
Abstract
Identifying the neural substrates underlying the personality traits is a topic of great interest. On the other hand, it is now established that the brain is a dynamic networked system that can be studied by using functional connectivity techniques. However, much of the current understanding of personality-related differences in functional connectivity has been obtained through the stationary analysis, which does not capture the complex dynamical properties of brain networks. In this study, we aimed at evaluating the feasibility of using dynamic network measures to predict personality traits. Using the electro-encephalography (EEG)/magneto-encephalography (MEG) source connectivity method combined with a sliding window approach, dynamic functional brain networks were reconstructed from two datasets: (1) resting-state EEG data acquired from 56 subjects; (2) resting-state MEG data provided from the Human Connectome Project. Then, several dynamic functional connectivity metrics were evaluated. Similar observations were obtained by the two modalities (EEG and MEG) according to the neuroticism, which showed a negative correlation with the dynamic variability of resting-state brain networks. In particular, a significant relationship between this personality trait and the dynamic variability of the temporal lobe regions was observed. Results also revealed that extraversion and openness are positively correlated with the dynamics of the brain networks. These findings highlight the importance of tracking the dynamics of functional brain networks to improve our understanding about the neural substrates of personality.
Introduction
Personality refers to a characteristic way of thinking, behaving, and feeling, that distinguishes one person from another (Back et al., 2009; Furr, 2009; Hong et al., 2008). Since personality traits are believed to be stable and broadly predictable (Canli and Amin, 2002; DeYoung, 2006), it is unsurprising that personality is linked to reliable markers of brain function (Yarkoni, 2014). In this context, the interest in the neural substrates underpinning personality has substantially increased in recent years. One of the most widely used and accepted taxonomies of personality traits is the five-factor model (FFM), or big-five model, which covers different aspects of behavioral and emotional characteristics (McCrae and John, 1992). It represents five main factors: conscientiousness, openness to experience, neuroticism, agreeableness, and extraversion.
On the other side, emerging evidence shows that most cognitive states and behavioral functions depend on the activity of numerous brain regions operating as a large-scale network (Bressler, 1995; Edelman, 1993; Fuster, 2010; Goldman-Rakic, 1988; Greicius et al., 2003; Mesulam, 1990; Sporns et al., 2004). This dynamic behavior is even present in the pattern of intrinsic or spontaneous brain activity (i.e., when the person is at rest) (Allen et al., 2014; Baker et al., 2014; de Pasquale et al., 2012, 2015; Kabbara et al., 2017; O'Neill et al., 2017). In particular, the dynamics of brain connectivity patterns can be studied at the millisecond time scale, for example using electro-encephalography (EEG) and magneto-encephalography (MEG).
However, although multiple studies have been conducted to relate the FFM traits to functional patterns of brain networks (Beaty et al., 2016; Li et al., 2017; Mulders et al., 2018; Tian et al., 2018; Tomeček et al., 2017; Toschi et al., 2018), we argue that the assessment of such relationships has been limited, in large part, due to an ignorance of networks variation throughout the measurement period. In this study, we hypothesized that investigating the dynamic properties of the brain network reconfiguration over time will reveal new insights about the neural substrate of personality. Our hypothesis was supported by many recent studies that demonstrate the importance of examining the temporal variations of brain networks in personality traits such as intelligence, creativity, executive function, and resilience (Kenett et al., 2020; Paban et al., 2019; Tompson et al., 2018).
Here, we tested our hypothesis on two datasets: (1) resting-state EEG data acquired from 56 subjects, and (2) resting-state MEG data provided from the publicly available Human Connectome Project (HCP) MEG2 release, including 61 subjects. Dynamic brain networks were reconstructed by using the EEG/MEG source connectivity approach (Hassan and Wendling, 2018) combined with a sliding window approach as in Kabbara and colleagues (2017), O'Neill and colleagues (2017), and Rizkallah and colleagues (2018). Then, based on graph theoretical approaches, several dynamic features were estimated. Correlations between individual FFM traits and network dynamics were assessed. Our findings reveal robust relationships between dynamic network measures and four of the big five personality traits (openness, conscientiousness, extraversion, and neuroticism).
Materials and Methods
The full pipeline of this study is summarized in Figure 1.
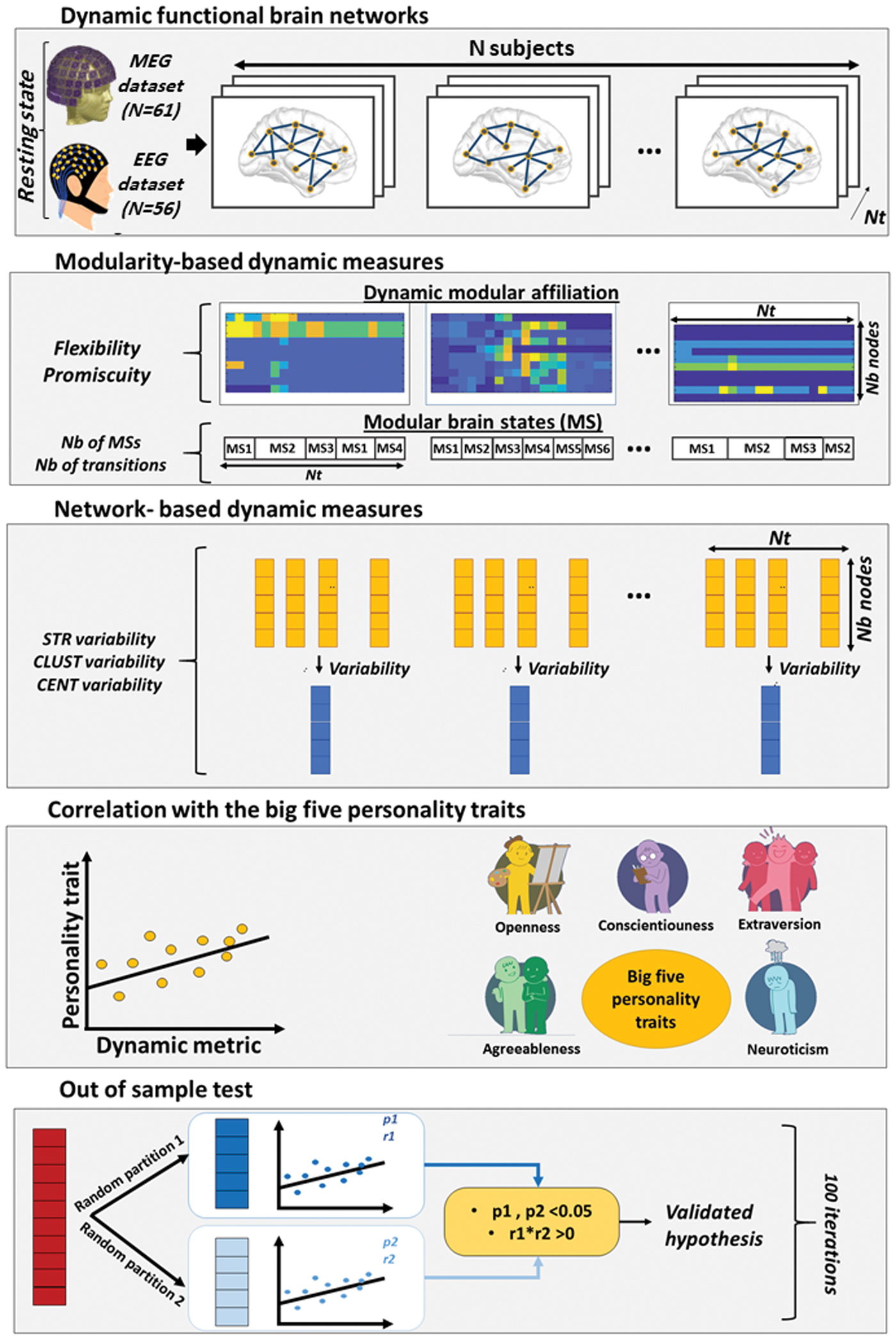
Full study pipeline. First, dynamic brain networks were reconstructed from resting-state EEG data of 56 participants and MEG data of 61 participants. Then, for each subject, dynamic features were extracted (modularity-based features and graph-based features). Correlations between FFM personality traits (agreeableness, extraversion, neuroticism, openness, conscientiousness) and the dynamic features were then evaluated. Finally, statistical tests were assessed by using a randomized out-of-sample test. CENT, betweenness centrality; CLUST, clustering coefficient; EEG, electro-encephalography; FFM, five-factor model; MEG, magneto-encephalography; STR, strength. Color images are available online.
Dataset 1: EEG dataset
Participants
A total of 56 healthy subjects were recruited (29 women). The mean age was 34.7 years old (SD = 9.1 years, range = 18–55). Education ranged from 10 years of schooling to a PhD degree. None of the volunteers reported taking any medication or drugs, nor suffered from any past or present neurological or psychiatric disease. The study was approved by the “Comité de Protection des Personnes Sud Méditerranée” (agreement no. 10–41).
EEG acquisition and preprocessing
Each EEG session consisted of a 10-min resting period with the participant's eyes closed (Paban et al., 2018). Participants were seated in a dimly lit room, were instructed to close their eyes, and then to simply relax until they were informed that they could open their eyes. Participants were informed that the resting period would last ∼10 min. The eyes-closed resting EEG recordings protocol was chosen to minimize movement and sensory input effects on electrical brain activity. The EEG data were collected by using a 64-channel Biosemi ActiveTwo system (Biosemi Instruments, Amsterdam, The Netherlands) positioned according to the standard 10–20 system montage, one electrocardiogram, and two bilateral electro-oculogram electrodes for horizontal movements. Nasion-inion and preauricular anatomical measurements were made to locate each individual's vertex site. Electrode impedances were kept below 20 kOhm.
The EEG signals are frequently contaminated by several sources of artifacts, which were addressed by using the same preprocessing steps as described in several previous studies dealing with EEG resting-state data (Kabbara et al., 2017, 2018; Rizkallah et al., 2018). Briefly, bad channels (signals that are either completely flat or contaminated by movement artifacts) were identified by visual inspection, complemented by the power spectral density. These bad channels were then recovered by using an interpolation procedure implemented in Brainstorm (Tadel et al., 2011) by using neighboring electrodes within a 5-cm radius. Epochs with voltage fluctuations between +80 and −80 μV were maintained. Five artifact-free epochs of 40-sec length were selected for each participant.
This epoch length was used in a previous study, and it was considered a good compromise between the needed temporal resolution and the results reproducibility (Kabbara et al., 2017).
Dynamic brain networks construction
Dynamic brain networks were reconstructed by using the “EEG source connectivity” method (Hassan and Wendling, 2018), combined with a sliding window approach as detailed in Kabbara and colleagues (2017, 2018) and Rizkallah and colleagues (2018). “EEG source connectivity” involves two main steps: (1) solving the inverse problem to estimate the cortical sources and reconstruct their temporal dynamics, and (2) measuring the functional connectivity between the reconstructed time-series.
Briefly, the steps performed were as follows: EEGs and magnetic resonance imaging (MRI) template (ICBM152) were coregistered through the identification of anatomical landmarks by using Brainstorm (Tadel et al., 2011). A realistic head model was built by using the OpenMEEG (Gramfort et al., 2010) software. A Desikan-Killiany atlas-based segmentation approach was used to parcellate the cortical surface into 68 regions (Desikan et al., 2006). The weighted minimum norm estimate (wMNE) algorithm was used to estimate the regional time series (Hamalainen and Ilmoniemi, 1994). The reconstructed regional time series were filtered in different frequency bands (delta: 1–4 Hz; theta: 4–8 Hz; alpha: 8–13 Hz; beta: 13–30 Hz; and gamma: 30–45 Hz). To compute the functional connectivity between the reconstructed regional time series, we used the phase locking value (PLV) metric (Lachaux et al., 2000) defined by the following equation:
where
Dynamic functional connectivity matrices were computed for each epoch by using a sliding window technique (Kabbara et al., 2017). It consists of moving a time window of certain duration δ along the time dimension of the epoch, and then PLV is calculated within each window. As recommended by Lachaux et al. (2000), the number of cycles should be sufficient to estimate PLV in a compromise between a good temporal resolution and a good accuracy. The smallest number of cycles recommended equals to 6. In each frequency band, we chose the smallest window length that is equal to
Functional connectivity matrices were represented as graphs (i.e., networks) composed of nodes, represented by the 68 regions of interest (ROIs), and edges corresponding to the functional connectivity values computed over the 68 regions, pairwise.
To ensure equal network density for all the dynamic networks computed across time, a proportional (density-based) threshold was applied in a way to keep the top 10% of connectivity values in each network.
Dataset 2: MEG dataset (HCP)
Participants
As part of the HCP MEG2 release (Larson-Prior et al., 2013; Van Essen et al., 2012), resting-state MEG recordings were collected from 61 healthy subjects (38 women). The release included 67 subjects, but 6 subjects were omitted from the analysis as their recordings failed to pass the quality control checks (including tests for excessive SQUID jumps, sensible power spectra, correlations between sensors, and availability of sufficient good quality recording channels). All subjects were young (22–35 years of age) and healthy.
MEG recordings and pre-processing
The acquisition was performed by using a whole-head Magnes 3600 scanner (4D Neuroimaging, San Diego, CA). Resting-state measurements were taken in three consecutive sessions of 6 min each. Data were provided pre-processed, after passing through a pipeline that removed artefactual segments, identified faulty recording channels, and regressed out artefacts that appear as independent components in an independent component analysis decomposition with clear artefactual temporal signatures (such as eye blinks or cardiac interference).
Dynamic brain networks construction
Here, we adopted the same pipeline used by the previous studies dealing with the same dataset (Colclough et al., 2016). Thus, to solve the inverse problem, we have applied a linearly constrained minimum variance beamformer (Van Veen et al., 1997). Pre-computed single-shell source models were provided by the HCP, and the data covariance was computed separately in the 1–30 and 30–48 Hz bands as in Colclough and colleagues (2016). Data were beamformed onto a 6-mm grid by using normalized lead fields. Then, source estimates were normalized by the power of the projected sensor noise. Source space data were filtered in delta: 1–4 Hz; theta: 4–8 Hz; alpha: 8–13 Hz; beta: 13–30 Hz; and gamma: 30–45 Hz (as in EEG dataset).
After obtaining the regional time series on the basis of the Desikan-Killiany atlas, a symmetric orthogonalization procedure (Colclough et al., 2015) was performed for signal leakage removal. To ultimately estimate the functional connectivity between regional time series, we used the amplitude envelope correlation measure (Brookes et al., 2012). This method briefly consists of (1) computing the power envelopes as the magnitude of the signal, using the Hilbert transform, and (2) measuring the linear amplitude correlation between the logarithms of ROI power envelopes. Finally, a sliding window (length = 6 sec, step = 0.5 sec) was applied to construct the dynamic connectivity matrices. This sliding window was previously used to reconstruct the dynamic networks derived from MEG data (O'Neill et al., 2016).
Also, matrices were thresholded by keeping the strongest 10% connections of each network.
Dynamic measures
Although functional connectivity provides crucial information about how the different brain regions are connected, the graph theory offers a framework to characterize the network topology and organization. In practice, many graph measures can be extracted from networks to characterize static and dynamic network properties. Here, we focused on measures quantifying the dynamic aspect of the brain networks/modules/regions and their reconfiguration over time.
Graph-based dynamic measures
Most previous studies attempt to average the graph measures derived from temporal windows (de Pasquale et al., 2015; Kabbara et al., 2017). However, such strategy constrains the dynamic analysis. Distinctively, we aimed here at quantifying the dynamic variation of node's characteristics inferred from graph measures (including strength, centrality, and clustering). The graph measure's variation
where
In this study we focused on three graph measures:
Strength: The node's strength is defined as the sum of all edges weights connected to a node (Barrat et al., 2004). It indicates how influential the node is with respect to other nodes.
Clustering coefficient: The clustering coefficient of a node evaluates the density of connections formed by its neighbors (Watts and Strogatz, 1998). It is calculated by dividing the number of existing edges between the node's neighbors by the number of possible edges. The clustering coefficient of a node is an indicator of its segregation within the network.
Betweenness centrality: The betweenness centrality calculates the number of shortest paths that pass through a specific node (Rubinov and Sporns, 2011). The importance of a node is proportional to the number of paths in which it participates.
An illustrative example of strength variability on a toy dynamic graph is presented in Figure 2B.
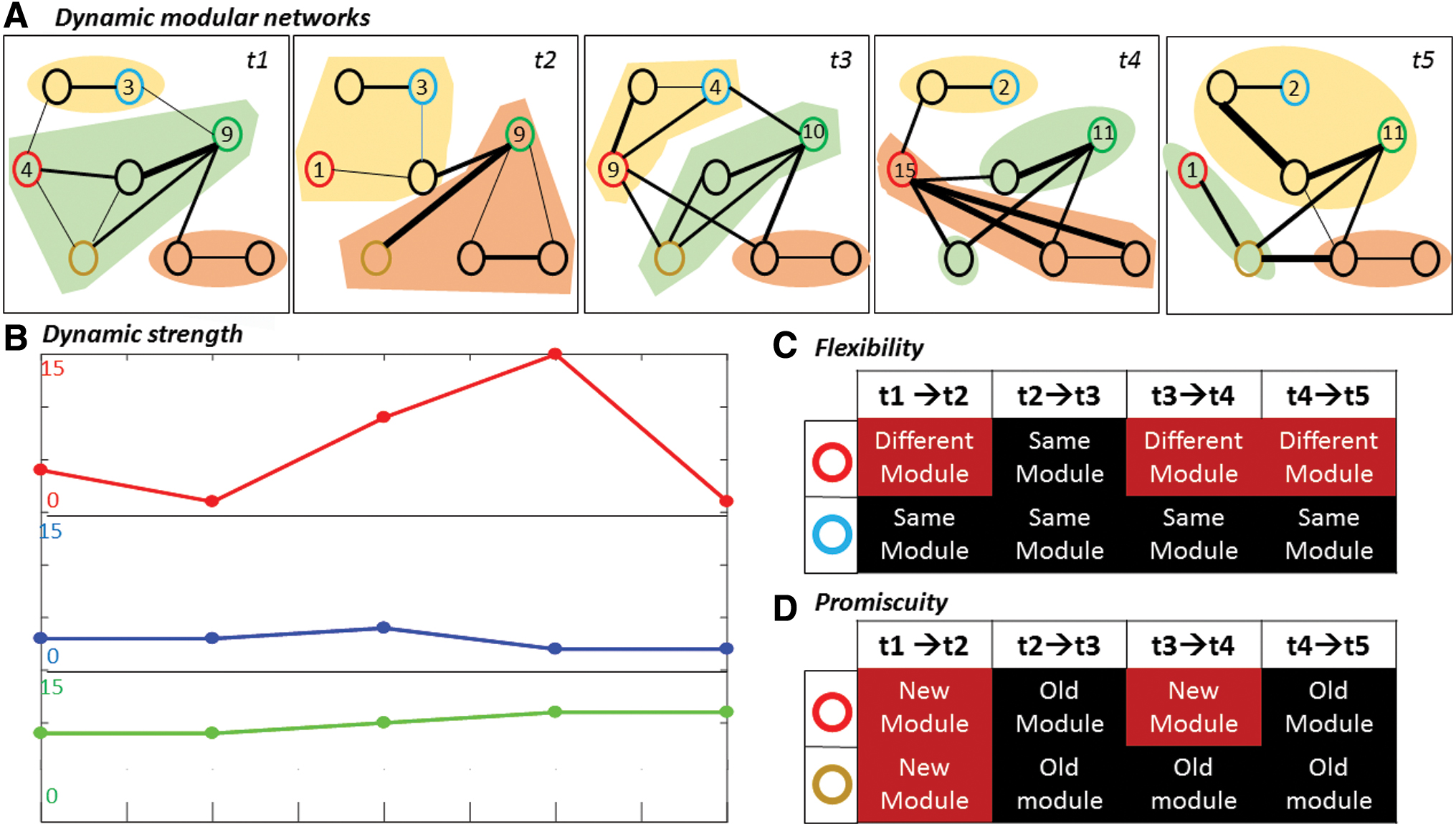
An illustrative example of the dynamic features extracted from a toy dynamic graph.
Modularity-based dynamic measures
Modularity describes the tendency of a network to be partitioned into modules or communities of high internal connectivity and low external connectivity (Sporns and Betzel, 2016). To explore how brain modular networks reshape over time, we detected the dynamic modular states (MSs) that fluctuate over time by using our recent proposed algorithm (Kabbara et al., 2019). Briefly, it attempts to extract the MSs that fluctuate repetitively across time. MSs reflect unique spatial modular organization, and they are derived as follows:
Decompose each temporal network into modules by using the consensus modularity approach (Bassett et al., 2013; Kabbara et al., 2017). This approach consists of generating an ensemble of partitions acquired from the Newman algorithm (Girvan and Newman, 2002) and Louvain algorithm (Blondel et al., 2008) repeated for 200 runs. Then, an association matrix of N × N (where N is the number of nodes) is obtained by counting the number of times two nodes are assigned to the same module across all runs and algorithms. The association matrix is then compared with a null model association matrix generated from a permutation of the original partitions, and only the significant values are retained (Bassett et al., 2013). To ultimately obtain consensus communities, we re-cluster the association matrix by using the Louvain algorithm.
Assess the similarity between the temporal modular structures by using the z-score of Rand coefficient, bounded between 0 (no similar pair placements) and 1 (identical partitions) as proposed by Traud and colleagues (2008). This yields a T × T similarity matrix where T is the number of time windows.
Cluster the similarity matrix into “categorical” MSs by using the consensus modularity method. This step combines similar temporal modular structures into the same community. Hence, the association matrix of each “categorical” community is computed by using the modular affiliations of its corresponding networks.
Once the MSs were computed, two metrics were extracted:
The number of MSs
The number of transitions: It measures the number of switching between MSs.
In addition, after obtaining the dynamic modular affiliations, two dynamic nodal measures were calculated:
Flexibility: It is defined as the number of times that a brain region changes its module across time, normalized by the total number of changes that are possible. We considered that a module was changed if more than 50% of its nodes have changed (Fig. 2C).
Promiscuity: It is defined as the number of modules that a node participates in during a period of time (Fig. 2D)
Statistical analysis
Dynamic measures were extracted at the level of each brain region (node-wise analysis), and at the level of the whole network. At the network level, flexibility, promiscuity, strength variation, clustering variation, and centrality variation were averaged over all brain regions. At the node level, the values of each node were maintained. To investigate the associations between the dynamic network measures and FFM personality traits, Pearson's correlation analysis was assessed. To consider the multiple comparisons problem (between the five frequency bands, five personality traits, and 68 ROIs), p values were corrected by using Bonferroni and false discovery rate (FDR) procedures (Bland and Altman, 1995). Bonferroni correction yields an adjusted threshold of
To avoid data dredging problem, we conducted randomized out-of-sample tests repeated 100 times. The out-of-sample test consists of randomly dividing data into two random subsets. If significant correlations were obtained from the two subsets for more than 95% of the iterations, the correlation was considered statistically significant on the whole distribution.
Evaluating the FFM personality traits
The five-factor model (FFM) represents five major personality traits: (1) conscientiousness, which describes an organized and detailed-oriented nature, (2) agreeableness, which is associated to kindness and cooperativeness, (3) neuroticism, which indexes the tendency to have negative feelings, (4) openness, which is related to intellectual curiosity and imagination, and (5) extraversion, which refers to the energy drawn from social interactions.
For the EEG dataset, personality traits were assessed with the French Big Five Inventory (BFI-Fr) (Plaisant et al., 2010). The BFI-Fr was composed by 45 items in which respondents decided whether they agreed or disagreed with each question, on a 1 (strongly disagree) to 5 (strongly agree) Likert scale. Responses were then summed to determine the scores for the five personality constructs.
According to the MEG dataset, the FFM personality traits were assessed via the NEO five-factors inventory (NEO-FFI) (McCrae and John, 1992; Terracciano, 2003). The NEO-FFI was composed by 60 items in which participants reported their level of agreement on a 5-point Likert scale, from strongly disagree to strongly agree.
Results
In each dataset, the dynamic functional networks were reconstructed by using a sliding window approach for each subject. Then, dynamic measures were extracted at the level of each brain region (node-wise analysis), and at the level of the whole network. At the network level, flexibility, promiscuity, strength variation, clustering variation, and centrality variation were averaged over all brain regions. At the node level, the values of each node were maintained.
Dataset 1: EEG
The correlation between FFM personality traits and the network-level parameters is presented in Figure 3. Neuroticism showed a negative correlation with the number of transitions (
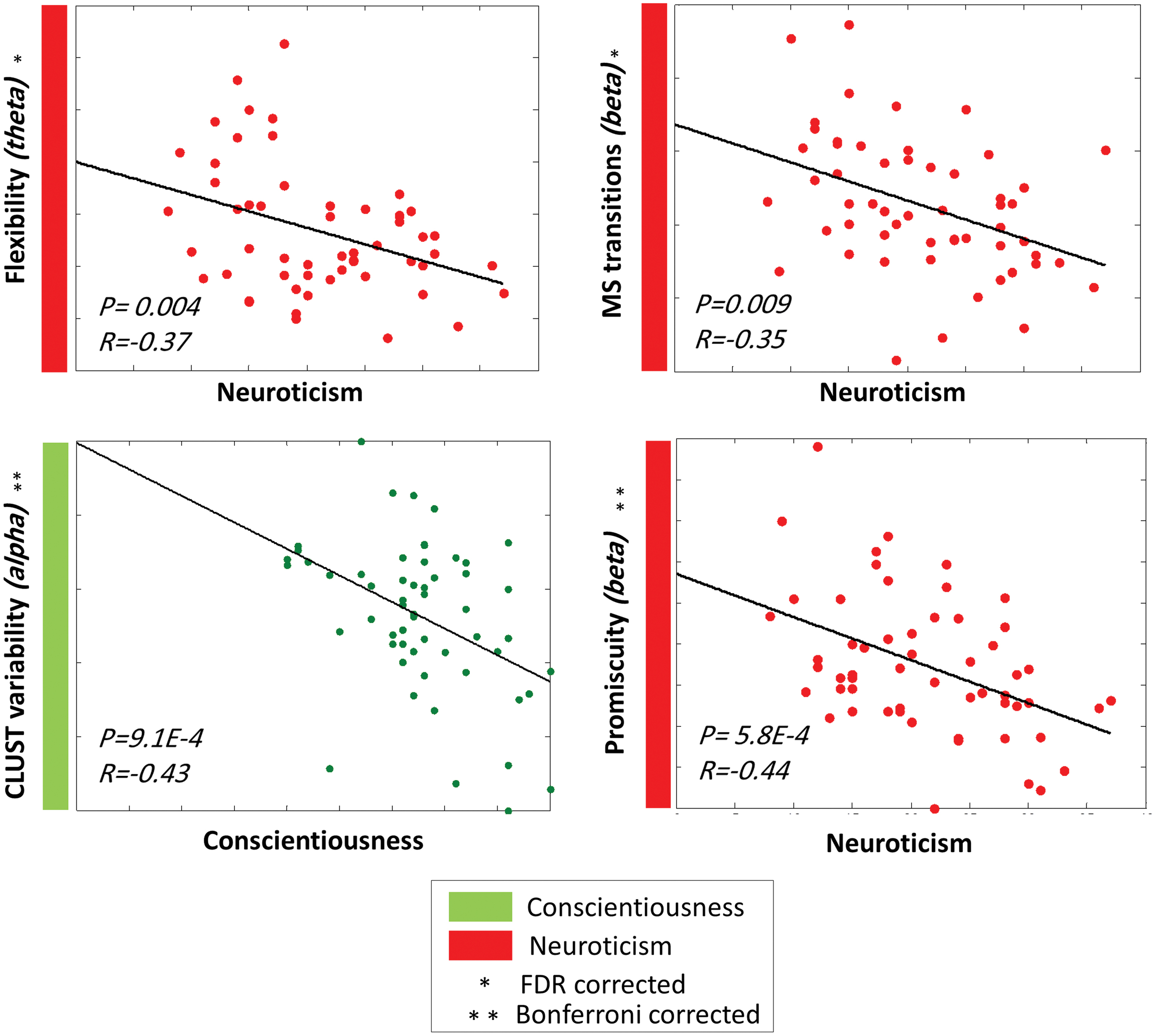
Significant correlations between the FFM traits and the dynamic graph measures computed on the network level by using the EEG dataset. Color images are available online.
Figure 4 illustrates the correlation between FFM traits and nodal characteristics in terms of dynamic features. Results show that higher extraversion was correlated with higher clustering variability of the superior parietal lobule (SPL) in the theta band (

The cortical surface illustrating the brain regions for which the dynamic measures significantly correlated with FFM traits by using the EEG dataset. FLEX, flexibility; FUS, fusiform; MTG, middle temporal gyrus; PCC, posterior cingulate cortex; SPL, superior parietal lobule; STG, superior temporal gyrus; TT, transverse temporal. Color images are available online.
Dataset 2: MEG
Figure 5 illustrates the correlation between FFM personality traits and network-level parameters for the MEG analysis. One can notice that neuroticism showed negative correlations with flexibility in the theta (
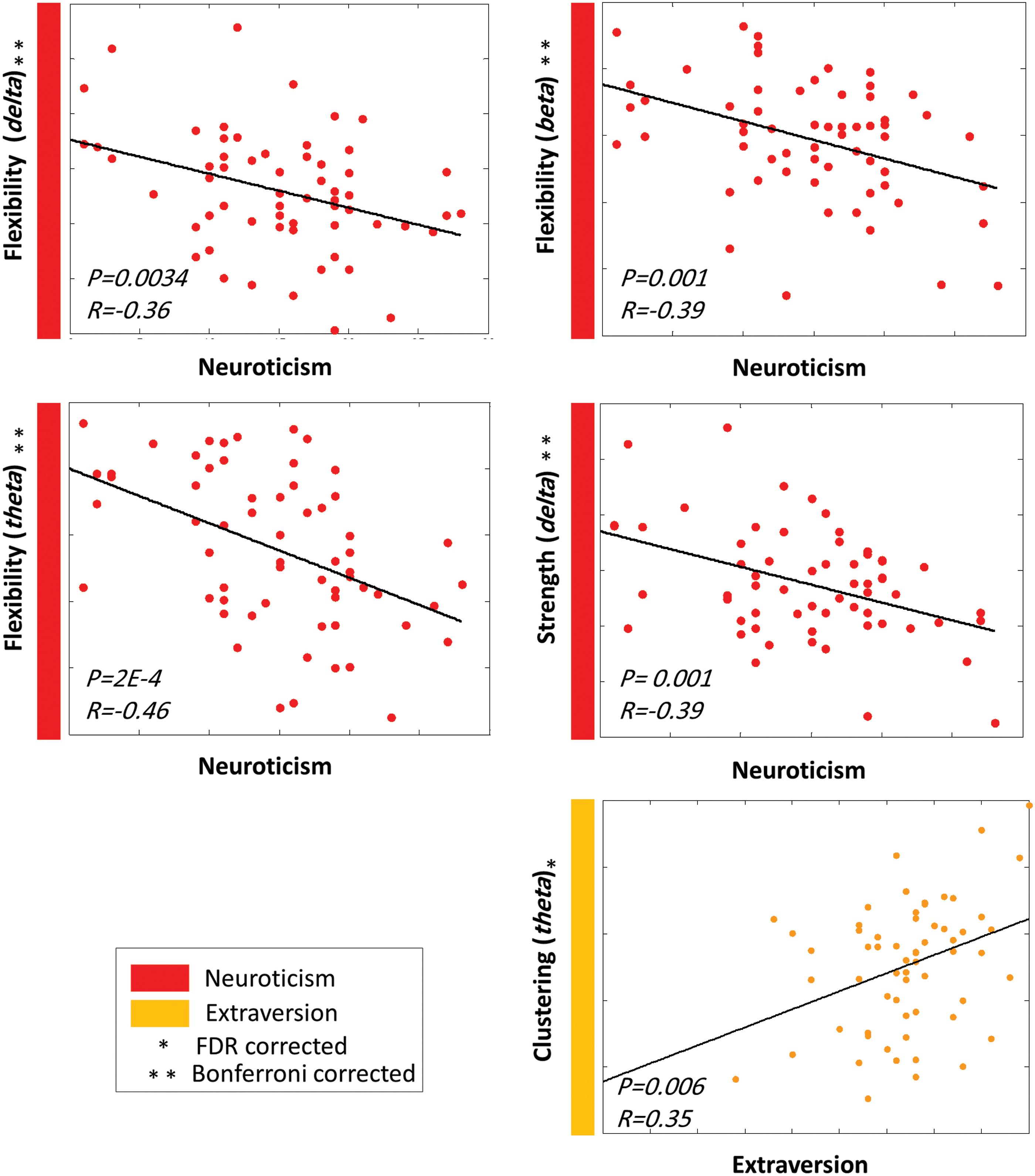
Significant correlations between the FFM traits and the dynamic graph measures computed on the network level by using the MEG dataset. Color images are available online.
Results in Figure 6 show that openness was positively correlated with the strength variability of the superior frontal gyrus in the beta band (PFDR corrected < 0:05; r = 0:48. However, negative correlations were observed between neuroticism and the strength variation of the left temporal pole in the alpha band (
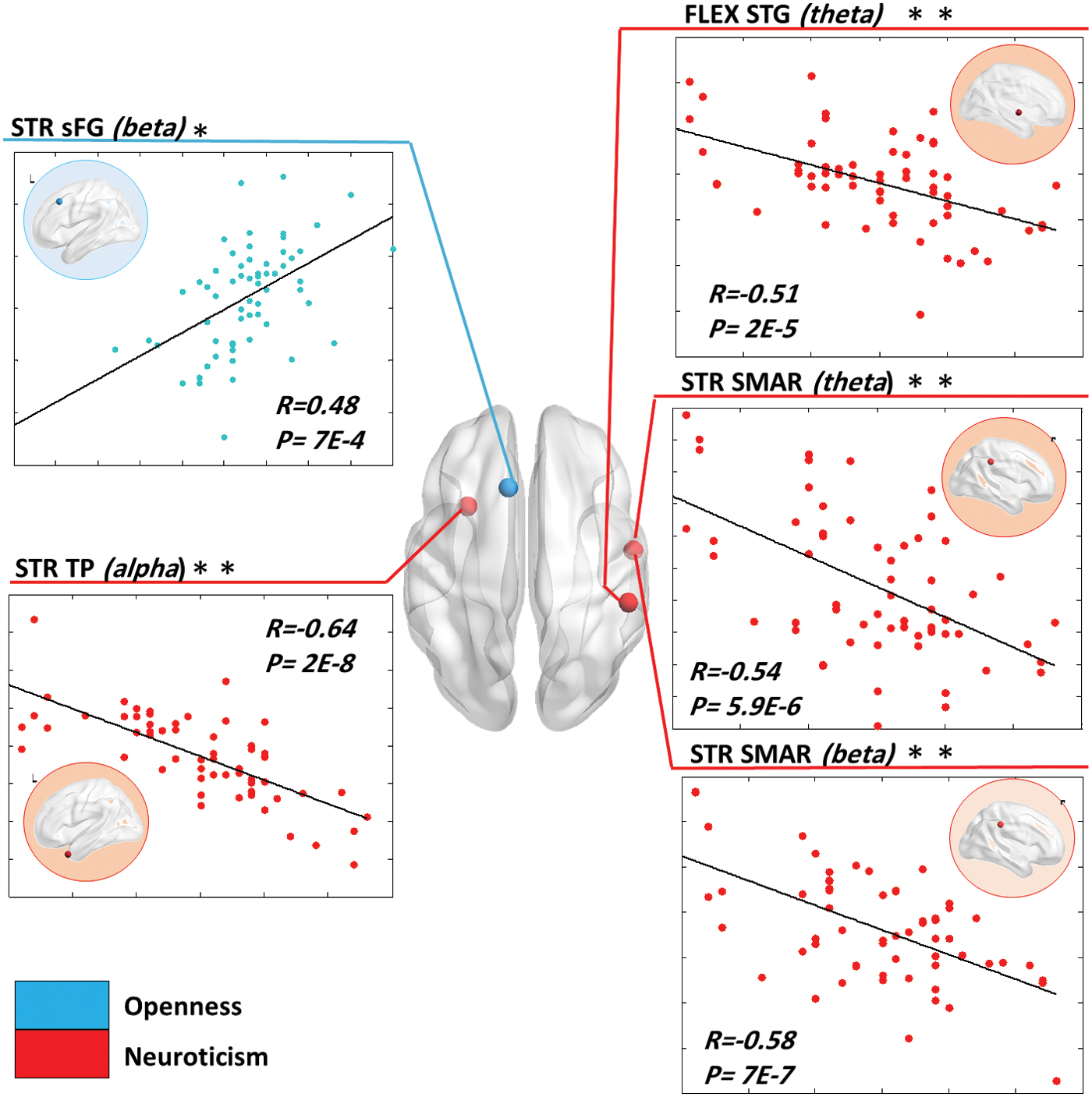
The cortical surface illustrating the brain regions for which the dynamic measures significantly correlated with FFM traits by using the MEG dataset. SMAR, supramarginal; TP, temporal pole. Color images are available online.
Randomized out-of-sample tests
For each feature, a distribution of 200 values (100 p values for each random subset) was obtained as a result of the correlation between the FFM personality traits and the network feature. We report in Tables 1 and 2 the results of randomized tests for all features mentioned as significant for the two datasets.
Results of the Randomized Tests for Dataset 1
A, alpha; B, beta; C, conscientiousness; D, delta; E, extraversion; MTG, middle temporal gyrus; N, neuroticism; O, openness; STG, superior temporal gyrus; SPL, superior parietal lobule; T, theta; TT, transverse temporal.
Results of the Randomized Tests for Dataset 2
sFG, superior frontal gyrus; SMAR, supramarginal; TP, temporal pole.
Discussion
This study provides evidence that dynamic features (derived from graph measures) based on resting-state EEG data are significantly associated with FFM personality traits (derived from the BFI-Fr questionnaire).
The majority of studies in personality has mainly examined the interaction between neuropsychological traits and brain features in a static way. In particular, multiple previous studies focused on investigating how personality traits are linked to differences in morphological brain properties (DeYoung, 2010; Gray et al., 2018; Liu et al., 2013; Omura et al., 2005; Riccelli et al., 2017). Another traditional way was to perform brain activation analysis to understand the neural basis of personality (Cooper et al., 2015; Falk et al., 2015). However, these strategies ignore useful information about the way in which brain regions interact with each other (Markett et al, 2018). Moving forward, multiple connectivity studies have been recently conducted to understand the neural substrates of human personality (Adelstein et al., 2011; Aghajani et al., 2013; Beaty et al., 2016; Bey et al., 2015; Dubois et al., 2018; Gao, 2013; Kyeong et al., 2014; Markett et al., 2013, 2016; Tompson et al., 2018).
Interestingly, graph theoretical assessment derived from networks was applied to link topological brain features to the Big Five personality traits (Beaty et al., 2016; Bey et al., 2015; Gao, 2013; Toschi et al., 2018). As an example, Toschi and colleagues (2018) show that conscientiousness is linked to nodal properties (clustering coefficient, betweenness centrality, and strength) of fronto-parietal and default mode network regions. Nevertheless, recent evidence revealed that dynamic analysis of functional data provides a more comprehensive understanding of neural implementation in personality (Tompson et al., 2018).
The main originality of this work is that it extends the traditional static view of brain networks to explore the time-varying characteristics associated to FFM traits. Particularly, we hypothesized that fast brain dynamics in EEG and MEG resting-state networks are correlated with FFM personality. Our hypothesis is based on many recent studies suggesting that personality-related differences in functional connectivity are discernable during rest (Adelstein et al., 2011; Beaty et al., 2016; Bey et al., 2015; Gao, 2013; Li et al., 2012, 2017; Markett et al., 2013; Mulders et al., 2018; Sheu et al., 2011). Such a finding is advantageous since collecting brain data during rest is more feasible. Also, this hypothesis is supported by the evidence that resting-state brain dynamics fluctuates at a sub-second timescale (less than 300 ms) (Baker et al., 2014; Damborská et al., 2019; Kabbara et al., 2017).
At the level of the whole network, both EEG and MEG analyses showed common observations according to the neuroticism personality trait.
This latter appeared to be the most sensitive to the analysis through dynamic approaches. Importantly, the EEG study showed negative correlations between neuroticism and centrality variation, number of transitions, promiscuity, and flexibility. Similarly, the MEG study showed negative correlations between neuroticism, flexibility, and strength variation. This suggests that the more individuals had a strong tendency to experience negative affection, such as anxiety, worry, fear, and depressive mood (Ormel et al., 2013), the less their brain showed dynamic characteristics in terms of modular organization over time. In other words, one may speculate that individuals with low dynamic measures of brain networks did not have enough capacity to get over their tendency to experience negative emotions and their psychological distress.
More particularly, at the node level, the degree of neuroticism was associated with low dynamic variation of temporal regions using the two modalities (mainly STG, MTG, and TT in EEG study; STG in MEG study). Importantly, the temporal lobe is known to be involved in processing sensory input related to visual memory, language comprehension, and emotion association (Kosslyn, 2007). In particular, the STG is involved in the interpretation of other individuals' actions and intentions (Pelphrey and Morris, 2006). Others stated that the STG plays an important role in emotional processing and effective responses to social cues, such as facial expressions and eye direction (Pelphrey and Carter, 2008; Singer, 2006). These findings are in agreement with a recent study showing that neurotic individuals present delayed detection of emotional and facial expressions (Sawada et al., 2016).
Using the MEG dataset, extraversion was showed to be positively correlated with the clustering variation of the whole network. Similar dynamic behavior was also found by using the EEG dataset where a positive correlation was established between extraversion and the clustering variation of SPL, which is involved in attention and visuomotor integration (Iacoboni and Zaidel, 2004). These findings highlight the complementary information that can be provided by the two modalities (de Pasquale et al., 2018). In line with Suslow and colleagues (2010) showing that extraverts displayed enhanced sensitivity and efficiency in sensory information processing compared with introverts, our data add to our neurobiological underpinning knowledge of extraversion highlighting the involvement of the SPL in such processes. Thus, the SPL would play a central role promoting segregation within the network of extraverted individuals.
Besides these similar observations led by both MEG and EEG analyses, conscientiousness revealed a significant correlation with dynamic metrics only using EEG, whereas openness showed a significant correlation with the dynamic measures using MEG solely. This discrepancy can be due to the fact that MEG-EEG differences particularly arise when investigating the transient resting-state functional connectivity patterns (Coquelet et al., 2020). It may also be due to the difference in the sample analyzed by the two modalities, as well as the pre-processing, source reconstruction, and connectivity methods used to reconstruct underlying networks. Moreover, several studies show that openness to experience and conscientiousness traits appear to differ across different samples (Hofstee et al., 1992; Johnson and Ostendorf, 1993).
Still, the impact of these differences was less drastic on the neuroticism and the extraversion traits. Importantly, these two traits are universally accepted and appear in all major models of personality traits (Zelenski and Larsen, 1999). Thus, the most consistent and significant result obtained shows that the dynamic flexibility in functional networks could plausibly contribute to increased emotional reactivity, particularly linked to neuroticism and extraversion (Yarkoni, 2014).
Results show that among the five frequency bands studied, most changes were observed within slow oscillations (namely, delta, theta, and alpha bands). As suggested by Knyazev (2012), these oscillations might play a major role in integration across diverse cortical sites by synchronizing coherent activity and phase coupling across spatially distributed neural assemblies, so that it might not be surprising that network properties related to personality traits were affected only within slower frequency bands.
Overall, this study adds to our recent paper (Paban et al., 2019) in providing new evidence that the dynamic reconfiguration of brain networks is of particular importance in shaping behavior.
Limitations
In this study, we have assessed the personality traits using FFM. One common limitation of FFM is that it does not provide an adequate coverage of all personality domains (McAdams, 1992). As an example, it lacks the description of religiosity, honesty, sense of humor, and many other domains. However, there is no consensus about the exact number of broad personality dimensions (Boyle, 2008). Second, FFM self-reports are sometimes subjective and may be influenced by many moderator factors such as cultures and situations (Boyle, 2008; The Five-Factor Model of Personality Across Cultures, 2002). Some studies also show that many personality traits (such as openness to experience and conscientiousness) are not replicable across different samples (Hofstee et al., 1992; Johnson and Ostendorf, 1993).
Despite all these limitations, the FFM has potentially been considered a useful structure for describing the personality constructs. Moreover, in this article, we have investigated the dynamic brain networks during resting state. We believe that the use of cognitive tasks that stimulate the related networks for each personality trait may advance our understanding of individual differences in dynamic network features.
Methodological considerations
First, in MEG analysis, the head model was computed from the individual MRI of each subject. Nevertheless, in EEG analysis, we used a template generated from MRIs of healthy controls, instead of a native MRI for EEG source connectivity. Recently, a study showed that there is no potential bias in the use of a template MRI as compared with individual MRI co-registration (Douw et al., 2018). In this context, a considerable number of EEG/MEG connectivity studies have used the template-based method due to the unavailability of native MRIs (Hassan et al., 2017; Kabbara et al., 2018; Lopez et al., 2014). However, we are aware that the use of subject-specific MRI is more recommended in clinical studies.
Second, we have adopted in each dataset the same pipeline (from data processing to networks construction) used by the previous studies dealing with the same datasets. Thus, for the EEG dataset, we used the wMNE/PLV combination to reconstruct the dynamic networks, as it is supported by two comparative studies (Mahmoud Hassan et al., 2014, 2016). For the MEG dataset, beamforming construction combined with amplitude correlation between band-limited power envelopes was sustained by multiple studies (Brookes et al., 2011; Brookes et al., 2012; Colclough et al., 2015, 2016; O'Neill et al., 2016).
Third, choosing the suitable window width is a crucial issue in constructing the dynamic functional networks. On the one hand, short windows do not contain sufficient information to accurately estimate connectivity. On the other hand, large windows may fail to capture the temporal changes of the brain networks. Hence, the ideal is to choose the shortest window that guarantees a sufficient number of data points over which the connectivity is calculated. This depends on the frequency band of interest that affects the degree of freedom in time series. In this study, we adapted the recommendation of Lachaux and colleagues (2000) in selecting the smallest appropriate window length that is equal to where 6 is the number of “cycles” at the given frequency band. The reproducibility of resting state results while changing the size of the sliding window was validated in a previous study (Kabbara et al., 2017).
Footnotes
Acknowledgment
The authors would also like to thank the Lebanese Association for Scientific Research (LASER) for its support.
Authors' Contributions
Author contributions included conception and study design (V.P., M.H.), data collection or acquisition (A.W., V.P.), statistical analysis (A.K., M.H., J.M.), interpretation of results (A.K., V.P., M.H., J.M.), drafting the article or revising it critically for important intellectual content (A.K., M.H., J.M., V.P.), and approval of the final version to be published and agreement to be accountable for the integrity and accuracy of all aspects of the work (all authors).
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was financed by the Rennes University, the Institute of Clinical Neuroscience of Rennes (Project named EEGCog), and AMU. The study was also funded by the National Council for Scientific Research (CNRS) in Lebanon.