
Abstract
Due to technological advances, spatially indexed objects, such as blood oxygen level-dependent time series or electroencephalography data, are commonly observed across different scientific disciplines. Such object data are typically high dimensional and therefore challenging to handle. We propose a new approach for spatially indexed object data by mapping their spatial locations to a targeted one-dimensional interval so objects that are similar are placed near each other on the new target space. The proposed alignment not only provides a visualization tool for such complex object data but also facilitates a new way to study brain functional connectivity. Specifically, we introduce a new concept of path length to quantify the functional connectivity and a new community detection method. The advantages of the proposed methods are illustrated by simulations and in a study of functional connectivity for Alzheimer's disease.
Introduction
Neuroimaging data, such as functional magnetic resonance imaging (fMRI) and electroencephalography data, often appear in the form of high-dimensional time series, where the time series are collected over a grid of spatial locations and the number of spatial location is large. These high-dimensional time series data can be handled as spatiotemporal data (Castruccio et al., 2016; Park et al., 2016; Shinkareva et al., 2006) with the time series representing the temporal component. A key example is fMRI, a neuroimaging technique that measures brain activity over a short period of time through the changes of blood oxygen levels. These blood oxygen level-dependent (BOLD) signals are repeated measurements on each voxel location of a regular grid of the three-dimensional (3D) brain. The result is a time series of BOLD signal at each voxel location. In this regard, fMRI, or synonymously, BOLD data can be viewed as four-dimensional (4D) spatiotemporal data with a time series object at each of the 3D spatial (voxel) locations. We regard each time series as an “object” and the 4D fMRI data as “spatially indexed object data.”
By leveraging the information on spatial adjacency and the rich literature for spatiotemporal data, one can tease out the information in these spatially indexed object data. Such an approach, however, is not effective to study functional connectivity (Friston et al., 1993), where the focus is on the strength of statistical associations of the BOLD objects among different brain regions, because regions that are anatomically far from each other could be highly connected.
Brain functional connectivity is a vast field (Friston, 2011; Preti et al., 2017; Van Den Heuvel and Pol, 2010). A popular approach is to develop brain networks based on a functional connectivity map. Such approaches often involve the choice of a threshold, which may affect subsequent analysis (Sporns, 2010). Moreover, it is not easy to develop statistical tests to detect group differences in the network structure. To tackle these challenges, we propose a new network approach that does not involve thresholding and provides a new approach to detect communities.
Our approach consists of two steps. In the first step, we employ multidimensional scaling (MDS) (Borg and Groenen, 2005) to align the BOLD objects along a one-dimensional (1D) interval, which then leads to a new brain map. In the next step, we propose a new network approach that is based on the new brain map constructed in the first step. A by-product of the approach is a novel way to visualize the 4D brain data of a subject by compressing the data to a two-dimensional (2D) image.
MDS is a popular approach for dimension reduction but has not been fully explored in the area of brain connectivity. It has been used for clustering or visualizing cortical regions (Horwitz, 2003; Welchew et al., 2005, 2002) as well as to map the brain anatomy to a functional connectivity 3D space (Friston et al., 1996). In the functional connectivity space, the proximity of regions of interest (ROIs) will be determined by the strength of their functional connectivity, not by their anatomic location, and the projection onto a new 3D space with functional connectivity as the metric will allow scientists to visually explore the underlying mechanism of cortical regions.
Inspired by this visualization advantage (Friston et al., 1996) and a previous success (Chen et al., 2011) where one rearranges high-dimensional scalar objects into functional data (Wang et al., 2016), we propose to project the BOLD time-course objects, which were originally spatially located on the 3D brain, onto a 1D space. This projection aligns all brain regions on an interval, for example, [0, 1], so that the BOLD object at each spatial location is assigned to a new horizontal location in [0, 1]. A 2D brain image is then formed for each subject by lining up the BOLD time-course objects vertically on their new locations. We design a new statistical method to rearrange those spatially indexed temporal data in such a way that “similar” objects (temporal data in this case) are placed near each other to facilitate data visualization and subsequent data analysis. Because the multivariate spatial locations are collapsed into 1D spatial locations, dimension reduction is thereby accomplished. While this may seem daring at first, the simulation and data analysis in the sections “Alignment of Time Series” and “Results” show the advantages of this approach.
The 1D spatial locations then facilitate a new network approach as nearby objects have stronger connectivities and naturally form a community. A new community detection method is then applied to the aligned fMRI time series. We demonstrate the advantages of such an object alignment approach through a study of the functional connectivity for Alzheimer's disease. Beyond fMRI data, the proposed approach is broadly applicable to any high-dimensional or spatial temporal data.
Finally, the proposed alignment approach leads to two new approaches (see sections “Path Length of the Aligned Data” and “A New Community Detection Method”) to quantify the brain connectivity, which were successfully applied to BOLD data as described in the section “Resting-State fMRI Data.”
Materials and Methods
Resting-state fMRI data
The resting-state fMRI data are from a study of Alzheimer's disease at the University of California, Davis. The study included 172 Alzheimer patients and 67 normal subjects, for whom resting-state fMRI data were obtained for 8 min, resulting in 240 time points of image acquisitions, but the first four observations were discarded to let the scanner magnetization achieve a steady state, so the final length of the BOLD time series was m = 236. The data were preprocessed with SPM8 according to a standard protocol: time-slicing correction; head motion correction; co-registration (by minimizing the normalized mutual information); normalization; regressing out nuisance parameters, which include six motion parameters, cerebrospinal fluid (CSF) signal, white matter signal, and global signal; and bandpass filtering with cutoff frequencies of 0.01 and 0.08 Hz. The normalization step follows the standard settings of SPM8, and the affine transformation is performed through a nonlinear deformation to align with the MNI template provided by SPM8. The whole-brain BOLD signals were then summarized into 90 cortical regions based on the automated anatomical labeling (AAL) system. The signals representing these 90 regions were obtained by averaging BOLD signals around the seed voxels, defined by locally regional homogeneity (Zang et al., 2004).
Alignment of time series
Consider n spatially indexed objects, X
1, …, Xn
, whose spatial location are in a p-dimensional space. These n objects are endowed with a distance (or disparity) matrix D = [djk
], where djk
= d(Xj
, Xk
) is a distance function between two objects Xj
and Xk
. MDS is a visualization and dimension reduction tool to map these n objects to n new locations,
Here, q < p and q is usually preselected as q = 1, 2, or 3 for visualization purpose, so that the original data, which could be abstract objects on a higher (p)-dimensional space, can now be visualized in a lower (q)-dimensional Euclidean space. The special case q = 1, which is called unidimensional scaling (UDS), is the focus of this article. It has the attractive feature that it aligns all the objects to line up on an interval, creating a total ordering for these n objects. Alternatively, one may assume that the original objects have a latent order and that UDS aims at recovering this order. For BOLD objects, the alignment produces a 2D image for each subject, where the horizontal axis marks the new aligned location of each region and its corresponding (temporal) BOLD signals are displayed on the vertical axis.
For the functional connectivity application, the object is the BOLD time series at a brain region and there are 90 such regions rendering n = 90 objects, spatially indexed in a p = 3 dimensional space. For the jth object, it is the BOLD time series Xj = (Xj 1, …, Xjm ) at the jth brain region, measured at m time points. For the distance djk, we use a function of the Pearson correlation (PC) between the BOLD time series at the jth and kth brain regions since PC is arguably the most popular measure of the functional connectivity for BOLD data (Bandettini et al., 1993; Biswal et al., 1995; Cordes et al., 2001; Greicius et al., 2003; Hampson et al., 2002). However, our approach can be based on other correlations, such as the Spearman rank correlation (Hollander et al., 2013; Spearman, 1904) or Kendall's τ (Kendall, 1938, 1962), and other similarity measures for time series (Ferreira and Zhao, 2016).
Let Xj
= (Xj
1, …, Xjm
) and Xk
= (Xk
1, …, Xkm
) be two BOLD time series objects measured at m time points. We assume that they have been normalized by their respective temporal means and variances, so
Distance measure
While PC is originally designed for independent and identically distributed paired data, it has been used for paired time series under the stationary assumption. However, the PC should not be viewed as a measure of a statistical correlation when the time series data are not stationary, as spurious correlations may be triggered for a pair of nonstationary time series (Granger and Newbold, 1974). To overcome this shortcoming and to avoid the stationary assumption, we took a novel view that regards the BOLD time series as a discrete realization of a stochastic process in L
2 (Hsing and Eubank, 2015) and use the cosine of the angle between two centered L
2-processes as a measure of similarity. Here, for any two L
2-processes, X and Y, the cosine of the angle for the two centered processes is defined as
The advantage of this holistic view is that there is no need to assume stationarity of the time series, and a distance (or disparity) measure between Xj
and Xk
can be constructed as
Implementation of MDS
Let
We note here that although MDS aims at finding a global minimizer to achieve a perfect total ordering, it is not critical to have a perfect ordering for a method to be effective in data applications. First, MDS is a powerful visualization tool as demonstrated by the heat maps in Figure 3. Second, a key purpose of aligning objects is to create smoothly transitioned objects to facilitate further data analysis. Figure 1 demonstrates this concept, where a smooth process Z(s, t) in Figure 1a represents smoothly transitioned objects Zs
(t) = Z(s, t), which are indexed by s and lined-up vertically along the horizontal s-axis. These initial indices s are then randomly permuted, and the corresponding perturbed objects are displayed in Figure 1b, which are subsequently aligned using our MDS method. The result is an imperfectly aligned data, where the aligned objects in Figure 1c, which are reconstructed from the randomly perturbed data in Figure 1b, appear to be smoothly transitioned similar to the smooth process in Figure 1a. For this reason, one can work with just the information of the order of the configuration

For the remainder of the article, we assume, for simplicity, that MDS maps the original objects {X
1, …, Xn
} to new locations
Simulation
To evaluate the effectiveness of the alignment method, we conduct the following simulation.
1. Generate n = 100 equidistant spatial grid points in
2. Generate BOLD signals Zj
(t) = Z(t, sj
) for t = 1, …, 236 as follows:
where r is an integer,
where
3. Perturb Zj (t) to Zπ (j)(t), where π is a permutation function on 1, …, 100 and let Xj (t) = Zπ (j)(t) be the objects to be aligned in the next step.
4. Perform UDS as described in the section “Alignment of Time Series” to align X
1, …, X
100 to new reconfigured locations
The Fourier basis functions in step 2 were selected to mimic the fMRI data in the section “Resting-State fMRI Data,” which were measured at m = 236 time points. Note that we have added noise, εj
(t), in step 2 to the BOLD signals to reflect reality. The resulting BOLD signals Zj
(t) = Z(t, sj
) in step 2, although noisy, are endowed with a natural ordering from the original process Z(t, s), which we take to be the latent order of Zj
(t). The perturbation of this latent order in step 3 disturbs the ordering of these smoothly transitioned objects, and the UDS in step 4 aims at recovering the original latent order. One challenge with the simulation is that there is no genuine true order for the objects generated from this experiment since djk
= 2(1−ρjk
), where ρjk
is the PC in Equation (1), is not necessarily the best metric to capture the total order behind the stochastic process. However, it is reasonable to use the spatial order of the smoothly transitioned objects as a surrogate for the true order. We adopt this approach but emphasize that this is a challenge in the simulation, as one cannot expect perfect alignment when evaluating the simulation results. Despite this challenge, the proposed alignment procedure performs well in the simulation based on 500 runs mimicking 500 random subjects. To evaluate the performance of the simulation, we use a criterion based on the relative ordering error (ROE),
where n is the number of objects;
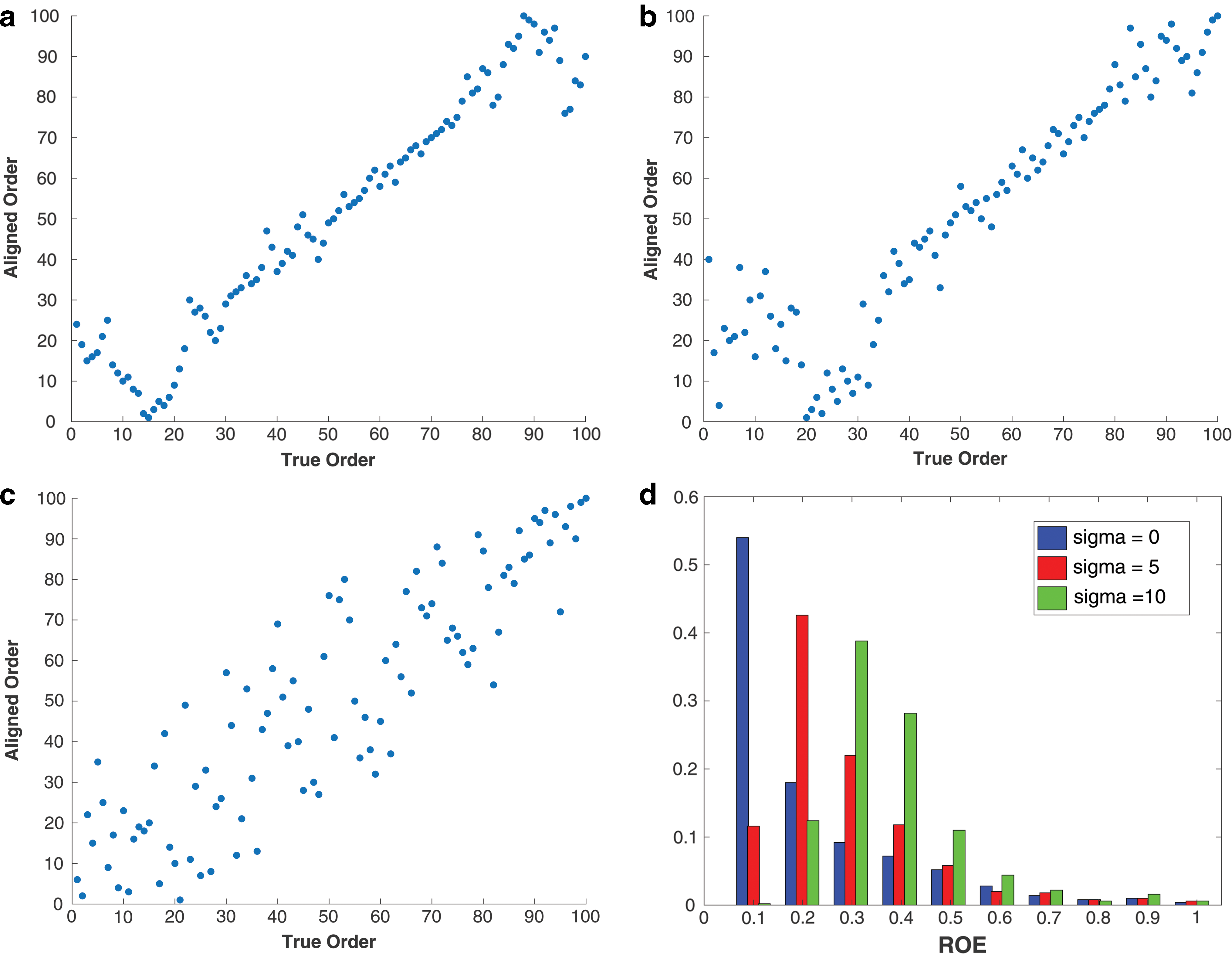
Simulation results with median ROE under different noise levels (σ = 0, 5, 10) are reported in
Path length of the aligned data
In network modeling, a popular approach to study the brain connectivity, ROIs are considered as nodes, and edges between them are established based on their functional connectivity. Many statistics have been proposed to characterize networks, and path length is often used to summarize the efficiency of the network. Recently, the minimum spanning tree (MST) of brain networks has been proposed to summarize the global efficiency of brain function (Olde Dubbelink et al., 2014; Stam et al., 2014; van Dellen et al., 2014). It selects the most efficient and essential path, which connects all the nodes. While this is an appealing way to summarize a network, it fails to detect the differences between the brain connectivity of the normal and demented subjects for the Alzheimer data in the section “Resting-State fMRI Data.” We thus propose a new approach that compares the two groups on a different path, where adjacency of the objects/nodes is determined by the order of the reconfigured brain locations. Specifically, the alignment facilitates an ordered path, ψ(1) → ψ(2) → ψ(3)… → ψ(n − 1) → ψ(n), of the brain connectivity of a subject. We use their ordered path to summarize the efficiency of the brain connectivity, defining a path length:
A traversal distance of a network is usually used to gauge network efficiency. Similar to MST, which is a breadth-first traversal algorithm, path length can be considered as a version of depth-first traversal algorithm, which can be used to measure network efficiency. It is conceivable that an inefficient brain will take longer to travel through, thus it is plausible that path length can capture the deficiency of the brain function of the Alzheimer patients. The data application in the section “Results” underscores the usefulness of this new notion of path length LA , as it is more effective in detecting the differences between the normal and demented subjects.
A new community detection method
Since the alignment in the section “Alignment of Time Series” arranges similar objects to nearby locations on an interval, one could view these similar objects as a community. An interesting question is how to detect communities by leveraging the reconfiguration after the alignment. The most widely used method to detect communities is to maximize the modularity on networks through thresholded or weighted adjacency matrices. In brain research, these adjacency matrices are usually constructed through functional connectivity maps of ROIs. We proposed a different approach here that takes advantage of the alignment. First, we consider the modularity criterion (Newman, 2006),
where wjk
is the weight of the edge between the jth and kth objects;
This criterion does not utilize the characteristics of the alignment based on the new reconfiguration proposed above. In the new configuration, one may postulate that objects that are close to each other belong to the same community. With this information, we set out to detect the community structure by locating the boundaries of communities on the new configuration. Since the reconfiguration lies on the 1D space, we formulate the problem as a change-point detection problem. Thus, the modularity criterion is changed to:
where (b
1, ⋯, bB
−1) are the change-points that serve as the boundaries of communities. That is, (b
1, ⋯, bB
−1) is a subset of
To tackle these challenges, we apply a genetic algorithm (Goldberg et al., 1989) to solve the optimization of the objective function [Eq. (6)]. In a genetic algorithm, candidate solutions are represented by finite-length of strings of alphabets, which are referred to as genes. Each gene is encoded as a binary value 0 or 1, with 1 denoting the boundary of a community. The candidate solutions (b 1, ⋯, bB −1) of the criteria function [Eq. (6)] are thus the locations of those genes encoded as 1. More details about the algorithm can be found in a later monograph (Goldberg, 2006).
Results
We apply the methods proposed in the section “Materials and Methods” to resting-state fMRI data from a study of Alzheimer's disease at the University of California, Davis. The study included 172 Alzheimer patients and 67 normal subjects, for whom resting-state fMRI data were obtained for 8 min, resulting in 240 time points of image acquisitions, but the first four observations were discarded to let the scanner magnetization achieve a steady state, so the final length of the BOLD time series was m = 236. The data were preprocessed with SPM8 and subsequently partitioned into 90 cortical regions (based on the AAL system) as described in the section “Resting-State fMRI Data.”
For each subject, this resulted in n = 90 time series, which were subsequently normalized to (X 1(t),⋯, X 90(t)), where the region locations were arranged in the same order as in the AAL system. The functional connectivity maps of the 90 brain regions are presented in Figure 3a and b for a normal and demented subject, respectively. These subjects are representative of their respective group in the sense that they have the median number of change-points within each group. We then applied the alignment approach to both subjects. The resulting functional connectivity maps based on the reconfiguration are shown in Figure 3c and d. We observe that the information in panels (a) and (b) is difficult to extract, whereas a clearer pattern emerged from each of the panels (c) and (d).

Brain image for a representative normal (
Length of selected path
The path length LA is computed for each subject, and the average path lengths for the normal and demented groups are 114.69 and 117.47, respectively. The Wilcoxon test reported in Table 1 suggests a marginal significant difference (p = 0.0934 for a two-sided alternative hypothesis) between the two groups and that Alzheimer's disease reduces brain efficiency (shorter path length is more efficient). Here, the Wilcoxon test is used since it does not rely on any parametric assumption and is fairly powerful in comparison with the t-test (Lehmann, 2012). In addition, we apply the MST method (Olde Dubbelink et al., 2014; Stam et al., 2014; van Dellen et al., 2014) to summarize the efficiency of brain network. The average shortest path for the normal and demented groups is 60.37 and 62.29, respectively, but the result is less significant (p = 0.1361 for the Wilcoxon test). This suggests that the new measure LA is a more effective summary than the MST in existing literature to distinguish normal from demented subjects for brain efficiency.
Test of Network Efficiency
Normal group is indexed by 1 and demented group by 2.
The p-values are calculated from the Wilcoxon rank-sum test.
MST, minimum spanning tree.
Community detection
In addition to using the selected path length to measure the efficiency of brain function, we are also interested in exploring another important topological property of brain networks, the community structure in terms of modularity. A modular organization characterizes many important biological systems, including brain networks. The modularity is considered as a topological structure that can combine advantages of high clustering and high efficiency and optimize between the brain wiring cost and efficiency (Bullmore and Sporns, 2012). This concept is then utilized to detect community structures by maximizing Q as commonly performed (Newman, 2006). Given the new configuration obtained through the proposed alignment method and based on the discussion in the section “A New Community Detection Method,” we adopted the modified Q in Equation (6) to detect the community structure. This modularity is used to measure how well the defined communities maximize the within-group connection and minimize the between-group connection.
Inspired by the framework of exponential random graph modeling (Simpson et al., 2011; Van Wijk et al., 2010), we apply a transformation wjk
= exp(ρjk
) to transfer the cosine angle into a nonnegative weight. We applied the change-point analysis in the section “A New Community Detection Method” to each subject and observed block diagonal structures for both the normal and demented subjects in Figures 4c and d. Furthermore, we applied a threshold
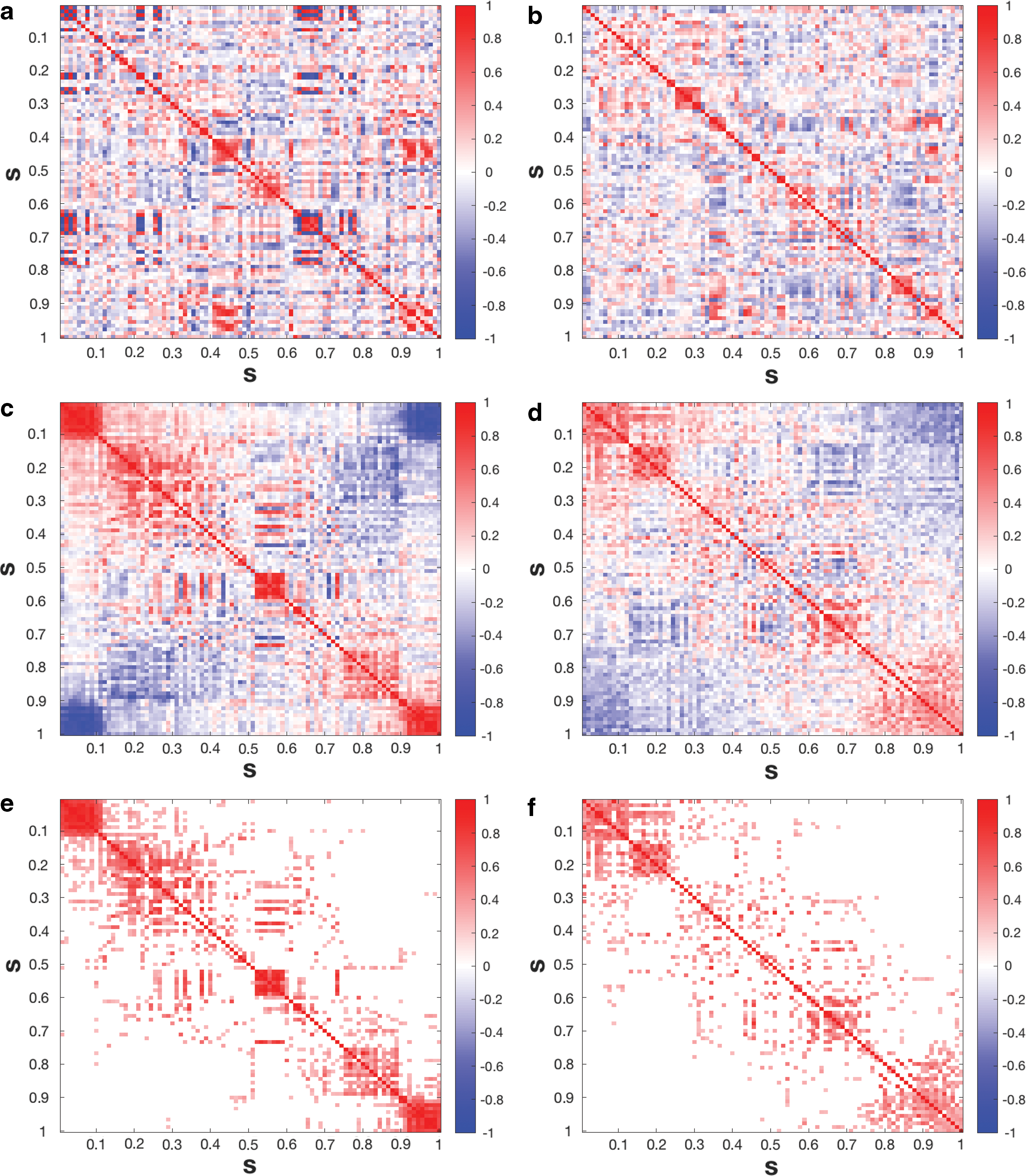
Brain image for a representative normal (
We also compared our approach with the algorithm from the brain connectivity toolbox [BCT; Rubinov and Sporns (2010)] on all subjects. The results shown in Figure 5a are based on our proposed algorithm and the Louvain Method for community detection and confirm that the communities detected by our approach are as good as those by BCT. In conclusion, these results indicate that our community detection approach conveys valuable information.
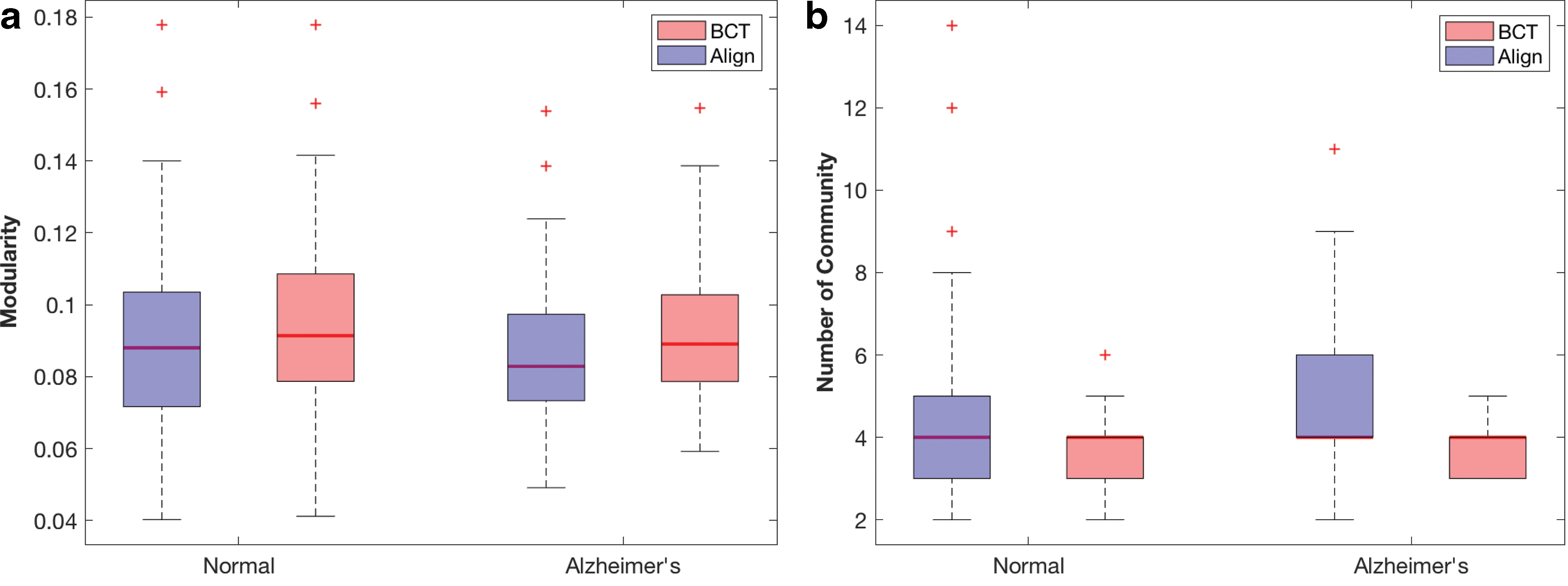
Finally, we compared modularities and numbers of communities between the normal and demented subjects. The average modularity, as shown in Table 2, is 0.0896 and 0.0849 for the normal and demented groups, respectively. Although not statistically significant, our method led to a smaller p-value (p = 0.1210) than the one from BCT (p = 0.1539), suggesting that the demented group seems to have less efficient community structures.
Test of Modularity
Normal group is indexed by 1 and demented group by 2.
The p-values are calculated from the Wilcoxon rank-sum test.
BCT, brain connectivity toolbox.
The average number of communities, as shown in Table 3, is 4.2500 and 4.8955 for the normal and demented groups, respectively. Our method shows that the two groups differ significantly (p = 0.0032) in terms of the number of communities with the demented group split into more communities. But BCT is less effective to detect this difference (p = 0.0625). Combining the results in Tables 2 and 3 and Figure 5 with the findings on the path length, we conclude that in comparison to the normal subjects, the demented subjects not only have less global efficiency but also have less intact local functional modules.
Test of the Number of Communities
Normal group is indexed by 1 and demented group by 2.
The p-values are calculated from the Wilcoxon rank-sum test.
Discussion and Conclusion
We propose a new approach to visualize neuroimaging data using BOLD signals as the platform. The proposed approach compresses the 3D spatial locations of the brain regions into spatial locations of a 1D space, so that the temporal signals of fMRI time series can be aligned along a horizontal axis and the heat map (Fig. 3) can be used to visualize the aligned objects. In addition to visualization, two new approaches emerged from the alignment idea. The first is a new summary for the global efficiency of the functional connectivity of an individual brain. This new summary is based on the path length along the aligned brain location and is shown in the section “Results” to be effective in distinguishing Alzheimer's and normal subjects, thereby providing new insights in how Alzheimer's disease alters the structure of brain connectivity.
The second by-product is a new method to detect communities through change-points locations of the functional connectivity as brain regions between two change-points are more similarly connected than those across change-points. Thus, instead of examining a 90 × 90 adjacency matrix, the alignment reduces the adjacency matrix to diagonal subblocks of sizes n 1, …, nB . Such a data reduction is feasible because we have reconfigured the objects by aligning them. When using the weighted modularity criteria, our community detection method performs competitively with the BCT approach.
In summary, the alignment approach opens a new way to explore complex spatially index objects. While we illustrate our approach with time series objects, the proposed method is applicable to any functions or abstract objects. As we have only performed the alignment at the subject level, an interesting question is whether it makes sense to do a unified alignment for a group of subjects, that is, whether it is reasonable to use the same ordering for all subjects in a group. The answer is likely to depend on the homogeneity of the group. One major advantage of the alignment method is that it alleviates or overcomes the curse of high dimensionality for object data. For instance, by aligning the BOLD signal of a subject, we provide an ordering for spatially index signals, so the realigned BOLD signals could be viewed as longitudinal functional data (Chen and Müller, 2012) or smooth 2D functional data.
Since methods to deal with such functional data are plentiful due to the fast growing interest in functional data analysis (Ramsay and Silverman, 2007; Wang et al., 2016), the alignment approach proposed in this article bridges a gap between the analysis for high-dimensional object data and smooth object data. There is clearly a vast array of methods one can borrow from the statistical literature of functional data analysis once the alignment has been completed, including functional principle component analysis, classification, clustering, and prediction. This could be an interesting future direction to explore.
Footnotes
Acknowledgment
The research of J.-L.W. is supported by NSF grant DMS-15-12975 and NIH Grants 7UG3OD023313 and 1UH3OD023313.
Author Disclosure Statement
No competing financial interests exist.