
Abstract
Background:
Synchronous and antisynchronous activity between neural elements at rest reflects the physiological processes underlying complex cognitive ability. Regional and pairwise connectivity investigations suggest that perturbations in these activity patterns may relate to widespread cognitive impairments seen in bipolar disorder (BD). Here we take a network-based perspective to more meaningfully capture interactions among distributed brain regions compared to focal measurements and examine network-cognition relationships across a range of commonly affected cognitive domains in BD in relation to healthy controls.
Methods:
Resting-state networks were constructed as matrices of correlation coefficients between regionally averaged resting-state time series from 86 cortical/subcortical brain regions (FreeSurferv5.3.0). Cognitive performance measured using the Wechsler Adult Intelligence Scale, Cambridge Automated Neuropsychological Test Battery (CANTAB), and Reading the Mind in the Eyes tests was examined in relation to whole-brain connectivity measures and patterns of connectivity using a permutation-based statistical approach.
Results:
Faster response times in controls (n = 49) related to synchronous activity between frontal, parietal, cingulate, temporal, and occipital regions, while a similar response times in BD (n = 35) related to antisynchronous activity between regions of this subnetwork. Across all subjects, antisynchronous activity between the frontal, parietal, temporal, occipital, cingulate, insula, and amygdala regions related to improved memory performance. No resting-state subnetworks related to intelligence, executive function, short-term memory, or social cognition performance in the overall sample or in a manner that would explain deficits in these facets in BD.
Conclusions:
Our results demonstrate alterations in the intrinsic connectivity patterns underlying response timing in BD that are not specific to performance or errors on the same tasks. Across all individuals, no strong effects of resting-state global topology on cognition are found, while distinct functional networks supporting episodic and spatial memory highlight intrinsic inhibitory influences present in the resting state that facilitate memory processing.
Impact Statement
Regional and pairwise-connectivity investigations suggest altered interactions between brain areas may contribute to impairments in cognition that are observed in bipolar disorder. However, the distributed nature of these interactions across the brain remains poorly understood. Using recent advances in network neuroscience, we examine functional connectivity patterns associated with multiple cognitive domains in individuals with and without bipolar disorder. We discover distinct patterns of connectivity underlying response-timing performance uniquely in bipolar disorder and, independent of diagnosis, inhibitory interactions that relate to memory performance.
Introduction
Bipolar disorder (BD) is a chronic mood disorder associated with widespread impairments in cognitive function, including general intelligence, executive function, memory, social cognition, and response timing (Bora et al., 2016; Bourne et al., 2013; Brotman et al., 2009; Mann-Wrobel et al., 2011). Cognitive impairments contribute to poorer functional and quality-of-life outcomes (Andreou and Bozikas, 2013; Baune and Malhi, 2015), persist during mood symptom remission (Mann-Wrobel et al., 2011; Volkert et al., 2016), and are not solely accounted for by medication use (Vrabie et al., 2015) and thus represent an important yet poorly understood target for future therapies.
Brain imaging studies to date have placed focus on regional activation and pairwise connectivity-based understandings of these impairments. However, how cognitive processing emerges from a whole brain of interacting elements may be more meaningfully probed using a network theory framework, which enables examination of multivariate patterns of functional connectivity across the brain. The application of this framework to intrinsic resting-state connectivity, which reflects the most frequent coupling patterns between regions (Cole et al., 2014), can provide a context-independent marker of cognitive impairment that potentially represents core functional circuit dysfunction in BD.
Consistent findings from localized approaches, which involve the specification of regions of interest, find functional dysconnectivity between and within regions of the prefrontal cortex and limbic system in BD (Chase and Phillips, 2016; Syan et al., 2018). Parallel to these seed-based analyses, findings from whole-brain network approaches suggest altered functional integration and segregation globally (Dvorak et al., 2019; Spielberg et al., 2016; Wang et al., 2017a), and locally among frontal cortex, limbic, and default-mode areas (Doucet et al., 2017; Dvorak et al., 2019; He et al., 2016; Roberts et al., 2017; Spielberg et al., 2016; Wang et al., 2016, 2017a, 2017b; Zhao et al., 2017).
There is corroborative evidence of underlying anatomical dysconnectivity both in terms of whole-brain integration and segregation (Nabulsi et al., 2019; O'Donoghue et al., 2017; Wang et al., 2019) and connectivity between prefrontal, basal ganglia, and limbic regions (Nabulsi et al., 2019; O'Donoghue et al., 2016; Perry et al., 2019) suggesting a structural foundation for altered functional connectivity in the illness. Recent work suggests impairments in intelligence (IQ), executive function, and processing speed associate with global anatomical network integration, segregation, and interhemispheric connectivity in BD (Ajilore et al., 2015; McPhilemy et al., 2019). However, an investigation of intrinsic functional network-level interactions relating to cognitive deficits in BD is yet to provide further context to understand their neural basis.
In BD, activation and seed-based resting-state approaches have investigated executive function (Alonso-Lana et al., 2016; Diler et al., 2013; Favre et al., 2013; Nguyen et al., 2017; Pompei et al., 2011), episodic memory (Oertel-Knöchel et al., 2015), and social cognition (Grant et al., 2018; Kim et al., 2009; Malhi et al., 2008), implicating specific patterns of functional disconnection within and between the frontoparietal, striatal, cingulate, and temporal areas. These approaches have been useful in understanding the neural underpinnings of these cognitive impairments in isolated brain regions, but are insensitive to complex functional interactions between neural elements across the brain and have often been restricted in using few cognitive performance measures. Here, we aim to determine resting-state network-level connectivity associated with cognition across a broad range of domains in healthy control (HC) and BD samples, to determine shared or differential relationships indicative of a breakdown in the resting-state network architecture supporting cognitive functioning in BD.
Based on previously reported associations in controls, we hypothesized that executive function, episodic memory, and IQ would associate positively with global efficiency and negatively with characteristic path length (Baum et al., 2017; Sheffield et al., 2015; van Den Heuvel et al., 2009) and that IQ would associate with specific patterns of functional connectivity between frontoparietal and default-mode regions (Hearne et al., 2016). Given the established deficit in these cognitive domains and alterations in global efficiency, characteristic path length, and subnetwork connectivity in BD, we hypothesized that differential brain-cognition relationships would exist in BD that may explain these cognitive deficits present in the disorder (Malhi et al., 2008; McPhilemy et al., 2019; Nguyen et al., 2017). In contrast, we explored relationships between short-term memory, spatial memory, social cognition, and the above cognitive domains with other commonly used measures of global integration and segregation and explored the variance in connectivity patterns underlying executive function, memory, social cognition, and a general factor of response time, all commonly impaired in BD.
This study represents an important extension of our previous work that examined neuroanatomical network features supporting cognition in this BD cohort (McPhilemy et al., 2019) and may provide a resting-state marker of functional connectivity useful in understanding cognitive deficits and brain function in BD.
Materials and Methods
Participants
Individuals with a diagnosis of BD and psychiatrically healthy individuals between 18 and 65 years of age were recruited through mental health services of the Western region of Ireland. The Diagnostic and Statistical Manual (DSM-V-TR) criteria for BD were confirmed by a psychiatrist using the Structured Clinical Interview for DSM-V (SCID) (American Psychiatric Association, 2013). Healthy volunteers had no personal history of psychiatric illness confirmed using the SCID, nonpatient edition, and no first-degree family history. Exclusion criteria included neurological disorders, learning disability, comorbid substance or alcohol abuse, history of head injury resulting in a loss of consciousness for over 5 min, or any illnesses potentially affecting cognitive function. The Hamilton Rating Scale for Depression (HAM-D-21) (Hamilton, 1959) and Young Mania Rating Scale (YMRS) (Young et al., 1978) assessed mood symptoms on the day of scanning and cognitive testing; euthymia was defined as scores of <8 and <7, respectively. All participants provided fully informed written consent. Ethical approval was obtained from The Clinical Research Ethics Committees of Galway University Hospital and St. James's Hospital Dublin.
Cognitive testing
Selected subtests of the Wechsler Adult Intelligence Scale (WAIS-III) (vocabulary, similarities, block design, and matrix reasoning) were combined to obtain full-scale IQ (Wechsler, 1997). The Cambridge Automated Neuropsychological Test Battery (CANTAB) was used to measure executive function (Intra/Extradimensional Shift, IED), episodic memory (Paired-Associates Learning), short-term memory (Delayed Match to Sample [DMS]), and spatial recognition memory (SRM) (Cambridge Cognition, 2018). The “Reading the Mind in the Eyes” assessed social cognition (Baron-Cohen et al., 2001). Principal component analysis (PCA) was used to identify variability factors from four available CANTAB response time measures; motor screening task (MOT) mean latency, DMS mean correct latency, IED total latency, and SRM mean correct latency. Bartlett's test of sphericity was significant (approximately chi square = 79.87, p = 3.81 × 10−15) and the Kaiser–Meyer–Olkin value of 0.76 exceeded the defined 0.6, suggesting an underlying latent structure in the response time data (Kaiser, 1974). One component exceeding an eigenvalue of 1 and explaining 58.34% of the total variance in response time was used to generate a factor score for subsequent analysis (Supplementary Fig. S1). Comparison of cognitive performance between diagnostic groups used multivariate analysis of covariance or Mann–Whitney U (SPSS v23).
Imaging acquisition and processing
Resting-state functional images were acquired with a single-shot echo-planar imaging sequence (repetition time [TR] 2 sec; echo time [TE] 28 ms; flip angle 90°; field of view 240 × 240 × 133 mm; a 3 mm2 in-plane resolution; 38 ascending slices covering the whole brain; 3.2 mm slice thickness with a 0.3 mm gap) using a 3T Philips Achieva scanner (Philips, The Netherlands) at the Centre for Advanced Medical Imaging, St. James's Hospital, Dublin, before the acquisition of any task-based functional images in the same scanning session. A total of 180 volumes were acquired along with four dummy scans to establish a steady-state longitudinal magnetization. Individuals were instructed to lie still with eyes open focusing on a red cross on the magnetic resonance imaging (MRI) screen for the 6-min scan duration. To register resting-state scans, T1-weighted images were acquired using a 3D magnetization-prepared rapid gradient echo sequence (TR/TE 8.5/3.9 ms; flip angle 8°; inversion time 804.3 ms; 1 mm3 isotropic voxel size). Preprocessing of resting-state functional images was performed to obtain functional correlation estimates with minimal sensitivity to motion, modeled on recommendations from Jo et al. (2013), including (1) despiking (3dDespike, Analysis of Functional NeuroImages [AFNI] v18.1.09
Functional network reconstruction
Eighty-six regions (34 cortical, 8 subcortical per hemisphere, and bilateral cerebellar hemispheres) were defined in a subject-specific manner (FreeSurfer v5.3.0,
Measures of global functional network connectivity
As hypothesized, executive function, IQ, and episodic memory were investigated in relation to global efficiency and characteristic path length. Exploratory investigations included relationships between these cognitive measures and positive/negative strength, global clustering, betweenness centrality and assortativity, between short-term and spatial memory, and social cognition and all global network measures (Brain Connectivity Toolbox v2017-15-01,
Subnetwork patterns of resting-state connectivity
Resting-state connectivity patterns associated with IQ, executive function, episodic, short-term, and spatial memory, social cognition, and the PCA-derived response time factor were investigated using a permutation-based method that controls for the familywise error rate (FWER) at the network level, covarying for age, gender, and diagnosis (network-based statistic, v1.2) (Zalesky et al., 2010). Unthresholded resting-state matrices retaining both positive and negative weights were used in this analysis. Interactions between cognitive performance and diagnosis were modeled to test for resting-state connectivity patterns associated with diagnosis-based variation in cognitive performance. A T-statistic representing the main effect of cognitive performance or interaction between cognitive performance and diagnosis for each connection was calculated and thresholded using T-statistic values of 1.5, 2, 2.5, and 3. FWER-corrected p-values (pFWE) were assigned to resulting subnetworks using a null distribution of maximum component size obtained via 5000 permutations.
Results
Participants
Analysis included 35 BD outpatient participants (28 BD type-I and 7 BD type-II) and 49 psychiatrically healthy individuals (HC) balanced for age, gender, and education (Table 1). Of the 35 BD participants, 24 met criteria for euthymia; 11 met criteria for mild-moderate depression (31%; HAM-D mean = 15.64, standard deviation [SD] = 4.80, range = 11–26), and 2 met criteria for hypomania (YMRS score = 10 for both individuals). Mood scores did not differ between the day of scanning and cognitive testing (HAM-D; T = −0.12, p = 0.91, YMRS; T = −0.77. p = 0.45). At cognitive testing, 20 BD participants were treated with mood stabilizers (11 lithium); 20 antipsychotic medications (19 atypical antipsychotics); 11 antidepressant medications; 1 benzodiazepine; 6 other psychotropic medications, and 3 antiepileptic mood-stabilizing medication; 4 were medication free (Supplementary Table S1). Over half of individuals with BD (n = 23) were taking a combination of at least two of the above classes of drugs. The time between scanning and cognitive testing was similar between diagnostic groups (HC mean ± SD = 5.73 ± 5.24 months, range = 0–20 months; BD mean ± SD = 6.83 ± 5.57 months, range = 0–16 months; T = −0.92, p = 0.36).
Demographic and Clinical Characteristics
Mood scores provided are from the day of scanning.
Significant difference at p < 0.05.
SD, standard deviation.
Cognitive differences between diagnostic groups
BD participants exhibited poorer neurocognitive performance than controls in executive function (U = 619.00, p = 0.03), episodic memory (F = 4.79, p = 0.03), short-term memory (F = 6.15, p = 0.02), and social cognition (F = 5.91, p = 0.02). No difference was demonstrated between groups for IQ (F = 3.14, p = 0.08), PCA-derived response time (F = 1.01, p = 0.32), spatial memory accuracy (F = 2.08, p = 0.15), or response time (F = 2.36, p = 0.13; Supplementary Table S2).
Global resting-state connectivity
We found an inverse relationship between episodic memory errors and characteristic path length but not global efficiency that did not survive false-discovery rate (FDR)-correction, and no relationships between IQ or executive function and these global network measures (Fig. 1 and Table 2). No interaction effects between diagnosis and global network measures on episodic memory, IQ, or executive function were detected (Table 3). Exploratory analysis found direct relationships between executive functioning errors and positive (F = 6.32, p = 0.01) and negative (F = 5.81, p = 0.02) strength; direct relationships between episodic memory score and positive (F = 5.96, p = 0.02) and negative (F = 8.26, p = 0.01) strength; direct and inverse relationships between episodic memory total errors and positive strength (F = 4.48, p = 0.04) and assortativity (F = 4.31, p = 0.04), respectively; and a direct relationship between spatial memory accuracy and betweenness centrality (F = 3.89, p = 0.05), all of which did not survive FDR correction. Group × global measure interactions were found for episodic memory errors and assortativity (F = 4.08, p = 0.05; HC r = −0.06, BD r = 0.24) and episodic memory score and betweenness centrality (F = 11.12, p = 0.001; HC r = 0.22, BD r = −0.51), with the latter surviving FDR correction.
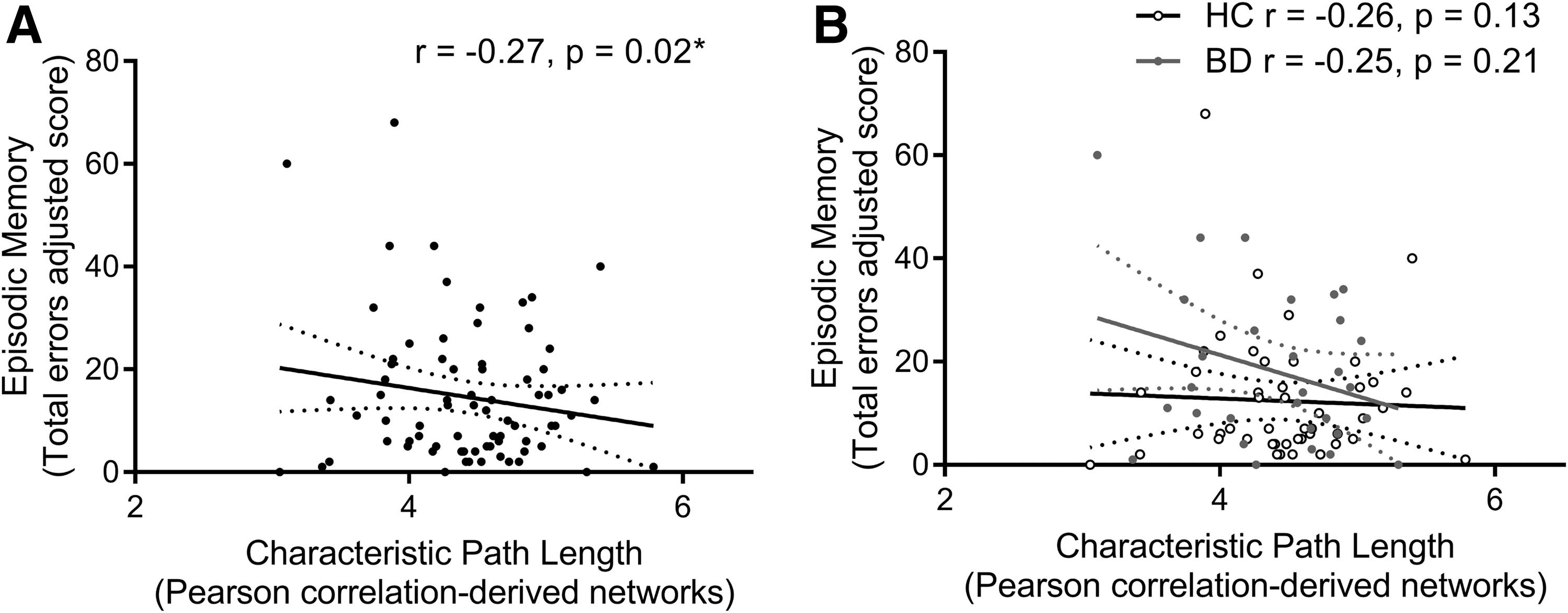
Relationship between episodic memory performance (total errors) and characteristic path length computed from Pearson-correlation-derived resting-state networks
Main Effect of Resting-State Global Efficiency on Cognitive Performance
Significant difference at p < 0.05.
Interaction Effects Between Diagnosis and Global Functional Connectivity Measures on Cognitive Performance
Resting-state subnetwork connectivity
Hypoconnectivity (decreased positive/increased negative connectivity) between distinct frontal, parietal, temporal, occipital, cingulate, insula, and amygdala regions related to lower episodic memory errors (T = 3, pFWE = 0.02; Fig. 2) and higher SRM accuracy (T = 2.5, pFWE = 0.02; Fig. 3) in the overall sample and similarly in BD compared with controls. We found no resting-state subnetworks related to IQ, executive function, short-term memory, or social cognition in the overall sample or in a manner that would explain deficits in these facets in BD.
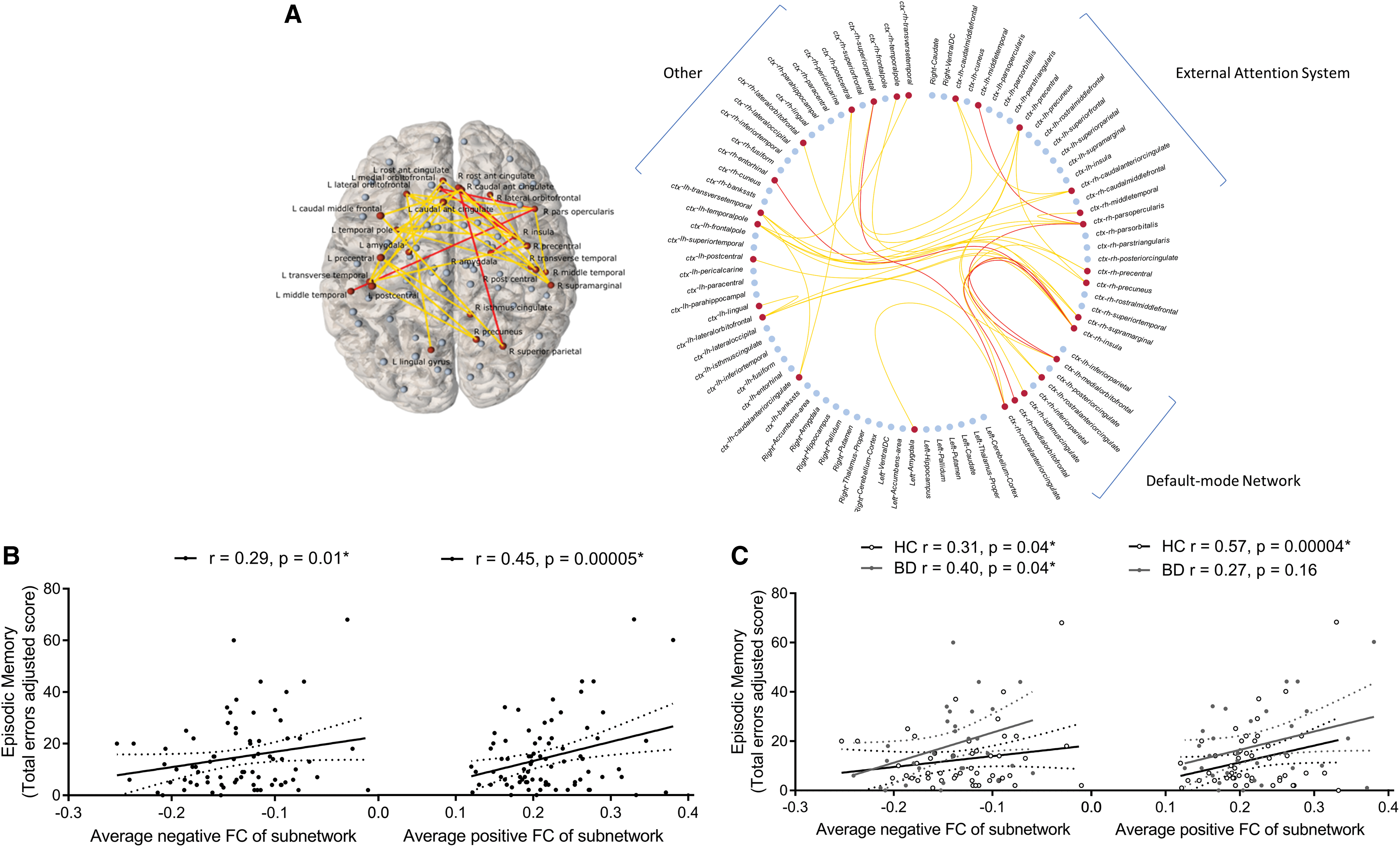
Episodic memory functional subnetwork (Pearson-correlation-derived networks) found over all subjects covarying for age, gender, and diagnosis (T = 3, p = 0.02).
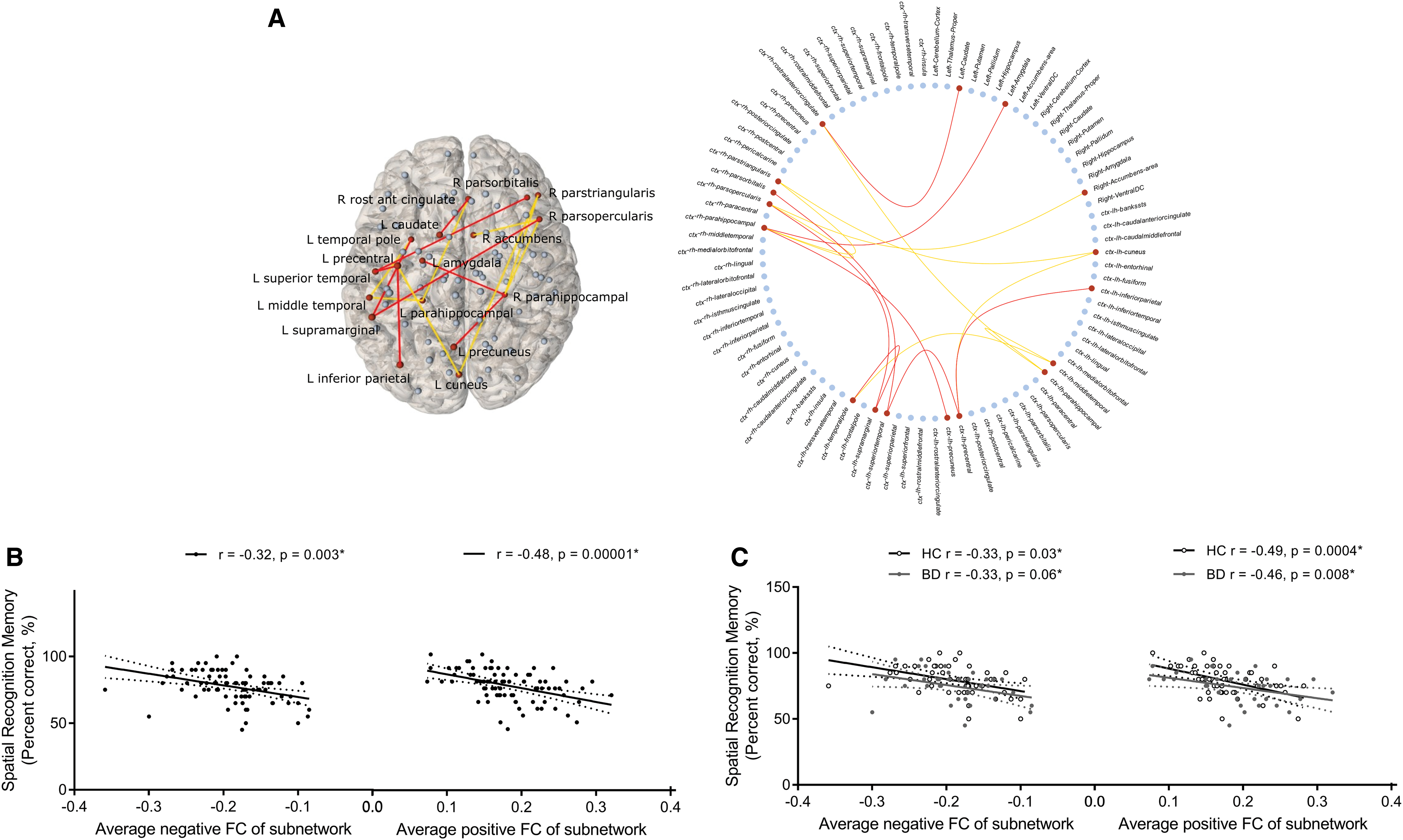
Spatial recognition memory accuracy functional subnetwork (Partial-correlation-derived networks) found over all subjects covarying for age, gender, and diagnosis (T = 2.5, p = 0.02).
Resting-state connectivity between frontal, parietal, cingulate, temporal, and occipital regions had a significantly different relationship with the PCA-derived response time factor in BD compared with controls (T = 3, pFWE = 0.03; Fig. 4); greater connectivity (increased positive/decreased negative) related to faster response times in the control group and an inverse relationship was detected in BD.
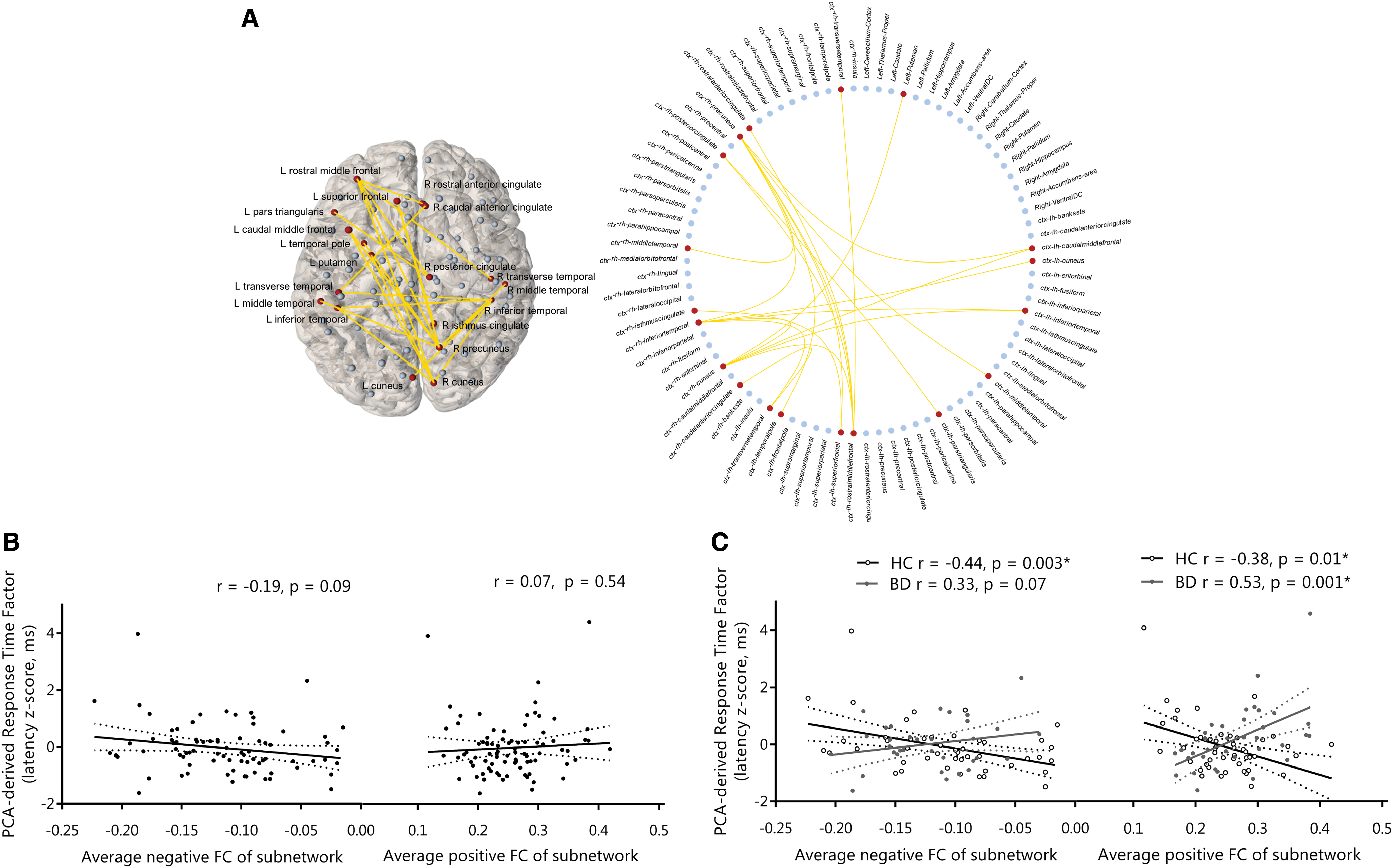
Response time functional subnetwork (Pearson-correlation-derived networks) that has a significantly different relationship with the PCA-derived response time factor in BD versus controls.
Discussion
We present for the first time evidence of alterations in the resting-state patterns underlying response timing in BD, which may prove important for understanding cognitive impairment generally in the disorder. Across all subjects, episodic memory related to global resting-state network integration and episodic and spatial memory to antisynchronous activity within distinct resting-state subnetworks; no relationships globally or in terms of subnetwork patterns explained variance in intelligence, executive function, or social cognition. Despite the presence of expected cognitive deficits in BD in executive function, episodic and short-term memory, and social cognition, we find no evidence implicating altered intrinsic functional connectivity patterns at the global or subnetwork level in these deficits.
Response timing
We find alterations in the resting-state connectivity patterns underlying response time variation in BD. Generally inconsistent with previous work (Bora et al., 2006; Brotman et al., 2009; Gallagher et al., 2015; Teixeira et al., 2013), reaction times were not significantly increased in the present BD cohort compared with controls, suggesting that reaction time deficits in BD may be sensitive to the attentional demands of the task (Townsend et al., 2012). Our findings corroborate previous task-based imaging of response time variation in controls, showing activation in the prefrontal, cingulate, and posterior parietal cortices, and suggest attentional mechanisms in the underlying neurobiology of response time deficits in BD (Bellgrove et al., 2004; Johnson et al., 2015; Simmonds et al., 2007; Yarkoni et al., 2009). Synchronous activity between these regions appears to support efficient response timing in healthy individuals, however, in BD, it is the antisynchronous activity that confers similar levels of functioning. Lower synchronous and greater antisynchronous activity between regions of this system has been identified in BD previously (Chase and Phillips, 2016) and in the present cohort (Nabulsi et al., 2020). Taken together, this suggests compensatory support due to a framework of functionally disconnected regions underlying response timing in BD.
Intelligence and executive function
Neither global efficiency nor distinct functional subnetworks related to intelligence or executive function generally or explained deficits in these facets of BD. We are consistent with a recent large-scale analysis reporting that resting-state efficiency does not explain variance in intelligence (Kruschwitz et al., 2018), and find that, while IQ and executive function impairments in BD may relate to altered integration and segregation within anatomical networks (McPhilemy et al., 2019), this does not extend to implicate resting-state connectivity. This may be expected given that the brain's intrinsic functional connections generally overlap with but are not identical to anatomical connections (Honey et al., 2009; Stam et al., 2015) and are not equally suited to the application of path-based measures when compared with anatomical networks (Honey et al., 2009; Petersen and Sporns, 2015; Stam et al., 2015). We were unable to detect expected network-level relationships with intelligence involving frontoparietal and default-mode regions, as recently reported in 317 healthy individuals using the same statistical approach (Hearne et al., 2016); or with executive function in overlapping neural systems detected using a priori and independent component analysis approaches (Reineberg et al., 2018; Vaidya and Gordon, 2013). Given brain-wide correction for multiple comparisons in network approaches, larger samples may be required to detect network-level relationships and subtle alterations in BD for these complex and integrative facets. Furthermore, a recent preliminary investigation of executive function deficits indicates that underlying functional connectivity differences may not be detected with aggregate connectivity patterns but rather connectivity dynamics (Nguyen et al., 2017).
Memory
We find distinct resting-state subnetworks supporting episodic and spatial memory across all subjects and none that explain cognitive deficits extant in BD. Longer average path length within resting-state networks related to less episodic memory errors; we note that the effect size of this relationship is moderate not surviving FDR-correction, however, potentially reflective of the specific inhibitory and not necessarily most topologically efficient pathways facilitating better memory performance. These findings corroborate a common inhibitory influence between external attention and default-mode systems supporting memory (Kelly et al., 2008) although is not exclusive to these systems at the network level and may not generalize to other actively engaging tasks. Specific to spatial memory performance, direct antisynchronous coactivation patterns between the right prefrontal, left parietal, and select limbic regions detected here reflect the separation of information transfer between distinct anatomical subnetworks that have been shown to relate to faster and slower response times in this spatial memory task previously in an overlapping cohort (see Supplementary Fig. S3 for visual comparison of these anatomical and functional networks) (McPhilemy et al., 2019). It is important to note that the precise nature of antisynchronous activity within functional networks remains unclear. While the present results may be indicative of inhibitory or competitive functional interactions between these brain areas (Fox et al., 2005), they may also represent the different spatiotemporal structures being produced on the underlying anatomical network rather than direct antagonistic relationships (Deco et al., 2011).
Social cognition
Despite an expected deficit in social cognition in BD, we find no resting-state, network-level connectivity patterns explaining this, building on our previous investigation, which identified no anatomical network-level basis (McPhilemy et al., 2019). Studies focusing on functional activation during social cognition tasks in BD implicate altered frontal, temporal, parietal, and insular cortex activation (Grant et al., 2018; Kim et al., 2009; Malhi et al., 2008; Willert et al., 2015) and lower seed-based functional connectivity between medial prefrontal and temporal cortices (Willert et al., 2015). It is therefore possible that coactivation patterns important for social cognition deficits are specific to task conditions and will be detected on examination of dynamic network configurations.
Methodological strengths, limitations, and future directions
In this study, we implement head motion correction and removal of white matter signal, cerebrospinal signal, and high motion time points from resting-state data, designed to mitigate physiological noise and head motion, which can spuriously contribute to variance in functional connectivity measures and thus sensitivity to detect relationships with cognitive ability (Satterthwaite et al., 2019). Global signal regression was not performed given that it mathematically induces spurious negative correlations, exacerbates the biasing effects that subject-motion can have on correlations between regions based on their distance, distorts resting-state differences between groups, and may remove behaviorally relevant information (Jo et al., 2013; Li et al., 2019; Murphy et al., 2009; Power et al., 2012; Saad et al., 2012). To guard against the problem of multiple comparisons, we addressed primary a priori hypotheses with additional exploratory analyses and used PCA dimensionality-reduction to analyze a parsimonious factor capturing most of the predictive information from the response time data. An important caveat to the present work is the influence that medication may have on both cognitive performance and fMRI measures in BD (Dandash et al., 2018; Gitlin, 2016; Hafeman et al. 2012; Torrent et al., 2011). The majority of individuals with BD in this investigation used medication, including lithium and antipsychotics; however, recent work indicates that medications have a limited effect on fMRI measures in BD (Hafeman et al., 2012). While this sample is reflective of the natural population one would expect, it remains possible that the findings are influenced by medication. In addition, our approach considers a single static network structure represented as the average resting-state connectivity over the course of scanning. We are therefore unable to assess how dynamic changes during the resting-state or task-specific network changes (Cole et al., 2014) relate to cognition and underlie cognitive deficits in BD. Functional network dynamics have been shown to represent an important driver of cognitive performance (Bassett et al., 2011, 2013; Cohen, 2018) and there is preliminary evidence to suggest that a reduced ability to reconfigure functional network architecture relates to processing speed and executive functioning deficits in BD (Nguyen et al., 2017). This is an exciting avenue for future neuroimaging investigations of cognitive impairments in BD.
Conclusions
Our findings suggest that commonly reported alterations in intrinsic connectivity patterns in BD relate to a specific breakdown in the support of response timing in contrast to performance or errors on the same executive function and memory tasks or intelligence or social cognition performance. Independent of BD, we identify distinct inhibitory resting-state patterns underlying memory performance, demonstrating the promise of network approaches in characterizing complex cognitive processing in the brain.
Authors' Contributions
D.M.C. designed, obtained funding for and supervised data collection, analysis, and interpretation; B.H. and
Footnotes
Acknowledgments
We gratefully acknowledge the participants, the support of the HRB-Clinical Research Facility Galway, the Centre for Advanced Medical Imaging at St. James Hospital Dublin, and funding support from the Irish Research Council Government of Ireland Postgraduate Scholarship.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This research was funded by the Health Research Board (HRA-POR-324) awarded to D.M.C.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Table S1
Supplementary Table S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.