
Abstract
During the early period of reading development, children gain phonological (letter-to-sound mapping) and semantic knowledge (storage and retrieval of word meaning). Their reading ability changes rapidly, accompanied by learning-induced brain plasticity as they learn to read. This study aims at identifying the neural bases of phonological and semantic processing in early childhood by using a combination of univariate and multivariate pattern analysis. Nineteen typically developing children between the age of five and seven performed visual word-level phonological (rhyming) and semantic (related meaning) judgment tasks during functional magnetic resonance imaging scans. Our multivariate analysis showed that young children with good reading ability have already recruited the left hemispheric regions in the brain for phonological processing, including the inferior frontal gyrus (IFG), superior and middle temporal gyrus, and fusiform gyrus. In addition, our multivariate results suggested that different sub-regions of the left IFG were recruited for the two tasks. Our results suggested the left lateralization of frontotemporal regions for phonological processing and semantic processing. In addition, we observed bilateral activations of parietal regions for semantic processing during early childhood. Our findings indicate that the neural bases of reading have already begun to be shaped in early childhood for typically developing children, which can be used as a control baseline for comparison of children at risk for reading difficulties.
Introduction
In today's society, learning to read as a child is the foremost step for developing high literacy skills. Teaching a child to read begins at birth with the reinforcement of pre-literacy skills, and most children officially learn to read between the ages of 5 and 7 years. Two common approaches to teach reading are sounding-out and sight-word reading methods. The sounding-out approach asks children to read aloud and to pronounce each letter or group of letters to recognize words by their sounds, which helps children to build letter-to-sound knowledge. Meanwhile, the sight-word approach requires children to memorize sight words or common vocabulary, which allows children to build their internal lexical dictionary. Thus, word reading can be achieved through grapho-phonological processing and lexico-semantic processing. The neural bases of these two processes have been studied mainly in older children and adults (Coltheart et al., 2001; Jobard et al., 2003; Vigneau et al., 2011), but not in young children (5–7 years old). Understanding the neural bases of early reading is critical for not only providing evidence on theoretical models of reading development but also building a control baseline to be used for examining how children with reading difficulties differ.
The two dominant theoretical models of reading are the dual-route cascaded (DRC) model (Coltheart et al., 2001) and the parallel-distributed-processing (PDP) connectionist model (Harm and Seidenberg, 2004). The DRC model suggests two distinct routes for word reading, including the grapho-phonological route transforming visual words into their sound representations (indirect route) and the lexico-semantic route transforming visual words into their meanings (direct route). According to the DRC model, skilled adult readers identify familiar words and words with irregular pronunciations such as “pint” by lexico-semantic processing (via the direct route) and pronounce newly encountered words and nonwords by rule-based grapheme-to-phoneme mapping (via the indirect route). In contrast, the PDP model postulates a single mechanism that generates pronunciation for all words. The PDP model suggests that word pronunciations are learned through repeated training with a corpus of written and spoken inputs. Different neural pathways have been identified to support the DRC model of reading (Friederici and Gierhan, 2013; Hickok and Poeppel, 2007; Price, 2012). A systematic meta-analysis of the DRC model of reading in adults' brain has proposed a dorsal route for grapho-phonological processing and a ventral route for lexico-semantic processing (Jobard et al., 2003). The dorsal route consists of the left superior temporal gyrus (STG), the left dorsal inferior parietal lobe (IPL, covering supramarginal gyrus, and angular gyrus), and the left opercular part of the inferior frontal gyrus (opIFG). The ventral route consists of the left fusiform gyrus (FG), the left basal inferior temporal area, the left posterior part of the middle temporal gyrus (MTG), and the left orbital part of IFG (orIFG). In addition, the triangular part of the left IFG (trIFG) has been suggested to be recruited in both routes. According to the PDP model, words are pronounced by using the same neural networks after a set of optimal connection weights have been learned (Binder et al., 2005; Harm and Seidenberg, 2004). The PDP model produces outputs stimulating children's learning behavior. In contrast, the DRC model suggests that young readers rely more on the indirect grapho-phonological route to translate letters into corresponding sounds and less on the direct lexico-semantic route to derive a meaningful representation of a given word. With practice over time, the young readers develop a larger internal lexicon dictionary storing words that can be recognized by sight via the lexical processing without the semantic processing. However, this study was not designed to differentiate between these two models. Instead, we hypothesize that both models would predict a lack of neural specialization of the two reading routes (direct vs. indirect route) for young children of 5–7 years of age.
The current literature on the neural representations of phonological and semantic processing in typically developing children is limited to late childhood (8–15 years old) (Bitan et al., 2007a, b; Blumenfeld et al., 2006; Cao et al., 2009; Hoeft et al., 2007; Li et al., 2018). Atypical brain structures and functions have been identified in children at risk for developmental dyslexia in early childhood (5–7 years old) (Im et al., 2016; Raschle et al., 2012, 2014; Wang et al., 2016; Yu et al., 2018) and even as early as in infancy (Langer et al., 2015; Zuk et al., 2019). However, there is limited knowledge of the neural bases of reading in typically developing children during early childhood when they begin to learn to read. This study will provide more evidence to improve our understanding of the neural bases of reading and might help us to identify children at risk for reading difficulties during early childhood. Early identification can lead to early intervention that is more effective at the time when brain plasticity is high.
For phonological processing, functional magnetic resonance imaging (fMRI) studies on older children (8 years old or older) have identified brain activation in the left IFG/superior frontal gyrus, left STG/MTG, bilateral FG, and bilateral middle frontal gyrus (MFG) by using English-word rhyme judgment tasks (Booth et al. 2004; Cao et al. 2006, 2008). These studies utilized high-level baseline tasks such as string matching as a control condition and have found left hemispheric brain activation in the fronto-temporal regions. However, a few studies have reported bilateral fronto-temporal activation by using a low-level task such as fixation as a control condition (Hoeft et al., 2006, 2007). For semantic processing, fMRI studies have identified the involvement of the left IFG/STG/MTG/FG during visual semantic decision tasks (Booth et al., 2001, 2003). However, Blumenfeld et al. (2006) used a semantic judgment task in 9- to 12-year-old children and observed that higher task accuracy was associated with more activation in posterior regions of the bilateral inferior temporal gyrus (ITG)/MTG. In contrast, lower task accuracy was associated with more activation in anterior regions of the bilateral IFG. Their findings suggested that the bilateral temporal cortices are related to better task performance during the meaning-based judgment tasks.
A few studies have directly compared brain activation for phonological and semantic processing in young children by using visual word pairs. One fMRI study directly compared brain activation between phonological and semantic processing and found greater brain activation in the left STG and angular gyrus comparing semantic processing versus phonological processing in 26 adolescents with a wide age range (9–19 years old) (Landi et al., 2010). However, other studies have reported that there were no significant differences between phonological and semantic processing in Chinese children (9–13 years old) (Cao et al., 2009; Liu et al., 2018). Their findings suggested that the neural representations of reading in children have not yet been specialized to the DRC model of reading. However, reading Chinese words is quite different from reading English words, which might contribute to the disparate findings. A recent longitudinal fMRI study provided neural evidence for the development of the DRC model of reading in older children (8–14 years old) (Younger et al., 2017). They reported that phonological decoding is initially used for reading, with a gradually decreased reliance on this strategy as reading becomes more automated for a child. Their fMRI results indicated that increases in reading ability were associated with decreases in brain connectivity of the dorsal route (indirect grapho-phonological route) and increases in brain connectivity of the ventral route (direct lexico-semantic route). In summary, past research suggests that the neural bases of reading supporting the DRC model are not specialized until adulthood, and the specialization of neural pathways is directly associated with individual reading ability.
The advancement of fMRI data analysis has led to the application of powerful pattern-classification algorithms to examine multi-voxel patterns of brain activation. The multi-voxel pattern analysis (multivariate pattern-analysis [MVPA]) has been suggested to be more sensitive and flexible for examining cognitive states (Norman et al., 2006). The conventional fMRI analysis uses a subtraction-based approach to identify brain activation related to certain experimental versus control conditions at a voxel-by-voxel level, which might overlook the brain activations if differences between conditions are across multiple voxels instead of a single voxel (Norman et al., 2006). To achieve greater sensitivity for discriminating conditions of interest with greater power and flexibility than the conventional univariate analysis (Kriegeskorte et al., 2006), we used searchlight analysis to identify differences of neural patterns across different conditions.
This study aimed at examining the neural bases of phonological and semantic processing in 19 young children (5–7 years old) by using fMRI during child-friendly visual rhyming and semantic judgment tasks. We used high-level control conditions such as other studies (Booth et al. 2004; Cao et al., 2006, 2008) for both tasks. For the visual rhyming task, we hypothesized the involvement of the left fronto-temporal activation based on past research studies (Booth et al. 2004; Cao et al. 2006, 2008). For the visual semantic judgment task, we hypothesized that bilateral fronto-temporal brain regions would be involved based on another study (Blumenfield et al., 2006). The use of both univariate and MVPA analyses can provide important complementary information (Poldrack and Farah, 2015). Thus, we conducted both univariate analysis and MVPA to understand the neural bases of phonological and semantic processing during early childhood.
Materials and Methods
Participants
Sixty healthy native English-speaking children were recruited for the study. The study involved two visits. During the first visit, a series of standardized behavioral tests were administered. The participant's parent was asked to fill out a questionnaire about the child's developmental history. Based on the questionnaire, participants with (1) a diagnosis of attention deficit hyperactivity disorder, (2) hearing or vision impairment, (3) neurological or psychiatric disorders, (4) a diagnosis of language disorder or reading disability, or (5) with any contraindications to be scanned in an MRI were excluded from the study and not invited for the second visit. The inclusion criteria for the second visit (imaging session) were: (1) native monolingual English speakers, (2) right-handed as measured by handedness questionnaire (Oldfield, 1971), (3) above 80 standard score of the non-verbal intelligence measured by the Kaufman Brief Intelligence Test, Second Edition (Kaufman, 2004), and (4) accuracy >70% in the familiarity task. The computer-based familiarity task was designed to ask the participant to read thirty 3–5 letter monosyllabic words and choose the corresponding picture that represents the word. The purpose of this customized picture naming task was to make sure that the participant would be able to complete both rhyming and semantic judgment tasks during fMRI scans. Twenty participants were invited back for the fMRI session based on the inclusion criteria mentioned earlier. The fMRI data from one participant were excluded due to excessive motion artifacts. Nineteen participants were included in the final analysis for this study. They were all right-handed (8 M, 11 F; mean age of 6.55 years old with age range from 5 year 4 months to 7 year 9 months). The Institutional Review Board approved all experimental procedures at the University of Nebraska at Lincoln. Written consent forms were obtained from the parent or guardian, and written assent forms were obtained from children who were older than 7 years old.
Behavioral measures
Behavioral measures were acquired during the first visit. Phonological awareness (PA) was measured by using the PA subtest of Woodcock Reading Mastery Test (WRMT)-III (WRMT-III-PA) (Woodcock, 2011), which consists of five sections, including first-sound matching, last-sound matching, rhyme production, blending, and deletion tasks. Word reading ability was measured by using the word identification subtest of WRMT-III (WRMT-III-WID) (Woodcock, 2011). Word association skill was measured by the word classes sub-test of the core language score in the Clinical Evaluation of Language Fundamentals (CELF)-5 (Wiig et al., 2013). The CELF score was only administered for 12 participants.
Imaging data acquisition
Paradigms
Three- to five-letter monosyllabic words suitable for young children were selected from the Medical Research Council psycholinguistic database (Wilson, 1988) to generate word-pair stimuli matched for concreteness, printed familiarity, word type (noun), and the number of syllables. Both rhyming and semantic judgment tasks presented word stimuli visually with a child-friendly image above the corresponding words (Fig. 1). Each word was presented for a duration of 800 msec with a 200-msec gap between word-pair presentations. Then, a 2200-msec response screen followed after the presentation of each word-pair stimulus, during which the word-pair stimulus remained on the screen along with a question mark in the middle of the two words (Fig. 1). Participants were instructed to respond as soon as they saw the question mark. For the rhyming judgment task, the participants were instructed to press the right button (color red) if the two words rhyme and press the left button (color blue) if the two words do not rhyme. For the semantic association judgment task, the participant pressed the right button (color red) if the two words associated semantically and pressed the left button (color blue) if they did not associate semantically. The control conditions were the same for both tasks and consisted of three to five non-alphabetic glyph characters of symbol strings. The two strings were also presented with child-friendly images to match the task condition for controlling visual inputs. During the control condition, the participants were asked to determine whether the symbol strings were matched or not. In addition, there were 10-sec baseline conditions consisting of a black cross on white screen. The participants were instructed to press either the left or right button as soon as the black fixation cross turned red (Fig. 1).

Schematic representation of functional magnetic resonance imaging task design illustrating the rhyming task
We administered two runs for each paradigm to minimize the effects of fatigue. A block design was used to achieve higher detectability of brain activation in pediatric fMRI data (Wilke et al., 2003). Each run consisted of eight blocks (four task blocks and four control blocks) starting with a fixation block (baseline) followed by a task block, a fixation block, and a control block (Fig. 1D). The order of runs was counterbalanced across subjects, such that nine participants completed the rhyming paradigm first, whereas the other 10 completed the semantic paradigm first. Each rhyming block consisted of eight conditions (four rhyming and four non-rhyming). Each semantic block comprised eight conditions (four semantically associated and four non-associated). Each control block consisted of four conditions (two matching strings and two non-matching strings). Within a block (either task or control block), stimuli were randomized. At the beginning of each run, a 5-sec fixation was presented as a baseline block and was not included in the fMRI data analysis to eliminate non-equilibrium effects of magnetization. Each run lasted for 4 min 37 sec (277 sec); each run consisted of 32 task conditions, 16 control conditions, and 8 fixation conditions.
Imaging acquisition protocol
Brain imaging data were acquired by using a 3.0-T Skyra Siemens scanner with a 64-channel head coil. The blood-oxygen-level-dependent (BOLD) signal was measured by using a multiband echo planar imaging (EPI) (University of Minnesota sequence cmrr_mbep2d_bold) sequence with the following parameters: repetition time (TR) = 1000 msec, time of echo (TE) = 29.80 msec, flip angle = 60°, matrix size = 210 × 210 mm2, field of view = 210 mm, slice thickness = 2.5 mm, number of slices = 51, and voxel size = 2.5 × 2.5 × 2.5 mm3. Before functional image acquisition, a high-resolution T1-weighted three-dimensional structural image was acquired for using a multi-echo magnetization prepared rapid gradient echo sequence (TR = 2530 msec, TE1 = 1.69 msec, TE2 = 3.55 msec, TE3 = 5.41 msec, TE4 = 7.27 msec, matrix size = 256 × 256 mm2, field of view = 256 mm, slice thickness = 1 mm, number of slices = 176, and TA = 6.03 min).
Before the imaging session, each participant underwent 30-min practice training in a child-friendly “MRI-like” room (a mock scanner room equipped with a nonmagnetic MRI simulator, Model#100355; Psychology Software Tools, Inc.). The literature suggests that pre-training sessions are crucial for young children, increase the success rates, and reduce the motion artifacts and anxiety (Leach and Holland, 2010). We achieved a 95% success rate by using a mock session. During the mock session, participants were exposed to different scanner noise and practiced the experimental tasks until they achieved an accuracy above 60%. The practice version of the experimental tasks used words that were different from the actual stimuli. During the actual imaging session, each participant was laid on the scanning table with his/her head secured with foam pads. A two-button box was placed on each side of the participant, and responses were collected through E-prime 2 (Psychology Software Tools, Inc.). We attached elastic straps across the head-coil apparatus to reduce the head motions. The state-of-art OptoActive™ active noise canceling headphones (OptoAcoustics, Mazor, Israel) was used to minimize the effects of the ambient scanner noise.
Imaging data analyses
Univariate analysis
Standard preprocessing steps were performed in SPM12 (Wellcome Department of Cognitive Neurology, London;
Multivariate pattern analysis
We employed a whole-brain searchlight analysis that is a recently developed MVPA technique for identifying locally informative areas of the brain. The searchlight analysis outperforms mass-univariate analyses due to its higher sensitivity to distributed information coding (Kriegeskorte et al., 2006). We performed searchlight analysis by using a linear discriminant analysis classifier implemented in CoSMoMVPA toolbox (Oosterhof et al., 2016). Two separate searchlight analyses were conducted to examine the spatial pattern of voxels in the brain that the classifier could reliably distinguish between (1) rhyme task from control task, and (2) semantic task from control task. The first-level SPMs from each run were stacked together into the searchlight analysis with 100 voxel searchlight spheres across the whole brain. Classification accuracies were obtained by using a leave-one-out cross-validation method with an eight-fold partitioning scheme for each subject. For each run, the dataset was split into eight chunks (each corresponding to one experimental block), and the classifier was trained on the data from seven chunks and tested on the remaining one. The procedure was repeated for eight iterations, using all possible train/test partitions. The average decoding accuracies across these iterations were calculated. At the group level, we performed a two-tailed one-sample t-test across individual maps where classification was significantly above chance (50%, since our classifiers were binary). The resulting SPMs were corrected for multiple comparisons by using a cluster-based Monte Carlo simulation algorithm with 1000 iterations implemented in the CoSMoMVPA toolbox (cluster-corrected threshold α = 0.01, two-tailed; z > 1.96, p < 0.05).
Results
Behavioral measures
The mean PA standard score of the 19 participants was 113.16 with a standard deviation (SD) of 13.85. The mean standard score of WRMT-III-WID was 112.76 with an SD of 14.41. The CELF word association subtest acquired for 12 participants had a mean scaled score of 12.83 with an SD of 2.37. The in-scanner performance data of accuracy and reaction time (RT) (for correct responses) are plotted in Figure 2 and summarized in Table 1. Children performed better in control conditions as compared with task conditions. A paired-sample t-test revealed that the accuracy was significantly higher in the control task than the rhyming task [t(18) = −3.23, p < 0.05] and the semantic task [t(18) = −3.52, p < 0.05]. The RT was significantly lower in the control task than the rhyming task [t(18) = 3.24, p < 0.05] and the semantic task [t(18) = 2.91, p < 0.05]. There were no significant differences in accuracy and RT between the rhyming and semantic judgment tasks.
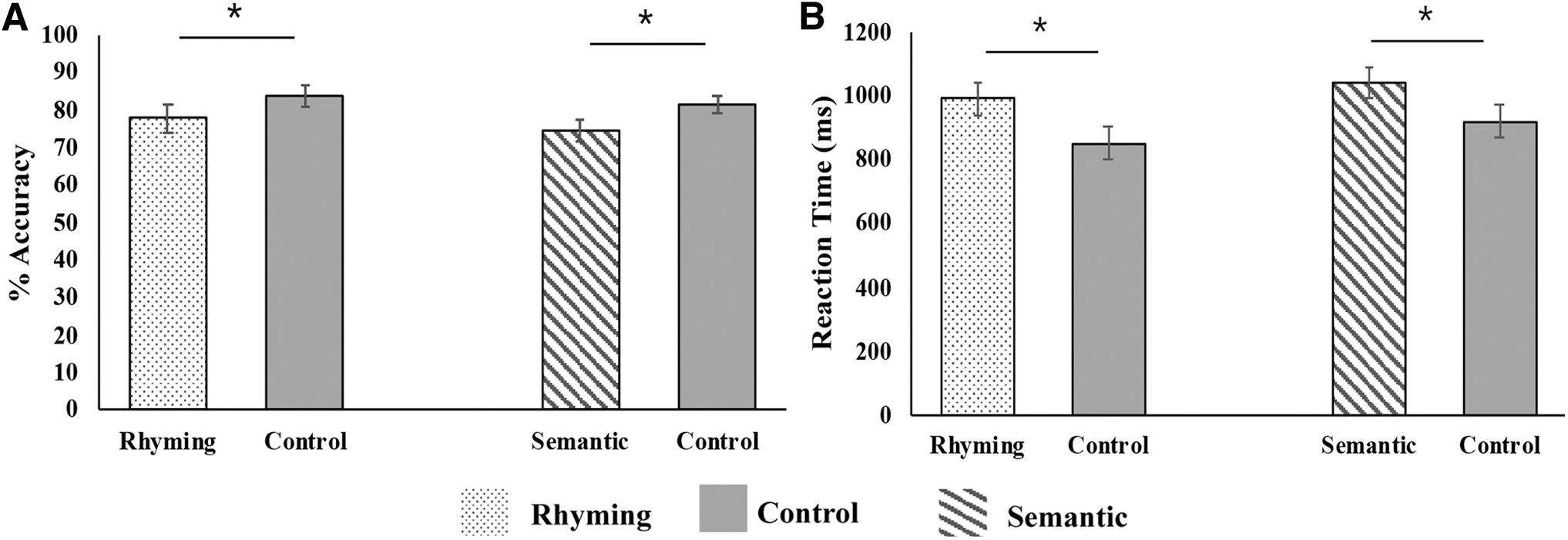
Average (N = 19) in-scanner task accuracy
In-Scanner Performance
Univariate analysis
The univariate results of rhyme and semantic judgment tasks are summarized in Tables 2 and 3 (Fig. 3). The contrast of rhyme > control identified significant activations in the left frontal regions, including the left IFG covering opIFG, trIFG, and orIFG, the left supplementary motor area (SMA), and the left precentral gyrus (Fig. 3A; Table 2). The contrast of semantic > control showed significant activations in the left IFG (opIFG, trIFG, orIFG), which also extended to the left precentral gyrus, left MFG, left SMA, and to the right IFG (trIFG and orIFG) and right precentral gyrus. There were significant activations in the bilateral temporo-parietal regions covering the right MTG/ITG and FG, as well as IPL including the supramarginal gyrus, angular gyrus, and precuneus (Fig. 3B; Table 3).
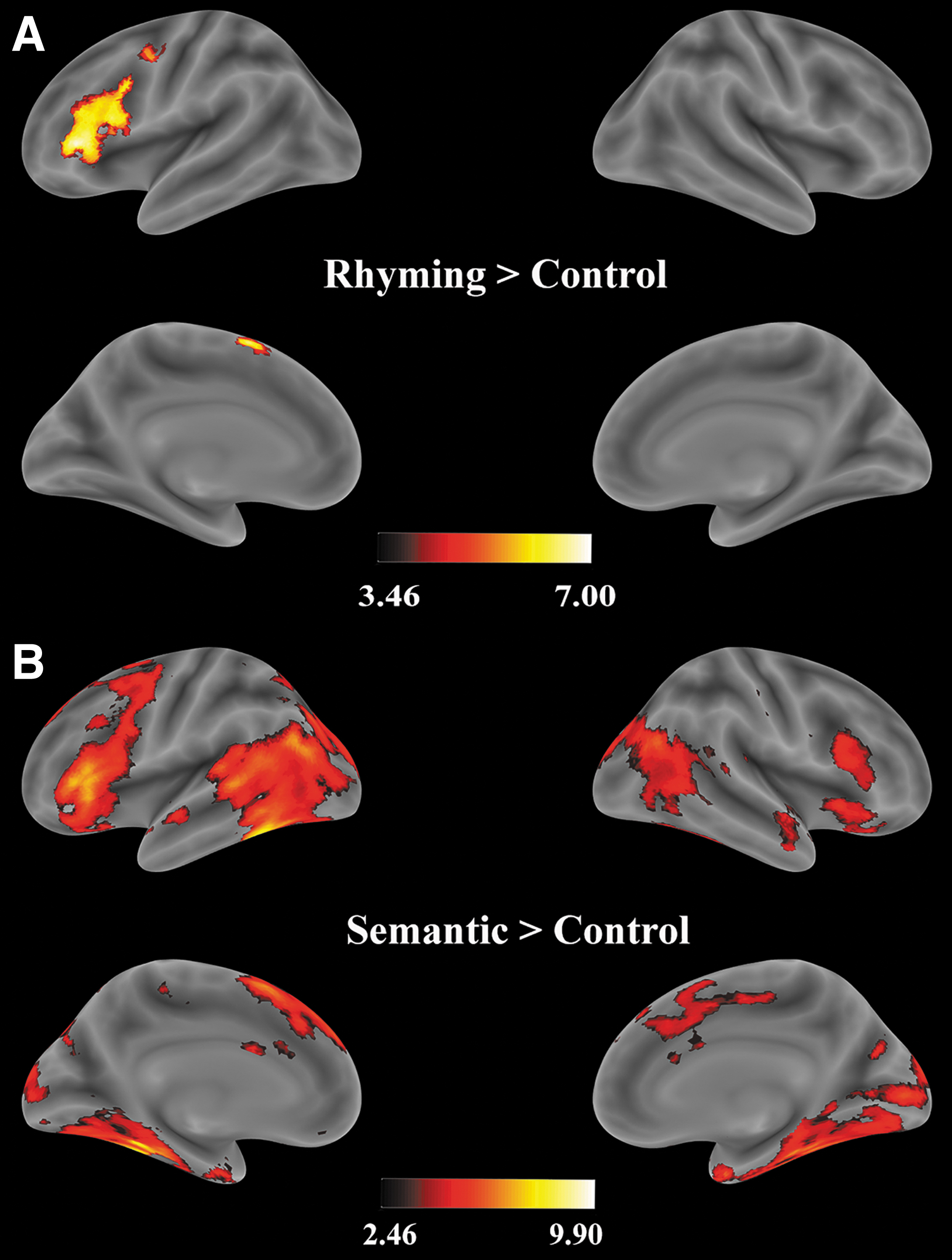
Voxel-wise significant activation, within the whole-brain anatomical mask for the contrast
Univariate Results for the Contrast Rhyme Versus Control
p-Value survived cluster-based (k > 20) FDR correction (q < 0.05).
BA, Brodmann area; FDR, false discovery rate; IFG, inferior frontal gyrus (opIFG, trIFG, orIFG); MNI, Montreal Neurological Institute; opIFG, opercular part of IFG; orIFG, orbital part of IFG; trIFG, triangular part of IFG; SMA, supplementary motor area.
Univariate Results for the Contrast Semantic Versus Control
p-Value survived cluster-based (k > 20) FDR correction (q < 0.05).
IFG, inferior frontal gyrus (opIFG, trIFG, orIFG); MFG, middle frontal gyrus; IPL, inferior parietal lobe; MTG, middle temporal gyrus; ITG, inferior temporal gyrus.
The direct comparison between the rhyming and semantic task did not show any significance in either direction (rhyming > semantic or rhyming < semantic) after FDR correction (q < 0.05) with a cluster threshold k > 20. The conjunction analysis of the two contrasts (rhyme vs. control and semantic vs. control) revealed an overlap in activations in the left IFG and the left SMA after FDR correction (q < 0.05) with a cluster threshold k > 20 (Supplementary Fig. S1; Supplementary Table S1).
Multivariate analysis
For rhyme versus control, searchlight analysis identified significant decoding accuracy in the left fronto-temporal regions, including the left opIFG, left pre/postcentral gyrus, and the left MTG/STG/ITG (including FG) (Fig. 4A; Table 4). For semantic versus control, searchlight analysis found significant decoding accuracy in the left fronto-temporal and bilateral parietal regions. Among the frontal regions, the left IFG (opIFG, trIFG, orIFG) cluster extended to the left precentral gyrus, and the left MFG. Among the temporal regions, a cluster extending from the left STG/MTG to the left ITG (including FG) significantly decoded semantic conditions from control conditions. The temporal cluster also extended to the bilateral parietal regions (IPL: supramarginal gyrus, angular gyrus, and precuneus) (Fig. 4B; Table 5).
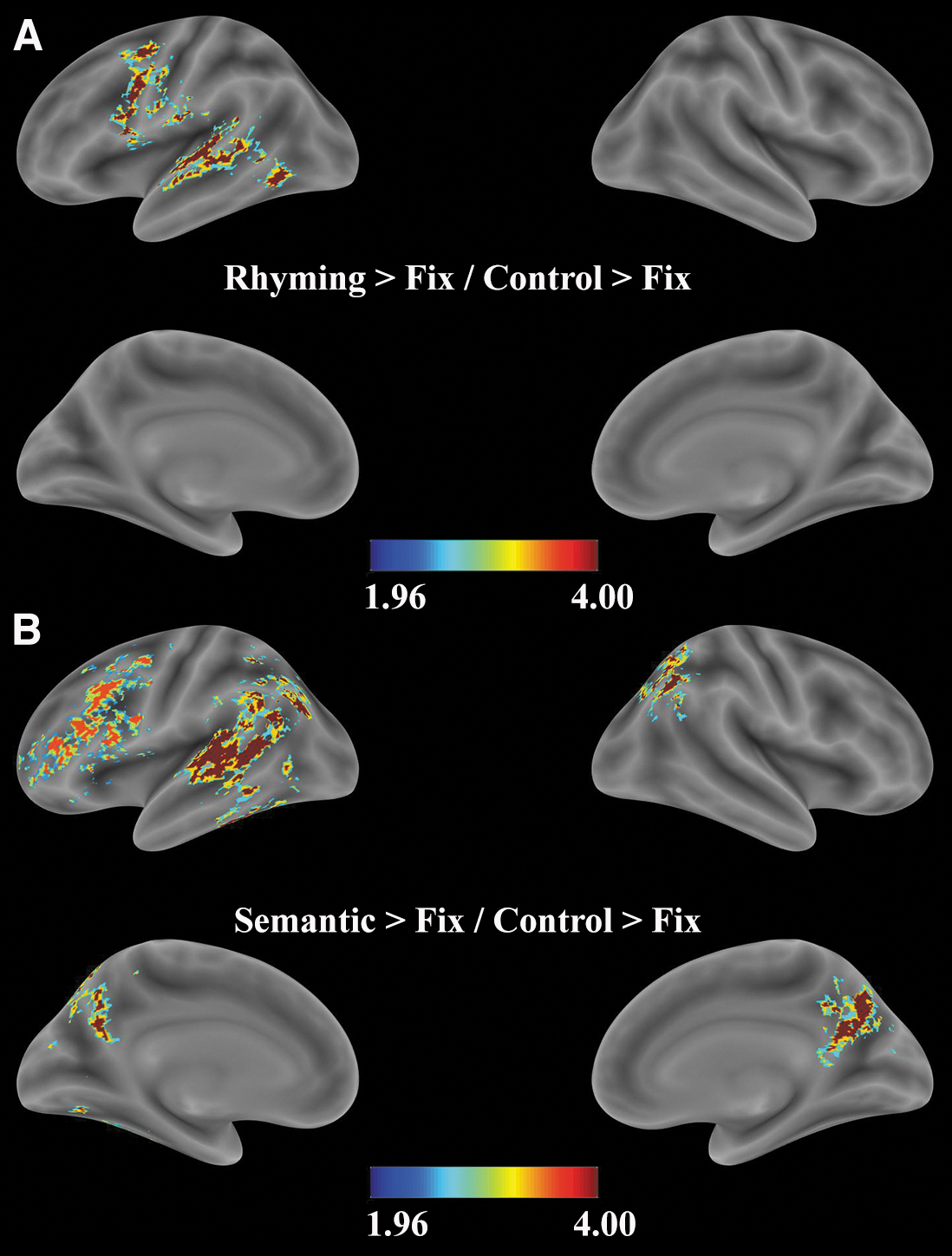
Multivariate pattern analysis results. Statistical group maps for the two-searchlight analysis performed with a 100-voxel searchlight by using a linear discriminant analysis classifier to identify regions that significantly decode above chance
Multivariate Pattern-Analysis Searchlight Results for Rhyme Versus Control
The table just cited lists the brain areas that successfully classify rhyme from control condition at a cluster-corrected threshold p < 0.05.
IFG, inferior frontal gyrus (op IFG); STG, superior temporal gyrus.
Multivariate Pattern-Analysis Searchlight Results for Semantic Versus Control
The table just cited lists the brain areas that successfully classify semantic from control condition at a cluster-corrected threshold p < 0.05.
IFG, inferior frontal gyrus (opIFG, trIFG, and orIFG).
Discussion
While performing the visual phonological judgment tasks, children have to access the sounds of the visually presented words and are therefore more likely to engage the grapho-phonological route of reading (Bitan et al., 2006, 2007a, b; Booth et al., 2007; Cao et al., 2006, 2009; Hoeft et al., 2007). Semantic association tasks require children to decide whether the two words presented are related and thus are more likely to engage in the lexico-semantic route of reading (Blumenfeld et al., 2006; Booth et al., 2001, 2003, 2004). In this study, we examined the neural representations of early reading skills by using visual rhyming and semantic judgment tasks during fMRI in young children (5–7 years old). Our univariate results indicated that the phonological processing in the brain had a left lateralized activation pattern, whereas the semantic processing in the brain involved a bilateral activation pattern. However, our MVPA results suggested that both processes in the brain involved left-lateralized fronto-temporal activations. This study provided evidence that young children with good reading ability have already established the grapho-phonological route for reading in the left hemisphere to support early stages of reading, including the left IFG, the left STG/MTG, and the left FG. In addition, our results suggested that the lexico-semantic processing also relies on the left IFG, the left STG/MTG, and the left FG, as well as the bilateral parietal regions covering IPL. The MVPA results provided a detailed account of neural representations of IFG sub-regions and suggested that the left opIFG was specifically involved in phonological processing, whereas none of the IFG sub-regions was specialized for semantic processing. In addition, the bilateral parietal regions showed specialization for semantic processing. All these findings are discussed later in greater detail.
Frontal lobe
Our univariate results showed left lateralization in the IFG, SMA, and precentral gyrus for both tasks and no significant differences through a direct comparison between tasks at FDR-corrected level. In line with a recent study on 35 typically developing children (5–6 years old) (Weiss et al., 2018), our direct comparison results supported that there is no specialization in the frontal regions yet during early childhood. However, our multivariate results indicated that phonological processing and semantic processing involved different sub-regions of the left IFG. The left opIFG was recruited for phonological processing, whereas no IFG subregions were specialized for semantic processing (Tables 4 and 5). In adults, it has been reported that phonological processing relied on the left opIFG (Jobard et al., 2003). Moreover, real-time transcranial magnetic stimulations to the left opIFG disrupted phonological processing (Gough et al., 2005), suggesting that the left opIFG as the posterior part of the left IFG was involved in phonological processing. Our MVPA results also supported that the left IFG is not a single function region, as the left IFG sub-regions (opIFG, triIFG, orIFG) were involved in semantic processing as well. Different from our univariate results, our MVPA findings indicated that young typically developing children already show some specialization of the sub-regions of the left IFG for phonological processing by the age of 5–7 years. The differences in results between the univariate and multivariate analyses may be caused by the higher sensitivity and greater power of the MVPA approach (Kriegeskorte et al., 2006). The MVPA compares the representation patterns of activity across conditions, whereas the univariate analysis compares spatial-average activation across conditions.
Both univariate and multivariate results showed left lateralization for IFG for phonological processing. Booth et al. (2004) used visual rhyming tasks in both adults and older children and only found activation in the right IFG in adults. Our findings on the left IFG are aligned with their results, suggesting that young children recruit a left lateralized activation pattern in the IFG during the visual rhyming task.
Temporal lobe
Previous research has identified the function of left MTG for semantic processing in both young (5–6 years old) and older children (9–12 years old) (Bitan et al., 2007b; Blumenfeld et al., 2006; Booth et al., 2004; Weiss et al., 2018). Studies that have directly compared phonological and semantic processing in adults' brains have reported a double dissociation between the tasks in the temporal regions (Jobard et al., 2003). In adults, the left STG was involved in phonological processing, whereas the semantic processing recruited the left MTG (Binder, 2016; Devlin et al., 2003; McDermott et al., 2003; Poldrack et al., 1999; Price, 2012). Weiss et al. (2018) directly compared the early specialization of brain regions for phonological and semantic processing of spoken language during early childhood (5–6 years old) by using auditory rhyme and semantic judgment tasks, respectively. By comparing the differences of brain activation between the two tasks using subtraction-based univariate analysis, they found specialization of the left STG and the left supramarginal gyrus for phonological processing, and the left MTG for lexical processing. Their findings suggested that the temporal regions were already specialized for spoken language by 5 years of age. However, our univariate results showed no FDR-corrected activation in temporal regions for phonological processing. Moreover, multivariate analysis revealed an overlap in left MTG/STG and left ITG (including FG) for both phonological and semantic processing. The discrepancy might be due to different task stimuli. Weiss et al. (2018) had used auditory presented words, whereas we used visually presented words.
The left lateralization for reading has been reported to be related to children's reading ability in early childhood (Yamada et al., 2011). Yamada et al. (2011) used a one-back letter reading task on 5-year-old children who received reading instructions in kindergarten and found that typically developing children with on-track pre-literacy skills recruited the left-lateralized temporal regions in the brain. In contrast, children at risk for reading difficulty showed more bilateral activation. Thus, the left hemispheric lateralization observed in our study could be a result of good pre-reading skills of the children recruited in our study.
Temporo-occipital lobe
The bilateral FG has been shown to be involved in processing visually presented words that require orthographic representations in older children (9–12 years old) (Booth et al., 2004). A letterbox, also known as the visual word form area (VWFA) in the left FG, has been suggested to decode letter strings to words (McCandliss et al., 2003). Turkeltaub et al. (2003) showed a decrease in activation in the right VWFA (anatomically homologous to the left VWFA) with an increase in age (6–22 years). They concluded that learning to read led to decreased activations in the right temporo-occipital regions accompanied by increased activations in the left IFG/MTG. Previous research suggested that the left VWFA is already specialized in the left occipito-temporal cortex by the age of 7 years (Gaillard et al., 2003; Parviainen et al., 2006). Our results showed the involvement of the left VWFA in both visual rhyming and semantic tasks and provided evidence that VWFA has already specialized in the left hemisphere by early childhood (5–7 years of age). Some authors postulate that the VWFA is dedicated solely to the lexico-semantic route (Levy et al., 2009), whereas others propose that VWFA is common for both grapho-phonological and lexico-semantic routes, and information is then passed on to the most appropriate route for reading a word (Goswami, 2008; Jobard et al., 2003). A recent study showed the involvement of the left VWFA during a PA task in 5- to 6-year-old children (Wang et al., 2018). Our findings support that the left VWFA is recruited for both pre-reading routes during early childhood.
Parietal lobe
We identified bilateral parietal regions (supramarginal, angular gyrus, and precuneus) for semantic processing, but not for phonological processing. The involvement of the bilateral parietal areas is related to the retrieval of semantic information in adults (Binder and Desai, 2011). For children of age 9 years and older, the left IPL and angular gyrus have been reported to be specialized for semantic categorization tasks (Booth et al., 2007; Landi et al., 2010). Our study provided evidence of bilateral parietal involvement (including the supramarginal gyrus, angular gyrus, and precuneus) for semantic processing and not for phonological processing, suggesting that the specialization of bilateral parietal regions for semantic processing presents even in young children (5–7 years of age). In contrast, Weiss et al. (2018) did not observe parietal specialization for semantic categorization tasks using auditory stimuli for young children (5–6 years of age). They argued that parietal regions might be specialized for semantic processing only later in development. However, our results are in line with another study that identified the bilateral inferior parietal cortex during implicit processing of visually presented words (Turkeltaub et al., 2003). Thus, we propose that the visual stimuli implemented in our study required high imageability for semantic decision making, and bilateral parietal areas were thus recruited for visual semantic categorization tasks in young children.
Implications on theoretical models of reading
The DRC model is a particularly good example of the weak phonological perspective that a direct lexical route takes precedence and is supported by a slow, secondary, and nonessential indirect phonological coding route (Coltheart et al., 2001). The neural pathways of the DRC model have been suggested to involve the dorsal and ventral pathways (Turkeltaub et al., 2003). This study provided the neural bases of pre-reading skills and supported both the models. Our results indicated the involvement of the dorsal pathway in phonological processing, as evidenced by the recruitment of the left opIFG, thus providing support for the DRC model. Our study also provided evidence for the PDP model illustrated by the common activation of fronto-temporal regions related to phonology and semantics in young children.
Limitations
In total, 60 children completed the first behavioral testing session, but only 20 out of 60 children were invited back to the second fMRI session (33%) based on their pre-reading skills, which might bias our sample. This is one of the limitations of this study. We did not observe any significant brain–behavior correlation, which can be attributed to the fact that all 19 children were good pre-readers. The behavioral measures of their pre-reading skills had ceiling effects, which makes brain-behavior correlation hard to be significant and limits the generalization of our results. As we were interested in the neural bases of rhyming and semantics, children were expected to complete tasks with above 60% accuracy. We could have tried to use lower-level cognitive tasks such as first-sound matching (Raschle et al., 2014; Yu et al., 2018) or letter identification (Yamada et al., 2011), but these tasks would have been too simple for those who were 7 years old in our study. We had also collected task-free resting-state fMRI data on our sample, but resting-state fMRI data could not provide task-specific information related to rhyming and semantics.
Conclusions and Future Directions
This study used a multivariate approach to understand the neural bases of phonological and semantic processing in early childhood (5–7 years of age), which has not been previously reported. Our MVPA results suggested that left lateralization for the indirect grapho-phonological route has already been shaped in young children with good reading ability. Moreover, the lexico-semantic route also relies on the left hemisphere regions, including the left IFG/STG/MTG/FG, and the additional recruitment of the bilateral parietal regions. Our MVPA results found that the left opIFG specialization for the phonological processing aligned with previous research in adults (Jobard et al., 2003) and school-age children (Bach et al., 2010). For future research, age-appropriate cognitive paradigms are required to identify pre-reading processes in even younger children (3–5 years of age). Moreover, within-subject design will help to determine the effects of modality (visual vs. auditory) in the neural specialization of phonological and semantic processes in early childhood.
Footnotes
Authors' Contribution
A.M. created the task paradigms by using E-prime 2, helped with data collection, analyzed the data, as well as drafted and revised the article with support from Y.W. Y.W. supervised the entire project; contributed to the conception, design, and data collection of this study; and revised the article for important intellectual content. D.S. provided consultation on MVPA analysis.
Acknowledgments
The authors thank the families for their participation. The authors are also grateful for the assistance from undergraduate research assistants: Cristal Franco-Granados, Makayla Gill, Emily Ann Grybas, Meredith Konkol, Linneaa Nguyen, Grace Oh, Michelle Rohman, Fatima Sibaii, and Thy Thy Trat Thai for helping with recruitment and data collection.
Disclaimer
The content of this article is solely the responsibility of the authors and does not necessarily represent the official views of the NIH.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by funds from the Barkley Trust, Nebraska Tobacco Settlement Biomedical Research Development, College of Education and Human Sciences, and the Office of Research and Economic Development at University of Nebraska
Supplementary Material
Supplementary Figure S1
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.