
Abstract
Posttraumatic stress disorder (PTSD) is associated with disrupted functional connectivity in multiple neural networks. Multinetwork models of PTSD hypothesize that aberrant regional connectivity emerges from broad network-level disruptions. However, few studies have tested how characteristics of network-level organization influence regional functional connectivity in PTSD. This gap in knowledge impacts both our understanding of the pathophysiology of the disorder and the development of network-targeted PTSD treatments. We acquired resting-state imaging from a naturalistic sample of patients with PTSD (n = 42) and healthy controls (n = 42). Group differences in functional connectivity were identified using region of interest analyses and estimations of within- and between neural network activity; PTSD patients demonstrated reduced amygdala–orbitofrontal connectivity and increased default mode network (DMN) connectivity compared with controls. We then used convergence—a novel measure representing the capacity for functional integration—to test whether differences in functional architecture underlie connectivity signatures of PTSD. This approach found that reduced frontoparietal network (FPN) convergence was associated with reduced amygdala–orbitofrontal connectivity. Furthermore, in controls only, increased DMN convergence was associated with reduced DMN-to-salience network connectivity, and increased FPN convergence was associated with reduced FPN-to-ventral attention network connectivity. These results suggest that FPN functional architecture may underlie insufficiencies in top–down control in PTSD, with results broadly supporting the notion that networks' functional architecture influences the breakdown of normative functional relationships in PTSD. This work also indicates the potential of convergence to be applied to clinical populations in future research studies.
Introduction
Posttraumatic stress disorder (PTSD) is an important public health problem associated with poor physical health and unemployment (Thomas et al., 2010). Lifetime prevalence of PTSD is 7% in the general U.S. population and afflicts 50–70% of military veterans (Hoge et al., 2004; Kessler et al., 2005). Given the limited effectiveness of current PTSD treatments (Watts et al., 2013), novel approaches to treatment are needed. Advancing our understanding of the underlying pathophysiology is critical for the generation of new biologically informed therapies.
Early neurocircuitry-based models of PTSD emphasized the key role of insufficient frontolimbic control in PTSD. Inadequate ventromedial prefrontal cortex “top–down” control over hyperactive fear processing was viewed as the disorder's central pathology (Milad and Quirk, 2012; Phelps et al., 2004; Rauch et al., 2006). While these models account for the threat sensitivity and intrusions characteristic of PTSD, they offer an incomplete explanation for the full range of clinical symptoms [e.g., dissociation and avoidance (Garfinkel and Liberzon, 2009)]. Nor do these early models offer an explanation for the observation of neuroimaging correlates of PTSD in other functional networks, including the frontoparietal network (FPN), salience network (SN), and default mode network (DMN) (Garfinkel and Liberzon, 2009; Hayes et al., 2012; Koch et al., 2016; Patel et al., 2012).
In response to neuroimaging evidence of broad aberrant neural activity, so-called triple-network models of PTSD have been adopted (Garfinkel and Liberzon, 2009; Hayes et al., 2012; Koch et al., 2016; Patel et al., 2012). DMN, SN, and FPN comprise the core networks and are implicated in self-referential thought, salience monitoring, and “top–down” cognitive control, respectively (Buckner et al., 2008; Seeley et al., 2007). These models propose that in PTSD, SN hyperactivity and enhanced threat monitoring disrupts DMN and FPN function. These disruptions give rise not only to maladaptive fear responses but also to broad deficits in memory, attention, and cognitive control that accompany PTSD (Hayes et al., 2012; Koch et al., 2016; Patel et al., 2012). Meta-analytic resting-state functional connectivity data are consistent with the triple-network model, suggesting that durable state-independent alterations in function underlie PTSD (Koch et al., 2016).
Although the accumulated neuroimaging literature underscores the centrality of SN, DMN, and FPN function to PTSD, it is unclear how the broader functional architecture of these networks relates to regional patterns of aberrant brain function in PTSD. Novel approaches like network convergence analysis may provide unique insight into these relationships. Convergence is a functional connectivity-based validated method for examining network-level functional architecture in the brain (Bell and Shine, 2015). In convergence analysis, voxel values represent the sum of strongly connected functional networks. Averaging these values within a network estimates the network's bias toward integrative versus segregated information processing, with higher convergence indicating greater potential for integration. Although convergence has been used to describe network architecture in healthy individuals, it has not been applied to clinical populations (Bell and Shine, 2015).
Here, we submitted resting-state data from a naturalistic sample of patients with PTSD to convergence analysis to evaluate whether differences in networks' functional architecture influenced PTSD-related patterns of regional functional connectivity. We focused our investigation on the PTSD triple-network, the ventral attention network (VAN), and affective network (AN). The VAN includes parts of the mid and posterior insula, lateral middle temporal gyrus (MTG), and midline paracentral regions (Yeo et al., 2011). VAN was included based on meta-analytic data linking posterior insula and lateral MTG to PTSD and anxiety disorders (Etkin and Wager, 2007; Tursich et al., 2015). AN was added to our investigated networks because prior functional connectivity studies have identified correlates of PTSD in the orbitofrontal cortex (OFC), an AN subregion (Jin et al., 2014; Philip et al., 2017; Sripada et al., 2012a,b), and because portions of VMPFC are in the AN (Yeo et al., 2011).
We hypothesized that the functional architecture of FPN in particular, would exert influence over the manifestation of regional PTSD correlates because our earlier study (Philip et al., 2017), found that responsiveness to TMS therapy targeting dorsolateral prefrontal cortex (part of FPN) for PTSD and depression was predicted by connectivity patterns among DMN and SN subregions. Previewing our results, we found that lower FPN convergence was associated with amygdala–hypoconnectivity in patients with PTSD. In addition, we found that although greater information flow through DMN and VAN was related to normative oppositional patterns of connectivity between internally and externally oriented networks, this relationship was absent in PTSD.
Materials and Methods
Participants
This is a secondary analysis of data from participants recruited from the Providence VA Medical Center and Butler Hospital, both affiliated with the Alpert Medical School of Brown University. Although results from some of these participants have been previously reported (Philip et al., 2017), analyses and results reported here are novel. Written informed consent was obtained from all participants. The institutional review boards at both hospitals approved all procedures. A total of 84 participants, 42 with current diagnosis of PTSD (mean age = 49.6 ± 11.3 years, female = 15) and 42 healthy individuals (mean age = 41.2 ± 14.3 years, female = 20), were recruited from the community or from ongoing studies at either institution. PTSD symptoms were measured with the PTSD checklist (PCL-5) (Weathers et al., 2013).
Additional details on participant inclusion/exclusion criteria and concurrent treatments appear in Supplementary Data (Supplementary Data are available online at
Magnetic resonance imaging data acquisition and preprocessing
Neuroimaging data were acquired at Brown University using either a Siemens 3T Trio or 3T Prisma MRI (Siemens, Erlangen, Germany) with a 32-channel head coil. Imaging acquisition included anatomical images followed by 8 min of standard resting-state echoplanar imaging. The CONN Toolbox (Whitfield-Gabrieli and Nieto-Castanon, 2012) was used to conduct standard magnetic resonance imaging (MRI) data preprocessing and functional connectivity preprocessing steps in volumetric space, and Freesurfer (
Overview of MRI data analyses
Resting-state functional MRI (fMRI) data collected from a naturalistic sample of patients with PTSD and from healthy controls were used to test (1) whether characteristics of functional architecture (convergence) account for PTSD symptom-related patterns of functional connectivity. Detailed definitions for region of interest (ROI), ROI-to-ROI analyses, and convergence statistics and analyses are supplied in subsequent Materials and Methods subsections.
A three-step process was used to address our hypotheses. First, we compared ROI-to-ROI functional connectivity of the amygdala, anterior hippocampus, and subgenual anterior cingulate (sgACC) with ROIs in the SN, VAN, DMN, AN, and FPN networks, between patients with PTSD and healthy controls. Hippocampus, amygdala, and sgACC were investigated given their prior association with responsiveness to therapeutic repetitive transcranial magnetic stimulation for PTSD (Philip et al., 2017). Second, we applied the convergence method to resting-state fMRI data to estimate functional connectivity density across the cortical surface. Third, to test whether networks' capacity for integrative processing influenced ROI-to-ROI correlates of PTSD, cortex-wise convergence statistics were averaged within functional networks of interest, and averages were entered into general linear models (GLMs) as predictors of patient versus control differences in functional connectivity. A similar approach was used to examine the relationships between within-network and between-network connectivity to convergence.
Definition of functional connectivity regions-of-interest
A set of functionally defined ROIs were used to measure connectivity in our networks of interest, the SN, VAN, DMN, AN, and FPN. Four 4-mm radius sphere ROIs were made for each network using the MarsBar Toolbox (
Region of Interest Functional Associations and Locations
ACC, anterior cingulate cortex; aHPC, anterior hippocampus; cACC, caudal anterior cingulate cortex; DLPFC, dorsolateral prefrontal cortex; MNI, Montreal Neurological Institute; mPFC, medial prefrontal cortex; OFC, orbitofrontal cortex; PCC, posterior cingulate cortex; PTSD, posttraumatic stress disorder; ROI, region of interest; sgACC, subgenual anterior cingulate; VMPFC, ventromedial prefrontal cortex.
ROI-to-ROI functional connectivity analyses
First, for each subject, we cross-correlated unsmoothed residual blood oxygen level-dependent (BOLD) signal time courses extracted from each seed in the ROI-to-ROI matrix. Correlations were converted to z scores using Fisher's r-to-z transformation. Z scores were entered into GLMs comparing ROI-to-ROI connectivity in patients versus controls. Models included age and data quality assessment (QA) statistics as covariates of no interest. Results from all ROI analyses were corrected for multiple comparisons using seed-level false discovery rate (FDR) correction (Hochberg and Benjamini, 1990). Subject-level QA statistics automatically generated by CONN during preprocessing were equal to the average correlation between all voxels in the brain. QA was regressed at the second level in post hoc sensitivity tests to ensure that group differences were not driven by residual effects of motion (Ciric et al., 2017; Saad et al., 2013). Results from all ROI analyses were corrected for multiple comparisons using seed-level false discovery rate correction (Hochberg and Benjamini, 1990). Additional post hoc tests were conducted to determine whether gender or scanner type influenced our results. Nonsignificant post hoc tests are not reported. Additional post hoc correlations examining the relationship between connectivity and symptom severity are summarized in Supplementary Data.
Within- and between-network functional connectivity analyses
Within-network and between-network connectivity values were generated by averaging connectivity Z-scores between all unique ROI combinations for each subject. Individuals' betas describing network connectivity were either contrasted between groups, or treated as dependent variables in subsequent GLM hypothesis testing.
Individual-level definition of resting-state functional connectivity networks
In preparation for convergence analysis, the 17 functional networks defined in Yeo and colleagues (2011) were projected onto the Freesurfer “fsaverage” surface (Fischl, 2012). Our use of atlas-based networks differed from methods described by Bell and Shine (2015), who used independent components analysis (ICA) to define networks in healthy controls. Given that our sample included both psychiatrically ill patients and healthy controls, we were concerned that ICA might yield components that are not representative of networks in either group. Although we could have defined ICA networks separately within each group, matching networks across groups may have introduced other inconsistencies. For these reasons, we substituted atlas-based definitions for ICA networks.
To create subject-level functional maps, we generated a reference signal for each network by averaging the BOLD time courses across vertices within the network's spatial boundaries. Then, to quantify associations between individual vertices and each network, we computed correlations between time courses from each vertex and each reference network signal, which yielded a Network (N = 17) × Vertex (V = 327,684) correlation matrix. Networks(s) were assigned to vertices if connectivity exceeded a threshold set at the 90th percentile (for details of empirical threshold determination, see Supplementary Data). A binary map for each network was then constructed by assigning a value of 1 to all vertices where network correlations exceeded threshold. Vertices could be associated with multiple networks.
Estimation of network convergence
Network binary maps were used to generate a network convergence metric (NCM) for each vertex on the cortical surface. The NCM at cortical vertex, v, was
where N is the sum of binary network maps at vertex v where connectivity surpassed threshold. Within-network convergence for each subject was then estimated as the average NCM within our networks of interest (DMN, SN, FPN, AN, and VAN). Subject-level convergence statistics were used for GLM hypothesis testing.
Testing the relationship between convergence and functional connectivity signatures of PTSD
The following analyses ask whether characteristics of functional architecture, that is, network-level capacity for integration, were associated with PTSD. We entered subject-level convergence statistics into GLMs as predictors of functional connectivity betas describing patients versus control differences in ROI-to-ROI functional connectivity. We did not include age as a covariate in these models since betas from the ROI analyses describe the contribution of group to connectivity after covariance. Additional post hoc sensitivity tests were conducted, however, to ensure that results were not driven by residual effects age, or QA, scanner, or gender. Marginal post hoc correlations examining the relationship between convergence and symptom severity are summarized in Supplementary Data.
Results
ROI-to-ROI functional connectivity in PTSD versus healthy individuals
Right amygdala-to-left OFC connectivity was weaker in individuals with PTSD (T(81) = −3.39, p < 0.005, p-FDR < 0.05). Follow-up within-group tests found evidence of significant positive connectivity between healthy controls (T(41) = 4.87, p < 0.001, p-FDR < 0.001), but not in patients (p-FDR > 0.1; Fig. 1A).
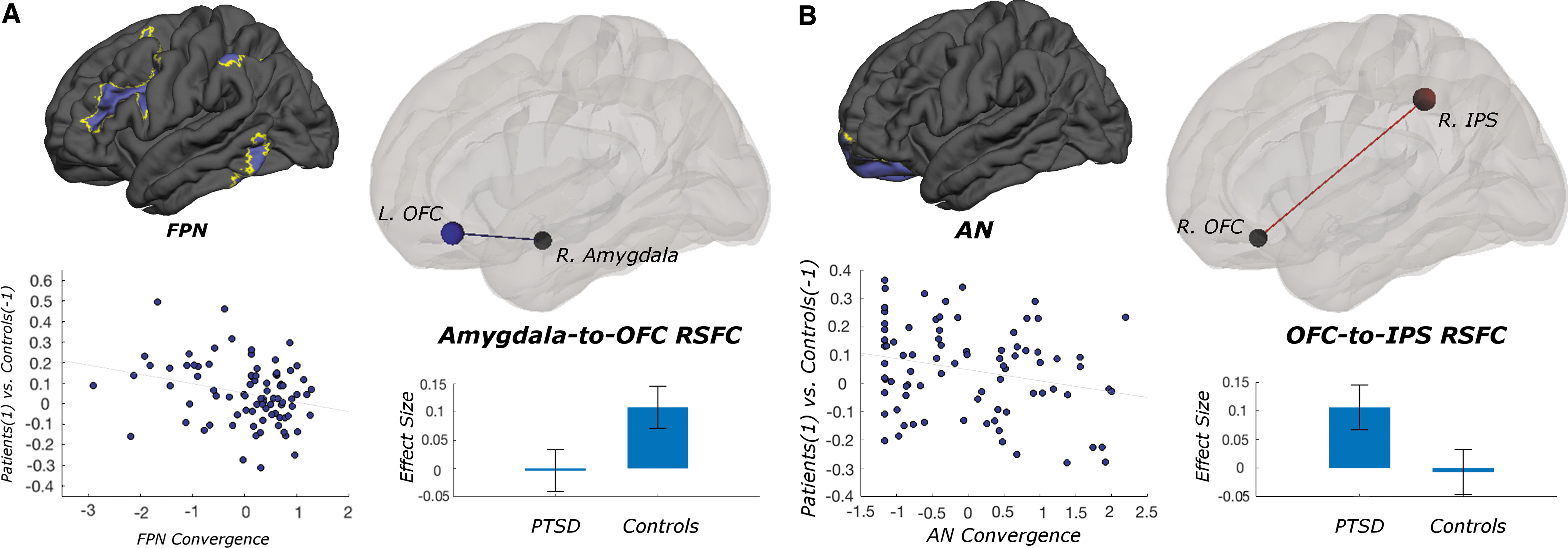
The functional architecture of FPN and AN influences ROI-to-ROI correlates of PTSD.
Functional connectivity between right OFC and right intraparietal sulcus (IPS) was stronger in patients than in controls (T(81) = 3.35, p < 0.005, p-FDR < 0.05; Fig. 1B). Although ROI time series were positively correlated in patients (T(41) = 4.5, p < 0.001, p-FDR < 0.001), ROI time series were uncorrelated in healthy controls.
Within- and between-network functional connectivity signatures of PTSD
Within-network connectivity between regions of the DMN was stronger in individuals with PTSD (T(82) = 1.73, p-FDR < 0.05) than in healthy controls. All other within-network comparisons were not significant (all p-FDR > 0.1).
Convergence and group differences in ROI-to-ROI functional connectivity
FPN convergence was negatively correlated with group differences in right amygdala-to-left OFC functional connectivity (T(1,81) = −2.53, p < 0.05) (Fig. 1A). In other words, disrupted connectivity in PTSD was associated with restricted FPN convergence. There was a trending negative correlation between AN convergence and group differences in right OFC-to-right IPS functional connectivity (T(1,81) = −1.97, p = 0.052) (Fig. 1B).
Convergence and group differences in between-network functional connectivity
DMN convergence differentially influenced DMN-to-SN connectivity by group (T(80) = 2.62, p = 0.01) (Fig. 2A). Specifically, increased DMN convergence was associated with reduced between-network connectivity in healthy controls only (T(41) = −3.50, p = 0.01). Despite this differential effect of convergence, group differences in DMN-to-SN cross-network connectivity were not observed (p > 0.1).
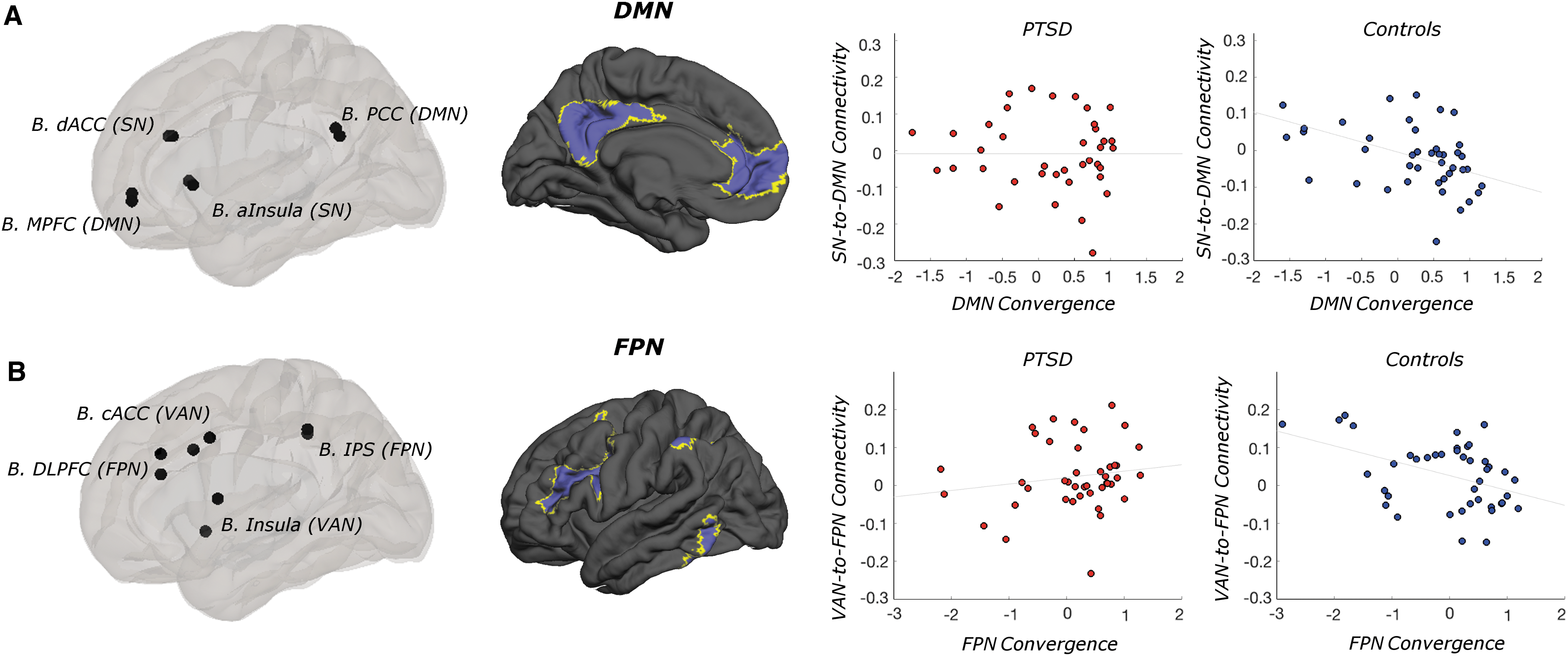
The functional architecture of DMN and FPN influences the between-network connectivity of internally and externally oriented functional networks.
Another statistically significant interaction indicated that FPN convergence differentially influenced connectivity between the FPN and VAN in patients versus controls (T(80) = 2.65, p < 0.01) (Fig. 2B). Greater FPN convergence was associated with reduced FPN-to-VAN connectivity in controls only (T(40) = −2.98, p < 0.005). Across subjects, VAN convergence was negatively correlated with DMN-to-VAN connectivity (T(80) = −2.15, p < 0.05).
Convergence and group differences in within-network functional connectivity
Within-network convergence did not influence group differences in within-network connectivity. However, in patients, within-network connectivity was significantly stronger in DMN (T(82) = 1.73, p < 0.05), and nominally stronger in FPN (T(82) = 1.35, p = 0.09).
Discussion
Our application of network convergence analysis provides new potentially important insights about the neural basis of PTSD. Like degree and global connectivity-based graph measures (Rubinov and Sporns, 2010), the NCM describes a network's potential to integrate and distribute information to other brain areas (Bell and Shine, 2015). In this study, we treated these convergence metrics as predictors of ROI-to-ROI functional connectivity in an effort to evaluate the impact of the functional architecture of individual networks on aberrant patterns of regional connectivity in PTSD. Critically, the results of our analyses indicate that lower convergence—restricted capacity for functional integration—in the FPN and DMN influences two broad patterns of dysregulated functional connectivity associated with PTSD: disrupted connectivity in limbic regions and a breakdown in normative patterns of oppositional functional connectivity between networks (e.g., SN and DMN). Our finding that broad characteristics of network architecture influence the expression of PTSD-related patterns of functional connectivity lends additional empirical support for the growing conceptualization of PTSD as a multisystem disorder associated with widespread differences in brain function (Garfinkel and Liberzon, 2009; Hayes et al., 2012; Kennis et al., 2016; Koch et al., 2016; Patel et al., 2012).
Functional architecture and limbic correlates of PTSD
Our observation of amygdala-to-OFC hypoconnectivity is consistent with previous reports of aberrant prefrontal-to-limbic functional connectivity in individuals with PTSD (Jin et al., 2014; Philip et al., 2017; Sripada et al., 2012a,b). Our results indicate that this hypoconnectivity is associated with reduced capacity for functional integration in FPN. This finding complements the long-standing hypothesis that insufficient prefrontal top–down modulatory control over fear or threat vigilance is a primary source of inappropriate fear expression (Milad and Quirk, 2012; Rauch et al., 2006). This work also lends empiric support to the use of interventions targeting this circuit, such as transcranial direct current stimulation delivered to the frontal cortex to improve fear extinction in patients with PTSD (van 't Wout et al., 2017; van 't Wout-Frank et al., 2018).
Our convergence analyses also revealed that significant ROI-to-ROI functional connectivity between right OFC and IPS in patients with PTSD was associated with greater functional segregation of the AN. Although resting-state methods do not provide the means to test this empirically, we speculate that comparatively elevated functional connectivity between OFC and IPS, a critical node of the FPN, may reflect tonic increases in the demand for compensatory cognitive control in response to symptoms of the disorder. Greater functional segregation of the AN as observed in this study may limit the impact of these compensatory signals on the regulation of maladaptive fear responses in PTSD. Segregation of the AN may also contribute to ventral prefrontal cortex hypoactivation during negative emotional processing, a robust neuroimaging correlate of PTSD (Etkin and Wager, 2007).
Functional architecture and dynamically oppositional networks in PTSD
The brain is organized into multiple distinct functional connectivity networks (Dosenbach et al., 2007; Greicius et al., 2003; Seeley et al., 2007; Vincent et al., 2008; Yeo et al., 2011), many of which exist in dynamic opposition [e.g., “task-positive” and “task-negative” networks (Fox et al., 2005), dorsal and ventral attention systems (Fox et al., 2006), SN and FPN vs. DMN (Sridharan et al., 2008)]. Notably, elevated functional connectivity between the SN and DMN has been previously reported in patients with PTSD (Sripada et al., 2012b). Although we did not find evidence of significantly elevated SN-to-DMN between-network connectivity in this study, convergence in the DMN did differentially impact the oppositional relationship between these two networks. Namely, we found that in healthy individuals only, so-called anticorrelations between the SN and DMN were stronger in those exhibiting greater DMN convergence.
Similarly, convergence within the FPN differentially impacted FPN-to-VAN functional connectivity in patients and controls. FPN is involved in the integration of information from the external environment with internal representations (Vincent et al., 2008). In this study, greater capacity of the FPN for functional integration was associated with stronger FPN-to-VAN connectivity in patients with PTSD, but greater anticorrelation in controls. Given that the VAN ROIs used in this study were associated with pain processing, increased functional coupling between these networks in patients may reflect the allocation of FPN resources toward nocioception or pain regulation. However, understanding the functional significance of this interaction in the context of PTSD symptomatology will require further examination using task-based study designs. Nonetheless, this result further underscores the rationale for interventions specifically targeting the FPN.
Together, these findings are consistent with previous reports of widespread disruptions in network topology (Kennis et al., 2016) and the characterization of both PTSD (Hayes et al., 2012; Koch et al., 2016; Patel et al., 2012) and psychopathology more generally (Menon, 2011), as multinetwork disorders. By demonstrating that PTSD-related differences in DMN and FPN network architecture impact the equilibrium between internally and externally directed functional networks, our findings support assertions that disequilibrium is a key aspect of PTSD pathophysiology (Sripada et al., 2012b). Whether this disruption in normative network functional architecture is a reflection of the pathology, or an indication of a failed compensatory response, remains unknown. Future studies, evaluating the impact of successful psychiatric treatment on network convergence, will be able to provide important insights into this question.
Limitations
Our convenience sample included individuals exposed to a mix of traumas (e.g., combat and sexual), but our sample size was insufficient to examine differential effects of trauma types. With few exceptions, psychiatrically ill participants were on medications. Although doses were stable for at least 6–8 weeks before scanning, the longer term effects of medications on scanning are unknown (Lanius et al., 2010). The lack of trauma-exposed healthy controls limits our ability to uniquely parse effects resulting from trauma exposure or PTSD. Variability in exposure to early life stress in both the PTSD and control groups also complicates the parsing of PTSD and trauma-exposure effects (see Supplementary Data for additional discussion). Our ROI selection was motivated by the fear/extinction literature in PTSD, although other networks (Kennis et al., 2016) and subnetworks (Miller et al., 2017) may be implicated. Although convergence has been validated against other graph measures (Martuzzi et al., 2011), the novelty of the method, and its use in a neuropsychiatric sample, limits direct comparisons with prior research.
Conclusion
We found that convergence provided a means to observe the loss of multinetwork relationships in PTSD compared with healthy controls. This work highlights how convergence methods can provide novel insights into large-scale network changes in PTSD, and suggests that this approach can provide unique insight into functional architecture in psychiatric illnesses more broadly. Future studies are needed to replicate and validate this work and evaluate whether convergence methods can provide neuroimaging predictors of treatment response or characterize how networks shift in response to psychiatric interventions.
Footnotes
Acknowledgments
We extend special thanks to all of the participants. We thank Causey Dunlap, BS; Sarah Albright, BA: and Eric Tirrell, BS, for their assistance with participant procedures. This study was supported by the U.S. Department of Veterans Affairs, Clinical Sciences Research and Development (IK2 CX000724 to N.S.P.); the Center for Neurorestoration and Neurotechnology at the Providence VA Medical Center; and an investigator-initiated grant from Neuronetics, Inc. to Butler Hospital (to N.S.P. and L.L.C.) that provided a portion of funds used for neuroimaging. The opinions herein represent those of the authors and not the U.S. Department of Veterans Affairs, or Neuronetics. Funders had no involvement in the collection, analysis, and interpretation of the data, or results reporting and dissemination.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.