 2016). Cue-related caudate activities have been related to the transition from moderate to heavy drinking in college students (Dager et al., 2014). In addition to impulse control, the caudate head supports procedural learning—the memory processes involved in encoding, storing, and retrieving information and automating actions (Hubert et al., 2007). Alcohol-addicted individuals showed impairment in procedural learning, in association with lower gray matter volume in the caudate head (Ritz et al., 2014).
A few studies reported specifically on thalamic caudate connectivity. In a resting-state fMRI study, participants with Alzheimer's disease showed greater caudate thalamic connectivity compared with controls (Kenny, 2013). The current findings of increased thalamic connectivity with bilateral caudate in link with higher AUDIT score appear to accord with the latter study, as problem drinking is associated with increased risk of dementia (Venkataraman, 2017). Another work demonstrated decreased caudate thalamic connectivity in ecstasy (3,4-methylenedioxymethamphetamine [MDMA]) abusers (Salomon, 2012). Thus, the excitatory and inhibitory effects, each of MDMA and alcohol on NMDA signaling, seem reflected in the contrasting findings of thalamic connectivity in chronic ecstasy and alcohol users (Finnegan et al., 1989; Nagy, 2008). Also in support of the current findings was a recent work showing increased thalamic functional connectivity with a wide swath of brain regions, including the caudate nucleus, following d-lysergic acid diethylamide (LSD) administration in healthy adults (Müller et al., 2017). As with alcohol, LSD inhibits NMDA receptor-mediated synaptic responses (Arvanov et al., 1999). Together, the current findings add to a literature in support of thalamus and caudate dysfunction as a developing sign of problem drinking.
In men alone, AUDIT score was positively correlated with motor, somatosensory, and premotor thalamic connectivity with the right SFG/MFG as well as motor thalamic connectivity with the left SFG. The SFG/MFG supports working memory and inhibitory control (Dambacher et al., 2014; du Boisgueheneuc et al., 2006; Hu et al., 2016a). The left SFG was less activated among nondependent drinkers, compared with nondrinking controls, during nogo inhibitions in a go/nogo task (Hatchard et al., 2017). In alcohol-dependent patients, dependence severity was negatively associated with activation of the right SFG during impulsive, relative to delayed decisions in the delay-discounting task (Lim et al., 2017). Finally, light drinkers exhibited greater BOLD response than did heavy drinkers in the left SFG and bilateral MFG during nogo inhibitions (Ahmadi et al., 2013). A positive correlation between thalamic connectivity with the SFG/MFG and AUDIT score may reflect a compensatory process during cognitive control in male problem drinkers.
Also, in men, AUDIT score was positively correlated with motor and somatosensory thalamic connectivity with the right IFG, a structure critical to saliency detection, response inhibition, and postconflict response slowing (Cai et al., 2017; Duann et al., 2009; Ide and Li, 2011; Li et al., 2006). Individuals with alcohol misuse showed steeper discounting of delayed rewards and greater activation of the IFG during selection of delayed rewards (Claus et al., 2011). More broadly, impulsivity, as assessed by the Eysenck Impulsiveness Scale, was positively correlated with gray matter volume of the IFG in young nondependent drinkers (Charpentier et al., 2016). Alcohol-dependent men showed greater fear-related responses in bilateral IFG and women showed the opposite (Padula et al., 2015). Motor and somatosensory thalamic connectivity with the IFG in relation to AUDIT score may underlie the role that these regions play in mediating impulse and affective control in male drinkers.
In women alone, AUDIT score was negatively correlated with premotor thalamic connectivity with the PCC. The PCC receives inputs from the thalamus as well as limbic and neocortical structures, and projects to multiple motor and premotor cortical areas (Child and Benarroch, 2013) to support emotional motor actions (Child and Benarroch, 2013; Pitel et al., 2015). Decreased PCC volume predicted relapse to alcohol use in dependent drinkers (Rando et al., 2011). A recent study reported blunted connectivity distribution—an index of connectivity with all other brain voxels—in the PCC during exposure to alcohol and stress compared to neutral cues in alcohol-dependent individuals (Zakiniaeiz et al., 2016). Furthermore, thalamic PCC connectivity was reduced in depression (Kong et al., 2018), supporting a link of problem drinking to depression in women (Hallfors et al., 2005; Harrell and Karim, 2008; Petit et al., 2017). More studies are warranted to examine sex differences in the role of PCC and PCC thalamic connectivity in affective psychomotor manifestations and problem drinking.
Thalamic connectivity, AE, and problem drinking
Alcohol use is enhanced by prior positive experiences with alcohol and the expectation of future positive effects with alcohol consumption (Brown et al., 1985). In this study, premotor and temporal association thalamic connectivity with the parietal cortex was positively correlated with AE in men and women combined. Furthermore, frontal association, premotor, and visual thalamic connectivity with the insula was negatively correlated with AE. The parietal cortex and insula have been implicated in AE in a balanced placebo study of working memory (Gundersen, 2008). Specifically, before the MR scan, half of the participants consumed a soft drink (half correctly informed and half incorrectly informed that they consumed an alcoholic beverage), and the other half consumed an alcoholic beverage (half correctly informed and half incorrectly informed that they consumed a soft drink). The authors showed that alcohol intoxication decreased, while expectancy of being intoxicated increased, brain activations. Moreover, compared with the control group receiving a soft drink and correctly informed, the alcohol-placebo group receiving a soft drink and incorrectly informed that it was alcohol demonstrated greater activation in bilateral precuneus, parietal cortex, and insula. Along with the latter study, the current findings support a role of thalamic connectivity with the parietal cortex and insula in AE.
Frontal association and premotor thalamic connectivity with the caudate head were positively correlated with both AUDIT and AE score. We identified the effect size of thalamic caudate connectivity and in mediation analyses showed that thalamic caudate connectivity related to problem drinking via enhanced AE. The alternative model that AE led to problem drinking and, in turn, changes in thalamic caudate connectivity was not supported. This finding suggested a role of increased thalamic caudate connectivity as a risk factor rather than a consequence of problem drinking.
Thalamic caudate connectivity supports frontal striatal thalamic circuit function, which is frequently implicated in the etiological processes of drug addiction (Huang et al., 2018). Dysfunction of the cortico/striatal/thalamic circuits may contribute to unbridled saliency of external drug cues and interoception in association with anticipation of drug use (Peters et al., 2016). The thalamus contributed to the generation of optimistic self-referential beliefs (Kuzmanovic et al., 2016) and anticipation of reward (Plichta et al., 2013). Young adults who started drinking at an earlier age showed higher caudate response to reward in the monetary incentive delay task (Boecker-Schlier et al., 2017). In accord with thalamic caudate connectivity increasing with AE, thalamus and caudate head coactivated during anticipation of emotional arousal (Greenberg et al., 2015; Metzger et al., 2010). Along with these earlier studies, the current findings suggest that resting-state thalamic caudate connectivity may conduce to positive alcohol expectancy, which in turn contributes to problem drinking in social drinkers.
Sex differences
Men and women showed differences in thalamic connectivity with a number of cortical and subcortical structures, as confirmed by slope tests (Zar, 1999), in relation to problem drinking and AE. Notably, men but not women showed thalamic connectivities with structures in the saliency and executive control circuits, whereas women but not men showed thalamic connectivities with the PCC—part of the default mode network, in correlation with AUDIT score. Thus, problem drinking appeared to be associated with thalamic connectivity with functionally opponent neural circuits between men and women. In men but not women, the thalamus responded more strongly to painful (Berman et al., 2006) and emotional stimulation (Filkowski et al., 2017; Seo et al., 2017). Increased thalamic connectivity with the caudate nucleus and parietal cortex may reflect higher arousal in link with AE in men. Thalamic connectivity with the PCC was reduced in depression (Kong et al., 2018), suggesting that problem drinking may be more likely to be associated with depression in women than in men (Hallfors et al., 2005; Harrell and Karim, 2008; Petit et al., 2017).
Notably, prenatal alcohol exposures appeared to diminish thalamic cortical connectivity in rats (Rodriguez et al., 2016) and humans (Donald et al., 2015) imaged during resting states. We previously showed that decreased gray matter volume of the thalamus contributed to problem drinking in men but not in women (Ide et al., 2017). Thus, consistent with the findings of mediation analyses, increased thalamic connectivity more likely did not reflect consequences of alcohol consumption but, along with thalamic volume, represented a risk factor disposing men to problem alcohol use. The current findings add to the literature that suggests potential sex differences in thalamic dysfunction in alcohol misuse (Ide et al., 2018).
Limitations of the study and conclusions
The study is the first to examine the relationship between subregional thalamic connectivity and problem drinking in nondependent social drinkers. As the thalamus is critical to cognitive and affective functioning and vulnerable to the effects of chronic alcohol consumption, these findings may advance research of thalamic dysfunction as a potential etiological process of early habitual drinking. An important limitation concerns the lack of behavioral tests. Thus, any suggested links to cognitive or affective dysfunction should be considered speculative. Second, our conclusions on sex differences need to be considered in the context of higher AUDIT score and monthly drinking frequency in men than in women and a slightly narrower range of AUDIT score in women than in men. On the contrary, one could also argue that problem drinking tends to be more severe in men and the results may adequately reflect the drinking populations (Flannery et al., 2007). A related issue is that we examined sex differences but have not documented menstrual cycles in female participants. Third, AE as assessed by questionnaires may involve logistic and methodological issues and behavioral tasks to circumvent these difficulties and to capture the association between alcohol-related knowledge and behavior would facilitate this direction of research (Kuntsche and Kuntsche, 2017). Fourth, the AUDIT is a screening instrument and may not address all relevant aspects of alcohol use behavior in social drinkers. Furthermore, we did not thoroughly assess anxiety, depression, and other potential emotional dysfunctions that may relate to drinking behavior. Alcohol influences not only cognitive and affective but also physiological processes such as sleep/wake behavior, as supported by the intralaminar and other nonspecific thalamic nuclei (Pereira de Vasconcelos and Cassel, 2015). More work is needed to understand the many potential roles of the thalamus in these physiological functions in relation to problem drinking. Finally, although the findings of mediation analyses suggested thalamic caudate connectivity as a risk factor for problem drinking, longitudinal studies are needed to confirm this proposition. In particular, the current findings should be considered specific to social drinking. Although 25 of the 107 participants showed an AUDIT score ≥8, a score considered a positive screen for alcohol use disorders, the cohort averaged at 5.2. More studies are required to understand how thalamic dysfunction may manifest differently in nondependent and dependent drinkers.
To conclude, thalamic functional connectivities are altered with respect to specific subregions and sex in relation to problem drinking and AE. Overall, problem drinking is associated with increased thalamic connectivities with other cortical and subcortical structures, whereas AE is associated with a mixed pattern of increased and decreased connectivities. Thalamic caudate connectivity may underlie positive AE and contribute to problem drinking. These findings may help more studies in understanding the thalamic cortical and subcortical pathways that conduce to the transition from social to habitual drinking and sex differences in these transitional processes.
Footnotes
Acknowledgments
The study was supported by NIH grants AA021449 and P50AA12870, and the VA National Center for PTSD.
Author Disclosure Statement
JK has individual consultant agreements at less than $10,000 per year with AstraZeneca Pharmaceuticals, Biogen, Idec, MA, Biomedisyn Corporation, Bionomics, Limited (Australia), Boehringer Ingelheim International, Concert Pharmaceuticals, Inc., Heptares Therapeutics, Limited (UK), Janssen Research & Development, L.E.K. Consulting; Otsuka America Pharmaceutical, Inc., Spring Care, Inc., Sunovion Pharmaceuticals, Inc., Takeda Industries, and Taisho Pharmaceutical Co., Ltd. JK is on the scientific advisory board of Bioasis Technologies, Inc., Biohaven Pharmaceuticals; Blackthorn Therapeutics, Inc., Broad Institute of MIT and Harvard, Cadent Therapeutics, Lohocla Research Corporation, Pfizer Pharmaceuticals, and Stanley Center for Psychiatric Research at the Broad Institute. JK has stocks or stock options on ArRETT Neuroscience, Inc., Blackthorn Therapeutics, Inc., Biohaven Pharmaceuticals Medical Sciences, Spring Care, Inc., Biohaven Pharmaceuticals Medical Sciences. JK receives income greater than $10,000 as the editor of Biological Psychiatry. JK has the following patents or patent applications: Dopamine and noradrenergic reuptake inhibitors in treatment of schizophrenia (US Patent #:5,447,948); Glutamate Modulating Agents in the Treatment of Mental Disorders (US Patent No. 8,778,979); Intranasal Administration of Ketamine to Treat Depression (No. 14/197,767 filed on March 5, 2014); Methods for Treating Suicidal Ideation (No. 14/197.767 filed on March 5, 2014); Composition and methods to treat addiction (No. 61/973/961. April 2, 2014); Treatment Selection for Major Depressive Disorder (filed June 3, 2016, USPTO docket number Y0087.70116US00); Compounds, Compositions and Methods for Treating or Preventing Depression and Other Diseases (No. 62/444,552, filed on January10, 2017); Combination Therapy for Treating or Preventing Depression or Other Mood Diseases (No. 047162-7177P1 (00754) filed on August 20, 2018). JK receives nonfederal research support: AstraZeneca Pharmaceuticals provides the drug, Saracatinib, for research related to NIAAA grant “Center for Translational Neuroscience of Alcoholism; Pfizer Pharmaceuticals provides an investigational drug, PF-03463275, for research related to NIH grant “Translational Neuroscience Optimization of GlyT1 Inhibitor.” The other authors do not have competing financial interests to declare.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.