
Abstract
Evolutionary-developmental theories consider the evolved mechanisms underlying adaptive behavioral strategies shaped in response to early environmental cues. Identifying neural mechanisms mediating processes of conditional adaptation in humans is an active area of research. Resting-state functional magnetic resonance imaging (RS-fMRI) captures functional connectivity theorized to represent the underlying functional architecture of the brain. This allows for investigating how underlying functional brain connections are related to early experiences during development, as well as current traits and behaviors. This review explores the potential of RS-fMRI of the amygdala (AMY) for advancing research on the neural mechanisms underlying adaptive strategies developed in early adverse environments. RS-fMRI studies of early life stress (ELS) and AMY functional connectivity within the frame of evolutionary theories are reviewed, specifically regarding the development of self-regulatory strategies. The potential of RS-fMRI for investigating the effects of ELS on developmental trajectories of self-regulation is discussed.
Introduction
Evolutionary-developmental theories consider the evolved mechanisms underlying adaptive behavioral strategies that are shaped in response to early environmental cues (i.e., experiences) (Belsky et al., 1991; Del Giudice et al., 2011; Figueredo et al., 2006; Geary and Bjorklund, 2000). Due to developmental plasticity in childhood, these cues shape development through a process of conditional adaptation, resulting in developmental trajectories that are dependent, in part, on conditions of the immediate environment (Belsky et al., 1991). Exposing the neural mechanisms mediating processes of conditional adaptation in humans is an active area of research (Del Giudice et al., 2011).
Rapidly advancing methodological techniques in resting-state functional magnetic resonance imaging (RS-fMRI) provide a means for measuring individual differences in brain circuitry (Thomason et al., 2011) and hold great promise for elucidating the neural mechanisms of conditional adaptation. RS-fMRI allows for measurement of functional connectivity between brain regions via low-frequency blood-oxygen-level-dependent (BOLD) signals (Biswal et al., 1995). Resting-state functional connectivity (RS-fc) is measurable independent of task paradigms, further allowing for examination of functional connectivity that is not driven by the immediate context but rather represents the underlying functional architecture of the brain (Biswal et al., 1995; Lee et al., 2013). This allows researchers to investigate how underlying functional connections relate to early experiences during development, such as early life stress (ELS; i.e., stress caused by trauma during early childhood), as well as current traits and behaviors.
This article explores the potential of RS-fMRI for advancing research investigating the neural mechanisms underlying adaptive strategies developed in adverse environments. To illustrate this potential, current RS-fMRI research on amygdala (AMY) functional connectivity in individuals with a history of ELS is reviewed in relation to current evolutionary-developmental hypotheses on the development of individual adaptive self-regulatory strategies.
Self-regulation involves the control of emotion, attention, and behavior in the face of competing stimuli and is particularly salient in complex social structures (Barkley, 2001). A long line of neurobiological research on the AMY indicates its involvement in fear conditioning, reward/punishment processing, motivation, addiction, emotional memory, and social cognition (Miskovic and Schmidt, 2012; RajMohan and Mohandas, 2007). These processes involve variations in self-regulation. Further, AMY dysfunction is implicated in autism, attention-deficit/hyperactivity disorder, dementia, affective disorders, and anxiety disorders (Miskovic and Schmidt, 2012; RajMohan and Mohandas, 2007), also representing variations in self-regulatory processes. It is this long line of research that implicates the AMY as primary for aversive learning (i.e., in the context of ELS), particularly during childhood (Gee et al., 2018). As such, this review article focuses on RS-fMRI research, specifically of the AMY and its potential role in adaptive self-regulatory strategies.
A brief overview of the corticolimbic system
The corticolimbic system comprises several brain regions that are involved in the processing of emotional stimuli: the AMY and hippocampus from the limbic system, the prefrontal cortex (PFC), and anterior cingulate cortex (ACC). The AMY plays a central role in the limbic system involving bottom-up emotion processing and regulation in response to socially or emotionally salient cues (Miskovic and Schmidt, 2012; Swartz and Monk, 2014). The AMY has bidirectional connections with the PFC, such as the lateral and medial PFC that are involved in the top-down regulatory processing of and response to stimuli (Miskovic and Schmidt, 2012; Ochsner and Gross, 2007). These bidirectional connections allow for information sharing between the AMY and PFC and the potential for both up- and downregulation of attention, emotion, and behavioral response in the face of salient stimuli [for more details about these processes, see Ochsner and Gross (2007)].
The ACC has connections to both the limbic system (e.g., AMY) and the PFC. Thus, the ACC has a role in mediating cognitive influences on emotion (e.g., regulation) (Benes, 2007). The hippocampus works with the AMY in aspects of learning and memory associated with emotional stimuli (Benes, 2007). In addition, although not typically included in the list of core structures of the corticolimbic system, the posterior cingulate cortex (PCC) is a relatively highly connected paralimbic structure that serves as a hub for information processing (Leech and Sharp, 2014). The PCC is involved in both internal and external focus of attention, particularly to emotional stimuli, such as threat (Leech and Sharp, 2014), and may play an important role in self-regulatory strategies.
The evolution of adaptive self-regulatory strategies
From an evolutionary standpoint, the ability to self-regulate emotion, attention, and behavior is especially important for the human species that evolved to rely on complex societal cooperative relationships and exchanges (Barkley, 2001; MacDonald, 2008). Self-regulation requires self-awareness, understanding societal norms, threat detection, and inhibiting one's prepotent response for another more desirable response (Barkley, 2001; Del Giudice, 2015; Heatherton, 2011; MacDonald, 2008). The umbrella of adaptive self-regulatory strategies (e.g., self-control, effortful control, executive functions, inhibition) has received much attention across academic disciplines (e.g., human development, psychology, education) (Blair, 2010; Diamond, 2006; Rothbart and Bates, 2006; Rothbart and Rueda, 2005; Zhou et al., 2012).
Evolutionary-developmental research concerns the study of adaptive strategies that develop within a specific environmental context (Geary and Bjorklund, 2000). Evolutionary-developmental theories [e.g., evolutionary theory of socialization (Belsky et al., 1991), adaptive calibration model (Del Giudice et al., 2011)] propose that early experiences serve as cues that shape developmental trajectories to best match the individual to the anticipated adult environment, thus maximizing the individual's outcomes given specific environmental opportunities and constraints. Indeed, much research indicates that individuals are differentially susceptible to the influence of early experiences (e.g., ELS) on specific aspects of development (e.g., self-regulation) (Belsky and Pluess, 2009; Boyce and Ellis, 2005).
Based on Belsky and colleagues' (1991) evolutionary theory of socialization, early environmental cues and experiences guide the development of particular strategies (i.e., life history strategies) (Ellis et al., 2009) and corresponding behaviors that have reliably led to adaptive outcomes for the species in a given context. Thus, no set of strategies is “best,” rather different strategies are adaptive or maladaptive depending on whether they match the environment in adulthood (Ellis et al., 2009). In regard to self-regulation, either higher or lower levels of self-regulatory behavior (e.g., inhibition) may be best fit for the environment depending on expected levels of adversity (Blair, 2010; Del Giudice, 2015; MacDonald, 2008).
ELS is biologically embedded to guide stress responsivity
Based on the adaptive calibration model (Del Giudice et al., 2011), early experiences calibrate the stress response system (SRS). Put simply, information gained from early environmental experiences are encoded in the SRS via neuroendocrine systems (e.g., hypothalamic pituitary adrenal [HPA] axis). This information is received by the SRS through limbic structures such as the AMY. Following this model, calibration subsequently mediates the individual's adaptive patterns of responsivity and behavior (e.g., to threatening stimuli) that are specific to the individual's expected environment. Within the SRS it is the reciprocal activation of the parasympathetic and sympathetic nervous system that is the basis of self-regulating one's responses.
Responsivity is further mediated by the likewise calibration of the HPA axis that is centrally controlled by limbic structures, especially the AMY. The HPA axis mounts a delayed response to stressful stimuli via release of cortisol. Chronic early stress, such as recurring experiences with ELS, results in sustained cortisol elevation (Del Giudice et al., 2011). Elevated cortisol prepares the individual for stressful events, such as elevating heartrate. However, repeated stressful exposures early in life often result in a blunted cortisol response (Del Giudice et al., 2011). Although a blunted cortisol response is often considered maladaptive or an indication of dysregulation, it may be an adaptive trade-off in an anticipated adverse environment (Del Giudice et al., 2011).
ELS shapes trajectories of self-regulation toward higher impulsivity
When considering the conditional adaptation of self-regulation, developmental trajectories shaped by early experiences equip an individual with cognitive-behavioral strategies that best fit the expected adult environment given available resources (Figueredo et al., 2006). Although high levels of self-regulation may be culturally regarded as ideal and leading to the “best” outcomes, there are environments (e.g., those high in adversity) where high self-regulation may be less adaptive or even maladaptive despite trade-offs (e.g., poorer school performance) (Blair, 2010; Del Giudice, 2015; MacDonald, 2008).
Self-regulation refers to the future-oriented processes that allow an individual to manage goal-directed behaviors over time and across changing contexts (Karoly, 1993). Future-oriented self-regulation is likely adaptive in a safe, predictable environment where life expectancy is long and future resources and opportunities are more plentiful and predictable. However, in an adverse environment (i.e., shorter life span, limited resources and opportunities), a proclivity toward higher impulsivity (i.e., reflexivity) versus self-regulation may be more adaptive (Blair, 2010; Del Giudice, 2015; MacDonald, 2008). The inclination to be impulsive (i.e., behavioral disinhibition and lack of future orientation) (Ross and Hill, 2002) may allow an individual to take the necessary risks to better compete for and secure limited resources as they become available (Blair, 2010).
Survival is an immediate necessity for future successful reproduction. Thus, the ability to both detect and appropriately respond to threat should be an adaptive priority. The ability to detect social threat is important for survival and reproduction (Barkley, 2001; Del Giudice, 2014; Del Giudice et al., 2011; MacDonald and Leary, 2005; Miskovic and Schmidt, 2012). The AMY plays a prominent role in threat detection and response [for review see MacDonald (2008), Miskovic and Schmidt (2012)]. Although individuals exhibit both top-down (i.e., explicit) and bottom-up (i.e., implicit) strategies in response to threatening stimuli [see also feedback and feedforward control in Del Giudice (2015)], individual variation exists in proclivity toward one strategy over another, coupled with proximal contextual factors (Del Giudice, 2015).
For individuals reared in harsh and unpredictable environments, threat detection abilities should be particularly salient, resulting in developmental trajectories that prioritize reflexive fear processing involving the bottom-up processes of salient threat cues (Del Giudice, 2015; MacDonald, 2008). In bottom-up reflexive fear processing, salient threat information is passed directly to the AMY, and it thus allows for the ability to react immediately, or without inhibition (MacDonald, 2008). For individuals reared in safer, more predictable early environments, the use of complex top-down approaches to fear processing may carry overall less risk (e.g., consequences of overreacting to a nonthreatening situation) and be more adaptive (Blair, 2010; Del Giudice, 2015). Prioritizing a top-down strategy allows the individual to take in and process information (via the prefrontal brain regions), and then strategically enact defensive or inhibitory processes as needed (Del Giudice, 2015; MacDonald, 2008). Therefore, based on adaptive strategies that are context dependent in their development, individuals whose early experience (such as with ELS) indicates a harsh anticipated adult environment are expected to have increased bottom-up (i.e., reactive) threat responses and this should be reflected in their neural profile (Blair, 2010).
In this article, self-regulatory behaviors are conceptualized along a continuum with impulsivity at the lowest, more reflexive end, and higher future orientation on the opposite end. Impulsivity can be seen as a strategy involving high risk with high potential benefits (MacDonald, 2008). Thus, in this article, impulsivity is considered along the continuum of self-regulatory strategies that minimize inhibition of behaviors, favor bigger immediate rewards (i.e., a present orientation to rewards and behaviors), and are generally adaptive in adverse environments.
Need for neuroimaging in evolutionary-developmental research
Methodologies in evolutionary-framed development research on adaptive strategies range from self-report inventories (e.g., ELS, reproductive histories, attachment histories) and measures of genetic polymorphisms and endocrine hormones to cross-species and cross-cultural comparisons and computer simulations (Schmitt and Pilcher, 2004). Progress in identifying the underlying physiological mechanisms of stress responsivity in early development [e.g., adaptive calibration model (Del Giudice et al., 2011), consilience model (Figueredo et al., 2006)] reinforces evolutionary-developmental research by pointing to rigorous methodological pathways for integrating evolutionary and developmental biology (Del Giudice et al., 2011). Stimulated by comparative animal research (Meaney, 2001; Meaney, 2010), the exploration of neural mechanisms of conditional adaptation in humans is now of vital interest (Del Giudice et al., 2011).
RS-fMRI offers a look into how brain regions are functionally connected devoid of response-inducing stimuli, and it represents an individual's intrinsic (i.e., underlying) functional organization of brain networks (Fox et al., 2005). Thus, RS-fMRI can provide a picture of the underlying functional relationships between regions of the brain in relation to ELS experiences. Brain imaging during a resting state is a noninvasive, methodologically simpler approach as compared with task-based fMRI (Lee et al., 2013). RS-fMRI studies have been shown to afford reliable and replicable results (Chou et al., 2012), indicating that RS-fMRI is a viable methodological approach for longitudinal research on development with infants and young children (Graham et al., 2015b; Lee et al., 2013; Shen, 2015).
The AMY is documented as having multifaceted roles, including control of attention such as in threat detection and processing, as well as emotional and behavioral inhibition (Miskovic and Schmidt, 2012), and therefore is key to understanding individual differences in adaptive self-regulatory strategies. Thus, RS-fMRI of the AMY may serve to inform evolutionary-developmental research about the underlying neural functional connections that correlate with adaptive self-regulatory strategies (e.g., impulsivity) shaped by early adversity. Currently, there are only preliminary attempts to explicitly link RS-fMRI methodology to evolutionary theoretically framed research endeavors (Ferrarini et al., 2009; Madeo et al., 2017; Santarnecchi et al., 2015; Thomas Yeo et al., 2011; Watanabe et al., 2014). Therefore, this article explores more fully and systematically the untapped potential for the joining of RS-fMRI with evolutionary-developmental investigations to stimulate new avenues of development research.
Materials and Methods
A search using Web of Science Core Collection, PsycINFO, PubMed, and MEDLINE was conducted to identify empirical articles from inception through April 2019 examining AMY functional connectivity measured by RS-fMRI in relation to ELS. This search was conducted by using combinations of the terms “resting state,” “intrinsic functional connectivity,” “early life stress,” “early trauma,” or “childhood trauma,” and “amygdala.”
Prospective articles were independently judged and first screened for inclusion based on their title and abstract. Screening criteria included (1) being an empirical article, (2) use of RS-fMRI methods, (3) including measurement of ELS, and (4) reporting of statistically significant findings regarding AMY RS-fc. Studies were not otherwise limited by study design or study sample. Studies that obviously did not meet the criteria based on title and abstract were excluded; all other studies were either marked for inclusion or further assessed by review of the article in its entirety. Additional studies listed as references in the included studies were likewise screened and marked for inclusion if they met the criteria. A PRISMA flow diagram depicting the search process and the number of studies identified and included in this review is shown in Figure 1.
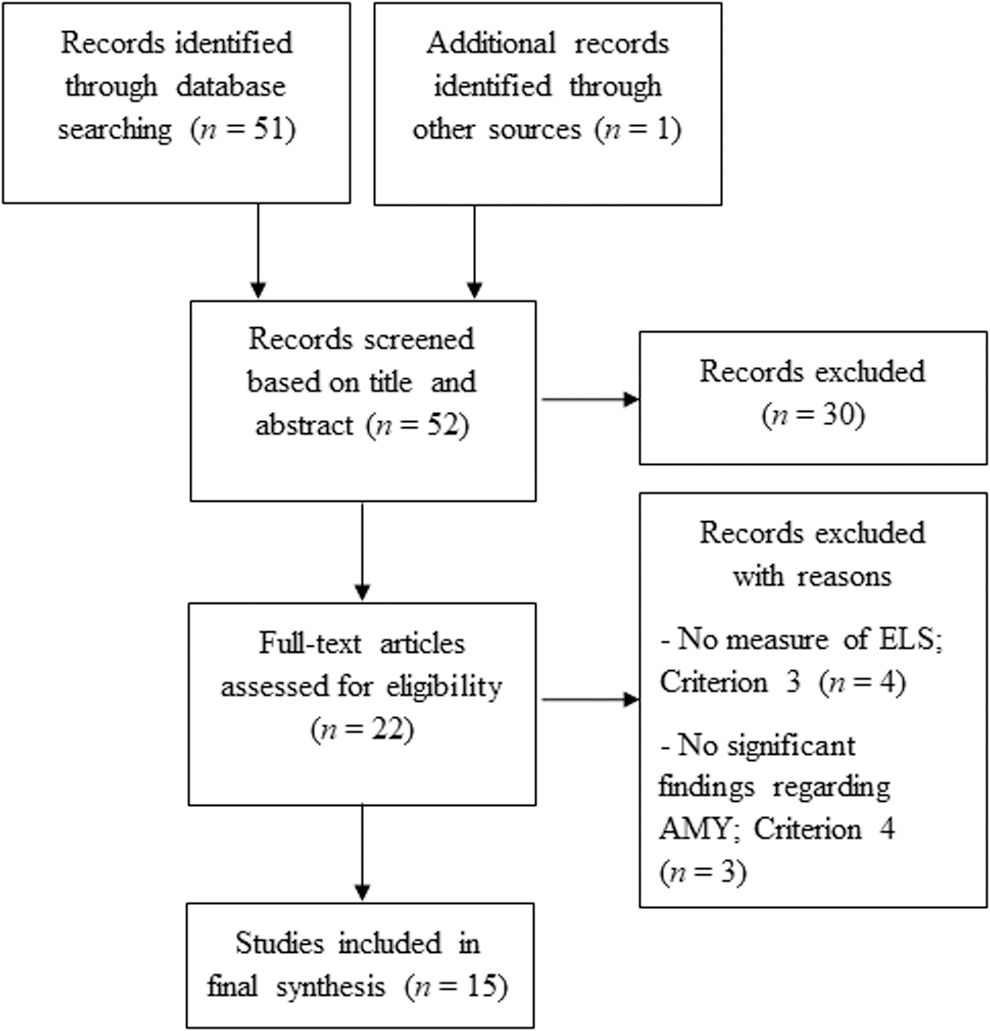
PRISMA flow diagram depicting number of studies identified and included in review. AMY, amygdala; ELS, early life stress.
Results and Discussion
Search results
Although one article (Vaidya and Gordon, 2013) was excluded due to not meeting criterion 1, it did lead to the identification of an additional article (Nooner et al., 2013). In sum, 15 articles remained eligible for review and are considered next in the context of evolutionary-developmental theories. An overview of each reviewed study describing the study sample, ELS measurement, RS-fMRI methodology, and findings related to AMY functional connectivity is presented in Table 1. To summarize, two studies measured ELS during prenatal/infancy, one during childhood, four during adolescence, and seven retrospectively measured ELS in adulthood. Two included studies conducted RS-fMRI during infancy, one during childhood, five during adolescence, and the remaining seven conducted RS-fMRI during adulthood. All studies used seed-based RS-fMRI analyses except for two that used graph analysis (Cisler, 2017; Cisler et al., 2013), and Du and colleagues (2016) who used amplitude of low-frequency fluctuation (ALFF) and fractional ALFF (fALFF).
Overview of Reviewed Studies
ACC, anterior cingulate cortex; ALFF/fALFF, amplitude of low-frequency fluctuation/fractional ALFF; AMY, amygdala; bl, basolateral; cm, centromedial; d, dorsal; decr., decreased; dl, dorsolateral; dm, dorsomedial; ELS, early life stress; GPS, genetic profile score; incr., increased; L, left; m, medial; MDD, major depressive disorder; OXY, oxytocin; PCC, posterior cingulate cortex; PFC, prefrontal cortex; pg, pregenual; r, rostral; R, right; RS-fc, resting-state functional connectivity; RS-fMRI, resting-state functional magnetic resonance imaging; SD, standard deviation; sf, superficial; sg, subgenual; vl, ventrolateral; vm, ventromedial.
Functional connectivity analyses applied in the included studies
A number of methods have been applied to estimate brain functional connectivity. Briefly, seed-based analysis, which is the most commonly used approach, calculates the correlation of time series between a seed voxel (region) and all other voxels in the brain (Biswal et al., 1995). Although static RS-fc is considered a traditional measure of functional connectivity assuming that functional networks are statics in time, dynamic RS-fc attempts to capture dynamic changes in connectivity between distinct regions (i.e., that may be driven by contextual factors) over a specific period (e.g., sliding window) (Hutchison et al., 2013). Studies measuring static RS-fc measure correlation between specific brain regions at a specific point in time whereas dynamic RS-fc measures connectivity (i.e., correlations between regions) at each sliding window.
Positive correlations reflect synchronized activity between these regions; negative correlations, also called anti-correlation, refer to a negative Pearson cross-correlation coefficient between brain regions. Importantly, reported increases in RS-fc refer to correlations moving further away from zero (i.e., no correlation), and therefore refer to either increasingly positive or increasingly negative correlation (i.e., more anti-correlation) (Vaidya and Gordon, 2013), with the opposite being true of reported decreases in RS-fc. Unfortunately, the distinction between whether the increase or decrease is in a positive or negative direction is not reported consistently. Increases in dynamic RS-fc indicate more fluctuations in connectivity between regions whereas decreases indicate less fluctuation.
Graph theory analysis provides a framework to investigate the topological organization (e.g., small-word, hub, efficiency) of brain connectivity (Rubinov and Sporns, 2010), including network structure. ALFF (Zang et al., 2007) and fALFF (Zou et al., 2008) are related measures that quantify the amplitude of slow fluctuations in RS-fMRI signals. Additional details regarding these RS-fMRI analyses are documented elsewhere [e.g., Vaidya and Gordon (2013)].
Overview of results
This review aims at exploring the potential for RS-fMRI in advancing evolutionary-developmental research by elucidating the neural mechanisms underlying individual variations in adaptive strategies of self-regulation shaped by adverse environments. The collection of 15 included articles are reviewed in turn under the two previously discussed evolutionary-developmental hypotheses of adaptive self-regulatory strategies: (1) ELS is biologically embedded, and (2) ELS shapes developmental trajectories of self-regulation toward higher impulsivity. Note that some of the reviewed articles speak to both hypotheses and, therefore, are included under both subheadings.
Resting-state fMRI and biologically embedded ELS
Evolutionary-developmental theories hypothesize that early experiences serve to calibrate the SRS to guide stress responsivity in ways aimed at best matching the individual to the expected adult environment. RS-fMRI research investigating brain functional connectivity developed in the context of ELS can assist in identifying the neuroendocrine mechanisms that are involved in this process. The AMY is particularly involved not only in fear response but also specifically in the processing of and emotional regulation in response to social threat (Miskovic and Schmidt, 2012; RajMohan and Mohandas, 2007), critical strategies in a social, cooperative species. Four studies that specifically included biological measures (e.g., cortisol, oxytocin, or genetic profiles) to investigate neuroendocrine mechanisms encoding the SRS are discussed next.
Burghy and colleagues (2012) conducted a prospective longitudinal study of 57 male and female adolescents investigating the associations between ELS in infancy, neuroendocrine function in childhood, and RS-fc measured in adolescence. ELS was used to predict AMY-PFC RS-fc in adolescence. This study considered the role of the HPA axis by including a measure of childhood basal cortisol as a potential mediator of adolescent AMY-PFC RS-fc. Results indicated that for females only, higher ELS in infancy significantly predicted increased basal cortisol levels in childhood, which then predicted decreased left AMY-PFC RS-fc in adolescence. In other words, results supported their hypothesis that basal cortisol levels in childhood mediate the relationship between ELS and decreased AMY-PFC RS-fc in adolescence, but only for the left AMY and only for females.
Burghy and colleagues connect these sexually dimorphic findings with previous work [e.g., Weinstock (2007)], which suggests that females may have heightened sensitivity to the effects of ELS on neuroendocrine function, resulting in more robust epigenetic change. Differential effects of ELS for males and females are not entirely surprising given that evolved reproductive strategies, which become salient during and after puberty (i.e., adolescence), are different for males and females (e.g., sex-differentiated mate selection pressures and levels of parental investment) (Del Giudice et al., 2011; Figueredo et al., 2006; MacDonald, 2008) based on multiple environmental factors.
The second study by Kaiser and colleagues (2017) also included basal cortisol measurement in the investigation of AMY-corticolimbic RS-fc for 70 females with a range of threat-related ELS experiences (e.g., peer aggression, abuse) and severity. A particular methodological contribution by the study is that both static and dynamic RS-fc were measured, thus investigating potential for “intrinsic flexibility” of AMY-corticolimbic relationships in relation to ELS. Kaiser and colleagues measured functional connectivity across sliding windows (i.e., sequential temporal segments) to create connectivity maps for each participant. They then used the standard deviation across sliding windows for each participant for analysis. Results indicated that threat-related ELS severity for females was associated with increased negative static RS-fc between the AMY and the left dorsolateral PFC, and increased dynamic RS-fc between AMY and rostral ACC. Both the PFC and ACC are implicated in top-down regulation of limbic structures.
Further, decreased static AMY-dorsolateral PFC RS-fc was associated with blunted cortisol response to stress in adulthood, which was moderated by dynamic AMY-rostral ACC RS-fc. In other words, females with decreased static RS-fc of a downregulating self-regulatory system (i.e., AMY-PFC) and more dynamic RS-fc of another downregulating system (i.e., AMY-rostral ACC) exhibited a less blunted cortisol response to stress in adulthood. Kaiser and colleagues raise the possibility that “intrinsic flexibility” of the AMY-rostral ACC indicates a compensatory mechanism for individuals who have blunted cortisol response due to severe ELS. Kaiser and colleagues' findings have potential for RS-fMRI methodology to further test evolutionary-developmental theories explaining individual differences to adverse environments [e.g., biological sensitivity to context theory (Belsky and Pluess, 2009; Boyce and Ellis, 2005), bet-hedging hypotheses (Del Giudice, 2015; Ellis et al., 2009)].
The third study by Fan and colleagues (2014) used a measure of oxytocin in their all-male sample of 18 individuals investigating AMY-prefrontal RS-fc and ELS. In their randomized within-subject design, males were administered either oxytocin or a placebo 45 min before RS-fMRI. This was followed by a psychosocial stress task administered during a final task-based fMRI to investigate whether ELS-related AMY-prefrontal RS-fc predicts state anxiety in response to psychosocial stress. Though there was neither a significant direct effect of oxytocin nor an interaction of oxytocin and ELS on AMY RS-fc, ELS did significantly moderate the association between oxytocin and an AMY-pregenual ACC rest
Oxytocin is described as the “tend-and-befriend” stress response hormone that is sex differentiated and guides females toward caretaking and support-seeking behaviors in the face of stress (Taylor et al., 2000). Cortisol ramps up stress response, whereas oxytocin often attenuates it, and androgens (i.e., male sex hormones) may inhibit oxytocin production under stress (Taylor et al., 2000). Decreased sensitivity to oxytocin for males in a harsh environment could be adaptive (Meaney, 2001), particularly regarding evolved sex-differentiated roles for males involving detection and response to threat and their typical “flight or fight” stress response (Del Giudice et al., 2011).
The fourth study that contributes to the discussion of the development of the HPA axis within adverse early environments is that by Pagliaccio and colleagues (2015). Researchers conducted a study of 120 school-age male and female children examining HPA axis genetic variation, ELS, AMY RS-fc, and emotion regulation (an aspect of self-regulation). Here, genetic risk profile scores reflected 10 single nucleotide polymorphisms within four HPA axis genes (i.e., CRHR1, NR3C2, NR3C1, and FKBP5) that are linked to depression, anxiety, and posttraumatic stress disorder. Regarding biological embedding of ELS, the study results showed that ELS and HPA genetic-risk profiles interacted to predict decreased negative left AMY RS-fc with the parahippocampal gyrus, caudate tail, and two regions of the left PFC (inferior frontal gyrus and middle frontal gyrus). In other words, increased experiences of ELS coupled with higher genetic risk profiles of stress-related psychopathology were associated with unilateral weakened RS-fc of the AMY with corticolimbic brain regions in childhood.
The potential interactions of genotypic variation with early environmental cues (i.e., gene X environment interactions) are foundational to current evolutionary-developmental theories (e.g., biological sensitivity to context theory) (Belsky and Pluess, 2009; Boyce and Ellis, 2005). Thus, coupling RS-fMRI research using both gene and environment aspects with an evolutionary-developmental framework can help further discern underlying neuroendocrine mechanisms of individual differences in adaptive strategies.
As a whole, the four reviewed studies demonstrate the explanatory potential of RS-fMRI within evolutionary-developmental research. Specifically, RS-fMRI allows for the investigation of neuroendocrine mechanisms encoding the SRS for individuals with (and without) ELS. Use of evolutionary theoretically framed models (e.g., adaptive calibration model) (Del Giudice et al., 2011) joined with RS-fMRI can elucidate the underlying neuroendocrine mechanisms of adaptive self-regulatory strategies and stress responsivity that best match the individual to the environment. In addition, hypotheses on the sex-differentiated roles of males and females can provide a theoretical framework to help explain emerging evidence of differences in males' and females' neural profiles related to stress-response calibration in adverse contexts.
Resting-state fMRI and trajectories toward higher impulsivity
Evolutionary-developmental theories predict that early adverse environments direct development toward higher impulsivity and a more reflexive approach to threat (Blair, 2010; Del Giudice, 2015; MacDonald, 2008). This reflexive approach (e.g., lower self-regulation) would allow individuals to better compete for and secure scarce resources and take advantage of fleeting opportunities (Blair, 2010). The functional connectivity of the AMY may be central to bottom-up versus top-down regulatory approaches to environmental stimuli (Miskovic and Schmidt, 2012). In this review of RS-fMRI literature, all 15 studies illustrate how neuroimaging during a resting state can support evolutionary-developmental research on adaptive self-regulatory strategies in adverse contexts.
Significant individual differences in AMY network topology
Findings from Cisler and colleagues' (2013) exploratory study of 38 women speak to overall AMY network topology. This study investigated differential RS-fc with a defined 21-node emotion processing/regulation network, including the right and left AMY, among resilient (i.e., no depressogenic effects) and susceptible individuals (i.e., reported depressogenic effects) in relation to ELS and a control group. In a between-group comparison, RS-fMRI of susceptible individuals showed significant differences in left AMY network topology within the emotion processing/regulation network: decreased local connectivity (i.e., efficiency) as compared with the control group, and increased hub-like properties (i.e., betweenness centrality) as compared with the resilient group.
Node efficiency indicates the ability of nodes to spread information to other nodes within a network. Decreased efficiency of the AMY, therefore, indicates that ELS for susceptible individuals may decrease other network nodes' ability to communicate with the AMY. Betweenness centrality also speaks to a node's role in the network in that it indicates the potential influence the node holds over the flow of information between other nodes in the network. In Cisler and colleagues' study, ELS for susceptible individuals was associated with an increase in the left AMY's potential influence on the flow of information and thus its role as a hub in the network. Cisler and colleagues explain that more “stable” networks are considered those in which information is distributed over multiple nodes versus relying on one hub.
Further, lowered efficiency suggests that information is less efficiently communicated to the left AMY, as it has to navigate through more paths to reach it. In contrast, one previous study by Ohashi and colleagues (2019) using diffusion tensor imaging to measure structural connectivity of the AMY in relation to ELS suggested lower nodal efficiency of the right AMY for resilient men and women compared with susceptible and control adults. Hence, more research is needed in this area to better understand the impact of ELS on AMY efficiency for susceptible and resilient individuals, and differing effects for men and women.
An additional study by Cisler (2017) further investigated whether the relationship between ELS and decreased AMY-medial PFC functional connectivity reported across studies [e.g., Birn et al. (2014), Burghy et al. (2012), El Khawli et al. (2018), Souza-Queiroz et al. (2016)] actually represents larger inter-network differences, specifically between the limbic and default mode networks. The default mode network consists of highly correlated brain regions, including the PCC and medial PFC; it is active during resting state and has often been shown to be deactivated during tasks (Greicius et al., 2003). Cisler's study included 26 female adolescents who had prior experiences with assault (i.e., physical and/or sexual assault) and 30 controls. Cisler's analyses demonstrated a negative correlation between ELS (specifically emotional abuse) and RS-fc of the left AMY and medial PFC.
Further, graph analyses indicated a significant positive correlation between emotional abuse and network modularity (i.e., the degree to which nodes in a network have denser intra-module connectivity but sparser inter-module connectivity) (Wang et al., 2010), suggesting that increased experiences with emotional abuse ELS correspond with increased network modularity of the limbic and default mode network hubs, and thus sparser links between these networks. Finally, Cisler and colleagues compared dynamic RS-fc (i.e., functional connectivity measured at each sliding window) for individuals with and without experiences of assault. Findings confirmed decreased positive correlation between dynamic RS-fc of the left AMY and medial PFC with the dynamic RS-fc of the overall limbic and default networks for the group of adolescents who experienced assault. Thus, experiences with ELS that correlate with decreased AMY-medial PFC RS-fc may represent overall greater network partitioning between the limbic and default mode networks (Cisler, 2017). Cisler's study demonstrates the benefits of including dynamic RS-fc in RS-fMRI studies, and it provides further insight on the underlying neural mechanisms linking ELS with decreased top-down self-regulatory processes.
An additional study by Du and colleagues (2016) contributes to understanding how ELS is associated with overall AMY network topology for individuals with major depressive disorder. In their study of 18 males and females with major depressive disorder and 18 healthy controls, Du and others measured ALFF and fALFF. Overall, individuals with major depressive disorder also reported higher ELS. With regards to the AMY, patients with major depressive disorder showed higher ALFF (i.e., higher intensity of spontaneous activity) in both the right and left AMY compared with healthy controls. Further, these differences persisted even when controlling for ELS. These results suggest that ELS serves as a confounding factor in fMRI studies investigating individuals with major depressive disorder.
In sum, differences in overall AMY network topology in the context of ELS provide preliminary indication of underlying neural mechanisms directing susceptible individuals toward more impulsive (i.e., bottom-up, reflexive) self-regulatory processes. For example, decreased efficiency of the left AMY and decreased network modularity between the limbic and default mode networks may serve to segregate the AMY from conflicting influence from other brain regions (e.g., regulatory effects of the PFC) when faced with recurrent threat, allowing the individual to remain reactive and perhaps better employ protective behaviors.
Also, with increased hub-like properties, the left AMY may have the upper hand at exerting influence on information passed through the network and thus biasing behavior responses. Increased AMY influence may be more adaptive and hence preferable for susceptible individuals reared in stressful environments given the probability that their environment in adulthood will be comparably stressful and risky. To better understand the implication of these (proposed) adaptive functional connectivity patterns, RS-fMRI research coupled with task-based fMRI measures is needed to further elucidate how these intrinsic patterns of unilateral connectivity relate to ecologically relevant scenarios of threat in harsh and unpredictable environments.
Individual differences in the corticolimbic system
Using a developmental approach and for ease of presentation, the reviewed studies regarding the corticolimbic system are grouped by age at RS-fMRI.
Infancy
Scheinost and colleagues (2016) present a study of ELS experienced in utero as measured by prenatal exposure to maternal stress indicated by physician's diagnosis of mother's depression or anxiety during pregnancy. In their study of 26 extremely premature male and female neonates (born at less than 28 weeks of gestation), Scheinost and collaborators investigated the association of ELS with AMY RS-fc. In their group comparison model, preterm neonates with ELS showed decreased RS-fc of the left AMY with the thalamus and hypothalamus (as well as the brainstem and fusiform) as compared with preterm neonates without ELS. Further, the effect of ELS on AMY RS-fc added to the influence of gestational age at birth. In short, although individuals born premature exhibited decreased RS-fc of the left AMY with other limbic structures, experiences with ELS during gestation exacerbated this effect.
A second study with RS-fMRI measured in infancy was conducted by Graham and collaborators (2015a). The study included a sample of 23 male and female infants in a seed analysis of RS-fMRI data collected during natural sleep. This study reported on RS-fc between the AMY and a paralimbic structure, the PCC. Results showed that infant right AMY-PCC RS-fc was increased when associated with a common form of ELS, nonviolent interparental conflict. Because the PCC plays a role in both internal and external focus of attention, including vigilant attention maintenance (Leech and Sharp, 2014), the study results show how adaptive self-regulatory mechanisms of attention are affected by early experiences. Changes to AMY-PCC RS-fc in the presence of parental conflict may be an indication of the heightened environmental sensitivity during the earliest years of life (i.e., evolutionary theory of socialization) (Belsky et al., 1991).
Childhood
One study by Park and collaborators (2018) used RS-fMRI with 79 male and female children ages 4–8 years to investigate whether the highly reported relationship between ELS and decreased RS-fc of the AMY and PFC [e.g., Birn et al. (2014), Burghy et al. (2012), Cisler (2017), El Khawli et al. (2018), Fan et al. (2014), Kaiser et al. (2017), Pagliaccio et al. (2015)] is evident during childhood. Not only did results, in fact, indicate that ELS was associated with less RS-fc between the bilateral AMY and medial PFC, but also AMY-medial PFC RS-fc was negatively correlated with parent-reported aggressive behavior and attention problems. This study suggests that ELS impacts AMY functional connectivity with PFC early in life and that ELS-related changes to neural profiles and connected self-regulatory-related behaviors are detectable via RS-fMRI in childhood.
Adolescence
One study of interest that measured RS-fMRI during adolescence is that by Burghy and colleagues (2012) (previously introduced in the resting-state fMRI and biologically embedded ELS section). Results indicated that higher ELS during infancy resulted in decreased RS-fc between the left AMY and the ventromedial PFC in adolescence, which was mediated by increased childhood basal cortisol levels for females. The ventromedial PFC is known to be integral in top-down regulation of the AMY such that increased activity is evident in the ventromedial PFC during an individual's self-regulation of emotions (Heatherton, 2011). As such, activity in the ventromedial PFC and AMY is inversely correlated.
Considering Burghy and others' results within an evolutionary-developmental framework, the decreased ventromedial PFC-AMY RS-fc points to less top-down regulation of the AMY by the ventromedial PFC (Heatherton, 2011) and hence neural mechanisms that may be associated with increased impulsivity, a potentially adaptive self-regulatory strategy in the context of adverse environments. However, as discussed earlier, this may be the case only for females.
Pagliaccio and colleagues (2015) (previously introduced in the Resting-state fMRI and biologically embedded ELS section) reported decreased negative RS-fc of the left AMY with ACC for youth who reported more experiences with ELS. AMY-ACC RS-fc was also negatively associated with externalizing behaviors (e.g., acting out, physical aggression) and positively associated with anxiety. The ACC has connections to the prefrontal area and thus has a role in mediating cognitive influences (e.g., regulation) on emotion (Benes, 2007). Therefore, decreased RS-fc between the AMY and ACC may increase emotionally driven behavior (e.g., impulsivity) as a result of decreased top-down self-regulation guided by the PFC (MacDonald, 2008).
Externalizing behaviors correlate with lower self-regulation [for review see Rothbart and Bates (2006)]. Weakened RS-fc between the AMY and ACC could represent the underlying neural mechanisms that shift developmental trajectories toward strategies favoring lower self-regulation and higher impulsivity given anticipated harsh contexts in adulthood. Cluster analyses also indicated an interaction effect such that youth with higher reported ELS and higher genetic risk for depression, anxiety, and posttraumatic stress disorder had weaker RS-fc between the AMY and the right parahippocampal gyrus, caudate tail, and two regions of the left PFC (i.e., middle and inferior frontal gyri). Further, this underlying weakened RS-fc between the AMY and corticolimbic structures correlated with lower emotion regulation and higher anxiety.
A study by Thomason and colleagues (2015) involved 42 male and female youth in an investigation of AMY RS-fc for low-income, trauma-exposed, and non-trauma-exposed children in urban environments. Analyses indicated decreased negative RS-fc, specifically regarding the centromedial (but not basolateral) AMY with the subgenual ACC for trauma-exposed youth. In other words, there was less negative covariance (i.e., anticorrelation) of BOLD activity between these regions. The centromedial AMY is involved in regulating attention and vigilance, whereas the basolateral AMY is associated with associative learning processes [see also Miskovic and Schmidt (2012), Thomason et al. (2015)]. The decreased negative RS-fc between an area of the AMY involved in threat vigilance and the ACC, a mediator between the AMY and PFC, thus may additionally represent the neural mechanisms responsible for increased threat detection and biasing perceptions toward threatening stimuli (Miskovic and Schmidt, 2012).
In addition, youth who had a reported history of trauma had decreased negative RS-fc between the superficial AMY and the anterior insula. The anterior insula has functional connections to brain regions, such as the AMY, that have been implicated in affective evaluation of salient stimuli (Miskovic and Schmidt, 2012). Thomason and colleagues' results support previous research showing increased AMY-insula functional connectivity for individuals with history of trauma (e.g., in adults with posttraumatic stress disorder), and their finding of AMY-insula covariance may indicate a primed response system to detect and react to threats.
Based on a secondary whole brain analysis, Thomason and colleagues (2015) also found decreased positive RS-fc between the superficial AMY and the dorsal ACC (a region of the ACC also involved in sympathetic nervous arousal) (Critchley, 2005), and decreased overall negative RS-fc in the fronto-AMY circuit (i.e., circuitry involving the ACC, orbitofrontal cortex, and the PFC) for individuals with ELS. Thus, these additional findings reinforce the view that ELS is associated with significant changes to the brain circuitry that are essential to threat detection and emotion regulation in the context of adverse environments.
Finally, a study by Nooner and colleagues (2013) also examined AMY RS-fc of 23 male and female youth with trauma exposure. Results showed that increases in ELS were associated with increased RS-fc between the right AMY and a local limbic cluster (i.e., hippocampus, parahippocampal gyrus, and lingual gyrus), along with decreased RS-fc between the left AMY and a long-range frontoparietal cluster (i.e., left middle frontal, inferior frontal, and angular gyri, superior and inferior parietal lobes, and lateral occipital cortex). This pattern of RS-fc suggests that ELS influences developmental trajectories toward more bottom-up approaches of threat detection and response, which is achieved by maximizing unilateral limbic connections and minimizing top-down regulation by prefrontal structures. Nooner and collaborators (2013) also mention that patterns of increased local and decreased long-range connectivity can be associated with “brain immaturity.”
Adulthood
To investigate emotion regulation malleability developed in the context of ELS, El Khawli and colleagues (2018) conducted an RS-fMRI study with 28 male and female adults who reported on their use of reappraisal (i.e., a cognitive therapy used to improve emotion regulation). Seed-based analyses detected an interaction between ELS (specifically emotional abuse) and reappraisal: Individuals with high reported ELS and high use of reappraisal had increased negative RS-fc of the left centromedial AMY with the ventromedial PFC, ventrolateral PFC, parahippocampal regions, and temporal regions, but decreased RS-fc between the right AMY and the right precuneus and temporal regions. However, individuals with low reported ELS and high use of reappraisal had decreased RS-fc of the left centromedial AMY with the left supplementary motor area, right orbitofrontal cortex, and left inferior temporal gyrus, as well as decreased RS-fc of the right centromedial AMY with the bilateral superior temporal cortex, right middle cingulate gyrus, and bilateral supplement motor area.
Thus, results from El Khawli and colleagues indicate that use of reappraisal by individuals with ELS (i.e., ELS × reappraisal) may play a role in the underlying unilateral functional connections of the AMY as seen in RS-fMRI. These results speak to the potential malleability of AMY neurocircuitry in adulthood related specifically to emotional abuse. Further, the associations between the use of reappraisal and RS-fc may have differential effects for the right and left AMY and these differences may be explained by levels of ELS (i.e., high vs. low ELS).
Next, Kaiser and colleagues' (2017) (previously introduced in the resting-state fMRI and biologically embedded ELS section) investigation of AMY RS-fc in the face of ELS used measures of both static and dynamic RS-fc in adulthood. Results indicated that ELS severity was associated with increased negative RS-fc of the AMY with the left dorsolateral PFC, and increased dynamic RS-fc with the rostral ACC. Analyses of moderation indicated a potential compensatory effect such that dynamic AMY-rostral ACC RS-fc attenuated the relationship between ACC-dorsolateral PFC static RS-fc and blunted cortisol response to stress during adulthood. Use of dynamic RS-fc here brought to light additional neural mechanisms not evident through the use of static RS-fMRI alone. Hence, both static and dynamic RS-fMRI should be used to more fully investigate neuroendocrine mechanisms as part of evolutionary-developmental hypotheses of adaptive self-regulation.
Next, Birn and collaborators (2014) explored the relationship between ELS and AMY RS-fc for 27 veterans with combat-related post-traumatic stress disorder (PTSD). Results regarding ELS indicated that ELS positively predicted negative RS-fc between the left and right AMY with the ventromedial PFC, rostral ACC, and dorsomedial PFC (i.e., more negative connectivity). Further, ELS also positively predicted negative RS-fc of the left AMY with the ventrolateral and dorsolateral PFC (i.e., more negative connectivity). Finally, ELS predicted increased RS-fc between the AMY and the cerebellum. Importantly, results suggested that ELS and trauma in adulthood (here measured by combat exposure) differentially predict AMY RS-fc with areas of the brain involved in top-downregulation.
Also, in a study of 18 adult males, Fan and colleagues (2014) investigated ELS as a modulator of AMY-prefrontal RS-fc. Results showed that level of severity of a specific subscale of ELS, emotional abuse, was negatively associated with right AMY RS-fc with prefrontal regions, including both pregenual and subgenual ACC, as well as the dorsal medial PFC. These results suggest that specific types of ELS, namely emotional abuse, and levels of severity have differential effects on unilateral AMY RS-fc, at least for males. In addition, RS-fc between the right AMY and left pregenual ACC predicted state anxiety (often considered self-dysregulation of emotion) in the face of psychosocial stress, which may be the result of a lowered regulatory function of the ACC over the AMY in stressful contexts. Finally, males with higher reported ELS and decreased right AMY-pregenual ACC functional connectivity displayed a decreased sensitivity to oxytocin, an attenuator of psychosocial stress. Once again, these results speak to the underlying neural circuitry related to stress responsivity and self-regulatory strategies, highlighting the potential of RS-fMRI measures in identifying neural mechanisms related to ELS.
Souza-Queiroz and colleagues (2016) investigated the impact of ELS on fronto-limbic RS-fc in a sample of 32 male and female adults with bipolar disorder and 47 healthy adult controls. Here, researchers defined fronto-limbic as including the bilateral AMY, hippocampus, and ventromedial PFC. Results indicated that for bipolar disorder patients only, overall ELS scores were negatively correlated with RS-fc of the AMY with the left ventromedial PFC. Investigation into specific subscores of ELS indicated that this negative relationship was statistically significant for physical neglect and emotional neglect, but not for emotional abuse, physical abuse, or sexual abuse. However, negative relationships between ELS and AMY volume, a measure of structural connectivity, did not vary between bipolar individuals and healthy controls. Further investigations into the relationships between specific aspects of ELS for clinical subpopulations within an evolutionary-developmental theoretical framework may help further explain neural mechanisms related to specific types of ELS and outcomes related to psychopathological diagnoses, which may be adaptive in a given context.
As a whole, the reviewed studies show that RS-fMRI can be a novel and vital component of evolutionary-developmental research by demonstrating how AMY network topology and RS-fc within the corticolimbic system are shaped by ELS. Importantly, and as demonstrated here, this methodology provides an opportunity to examine functional connections within the brain during a resting state across the lifespan. Existing research on AMY RS-fc for individuals with experiences of ELS shows alignment with hypotheses regarding the development of a more reflexive approach (e.g., lower self-regulation) to threat and provides methods for uncovering the neural mechanisms of these developmental processes.
Review limitations
There was a lack of consistency among the reviewed studies in RS-fMRI methods. For example, some studies conducted imaging of participants with eyes open whereas others conducted imaging with participants' eyes closed (see Table 1 for details). There is some evidence of significant differences in RS-fc between these two conditions such as for visual and auditory networks (Agcaoglu et al., 2019). However, it is less clear whether these differences exist for RS-fc of the AMY. Future studies should consider the potential variations between RS-fMRI conditions. In addition, reliability in RS-fc measurement may be impacted by scan length, with recommendations to increase scan length to as much as 13 min (Birn et al., 2013). In the reviewed studies, scan lengths ranged from 5 to 12 min (Table 1).
As previously acknowledged, the distinction between increased/decreased positive RS-fc and increased/decreased negative RS-fc is not always reported (Vaidya and Gordon, 2013). Thus, identifying general patterns of results across studies between the AMY and other specific areas of the brain is limited. Further, given that RS-fMRI research on the AMY and ELS is newly emerging, the ability to draw overall conclusions is restricted by the limited number of RS-fMRI studies. Hence, this review presents existing RS-fMRI methodology and findings within each study regarding ELS and AMY RS-fc to serve as a consolidated resource for informing and stimulating RS-fMRI evolutionary-developmental research.
Conclusion
RS-fMRI has the yet untapped potential to support research within an evolutionary theoretical framework for elucidating the neural underpinnings of adaptive strategies shaped by early adversity. Current evolutionary-developmental theories predict that impulsivity, versus higher self-regulation, may be more adaptive in some adverse contexts and this should be evident within neural profiles, including that of the AMY. In this review, the limited existing research using RS-fMRI to investigate RS-fc of the AMY for individuals who experience ELS is reviewed and it examined these alongside evolutionary-developmental hypotheses. Congruent with these hypotheses, individuals who experienced higher ELS often (1) exhibited changes in AMY RS-fc that may represent biological embedding of stress responsivity, and (2) they had patterns of AMY RS-fc that may indicate developmental trajectories aimed toward higher impulsivity. Not only does RS-fMRI offer feasible methodology to examine changes in functional connectivity of AMY and other corticolimbic regions for individuals of all ages, but it also provides an opportunity for examining timing effects of ELS (Graham et al., 2015b), an important developmental concern.
Specific to the reviewed findings of AMY RS-fc for individuals with ELS, there was an emerging pattern of decreased RS-fc with other corticolimbic structures (i.e., ACC and PFC) for the left AMY. However, for the right AMY, individuals with ELS displayed both increased and decreased RS-fc specific to other corticolimbic structures (i.e., hippocampus/PCC and ACC/PFC, respectively). Findings on overall AMY network topology also suggested unilateral effects with regard to node efficiency and betweenness centrality, and specifically for individuals with higher susceptibility to depression. Further work is needed to more deeply explore these unilateral effects, identify criteria for individual susceptibility, and determine how changes in AMY functional connectivity direct individual differences in self-regulatory behaviors. Studies linking intrinsic brain network functional connectivity (e.g., the extent to which brain networks have developed functional connections) and task-related functional connectivity (e.g., under what circumstances those functional networks are stimulated) may help advance this area of research.
Potential also exists in further exploring hypotheses on sex-differentiated roles that may be associated with differences in male and female neural profiles. Finally, measurement of dynamic RS-fc may offer additional information as to how ELS is linked to increases in (or decreases in) fluctuating connectivity between regions involved in self-regulatory processes and whether these fluctuations are associated with adaptive behaviors. Further development of methodologies involving RS-fMRI within an evolutionary-developmental theoretical framework is worthwhile in that they could allow more rigorous testing of hypotheses about the neural mechanisms for development of adaptive strategies shaped by adverse contexts.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the University of Arizona BIO5 Institute Team Scholars Award.