
Abstract
The thalamo-cortical circuit is important in the genesis of absence epilepsy. This circuit can be influenced by connecting pathways from various parts of central nervous system. The aim of the present study is to define the dento-thalamic connections in Wistar animals and compare the results with genetic absence epilepsy rats from Strasbourg (GAERS) using the biotinylated dextran amine (BDA) tracer. We injected BDA into the dentate nucleus of 13 (n = 6 Wistar and n = 7 GAERS) animals. The dento-thalamic connections in the Wistar animals were denser and were connected to a wider range of thalamic nuclei compared with GAERS. The dentate nucleus was bilaterally connected to the central (central medial [CM], paracentral [PC]), ventral (ventral medial [VM], ventral lateral [VL], and ventral posterior lateral [VPL]), and posterior (Po) thalamic nuclei in Wistar animals. The majority of these connections were dense contralaterally and scarce ipsilaterally. Contralateral connections were present with the parafascicular (PF), ventral posterior medial, ventral anterior (VA), and central lateral (CL) thalamic nuclei in Wistar animals. Whereas in GAERS, bilateral connections were observed with the VL and CM. Contralateral connections were present with the PC, VM, VA, and PF thalamic nuclei in GAERS. The CL, VPL, and Po thalamic nucleus connections were not observed in GAERS. The present study showed weak/deficit dento-thalamic connections in GAERS compared with control Wistar animals. The scarce information flow from the dentate nucleus to thalamus in GAERS may have a deficient modulatory role on the thalamus and thus may affect modulation of the thalamo-cortical circuit.
Introduction
The cerebellum is no longer considered as a brain area related solely to motor skills. Experimental and clinical studies showed involvement of the cerebellum in cognitive functions (Albert et al., 2017; Budisavljevic and Ramnani, 2012; Igelstrom et al., 2017; Ignashchenkova et al., 2009; Manto, 2015; Shakiba, 2014; Stoodley and Limperopoulos, 2016; Verly et al., 2014). The impairment of execution of motor, cognitive, and sensorimotor tasks and learning was reported as dysfunction of cerebello-striato-cortical loops (Caligiore et al., 2017; Desmond and Fiez, 1998; Tzvi et al., 2017; Schmahmann, 2010; Middleton and Strick, 1997). Involvement of the dentate nucleus in autism, psychogenic ataxia-like symptoms, and impaired resting-state loops were reported as dysfunctions of cerebello-thalamo-cortical loops (Danielson et al., 2011; Olivito et al., 2017; Salih et al., 2010).
The cerebellum is connected to wide regions of the central nervous system (Bostan et al., 2013; Courchesne and Allen, 1997). The density of input to the cerebellum is 40 times higher than its output (Kandel and Mack, 2014), which shows the importance of its integrative functions (Stoodley and Schmahmann, 2009). The majority of the inputs of the cerebellum are to the cerebellar cortex, the outputs are through the deep cerebellar nuclei (lateral, medial and anterior, and posterior [Po] interposed).
The dentate nucleus (lateral nucleus) of the cerebellum has well-organized output channels that project to the primary motor, premotor, oculomotor, and prefrontal cortical areas through the thalamus (Middleton and Strick, 1998). It has been reported that the dentate nucleus has specific organization, the dorsal parts of the nucleus project to the motor regions of the cerebral cortex through the motor relays of the thalamus, ventral parts of the nucleus project to nonmotor areas, and the prefrontal cortex through the ventral lateral (VL) thalamic nuclei (Bernard et al., 2013; Dum and Strick, 2003; Joyal et al., 2001; Kim et al., 1994; Middleton and Strick, 2001). Projections from both dorsal and ventral parts of the dentate nucleus take part in various important circuits, which have a role in both motor and nonmotor functions (Balsters et al., 2013; Bond et al., 2017; Middleton and Strick, 2001; Olivito et al., 2017;; Ramnani, 2012; Salih et al., 2010).
The majority of the information flow from the deep cerebellar nuclei to the cerebral cortex is through the thalamus. The thalamus is the main relay station of information before it reaches the cortex. The thalamic relay processes information that comes from the cerebellum and conveys it to its cortical target (Sherman and Guillery, 2009). However, what is it that the thalamus does to messages that flow from the dentate nucleus before it is transmitted to the cortex has not been demonstrated yet.
The important role of thalamo-cortical circuits in the genesis of absence epilepsy is well documented in both humans (Bertram, 2013; Crunelli and Leresche, 2002; Danober et al., 1998; Fletcher et al., 1996; Moeller et al., 2008; Yang et al., 2012) and experimental animals (Kandratavicius et al., 2014; McCormick and Contreras, 2001; Meeren et al., 2002). Studies have shown that thalamo-cortical circuits can be influenced by various parts of the central nervous system, which diverted researchers to examine the role of different parts of brain regions that influence the thalamo-cortical circuit.
Absence epilepsy is a neurological disease characterized by the presence of bilaterally synchronous spike-wave discharges (SWDs) in the encephalogram accompanied by loss of consciousness. The genetic absence epilepsy rats from Strasbourg (GAERS) are an inbred Wistar animal strain and are regarded as a reliable model of genetic generalized epilepsy and most of the genetic, pharmacological, and behavioral characteristics are similar to those observed in humans (Çarçak et al., 2009; Danober et al., 1998; Vergnes et al., 1986).
Disrupting thalamo-cortical activity patterns has proven to be a promising approach to stop generalized SWDs. Recent clinical and experimental studies showed the involvement of the cerebellum in the mechanism of absence epilepsy (Blumenfeld, 2012). Kros et al. (2015) showed that the cerebellar nuclei are potent modulators of pathological oscillations in thalamo-cortical network activity during absence seizures. Furthermore, Kros et al. (2015) provided strong evidence that cortical seizures can be stopped by optogenetic stimulation of the dentate and interposed cerebellar nuclei. Absence epilepsy is a brain network disease, thus in the present study, we aim to evaluate dento-thalamic connections using the biotinylated dextran amine (BDA) tracer in GAERS and control Wistar animals and compare the difference between the two strains.
Materials and Methods
In the present study, a total of 37 animals weighing 250–300 g were used for the BDA injections. Of the 37 injections, 13 (n = 6 Wistar and n = 7 GAERS) with minimal contamination of adjacent structures or along the pipette tract were included in the study (Fig. 1A, B). All animals were fed with a standard laboratory rat chow and tap water ad libitum and housed in Plexiglass cages with a 12-h light/12-h dark cycle in a temperature-controlled room (20°C ± 3°C). All procedures and animal housing before and after surgeries were approved by the Animal Care and Use Committee of Marmara University.
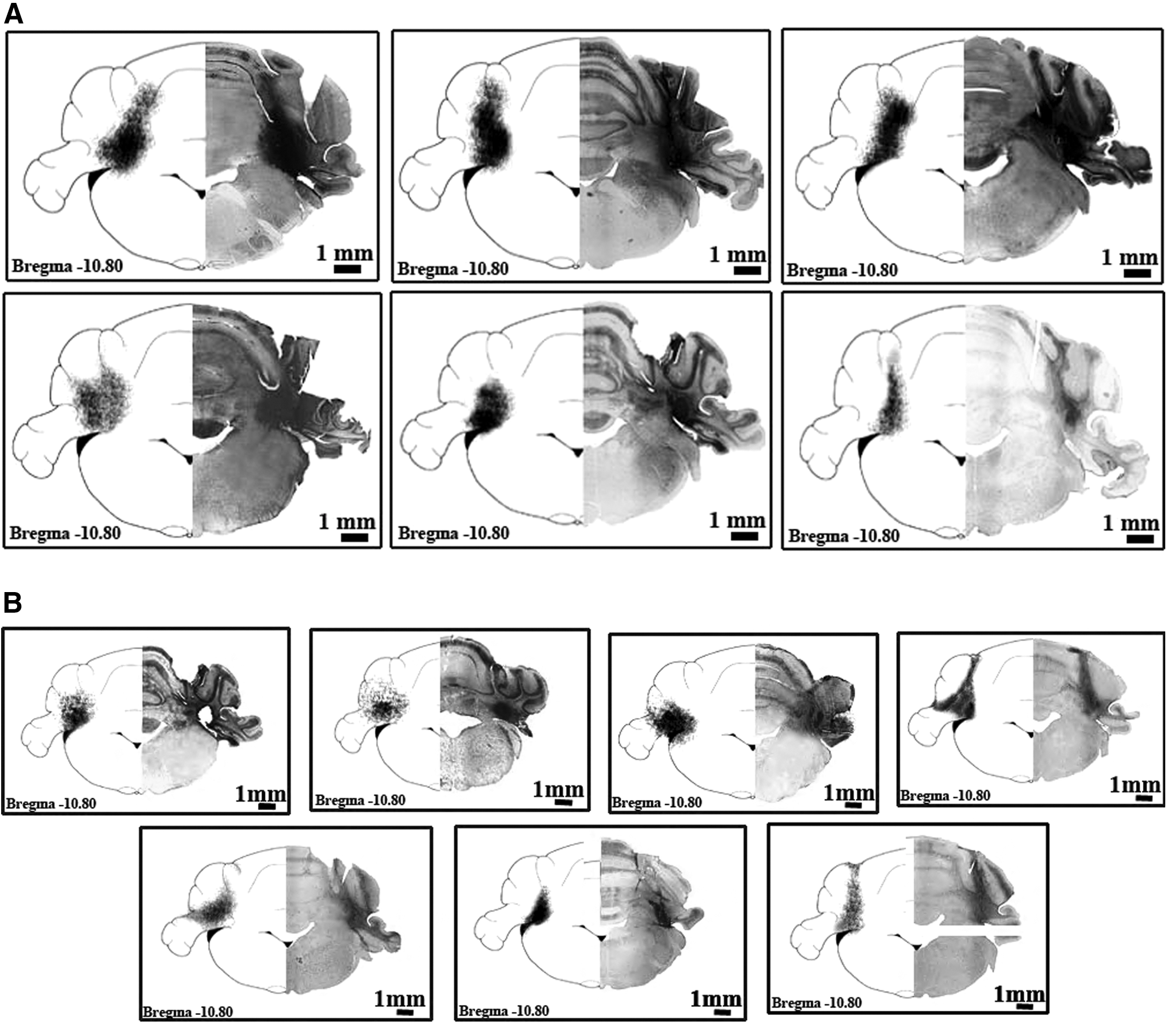
All animals were deeply anesthetized with intraperitoneal injection of the ketamine/xylazine mixture and surgery started after no cornea and extremity reflexes were observed. The 10 kDa BDA (Invitrogen D-1956, Dextran, Biotin, Lysine Flexible, BDA-10.000) of 0.4 μL was injected using a stereotaxic instrument to the coordinates of the dentate nucleus (AP: −10.8 mm, L: 3.6 mm, V: 6.5 mm) of adult Wistar rats (n = 6) and GAERS (n = 7; Fig. 1A, B). Injections were performed by using a Hamilton syringe (HAM-80008, 1701 SN 10 μL Syr) and Neurostar motorized stereotaxic frame (Neurostar, Tübingen, Germany). All animals survived for 7–10 days after BDA injections and animals were transcardially perfused with saline (0.9% NaCl, pH 7.45; 200 mL), followed by 500 mL of phosphate-buffered fixative containing 2% paraformaldehyde (PFA) and 1.5% glutaraldehyde (GA) solution (in 0.1 M phosphate-buffered saline [PBS], pH 7.45; PBS Amresco E404-200TABS, PFA MERCK 1.04005.1000, GA MERCK 8.20603.1000). The brains were removed and postfixed in the same fixative solution overnight. All brains were collected in 30% sucrose solution at +4°C. Coronal serial sections (50 μm) were cut using a cryomicrotome (Leica CM 1950) and free-floating sections were collected in well plates.
BDA histochemistry
Histochemistry was applied to every fourth section to visualize the BDA in the tissue, and sections were run in the avidin-biotinylated HRP (ABC) procedure, followed by the metal-enhanced diaminobenzidine (DAB-Ni) reaction. The procedure started with 10 min of sodium borohydride (Sigma-Aldrich S38714-486) incubation; PBS 0.1 M washing, twice for 10 min each; ABC standard solution (Vector Laboratories PK6100; Vectastain Elite ABC Standard Kit) incubation for 3 h at room temperature; PBS 0.1 M wash for 10 min; Tris buffer (TB) 0.05 M (pH: 8.0) washes, twice for 10 min each; DAB-Ni [DAB Sigma D5905, ammonium nickel(II) sulfate hexahydrate Sigma A1827] preincubation in a dark room for 15 min; H2O2 reaction for 3–15 min in a dark room; TB 0.05 M (pH: 8.0) wash for 10 min, followed by PBS 0.1 M wash for 10 min; and finally, the sections were mounted. The sections were examined at 40× magnification under a light microscope. Ipsilateral, contralateral, and bilateral labeling was documented on four sections for each nucleus and for each animal. The density of labeled axons was evaluated (labeled) as follows: * scarce for the fiber number below 10, ** moderate for between 10 and 20, and *** dense for the cell number above 20.
Results
A total of 13 animals were included in the study (Fig. 1A, B). Subsequent to BDA injections into the dentate nucleus, only constantly labeled axons and cells in thalamic nuclei were documented (Table 1). The labeled axons and cells in thalamic nuclei, which were present in at least four animals for each strain, are included in the results, see Table 1. The densities of bilateral, ipsilateral, and contralateral dento-thalamic connections were documented for both GAERS and Wistar animals (Table 1).
The Density of Ipsilateral and Contralateral Dento-Thalamic Connections of GAERS and Wistar Animals
scarce for the fiber number below 10, ** moderate for between 10 and 20, and *** dense for the cell number above 20.
CL, central lateral; CM, central medial; PC, paracentral thalamic nucleus; PF, parafascicular; Po, posterior; VA, ventral anterior; VL, ventral lateral; VPL, ventral posterior lateral; VM, ventral medial; VPM, ventral posterior medial.
Dento-thalamic connections in Wistar animals
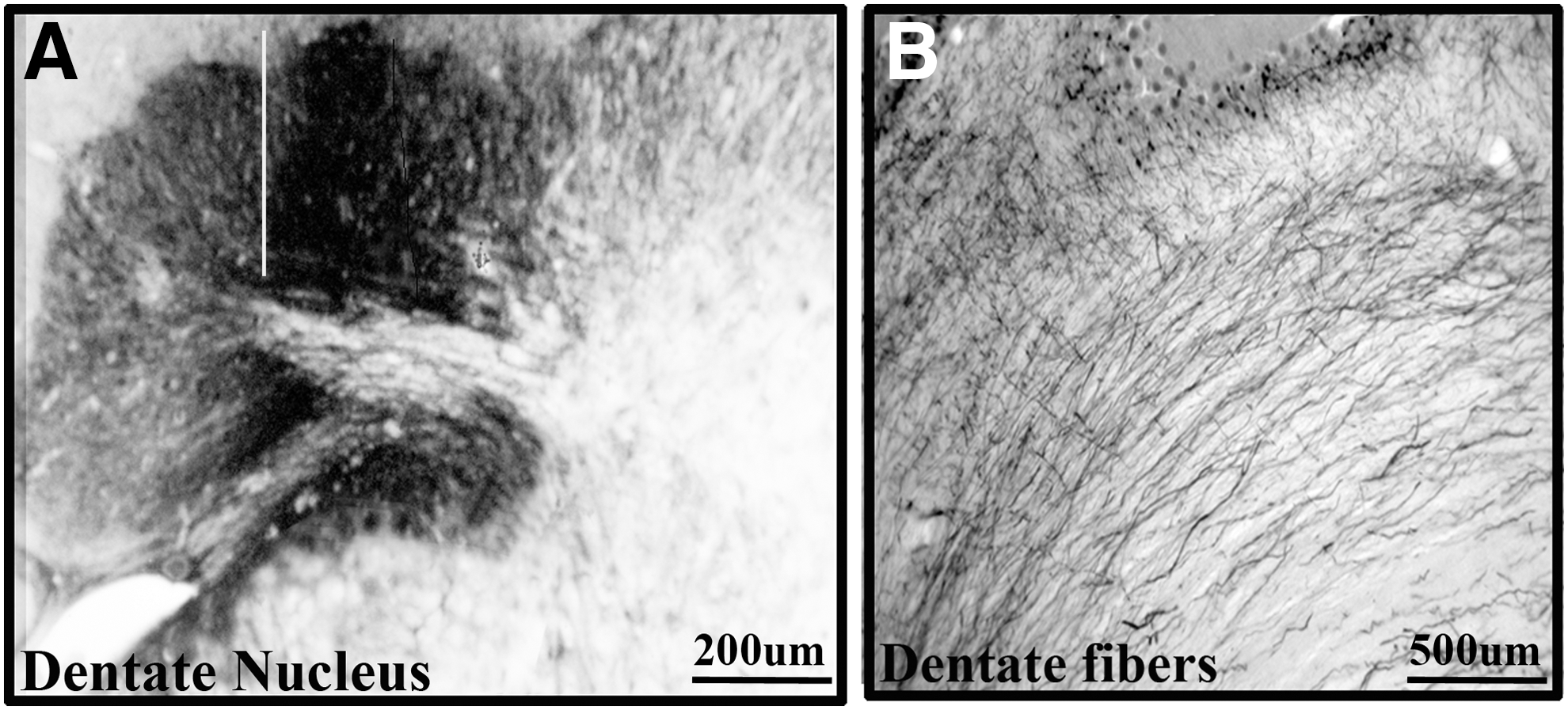
The dentate nucleus was connected to a wide range of thalamic nuclei in Wistar animals, see Table 1. The major outputs of the cerebellum are from the dentate nucleus. The dentate nucleus consists of a hilum directed medially and the axons exit through the hilum (Fig. 2A, B). The dentate nucleus was bilaterally connected to central (central medial [CM] and paracentral [PC]), ventral (ventral medial [VM], VL, and ventral posterior lateral [VPL]), and Po thalamic nuclei in Wistar animals (Fig. 3A–E). BDA labeling was denser in the contralateral compared with ipsilateral-related thalamic nuclei. Only contralateral connections were present with the central lateral (CL), ventral anterior (VA), ventral posterior medial (VPM), and parafascicular (PF) thalamic nuclei in Wistar animals (Fig. 4A, B). Among dento-thalamic connections, the dentate nucleus was most strongly connected to the VL and Po thalamic nuclei (Table 1).
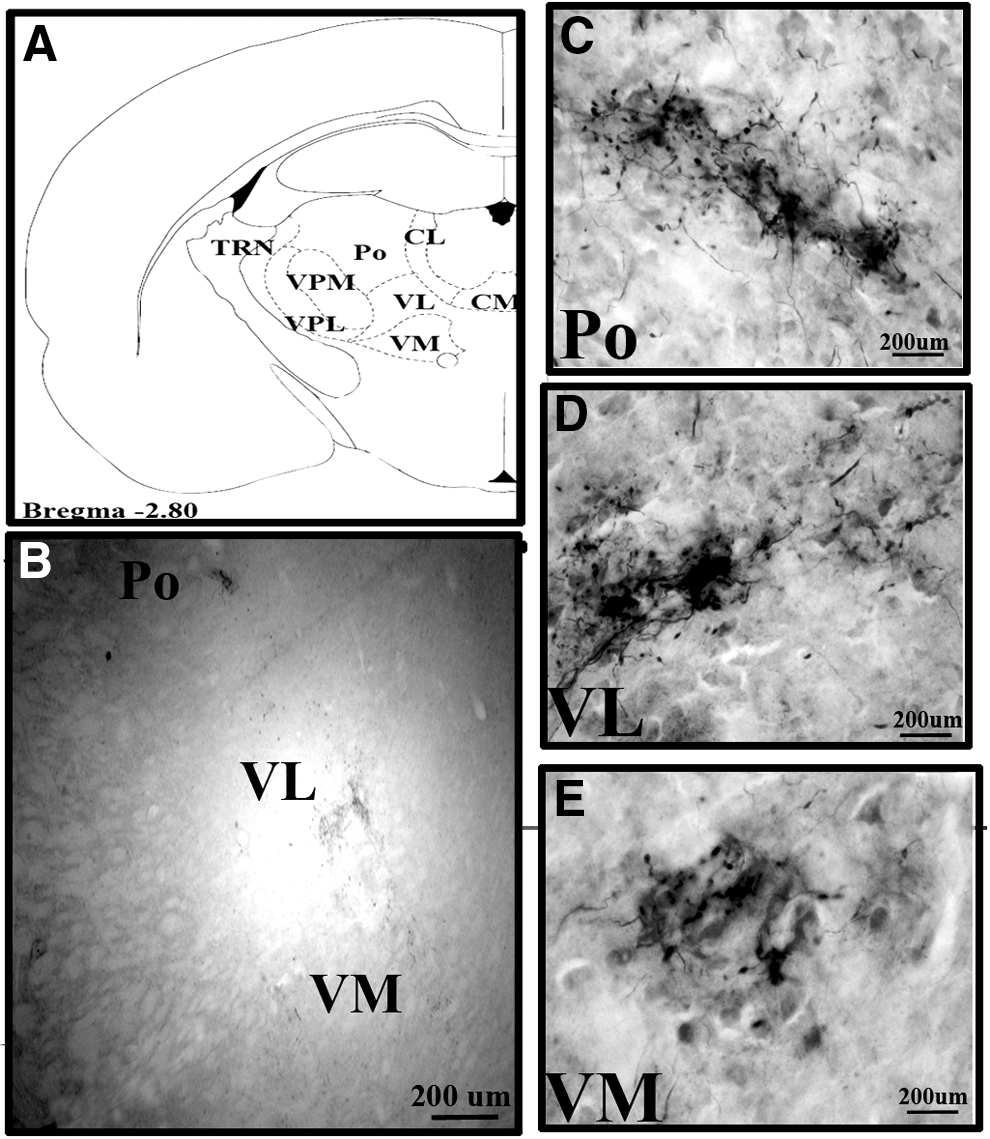
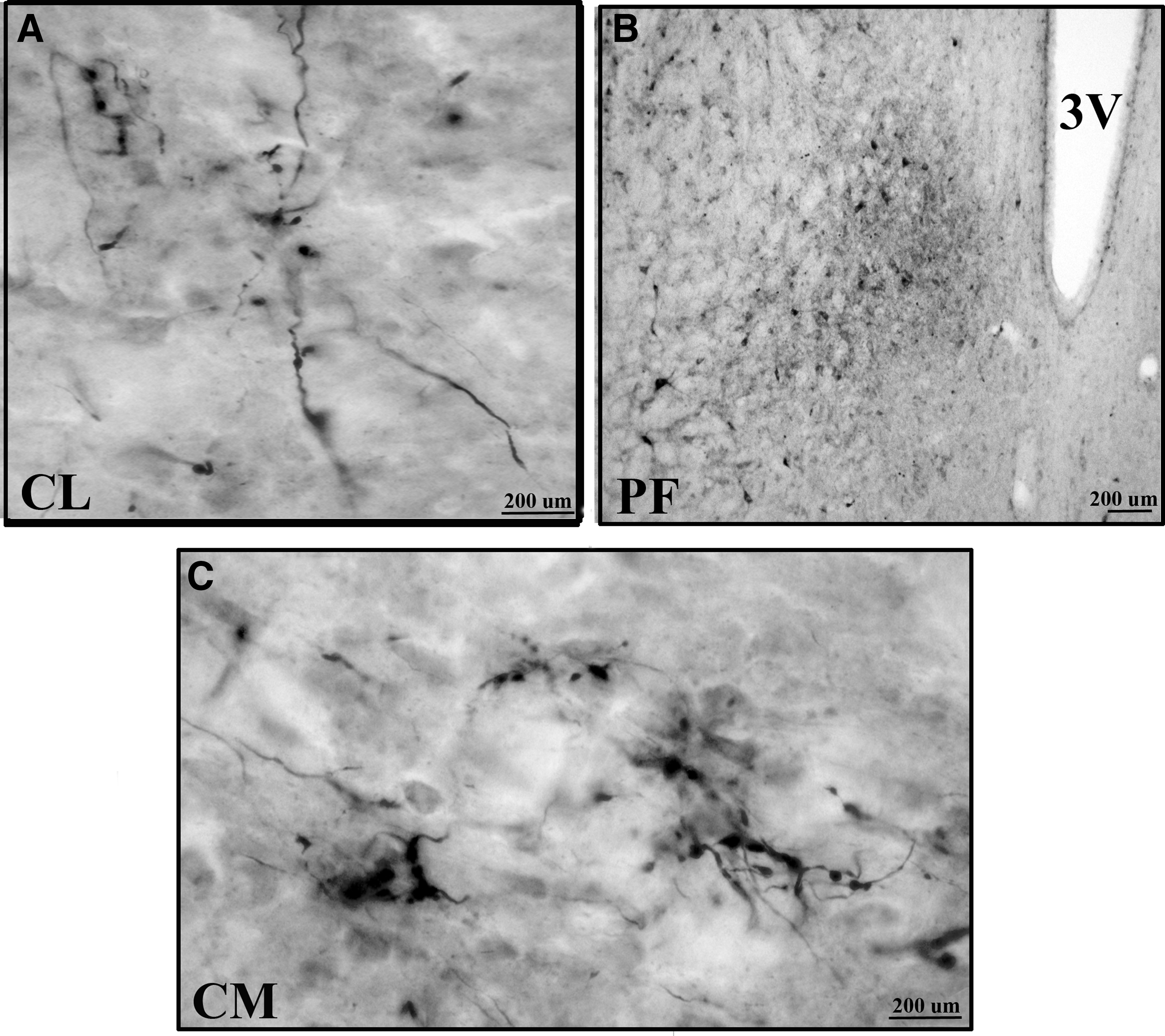
Dento-thalamic connections in GAERS
The dentate nucleus was connected to restricted thalamic nuclei in GAERS compared with Wistar animals, see Table 1. The dentate nucleus was connected to only VL and CM thalamic nuclei, bilaterally and contralaterally to the PC, VM, VA, and PF thalamic nuclei (Fig. 4C). Among the dento-thalamic connections, the dentate nucleus was strongly connected to the contralateral VL thalamic nuclei in GAERS (Table 1). The majority of dento-thalamic connections in GAERS were contralateral (Table 1). The CL, VPL, and Po thalamic nucleus connections were not observed in GAERS. The dento-thalamic connections in GAERS were weaker compared with Wistar animals (Fig. 5A, B).
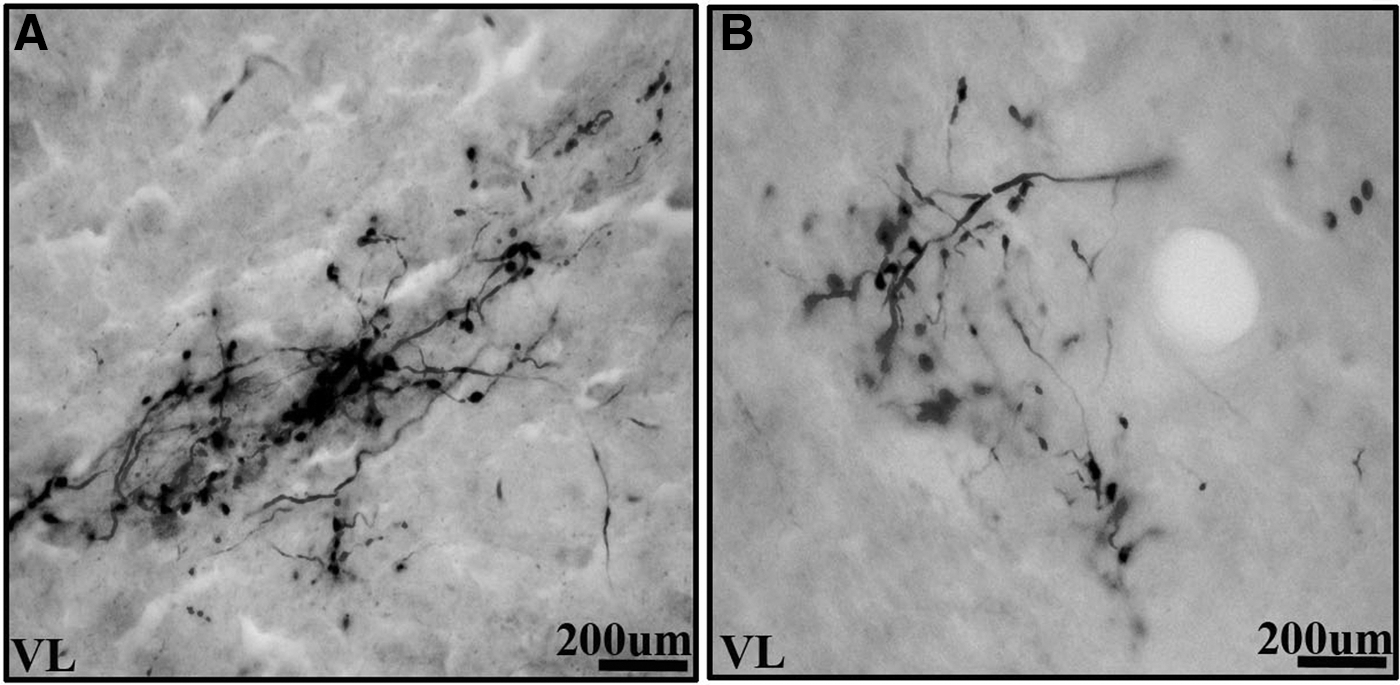
Comparison of the density of labeled axons in the VL thalamic nuclei of the two experimental groups
Discussion
The dento-thalamic connections were much denser and connected to a wider range of thalamic nuclei in Wistar animals compared with GAERS. The scarce information flow from the dentate nucleus to thalamus in GAERS may have a deficient modulatory role on the thalamus and thus may affect modulation of the thalamo-cortical circuit.
There are studies that report on connections of the dentate with specific thalamic nuclei in cats, monkeys, and humans using tract tracers and diffusion MRI (Bernard et al., 2013; Holsapple et al., 1991; Jimenez-Castellanos and Reinoso-Suarez, 1985; Pelzer et al., 2017). However, collective data regarding the dento-thalamic connections is rather sparse. The majority published connections highly correspond (VPM, VPL, VL, Po, IL, PF, and VM) with our results (Aumann et al., 1994; Bentivoglio and Kuypers, 1982; Bernard et al., 2013; Cavdar et al., 2018; Faull and Carman, 1978; Middleton and Strick, 1997). Gornati et al. (2018) reported that in vitro electrophysiological recordings revealed that optogenetic dentate nucleus stimulation resulted in enhanced charge transfer and action potential firing in VL neurons compared with VM or CL neurons. The present study has shown that VL connections were more dense compared with VM or CL, which may be the reason for the enhanced charge transfer and action potential firing in VL compared with VM and CL. Furthermore, single-axon reconstructions of cerebellar-recipient zones within VL, VM, and CL reveal that their axons also spread throughout other regions, highlighting that the cerebellar input can affect a wide range of thalamo-cortical networks and functions (Deschênes et al., 1996).
White matter serves a critical role in organization of neural networks (Catani et al., 2012). Advances in brain imaging have revealed white matter impairment in most forms of neurodegeneration diseases, including Alzheimer's (Zhang et al., 2014), Huntington's (Gatto et al., 2015), and Parkinson's diseases (Duncan et al., 2016; Zhai et al., 2016), and psychological and behavioral disorders (Lener and Iosifescu, 2015; Seitz et al., 2016). There are studies that demonstrate white matter abnormalities in patients with temporal lobe epilepsy (Rodríguez-Cruces and Concha, 2015) and absence epilepsy (Correa et al., 2017). The present experimental study showed the absence of or weak dento-thalamic connections in GAERS, which may have a role in the pathophysiology of absence seizures.
Lesion studies suggested that thalamic nuclei, particularly the VL, VA, and VPL, have a permissive effect on partial and generalized seizures in animal models (Gasteiger et al., 1985; Mirski and Ferrendelli, 1984). The results of our study showed that VPL connections were not present in GAERS. Furthermore, Vergnes and Marescaux (1992) induced cortical and thalamic lesions and reported the effects on SWDs in genetic absence rat models. These results demonstrate that lesions of the lateral thalamus, including the specific relays (VPL, VPM, Po, VL, MDl, MDc, CL, and PC) and reticular nucleus, suppressed ipsilateral SWDs. The present study showed weak dento-thalamic connections with the former thalamic relay nuclei in GAERS compared with control Wistar animals, which may suggest an insufficient modulation of the thalamic relay nuclei through the dentate nucleus. Feline generalized penicillin animal epilepsy models have shown that thalamo-cortical circuits can be influenced by connecting pathways for generation of typical SWD (Avoli and Kostopoulos, 1982; Norden and Blumenfeld, 2002). A recent study showed that cortical seizures can be manipulated by local injections of muscimol or gabazine to the interposed and dentate cerebellar nuclei, which strongly affected (increase and decrease, respectively) the occurrence of SWD at the cerebral cortex (Kros et al., 2015). Furthermore, Lüttjohann and van Luijtelaar (2012) studied interactions between the cortex and different thalamic nuclei in WAG/Rij rats for generation and maintenance of SWD using multisite cortical and thalamic local field potential recordings and reported that the earliest and strongest increases in coupling strength were seen between cortical layers 5/6 and the Po thalamic nucleus and conclude that Po seems crucial for occurrence of SWDs. The dentate nucleus sends inhibitory projections to thalamic nuclei and mediates the information flow to the cortex. The results of the present study showed the absence of dento-Po thalamic connections in GAERS, which may play an important role in the occurrence of SWDs. Furthermore, the Po thalamic nucleus is a higher order nucleus that is thought to be involved in corticocortical communication (Sherman and Guillery, 2009) and can play a role in generalization of the SWD activity.
Direct thalamic and cerebellar electrostimulation has been examined as a potential therapeutic modality for human epilepsy (Shenoy et al., 2016). The CM nucleus of the thalamus is a stimulation target because of its diffuse projections to the cerebral cortex. The electroencephalogram (EEG) evidence in humans suggests that CM participates in propagation of absence and other generalized seizure activity (Velasco et al., 2000). The results of the present study showed that in both GAERS and Wistar animals, the dentate nucleus was bilaterally connected to the CM thalamic nucleus. Bilateral connection of the dentate nucleus with the CM thalamic nucleus may contribute in propagation of seizures to the cerebral cortex during epileptic seizures.
Although the mechanism is still not clear, the interaction between cerebellum and epilepsy is well known. The cerebellum is of interest in epilepsy because of the increased incidence of cerebellar atrophy in patients with partial and generalized epilepsy (Crooks and Mitchell, 2000; Gessaga and Urich, 1985). Both the increased likelihood of cerebellar atrophy in epileptic patients and the potential for cerebellar stimulation to decrease seizure frequency imply an important pathophysiologic role. Thus, the dentate nucleus for these reasons represents potential therapeutic targets that must be aggressively and thoroughly investigated.
Absence epilepsy is not a focal disorder, but rather a brain network disease. Thus, we propose that anatomical structures that have connections with specific thalamic nuclei may be involved in the epileptic mechanism and can affect the thalamo-cortical circuit. The dentate nucleus has strong connections with the thalamus and cerebral cortex, it seems quite plausible that it may have epileptogenic potential and must not be ignored. Thus, abnormal dentate input to the thalamus can affect a wide range of thalamo-cortical networks and functions. What role the cerebellum plays remains to be elucidated.
Conclusion
The results of this study show weak/deficit dento-thalamic connections in GAERS compared with Wistar animals. These results may help to better understand the pathophysiology of absence epilepsy.
Footnotes
Acknowledgment
The authors would like to thank the Koç University Research Center for Translational Medicine (KUTTAM) for use of their facilities.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This research did not receive any specific grant from funding agencies in the public, commercial, or not-for-profit sectors.