
Abstract
Background:
Tinnitus and its mechanisms are an ongoing subject of interrogation in the neuroscientific community. Although most current models agree that it encompasses multiple structures within and outside the auditory system, evidence provided in the literature suffers from a lack of convergence. To further our understanding of contributions to tinnitus lying outside the auditory system, we explored a new model based on a proprioceptive hypothesis specifically in subjects experiencing chronic nonbothersome tinnitus due to acoustic trauma. The present study addresses the role of the right operculum 3 (OP3) involved in this model. It also investigates classical models of tinnitus.
Methods:
A seed-based resting-state magnetic resonance imaging study explored the functional connectivity in an acoustic trauma group presenting slight to mild nonbothersome chronic tinnitus and compared it with a control group.
Results:
Group differences were found with two networks: with the sensorimotor–auditory and the frontoparietal, but not with the default mode network nor the limbic regions. In the auditory pathway, the inferior colliculus displayed group differences in connectivity with the right superior parietal lobule. Exploratory analysis elicited a significant increase in connectivity between two seeds in the right OP3 and two mirror regions of the dorsal prefrontal cortex, thought to correspond to the human homologue of the premotor ear–eye field bilaterally and the inferior parietal lobule involved in proprioception, in the tinnitus group.
Conclusions:
These new findings support the view that acoustic trauma tinnitus could bear a proprioceptive contribution and that a permanent cognitive control is required to filter out this chronic phantom percept.
Impact statement
Tinnitus studies are complicated by the large heterogeneity of etiologies in subjects that are recruited. Here, we included participants experiencing chronic non-bothersome tinnitus following acoustic trauma, thus having few comorbidities, to address the cerebral functional networks modifications induced by this percept. We found functional connectivity modifications in several networks, including in the auditory pathways supporting the idea that tinnitus is not a single auditory issue. In addition, a specific connectivity between the right OP3 region and bilateral superior frontal gyrus was found, in line with Job's recent hypothesis of an implication of the middle-ear pathway.
Introduction
Tinnitus (i.e., ringing or whistling in the ears) is a symptom that emerges with a variety of diseases (Baguley and McFerran, 2013), hence the definition of several types of tinnitus. Among those, acoustic trauma tinnitus has been considered as the direct result of cochlear injury and deafferentations, inducing maladaptive changes in the central nervous system. However, this generally accepted model has been contradicted by evidence of people having hearing loss without tinnitus and vice versa, whereas treatments based on this model to abolish tinnitus in humans have proved unsuccessful (Elgoyhen et al., 2015; Lehner et al., 2016; Vanneste et al., 2013).
Investigating tinnitus as a pathology of brain networks, many studies based on resting-state analysis of functional magnetic resonance imaging (fMRI), electroencephalography, or magnetoencephalography data were designed. They generated a large diversity of results showing the implication of regions extending far beyond the auditory network. Following alterations have been shown: in a network of the right parietofrontal and anterior cingulate cortex (Schlee et al., 2009); concomitant dysregulation of limbic and auditory networks (Chen et al., 2017; Leaver et al., 2016a); implication of the default mode network (DMN) as suggested through a review analysis (Elgoyhen et al., 2012); a reduced interhemispheric connectivity between auditory areas (Hofmeier et al., 2018); involvement of the right executive control network in tinnitus patients with distress (Kandeepan et al., 2019). Also, a decrease in information transfer between frontolimbic and medial temporal regions (Mohan et al., 2018); frontostriatal network dysfunction (Hullfish et al., 2019; Leaver et al., 2016b); modification of cortical and subcortical functional connectivity in tinnitus encompassing attentional, mnemonic, and emotional networks (Maudoux et al., 2012) and default mode, auditory, dorsal attention, and visual resting-state networks (Burton et al., 2012; Husain, 2014) were reported. The discrepancies in literature results may be partly due to the heterogeneity of the associated pathologies in the tinnitus population studied. Hence the importance highlighted by some authors of targeting homogeneous groups of subjects in tinnitus studies (Koops et al., 2019; Schmidt et al., 2017; Wineland et al., 2012). Focusing on the tinnitus percept while disregarding distress can be achieved by recruiting a subpopulation of chronic tinnitus subjects with similar etiology whose quality of life is not impacted. We aim to reinvestigate network-based functional connectivity in such a group.
Unlike current acoustic trauma tinnitus models, we showed in our previous fMRI studies that tinnitus may come, not from hearing loss itself, but from a somatosensory disturbance, most probably involving middle ear muscle proprioceptive dysfunction (Job et al., 2011, 2016b), leading to the erroneous integration of proprioceptive signals (i.e., illusory sound) in the brain. Indeed, our previous study aimed at stimulating the middle ear proprioceptors by applying specific click vibration rates to the ear (Job et al., 2016b), which resulted in a tinnitus-like perception in healthy subjects at a particular frequency. Neural correlates of this stimulation were found in a small brain region in the right operculum 3 (OP3; Job et al., 2016b). This region had also been found hyperactivated in the fMRI study comparing sound detection in a tinnitus group with a control group (Job et al., 2012). We hypothesize that this region may have been overlooked in previous fMRI studies due to merging with neighboring regions when performing spatial smoothing with large kernel sizes.
It has been shown that disturbed muscle proprioceptive inputs using mechanical vibration send erroneous messages to the brain as a form of perception illusion (Kavounoudias et al., 2008; Roll et al., 2004). Similarly, auditory perception illusions could occur in the auditory system as long as such proprioceptors are present in the middle-ear muscles, a result that was demonstrated by Kierner et al. (1999). In acoustic trauma, the physical stress generated in the whole auditory system, including the middle ear, is extreme (Job et al., 2016a; Wojtczak et al., 2017) and could overwhelm physiological adaptations.
In this study, we focused on a homogeneous group of subjects presenting chronic nonbothersome acoustic trauma tinnitus, free of comorbidities such as anxiety or depression. In continuity with our previous study on this region in the right OP3, we wondered whether it could be involved in a network of tinnitus, or be part of an abnormal interplay between the main resting-state networks. Thus, we first investigated how the main resting-state networks described in the literature were affected by tinnitus in our study population. Second, we questioned the existence of a network specifically related to the tinnitus percept by exploring the connectivity of this OP3 region in acoustic trauma tinnitus. We sought to achieve these objectives using a seed-based connectivity approach with resting-state functional magnetic resonance imaging (rs-fMRI), which allows for adjustment of the data to confounding factors. In this study, hearing loss was a confounding physiological factor and psychological disturbance related to tinnitus was a second confounding factor. Following our hypothesis, specific connectivity related to the percept of tinnitus per se should be observed since we are taking these factors into account.
We studied whole-brain functional connectivity differences between the tinnitus group and the control group using several seeds including regions documented as belonging to the main resting-state networks, those located at and around the previously determined region in the right OP3 (Job et al., 2016a), and those along the auditory pathway. The left OP3 seed, mirror of the right OP3 seed, and the extended cytoarchitectonically defined right OP3 were also investigated.
Materials and Methods
Participants
Nineteen male chronic tinnitus participants (mean age 42.5 ± 12 years old) and 19 male tinnitus-free age-matched controls (mean age 42.5 ± 11.9 years) were included. Inclusion criteria required nonbothersome tinnitus of acoustic trauma origin, with a duration ≥6 months (mean duration 12.2 ± 7.3 years, median = 13 years). Tinnitus participants were recruited through medical unit physicians in military regiments, through general announcements in the regiments and through the ear, nose, and throat department of military hospitals open to civilians. Controls were recruited through an announcement on a specialized call-for-volunteers website for scientific protocols and by word of mouth. Control participants were not to have reported frequent or permanent tinnitus but may have experienced it once or occasionally. All participants were different from those studied in our previous publications to avoid circular analysis.
The study was approved by the Local Ethics Committee CPP Sud-est V, Ref: 10-CRSS-05 MS 14–52, and conducted in accordance with the Declaration of Helsinki. Informed written consent was obtained from all participants.
Hearing levels, tinnitus description, and tinnitus handicap inventory
Both tinnitus participants and matched controls underwent audiograms. We assessed hearing performances by tonal audiometry at 0.25, 0.5, 1, 2, 4, and 8 kHz (Audioscan fx by Essilor©) on the day of the magnetic resonance imaging (MRI) acquisitions. Electroacoustic calibration of the audiometer equipped with Beyer dynamics DT48 earphones was performed per French AFNOR standard S3007. We used the automated Hughson–Westlake procedure, which assessed hearing thresholds by modifying sound intensity using 5 dB up and 10 dB down steps. Sound intensity levels were modified automatically according to the participant's responses. The threshold was defined as the lowest level at which at least two responses were obtained out of three presentations on an ascending run. Hearing loss levels were expressed in dB hearing levels.
Two questionnaires were completed by tinnitus participants. The first was an interview form that collected general information about the participant's tinnitus. The second was the French version of the tinnitus handicap inventory (THI), used internationally to robustly assess the intensity/intrusiveness of tinnitus and its associated handicap (Newman et al., 1996). THI scores are divided into severity grades (i.e., slight [THI 0–16], mild [THI 18–36], moderate [THI 38–56], severe [THI 58–76], and catastrophic [THI 78–100]; McCombe et al., 2001). According to this grading, the participants' tinnitus ranged from slight to moderate (Table 1).
Tinnitus Participants' Characteristics
Age (in years), ear: side of the tinnitus percept, duration: tinnitus duration (in years), THI score: from the tinnitus handicap inventory questionnaire. Origin of the acoustic trauma: M, AR, W. Hearing loss: frequency range (in kHz) where hearing loss >25 dB HL (hearing thresholds ≤25 dB HL in frequencies tested are denoted [—]).
AR, army riffles; HLs, hearing levels; M, music; THI, tinnitus handicap inventory; W, noisy workplace.
Autonomic status
Since chronic tinnitus is known to affect autonomic conditions with sympathetic overactivity (Choi et al., 2013), we wanted to make sure that group differences would not be attributed to an imbalance in autonomic nervous functions. To verify this, we measured the heart rate variability (HRV) in all subjects before, during, and after the MRI experiment. In the scanner, during the resting-state acquisition (13 min long), the finger photoplethysmography device of the scanner, sensitive to the hemodynamic pulse at fingertip, was registered at 100 Hz using the hardware delivered with the magnetic resonance (MR) system. The peaks were automatically detected and recorded with the data file for further analysis. These peaks are fully correlated with the R-peaks from electrocardiogram (r 2 > 0.99) but delayed in mean by 620 ms (Rubio et al., 2015). For all the data, HRV analysis was performed with the dedicated software, Kubios®. Time domain, frequency domain, as well as nonlinear analyses were applied on interbeats intervals measured between consecutive peaks. Standard autonomic parameters were computed and statistical comparison between groups was performed in SPSS using nonparametric Mann–Whitney U tests.
MRI data acquisition
A rs-fMRI study was performed in the search for a potential functional network specific to tinnitus perception per se. Scanning was conducted on a 3T Philips Achieva-TX scanner (Best, The Netherlands) at the Grenoble MRI facility-IRMaGe, equipped with a 32 channel-head coil. The protocol included a long rs-fMRI sequence (longer than current sequences used in tinnitus research investigations) for which participants were instructed to lie with their eyes open and let their minds wander without focusing their thoughts on anything in particular while a gray background image with a small white cross in the center was displayed. A high-resolution structural T1-weighted image was also acquired. The parameters of the gradient echo echo planar imaging sequence were adjusted to lower the scanner noise, thus limiting the acoustic dose received by the participants. This was achieved by decreasing the slope of the commutation gradients (i.e., SoftTone = yes). The frequency spectrum of the acquisition sequence presented a peak at a lower frequency (630 Hz) and a 10 dB reduction compared with standard sequences. High frequencies, above or equal to 1 kHz, presented an average reduction of 13 dB when compared with the standard sequence used in fMRI on this MR scanner (Fig. 1). This sequence was reported as acoustically comfortable by the participants. Other main parameters were 32 slices, 3.5 mm thick, acquired with a multiband factor of 2, in-plane voxel size = 3 × 3 mm2, repetition time (TR) = 2000 ms, echo time (TE) = 32 ms, flip angle = 75°, SENSE factor = 3, 5 dummy volumes, and 400 volumes. This long acquisition duration (13′20″) was chosen since it is directly related to the reliability of rs-fMRI connectivity estimates (Birn et al., 2013; Termenon et al., 2016). The T1-weighted structural image acquisition had the following parameters: 3D Magnetization Prepared Rapid Acquisition GRE, 0.9 × 0.9 × 1.2 mm3, inversion time = 800 ms, TR = 25 ms, TE = 3.9 ms, flip = 15°, SENSE factor = 2.2.
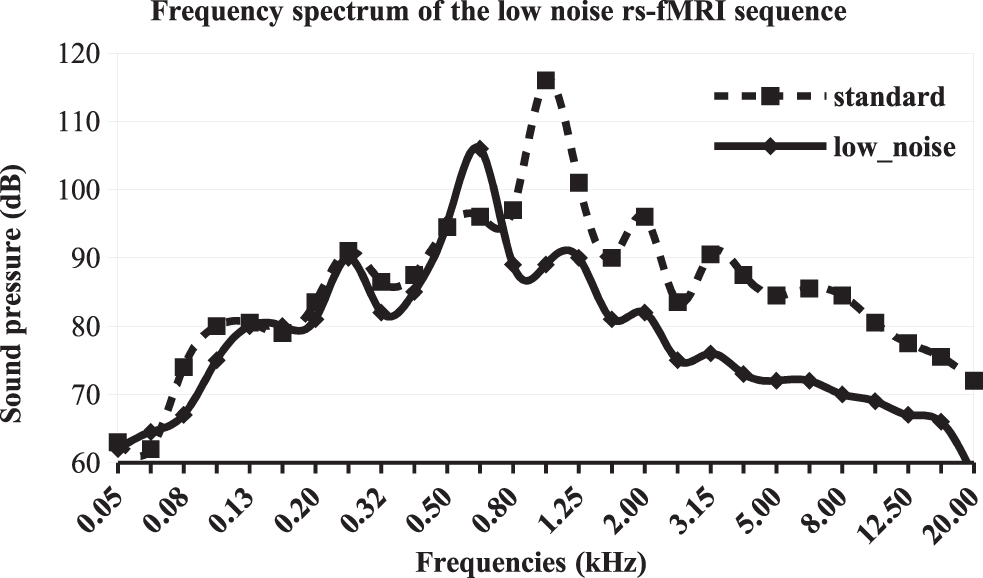
Frequency spectrum of the low noise rs-fMRI sequence used in this study (
MRI data preprocessing
As the seed of interest in the right OP3 is precisely localized within the brain, we designed the analysis pipeline to preserve spatial accuracy as much as possible. We preprocessed the fMRI data using Statistical Parametric Mapping 12 software. Preprocessing of fMRI data included motion correction, slice-timing, and coregistration of functional images on the individual structural image. We further realigned all individual anatomical and functional images to a common referential using the Diffeomorphic Anatomical Registration Through Exponentiated Lie Algebra (DARTEL) elastic registration algorithm (Ashburner, 2007) that allows accurate interindividual brain realignment in particular around the main sulci (Klein et al., 2009). To do this, we first segmented each structural image to extract the gray matter (GM), white matter (WM), and cerebrospinal fluid (CSF) images. We then calculated the deformation field that transforms the individual GM images to a highly resolved GM template compatible with the standard Montreal Neurological Institute (MNI) coordinates system. We chose the template provided with the Computational Anatomy Toolbox for sake of spatial accuracy. The deformation field image of each participant was further applied to the structural GM, WM, and CSF images. The mean structural image of all participants was computed for display purposes so that our results precisely matched the anatomy. The deformation field was also applied to the functional individual images with a small smoothing kernel size (1.5 mm3 isotropic) to limit blurring of the functional images. In an additional analysis, to better fit standard connectivity analyses, larger spatial smoothing (one with a kernel of [3 mm]3 and one with [5 mm]3) was applied to check whether the results found in the seed-based analysis with 1.5 mm smoothing were robust. All individual images were then in the same template space with the same spatial resolution (isotropic 1.5 mm) for further analysis.
Denoising was performed in all individual time series in template space within the connectivity (CONN) toolbox (Whitfield-Gabrieli and Nieto-Castanon, 2012; version 18.a). Outliers in functional time series were derived from the art toolbox, whereas physiological and noise sources were regressed from WM and CSF using CompCor (Behzadi et al., 2007). Six realignment parameters and their first-order temporal derivatives were also regressed at this point, but no global signal regression was performed since it can affect correlation patterns and introduce bias in group analysis (Saad et al., 2012). Before denoising, the histogram of voxel-to-voxel connectivity values varies between subjects and was shifted to the right. After linear regression of the confounds, the distribution of correlation values was centered around zero, with standard deviations across subjects ranging from 0.11 to 0.16 for 107 degrees of freedom, indicating good data quality. The results of the denoising step were visually inspected in each participant. Finally, band-pass filtering was applied within 0.008 to 0.08 Hz to all time series, to select the data matching the resting-state frequency band.
Seed definition for functional networks analysis
The network-based analysis included the DMN, first described by Fox et al. (2005) and six networks derived from the independant components analysis decomposition of resting-state data set of 497 individuals recruited in the Human Connectome Project. These networks correspond to the visual, the sensorimotor–auditory, the saliency, the language, the frontoparietal, and the dorsal attentional networks. A seed was defined at each node of the networks.
Definition of exploratory seed regions
We included as our main region of interest (ROI) the tinnitus-related region in the right OP3 derived from our previous study (Job et al., 2016a). This ROI and its spatial equivalent on the left hemisphere (as control) were boxes (9 × 4 × 4 mm3) built with Marsbar, centered at MNI: ±40–13 17, corresponding to the location previously found. For a more extensive exploration of the right OP3, we created two additional equal-sized boxes just anterior and posterior to the first ROI, respectively, centered at MNI: ±40–9 17 and ±42–17 17 (Fig. 2). We also defined an ROI that encompassed the whole right OP3 with the help of the Anatomy toolbox.
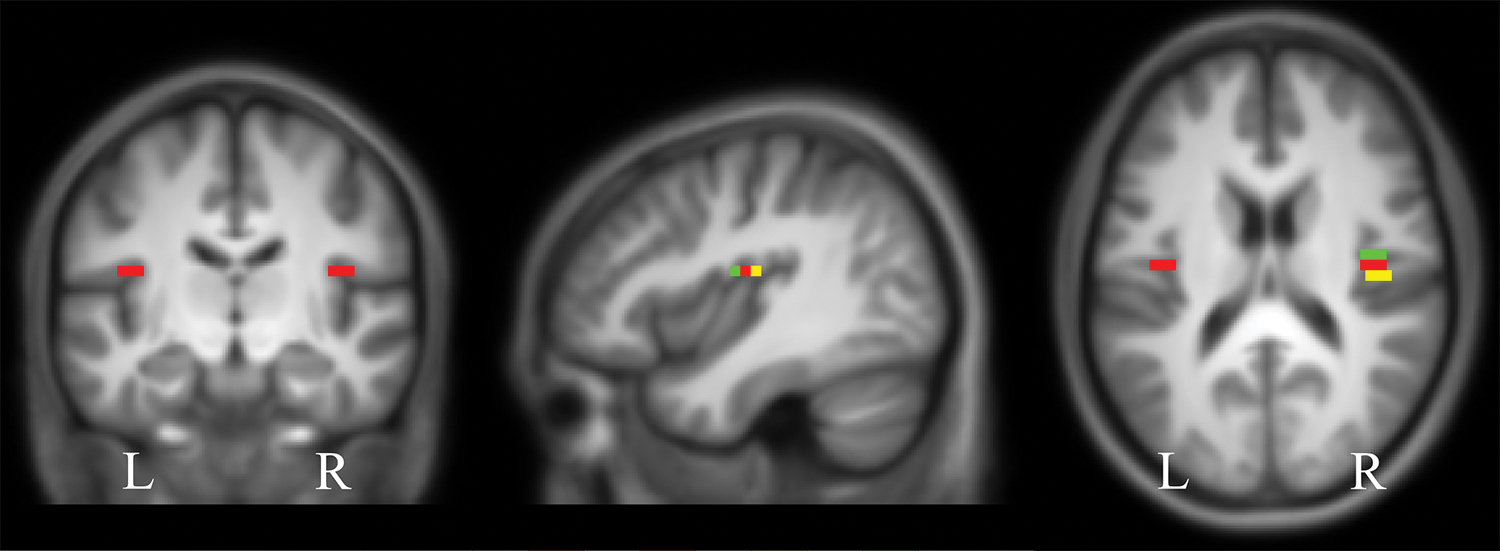
Seeds from the OP3 region used in this connectivity study: the tinnitus-related seed (in red; centered at MNI coordinates ±40, −13, and 17) and the seeds from the anteroposterior gradient: the anterior seed (in green) and the posterior seed (in yellow). Localizations are superimposed on the mean anatomical images of our group of 38 subjects. MNI, Montreal Neurological Institute; OP3, operculum 3. Color images are available online.
To address the connectivity of the auditory areas said to be involved in tinnitus perception, six other regions were chosen bilaterally along the auditory pathway. These seeds were defined based on the anatomy in the inferior colliculus (4 mm sphere centered at ±4, −36, and −10), the medial geniculate bodies (5 mm sphere centered at ±17, −24, and −10), and based on the Anatomy toolbox for Heschl's gyri by combining the Te1.0, Te1.1, and Te1.2.
Statistical analysis of functional connectivity networks
All connectivity analyses were performed using the CONN toolbox (v. 18a).
For each subject and each network, a seed-to-voxel connectivity map was computed from each seed of the network using Pearson's correlation. Individual maps were converted to z-maps using Fisher's transform to ensure data normality. In the network-based analysis, an F-test was performed across all seed maps and subjects in each group (within-group analysis), introducing THI and hearing loss as covariates of no interest. For within-group analysis, whole-brain false discovery rate (FDR), FDR-corrected threshold of p < 0.05 was retained for statistical significance. Differential functional connectivity (between-group analysis) is performed by weighing the F-test with a [1–1] contrast. For between-group analysis, p < 0.0001 and a cluster-size p-FDR corrected <0.05 were retained for statistical significance.
Statistical analysis of seed-based functional connectivity
For each subject and each seed, a seed-to-voxel connectivity map was computed using Pearson's correlation and converted to z-maps using Fisher's transform. For within-group analysis, a one-sample t-test was performed while regressing THI and hearing loss, considered as covariates of no interest. An FDR-corrected threshold of p < 0.05 was retained to assess statistical significance. In the between-group analysis, differential connectivity was calculated using a two-sample t-test with the contrasts [1–1] testing for stronger connectivity in tinnitus compared with controls and [−1 1] testing for weaker connectivity in tinnitus compared with controls. For exploratory analysis, we chose a statistical threshold (p < 0.0001, cluster-size p-FDR corrected <0.05) to elicit significant differential connectivity.
Results
Participants' characteristics
In tinnitus participants, the mean hearing threshold was significantly different from that of the control group, for frequencies ranging from 1 to 8 kHz (Mann–Whitney test, p < 0.05), with marked hearing loss at frequencies >4 kHz (Mann–Whitney test, p < 0.001), as usually observed with acoustic traumas (Fig. 3). At the frequency of the peak of noise in the acquisition sequence, no difference in hearing performance was found between the two groups, thus ruling out the possibility that differential connectivity arose from differences in acoustic perception related to scanner noise. All tinnitus participants described their tinnitus as high-pitched whistling, except for one participant who described a medium high-pitched sizzling. Tinnitus loudness was scored on an analog visual scale (mean 4.5 ± standard deviation 1.6 ranging from 2 to 8, from a total possible range 0–10). Bilateral tinnitus represented 68% of our population. There was the same amount of right and left tinnitus. Tinnitus duration ranged from 6 months to 25 years (mean 12.2 ± 7.3 years).
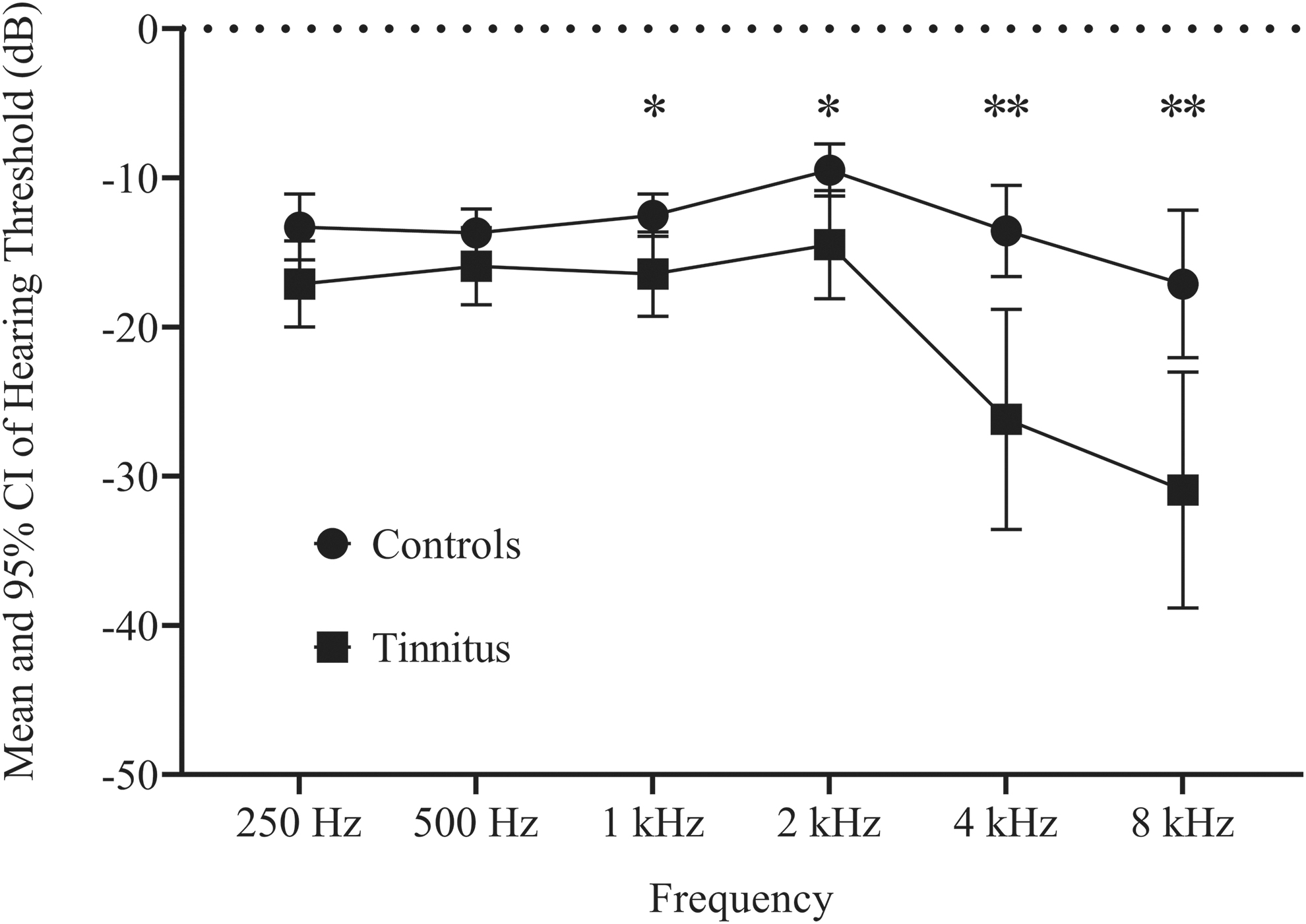
Audiograms of acoustic trauma tinnitus participants and control groups (mean ±95% CI) for both ears showing the hearing loss at high frequencies for the tinnitus group. Mann–Whitney tests results in the comparison of hearing thresholds (*p < 0.05, **p < 0.001) are displayed. CI, confidence interval.
THI scores described slight to mild tinnitus in all participants (mean 16.2 ± 10.5) except for one with moderate tinnitus (THI = 44), reflecting a tinnitus population less impacted than in the current literature in the field.
Detailed results from audiometry and neuropsychological testing are presented in Table 1 for tinnitus participants.
No significant differences in autonomic status were found between our two groups during rs-fMRI acquisition.
Functional network-based connectivity results
Within-group network-based results were able to replicate typical patterns of connectivity in all networks.
At network level, between-group differences were found with the sensorimotor–auditory and with the frontoparietal networks (Fig. 4). With the sensorimotor–auditory network, we found differences in connectivity with the right paracingulate gyrus (5, 26, 41), the right middle temporal gyrus (63, −51, 04), and the right precentral gyrus (−50, 4, 15). In the connectivity with the frontoparietal network, differential connectivity between the two groups was found with the right medial frontal gyrus (40, 18, 34). Coordinates are reported in MNI system after DARTEL registration (Table 2).

Between-group comparison of functional connectivity with the sensorimotor–auditory network
Significant Differential Functional Connectivity Results for the Sensorimotor–Auditory and Frontoparietal Networks
No other significant differential connectivity was found between both groups. Results were all obtained at height threshold p < 0.0001, extent threshold p-FDR corrected <0.05.
BA, Brodmann Area; FDR, false discovery rate.
No between-group differences were found in the connectivity with the DMN, visual, language, dorsal attentional, and saliency networks at the chosen statistical thresholds.
Functional connectivity with the auditory pathway seeds
Within-group analysis with Heschl's gyri showed bilateral connectivity with auditory regions, operculum, insula, dorsal anterior cingulate cortex, and the face representation of the sensorimotor cortex (Fig. 5).

Within-group connectivity with the right
Between-group analysis with Heschl's gyri seeds elicited a single significant differential connectivity between left Heschl's gyrus and the bilateral posterior cingulate cortex (0, −46, 32; Table 3).
Significant Differential Functional Connectivity Results for Tinnitus>Controls Groups Along the Auditory Pathway
No other significant differential connectivity was found between both groups. Results were all obtained at height threshold p < 0.0001, extent threshold p-FDR corrected <0.05.
Between-group analysis with the inferior colliculus seeds elicited a difference in the right superior parietal lobule (38, −40, 39; Table 3). No differential connectivity was found from the medial geniculate body seeds at the defined statistical thresholds.
Functional connectivity with OP3 seeds
Within-group analysis of the right OP3 box at (40, −13, 17) in controls and tinnitus (Fig. 6A) revealed a main connectivity with the sensorimotor network (bilateral precentral gyri, bilateral postcentral gyri, bilateral central and parietal opercular cortices, and supplementary motor area), but also with the salience network (bilateral insular cortex and anterior cingulate gyrus) and the auditory network (bilateral Heschl's gyri, bilateral planum temporale, and bilateral planum polare). Within-group analysis of the contralateral left OP3 box in controls and tinnitus (Fig. 6B) provided a similar pattern as with the right OP3 box, reflecting a symmetrical pattern of connectivity. Within-group connectivity of the whole right OP3 region (Fig. 6C) presented a larger extent of connectivity than for the right OP3 box, justifying the precise localization of our seed.
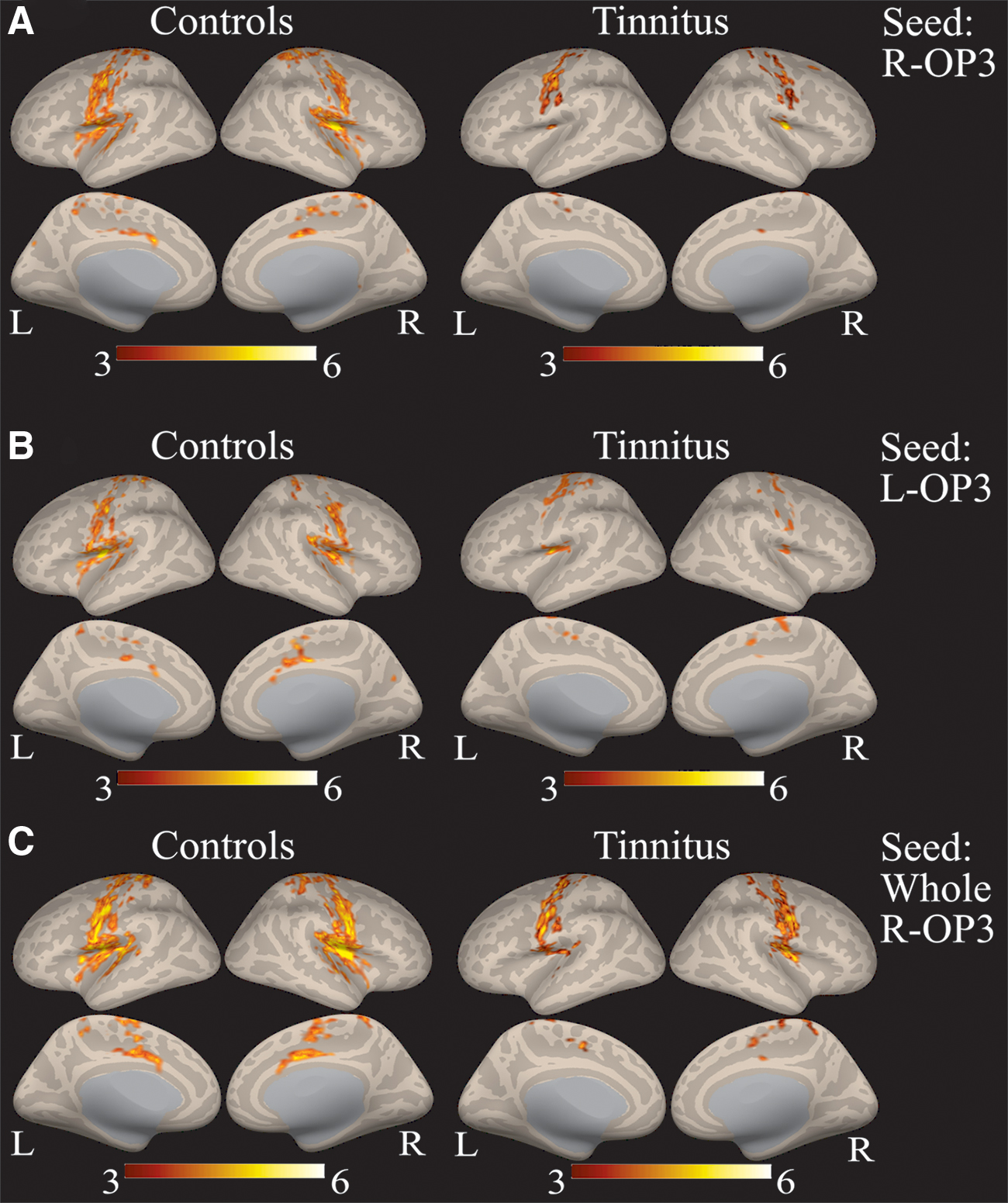
Within-group connectivity with the right
Between-group comparison revealed a specific connectivity in the tinnitus group compared with controls with the right OP3 seed. Increased connectivity was found with a region in the right superior frontal gyrus at the border of Brodmann Area (BA) 6 and 8 (Fig. 7A), with a maximum at MNI coordinates (20, 11, 47; Table 4). We could not elicit any differential connectivity from the homologous left hemisphere OP3 box, even at a lower threshold. No significant reduced connectivity was found in the tinnitus group as compared with the control group. The additional analysis performed with a spatial smoothing of 3 × 3 × 3 mm3 confirmed the finding in BA6/8 (same maximum location, T = 6.17 and extent = 28 voxels). The additional analysis with a 5 × 5 × 5 mm3 kernel provided no significant difference between groups, probably due to the mixing of signals originating from different regions (opercular, insular, and temporal regions) induced by spatial smoothing. These results confirm the need for a precise localization of the seeds.
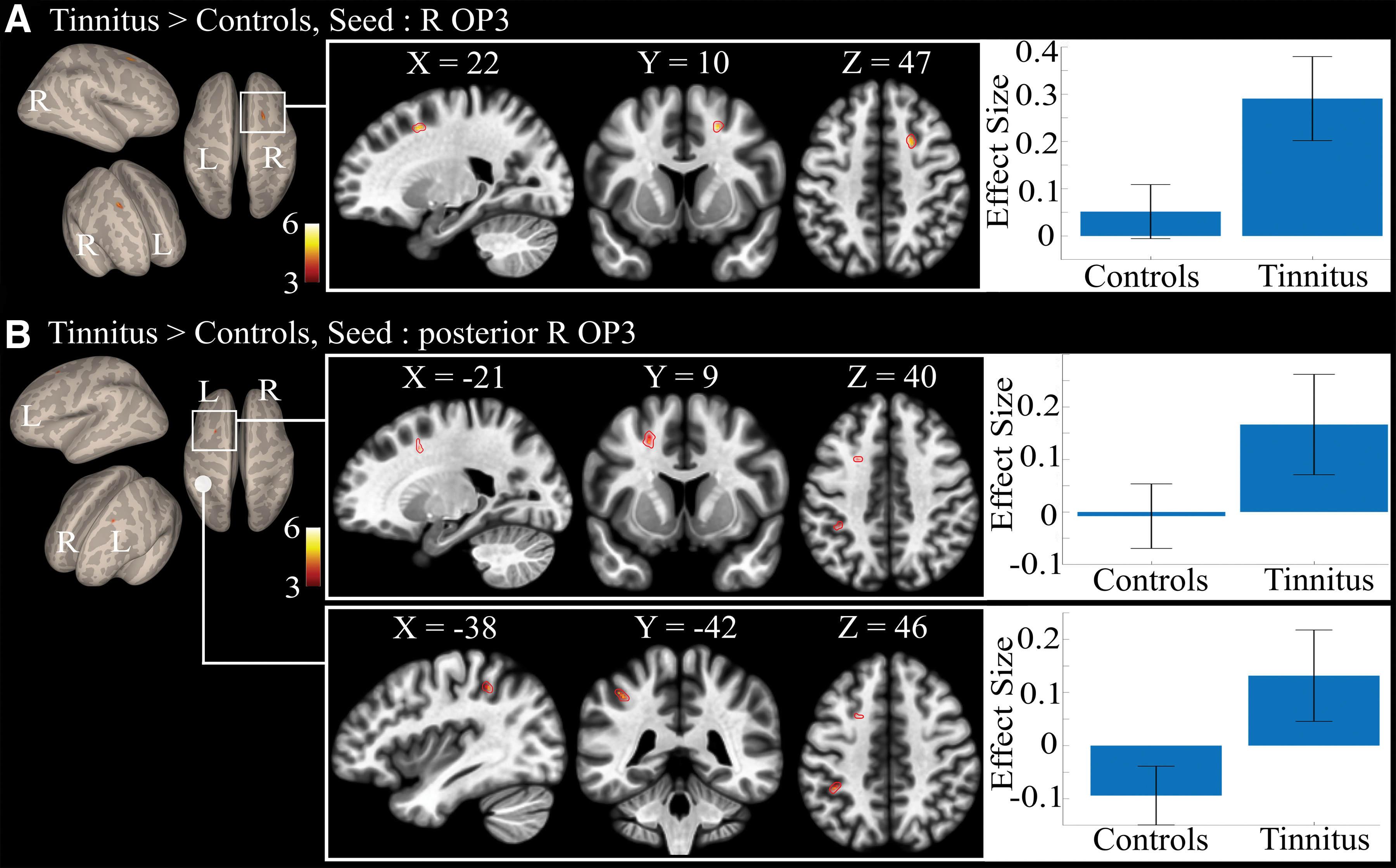
Between-group connectivity:
Significant Differential Functional Connectivity Results for Tinnitus>Controls Groups for the Right Operculum 3 (OP3) Seed and for the Right OP3 Posterior Seed
No other significant differential connectivity was found between both groups. Results were all obtained at height threshold p < 0.0001, extent threshold p-FDR corrected <0.05. Seeds location can be seen in Figure 2.
Region achieving above height threshold but only a tendency for statistical extent threshold.
OP3, operculum 3.
The anterior and posterior seeds in the right OP3 gave different between-group differential connectivity results. The anterior seed did not show any significant differential connectivity, whereas the posterior right OP3 seed displayed an increased connectivity with a region contralateral to the frontal region elicited earlier with the original R-OP3 box and with the left inferior parietal lobule (IPL; Fig. 7B; Table 4). The seed comprising the whole OP3 displayed no between-group differences in connectivity. No decreased connectivity in the tinnitus group was found compared with the control group.
Discussion
The chronicity of tinnitus may induce particular connections within the brain, which could correspond to a “tinnitus network.” Many investigations using fMRI were performed in an attempt to observe this phenomenon. However, facing the heterogeneity of literature results, the need for homogeneous groups of tinnitus participants has been recently pointed out. In this study, we conducted an fMRI study at rest with a group of subjects with nonbothersome chronic tinnitus of similar etiology, acoustic trauma, and compared resulting functional connectivity with a group of matched healthy subjects.
Differential connectivity with the main resting-state networks
Functional connectivity with the main resting-state networks revealed differences between our groups in two networks (Table 2). In the sensorimotor–auditory network, we found between-group differences in connectivity with the left inferior premotor area, the right associative auditory area, and the right paracingulate gyrus. The left inferior precentral gyrus is located within an area common to all kinds of motor illusions (Naito et al., 2017). This difference in connectivity in the tinnitus group reinforces our hypothesis that tinnitus could relate to a proprioceptive illusion linked to the middle ear muscles (Job et al., 2016b). The differential connectivity that we found with the right posterior middle temporal gyrus is associated with auditory processing, such as the detection and analysis of acoustic changes (Schönwiesner et al., 2007) or the tracking of sound in space (Da Costa et al., 2018). The paracingulate gyrus seems to be a key region for allocating cognitive resources to concurrent information (Gennari et al., 2018). This leads one to think that the chronic perception of phantom sounds must require a permanent allocation of cognitive resources to extract the relevant auditory input while minimizing the middle ear proprioceptive illusion corresponding to tinnitus. The frontoparietal network was also found to present a differential between-group connectivity with the right medial frontal gyrus, a core node for cognitive control. Interestingly, in this very region, the activity in response to a cognitively demanding task was found reduced in a group of chronic tinnitus subjects, suggesting a cognitive load permanently involved in maintaining tinnitus awareness (Trevis et al., 2017). Another explanation for this cognitive load could be the permanent filtering out of the tinnitus percept.
Unlike in previous studies, the lack of differences in other networks such as the DMN (Chen et al., 2018; Elgoyhen et al., 2012) or the attentional networks (Husain, 2014; Maudoux et al., 2012; Schmidt and Carpenter-Thompson, 2017) is notable here. In the DMN, this may be related to the lack of distress associated with tinnitus in the present participants. Indeed, the distress component of tinnitus has been shown to be associated with increased connectivity in the DMN (Chen et al., 2018) and between the limbic system and the auditory cortex (Chen et al., 2017; Leaver et al., 2016a). No difference in connectivity was found in our study with the dorsal attentional network, although Schmidt et al. (2017) found an increased connectivity of this network with the precuneus that correlated with tinnitus severity (Schmidt and Carpenter-Thompson, 2017). This discrepancy could be due to the low severity of tinnitus in our group of subjects compared with Schmidt's group.
Differential connectivity along the auditory pathway
Along the auditory pathway, we found differences in connectivity between the inferior colliculus, known for its role in integrating multiple auditory signals that help filter out sounds from vocalizing, breathing, and chewing activities and focus on important sounds, and the right superior parietal lobule, which is involved once again in cognitive control. This suggests that early along the auditory pathway, different cognitive resources are required for the filtering out of tinnitus. Interestingly, in rats with noise-induced tinnitus, deep brain stimulation of the inferior colliculus suppresses behavioral evidence of tinnitus (Smit et al., 2016). Although Davies et al. (2014) displayed no group differences in the auditory cortex in their study of nonbothersome tinnitus, we found increased connectivity in the tinnitus group compared with controls, between the left Heschl's gyrus and the posterior cingulate cortex. The latter is a node of the DMN that is thought to play a central role in supporting internally directed cognition. For tinnitus treatment purposes, this region was targeted in a neurofeedback intervention and showed a reduction of tinnitus-related distress (Vanneste et al., 2016). In our case, as tinnitus subjects did not present such distress, this observation would not explain the increased connectivity. No other significant differential connectivity was found between our two groups, supporting the view that mild tinnitus after acoustic trauma is not directly related to modified connectivity with auditory regions. This is consistent with results from Langers et al. (2012) who found that macroscopic tonotopic reorganization in the auditory cortex is not required for the emergence of tinnitus, and that this reorganization relates to hearing loss.
Exploratory findings with OP3 and nonauditory hypothesis
When considering the seed of the right OP3 box derived from previous studies (Job et al., 2012, 2016b), within-group connectivity was found to involve the somatosensory and the salience as well as the auditory networks. This suggests that our seed may play a pivotal role in the integration of somatosensory and auditory inputs presenting salient features. This result is in line with a recent functional connectivity study suggesting that the parietal operculum could be a connector hub linking auditory, somatosensory, and motor cortical areas (Tanaka, 2018). When checking for significant between-group differences, we found an increased connectivity between the tinnitus-related right OP3 seed and a region of the right superior frontal gyrus (at the limit between BA6/8). As this seed did not present any differential connectivity with sensorimotor–auditory networks, it seems to be a new connection. Furthermore, when testing the functional connectivity from the region just posterior to this seed, the increased connectivity found in the tinnitus group compared with controls in the left superior frontal gyrus mirrors that on the right (Table 4). This suggests the existence of a connectivity gradient within the right OP3. These bilateral regions are located anatomically just anterior to the bilateral frontal eye field. This seems to correspond to the region of the premotor ear–eye field (PEEF) documented in macaque monkeys (Lanzilotto et al., 2013), which is thought to play an important role in engaging the auditory spatial attention for the purpose of orienting eye and ear toward the sound source. The region found here could act as a human homologue of the PEEF, so a human PEEF (H-PEEF). In macaques, the PEEF involves auditory motor cells for both eye and ear movements (Hutchison et al., 2012; Lanzilotto et al., 2013; Lucchetti et al., 2008). The human homologue to the macaque PEEF (H-PEEF), for which the connectivity with the right OP3 is disturbed in participants with tinnitus, is a new finding. It is possible, although speculative, that similarly to the PEEF in macaques, the H-PEEF might play a premotor role in the control of coordinated ocular movements and tympano–middle-ear movements, pinna movements being vestigial in humans. If so, the oculomotor muscles' proprioceptive system responsible for eye coordination (Donaldson, 2000) would be disturbed by its potential coupling with an altered middle-ear system.
Interestingly, these results could shed light on unexplained observations of poor binocular coordination in subjects with tinnitus. Oculomotor performance during saccades (Lang et al., 2013), fixation, smooth pursuit, optokinetic nystagmus movements, and vergence was reduced in tinnitus participants Jozefowicz-Korczynska, 2002; Kapoula et al., 2010; Lockwood et al., 2001; Mezzalira et al., 2007; Yang et al., 2010); however, signs of vestibular dysfunction were absent. Furthermore, there is mention in the literature of tinnitus subjects capable of modulating their tinnitus with eye movements (Coad et al., 2001). These lower performances do not seem to be related to the severity of the handicap because they do not correlate with neuropsychological tests measuring the emotional/attentional status Jozefowicz-Korczynska, 2002). A cross-talk with the visual system could also provide some answers to another poorly understood condition, that of visual snow reported by 63% of subjects with bilateral tinnitus (Renze, 2017).
An alternative explanation could be that these bilateral areas in the superior frontal gyrus correspond to the 6 ma region described in the atlas derived from the data of the Human Connectome Project (Glasser et al., 2016). Interestingly, a study of the recently identified fiber pathway, the frontal aslant tract, describes the fine structural connections of this tract and shows the existence of a WM link between OP3 and the 6 ma region (Briggs et al., 2018). Its function is still not well documented but a report suggests the right frontal aslant tract may support executive function, namely inhibitory control and conflict monitoring for action (Dick et al., 2019). Whether this tract is different in subjects experiencing tinnitus remains to be studied and lies beyond the scope of this study.
In addition, in the right posterior seed, increased connectivity was observed with the IPL. The IPL is a vast region participating in the integration of sensory information. For instance, the left IPL is involved in proprioceptive illusion (Kavounoudias et al., 2008; Naito et al., 1999, 2017), tactile illusion (Kavounoudias et al., 2008), as well as auditory illusions (Brancucci et al., 2017). These new findings support the view that acoustic trauma tinnitus could bear a proprioceptive contribution.
Tinnitus group homogeneity
Tinnitus population homogeneity appears as a key parameter to help disentangle tinnitus percept per se, associated auditory dysfunctions such as hearing loss, psychological, and cognitive impacts of tinnitus, and resulting alterations of the quality of life. In this report, we studied a group of subjects experiencing chronic slight-to-mild tinnitus (mean THI 16.2), of moderate loudness (mean Visual Analogic Score 4.5 on a 0–10 scale), with greater hearing loss in the high-frequency range than in the control group, and duration >6 months, indicator of the chronicity of the percept. We regressed out the THI and the hearing loss to limit their contribution. The influence of duration was tested (not described here) but did not show any significant results, indicating that chronicity rather than duration is the important feature in these subjects. Moreover, to rule out the potential influence of the autonomic system to the functional connectivity, we measured the HRV in both groups, finding differences in neither the low-frequency range nor the high-frequency range related to respiration rythm. We thus believe that the results correspond to the percept per se, and that the regions highlighted are involved in the coping strategies for living with tinnitus without discomfort.
Methodological issues
Common to all tinnitus subjects are the phantom percept and the related coping mechanisms. However, the acquisition conditions, the analyses used in the field are heterogeneous and consensual results are difficult to provide. We thus believe that future studies would be able to replicate at least part of our findings when using the same kind of data and processing (long-acquisition duration, precise normalization using DARTEL, and small smoothing kernel). To allow future researchers to benefit from our results, we deposit our preprocessed, denoised, and filtered data on the OpenNeuro platform.
Conclusion
In this study, we compared the functional brain networks of a group of subjects with chronic slight to mild nonbothersome tinnitus of acoustic trauma origin with a group of control subjects. We found differences in connectivity in the main networks with regions involved in cognitive control and proprioception. In an exploratory analysis, we found that the subregion in the right OP3 already shown to be related to tinnitus (Job et al., 2012, 2016a) and its immediate posterior region are connected to two mirror areas located bilaterally in the superior frontal gyrus, just anterior to the bilateral frontal eye fields. This altered connectivity does not reflect an abnormal interplay between the main resting-state networks. Instead, it seems focal because when we further tested a larger seed involving the whole cytoarchitectonic right OP3, there no longer was any abnormal connectivity.
These new findings support the view that tinnitus could have a proprioceptive contribution in acoustic trauma and that a permanent cognitive control is required to filter out this chronic phantom percept.
Footnotes
Authors' Contributions
Experiment conception and design were carried out by A.J. and C.D.-M. Experiment execution was done by A.J. and C.D.-M. Data analysis was performed by A.J., C.D.-M., and C.J. Discussion of the study was carried out by A.J., A.J., A.K., C.J., and C.D.-M. Article writing was carried out by A.J., C.D.-M., A.J., A.K., and C.J.
Acknowledgments
The authors are very grateful to Dr. F.J. Hemming for proofreading the article and to Drs. J.Gauthier, E. Ressiot, H. Michel, and C. Klein for selecting tinnitus participants.
Disclaimer
Part of this work has been deposited on BioRXiv under the reference BIORXIV/2019/676429 “A.J., A.K., C.J., A.J., C.D.-M., Specific connectivity with Operculum 3 (OP3) brain region in acoustic trauma tinnitus: a seed-based resting state fMRI study.”
Author Disclosure Statement
The authors declare that no competing financial interests exist.
Funding Information
This study was supported by French government grant number PDH1-SMO-3-0811 (A.J.). This study was performed on the IRMaGe platform member of France Life Imaging network (Grant ANR-11-INBS-0006).