
Abstract
Among various hippocampal rhythms, including sharp-wave ripples, gamma, and theta, theta rhythm is crucial for cognitive processing, particularly learning and memory. Theta oscillations are observable in both humans and rodents during spatial navigations. However, the hippocampus (Hip) is well known as the generator of current rhythm. Other brain areas, such as prefrontal cortex (PFC), can be affected by theta rhythm, too. The PFC is a core structure for the execution of diverse higher cortical functions defined as cognition. This region is connected to the hippocampus through the hippocampal/prefrontal pathway; hereby, theta oscillations convey hippocampal inputs to the PFC and simultaneously synchronize the activity of these two regions during memory, learning and other cognitive tasks. Importantly, thalamic nucleus reunions (nRE) and basolateral amygdala are salient relay structures modulating the synchronization, firing rate, and phase-locking of the hippocampal/prefrontal oscillations. Herein, we summarized experimental studies, chiefly animal researches in which the theta rhythm of the Hip-PFC axis was investigated using either electrophysiological assessments in rodent or integrated diffusion-weighted imaging and electroencephalography in human cases under memory-based tasks. Moreover, we briefly reviewed alterations of theta rhythm in some CNS diseases with the main feature of cognitive disturbance. Interestingly, animal studies implied the interruption of theta synchronization in psychiatric disorders such as schizophrenia and depression. To disclose the precise role of theta rhythm fluctuations through the Hip-PFC axis in cognitive performances, further studies are needed.
Introduction
Cognitive processes are a product of the simultaneous activity of highly inter-related neural networks in different parts of the brain that execute high-level cognitive functions together. The hippocampus, an integral portion of cognition, contributes to the control of emotion as a part of the Papez circuit and is also primarily involved in memory formation and spatial exploration (Bubb et al., 2017; Choi et al., 2019).
Based on functional magnetic resonance imaging (fMRI) findings, through specific cognitive tasks, different areas of the brain are activated, such as the dorsolateral prefrontal cortex (DLPFC), anterior cingulate cortex, posterior parietal cortex, and medial PFC (mPFC) (Buff et al., 2016; Nakao et al., 2010). These areas form complex cognitive networks that manage and control various domains of the cognitive function, including working memory, attention, planning, inhibition, and other executive functions (Niendam et al., 2012). The cognitive control network concept has provided a new approach in the circuit level to address how the cognitive process is performed in both healthful conditions and problematic psychiatric disorders (Williams, 2016).
Among the abundant circuits of cognitive networks, the PFC and its close bidirectional connections to the hippocampus, hippocampal/prefrontal pathway, seem critical. Direct communication between the hippocampus and PFC, along with other circuits, is in charge of various cognitive functions such as decision-making, memory, and fear (Sigurdsson and Duvarci, 2016). This connection is mainly formed between the cornu ammonis1 (CA1) and mPFC; other circuits consist of different cerebral regions such as the basolateral amygdala (BLA), nucleus accumbens (NAc), nucleus reuniens (nRE), and the ventral tegmental area (VTA) (Godsil et al., 2013). Impairment of the hippocampal/prefrontal pathway triggers a variety of cognitive and psychological disorders (Bast et al., 2017).
Besides the vital role of the hippocampus in cognition, its role in the generation of brain rhythms such as gamma (Colgin and Moser, 2010), sharp-wave ripples (Maier et al., 2003), and theta has been proven (Buzsáki, 2002). It is believed that these rhythms perform a fundamental duty in the coordination of the hippocampus with related structures (Colgin, 2016; Klausberger et al., 2003; Tort et al., 2008). Also, rhythmic activities may contribute to the organization of higher cortical functions, such as speech (Murphy and Benítez-Burraco, 2017) and social communication (Tendler and Wagner, 2015), which are affected in mental disorders, such as autism and schizophrenia (Jochaut et al., 2015; Yu et al., 2018).
Characteristics of theta rhythm, as the well-established rhythm, have been extensively studied during active behavior (Korotkova et al., 2018) and multiple cognitive processes such as memory (Alekseichuk et al., 2017; Berens and Horner, 2017). In the context of excitatory and inhibitory neuronal connections from different hippocampal subregions, theta rhythm (4–12 Hz) appears (Colgin, 2016) in the local field potential (LFP) recordings (Chatzikalymniou and Skinner, 2018). Based on the evidence, the hippocampus integrates acquired information through neocortical circuits using oscillations such as theta (Tingley and Buzsáki, 2018; Zheng et al., 2018). Along with theta activity, other rhythms, especially gamma, may oscillate with theta frequency through hippocampal/prefrontal synchrony during the cognitive performance, in a coupled manner (Kupferschmidt and Gordon, 2018).
As mentioned earlier, among neocortical areas, the PFC is closely related to the hippocampus (Carr and Sesack, 1996) so that it receives remarkable excitatory glutamatergic inputs from this structure (Parent et al., 2009). Given the importance of hippocampal/prefrontal coordination, in this review, we focus on synchrony between these two regions in terms of theta rhythm during cognitive functions.
Hippocampal/Prefrontal Pathway
In this section, we outline the anatomical connections that join the hippocampus to the PFC and then highlight the mechanisms of synaptic plasticity in this pathway.
Anatomical connections
Several pathways are responsible for conveying data from the hippocampal formation to diverse parts of the PFC. According to primates and rodent studies, these projections are mainly terminate to the medial and orbital PFC (mPFC, OPFC), including 11, 13, 14, 24a, 32, and 25 areas (Aggleton et al., 2015). It should be noted that connecting pathways exchange information directly, unilaterally, or bilaterally between the hippocampus and PFC (Öngür and Price, 2000). mPFC, as a more connected part of PFC to the hippocampus (Cenquizca and Swanson, 2007; Jay and Witter, 1991), is divided into four key regions from the ventral to the dorsal part: the medial agranular, anterior cingulate, prelimbic (PL), and the infralimbic (IL) (Heidbreder and Groenewegen, 2003). Ventral parts (PL, IL) are associated with emotional, cognitive, and memory processing (Nelson and Guyer, 2011; Öngür and Price, 2000; Sallet et al., 2013). The mPFC circuits consist of mPFC along with the hippocampus, NAc, insular cortex, mediodorsal, and reuniens nuclei of the thalamus, BLA, and VTA of the midbrain (Vertes, 2006) (Fig. 1).
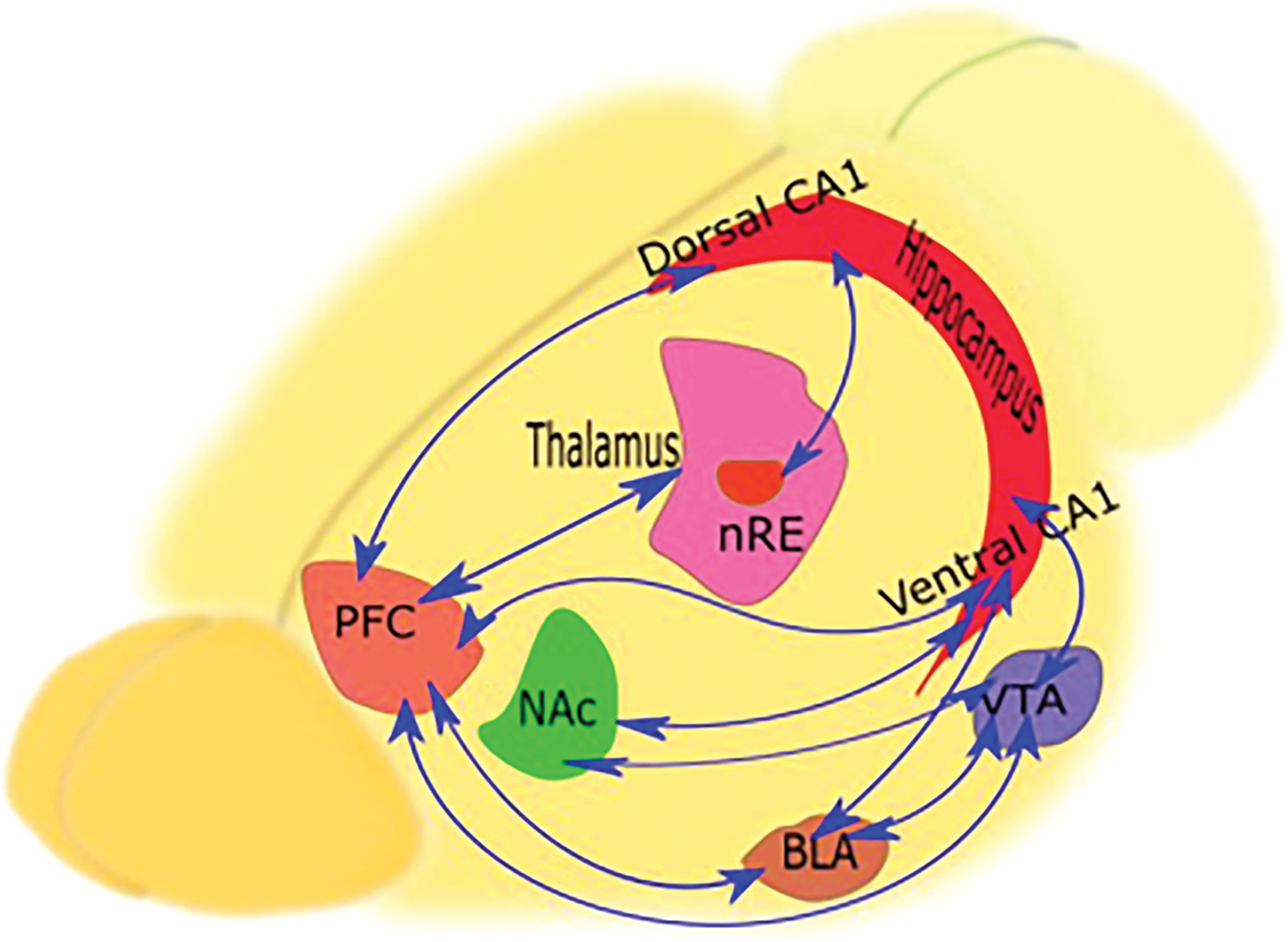
Anatomical connections between the hippocampus and PFC. As illustrated schematically in the figure, there are direct and indirect routes connecting the hippocampus to the PFC. In addition to the direct monosynaptic pathway, the polysynaptic route traverses through various structures, especially NAc and VTA, creating the Hip-NAc-VTA-PFC circuit. In the recent circuit, VTA and hippocampus bidirectionally influence each other. In the Hip-BLA-PFC circuit, the PFC can affect the hippocampus through the amygdala. In this circuit, the amygdala is associated bilaterally with the PFC and hippocampus. Instead, in the PFC-nRE-Hip circuit, the PFC can affect the function of the hippocampus through the nRE of the thalamus. This nucleus is primarily associated with the dorsal hippocampus. BLA, basolateral amygdala; Hip, hippocampus; NAc, nucleus accumbens; nRE, nucleus reuniens; PFC, prefrontal cortex; VTA, ventral tegmental area. Color images are available online.
In the rat brain, fibers originating from the CA1 and subiculum areas of the hippocampus project mainly to the infralimbic, medial orbital, PL, and anterior cingulate regions (Rosene and Van Hoesen, 1977; Swanson, 1981). These fibers pass through the fornix to reach these areas and form a direct relationship between the hippocampus and the PFC (direct-Hip-PFC pathway) (Cenquizca and Swanson, 2007; Godsil et al., 2013; Hoover and Vertes, 2007; Jay and Witter, 1991). In addition to the direct Hip-PFC pathway, there are multisynaptic pathways between these two structures. In the mutual Hip-NAc-VTA-PFC circuit, the hippocampus, receiving direct input from the VTA (Nazari-Serenjeh et al., 2011), stimulates VTA neurons through direct glutamatergic efferent or indirect NAc projections (Tan, 2008). At the end of their journey, a fraction of the ventral hippocampal neurons project to both BLA and mPFC (Sierra-Mercado et al., 2011), producing the Hip-BLA-PFC circuit. This circuit is responsible for coordination between the amygdala and PFC (Serkov et al., 2016). Moreover, the PFC also can manipulate the hippocampal activity via the nRE, a thalamic relay nucleus, in the context of the PFC-nRE-Hip circuit (Bokor et al., 2002). These complex anatomical connections are accompanied by synaptic plasticity that affects their communication, as described below.
Synaptic transmission
According to the intracellular recordings, hippocampal pyramidal terminals form asymmetric synapses with prefrontal pyramidal neurons, which are divided into three classes: regular spiking, inactivating bursting, and noninactivating bursting (Carr and Sesack, 1996; Dégenètais et al., 2002). Extracellular recordings and neural pathways' tracing by d-[3H]aspartate revealed that the hippocampal/prefrontal pathway is an AMPA-mediated glutamatergic route (Jay et al., 1992).
There are two kinds of postsynaptic potentials from prefrontal neurons, including inhibitory and excitatory postsynaptic potentials. As a result of paired-pulse stimulation in the hippocampus, short-term plasticity emerges in prefrontal neurons, which can play a role in the temporal processing of the information. In contrast, a tetanic stimulation produces a long-term potentiation in prefrontal neurons, followed by an inhibitory postsynaptic potential. These two short- and long-term phenomena, using the hippocampal/prefrontal pathway, contribute to the formation and consolidation of memory, as well as high-level cognitive processes (Dégenètais et al., 2003).
Stimulation of the ventral hippocampus of the awake and freely moving rats by two different high-frequency stimuli (HFS) led to a long-term potential (LTP) in the mPFC, lasting more than a week. It is worth mentioning that rats showed a more significant hippocampal/prefrontal response lasting 9 days after being exposed to enriched environments throughout the night (Taylor et al., 2016). The occurrence of the LTP in PFC depends on NMDA receptors and further activation of cAMP-dependent kinase (PKA), associated with an increase in dopamine levels.
Given that mesocortical dopamine inputs are regarded as influencing factors in the hippocampal/prefrontal pathway, elevation of dopamine and subsequent dopamine receptor stimulation induce a long-term enhancement in the hippocampal/prefrontal LTP. Therefore, blocking of D1, not D2, receptors impairs LTP (Jay et al., 2004) but facilitating of D2 receptors terminates to long-term depression (Banks et al., 2015). Synaptic plasticity in this pathway leads to oscillatory changes, mostly in the theta range; in the next section, we uncover hippocampal/prefrontal theta oscillations.
Hippocampal/Prefrontal Oscillations
Dynamic coupling between different regions is a necessary part of rhythm functions and observable during local field potential and single-unit recordings (Fell et al., 2001; Tabuchi et al., 2000; Varela et al., 2001; Womelsdorf et al., 2007). The outcome of this type of coordination is that diverse areas can accompany or block others' activities (Womelsdorf et al., 2007). Since the hippocampus and PFC are not excluded from this rule, synchronization of theta rhythm between these two areas can affect various cognitive states such as anxiety, memory, sleep, seizure, and schizophrenia (Backus et al., 2016; Kleen et al., 2011; Sigurdsson et al., 2010; Tang et al., 2017).
The hippocampus produces three main types of rhythms, including sharp-wave ripples, gamma, and theta. Theta rhythms (4–12 Hz) are detectable during behaviors such as free movement, rapid eye movement sleep, and spatial memory tasks. Gamma (25–140 Hz) rhythm usually occurs along with theta rhythm. The slow and irregular sharp waves (1–10 Hz) are a series of oscillations that accompany fast ripple oscillations (150–300 Hz). Combining these recent two waves brings about sharp-wave ripple, which predominates during deep sleep (Colgin, 2016). These rhythms can all coordinate and regulate the relationship of the hippocampus with other brain structures.
Hippocampal/prefrontal theta oscillations throughout cognitive functions
Hippocampal/prefrontal theta rhythm may be affected by various cognitive statuses such as anxiety, memory formation, and psychological diseases, which is discussed in more detail below.
Working memory
Theta oscillations are observable in both humans and rodents during spatial navigations, but in humans, they occur at low frequencies. Hippocampal rhythms of rats are centered nearby ∼8 Hz, while that of humans is about ∼3 Hz (Watrous et al., 2013) (Fig. 2).

Comparison of low-frequency oscillations in rats and humans through spatial navigation.
Communication between the hippocampus and PFC is necessary for both working and long-term memory (Axmacher et al., 2008; Fuster, 2009). There is no possibility for a direct electrophysiological recording of the hippocampus and PFC in humans, but animal studies have revealed that numerous prefrontal neural networks are involved in spatial working memory (SWM) synchronizing with the hippocampus (Jin and Maren, 2015; Lara and Wallis, 2015). In human subjects, using integrated diffusion-weighted imaging and electroencephalography, close anatomical connections have been found between the hippocampus and right ventrolateral PFC (area 46) during working memory tasks, such that low-frequency oscillations appear while the task is carried out (Cohen, 2011). So, neural fluctuations may provide integrity between the hippocampus and PFC during working memory tasks.
Since the mPFC plays a crucial role in the processing of SWM, theta oscillations in the mPFC are synchronized with the hippocampal theta oscillations throughout SWM tasks. The in vivo LFP recordings of the ventral hippocampus (vHPC), dorsal hippocampus (dHPC), and mPFC, while a mouse is performing the T-maze, exhibit strong theta power in the mPFC that changes during various stages of the T-maze, but theta power (amplitude as a voltage) and peak frequency are more prominent in the vHPC and dHPC compared with the mPFC. Following infusion of muscimol into the vHPC, gamma frequency power decreases more notably compared with the theta frequency power (O'Neill et al., 2013). Every success in the T-maze is directly related to the coherence of theta rhythm in the mPFC and dHPC. During the task, theta oscillations in the dHPC precede the mPFC theta about 30 ms for every right answer, but not for the wrong trials, and mPFC low gamma precedes the dHPC in correct trials contrary to the incorrect trials. Moreover, theta/gamma phase/amplitude coupling during choice-point traversals differs as a function of task performance. Considering the nRE as an important relay structure between the mPFC and hippocampus, inactivation of this nucleus diminishes the single unit firing rate of the mPFC (Hallock et al., 2016).
Studies typically prove that the hippocampal theta phase is correlated with the mPFC neural spiking. Neurons in an assumed network may respond by a spike or frequency at times when they receive a neuronal input. This response in the context of fixed time delays to a periodic input (such as theta) is mentioned as phase-locking, which is fundamental for the synchronization within different regions of the brain (Fig. 3) (Stoop et al., 2004).
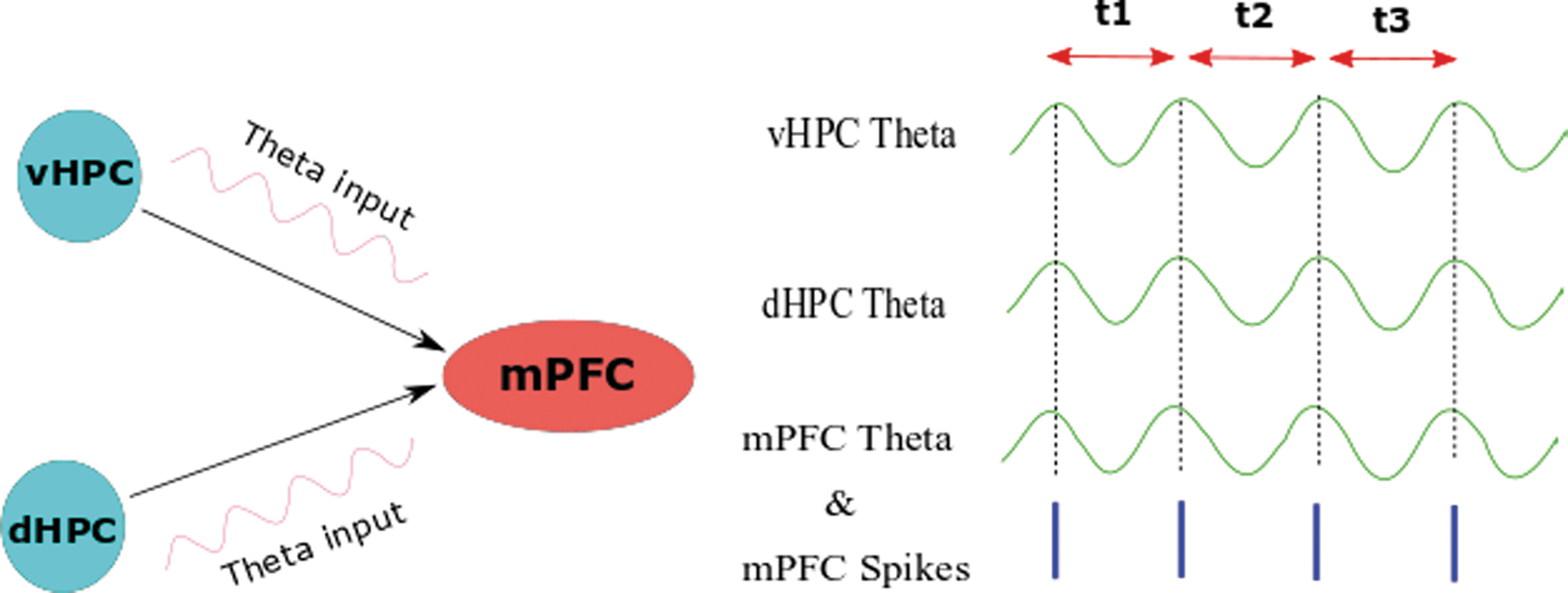
Phase-locking in the hippocampal/prefrontal pathway. Responses of mPFC neurons in a pattern of fixed time delays (t1 = t2 = t3) to a periodic hippocampal input (theta) lead to phase-locking between two regions. mPFC, medial PFC. Color images are available online.
Interestingly, there is phase-locking between the mPFC and CA1 so that mPFC spikes occur within constant time windows imposed by continuous CA1 theta rhythm, and this phase-locking between the prefrontal cells and CA1 is predicted by significant correlations of them at delays about 150 ms (Siapas et al., 2005). Overall, theta rhythm is a general coordinator mechanism; hence, while the behavior is ongoing and requires interactions between structures, it synchronizes different neural activities and facilitates the encoding of information (Jones and Wilson, 2005).
Inferential reasoning is a phenomenon in which different memories integrate, which is necessary to perform complex cognitive activities in humans (Kumaran et al., 2009; Wood et al., 2000; Zeithamova et al., 2012). Based on magnetoencephalography recordings, while participants are doing inference tasks, hippocampal theta power increases and communicates across the hippocampus and mPFC, indicating that memory is being integrated. Thus, hippocampal theta oscillations, by spreading into mPFC, correspond to dynamic exchanging between the encoding process and retrieval of information. According to the time-frequency window of the peak theta power, theta coherence of the hippocampal region to the mPFC elevates during the task (Backus et al., 2016).
The cross-frequency coupling of brain rhythms is observable in various parts of the brain during diverse cognitive activities such as working memory. For example, this kind of coupling can be dramatically manifested between the mPFC and hippocampus as theta/gamma coupling in a transgenic mouse model of cognitive problems. Any success in the SWM test leads to the enhancement of cross-frequency coupling. The interference in the encoding stage by optogenetic tools or increasing the task difficulty potentiates the theta/gamma coupling for each correct trial. Eventually, as neuron synchronization in the mPFC elevates, the coupling of high theta and low gamma in the hippocampal/prefrontal pathway increases too. This coupling is a mechanism that acts as a compensator, and as the SWM difficulty intensifies, its settings change. Besides, there is a phase-power coherency between the gamma power of mPFC and theta phase in the mPFC, so that theta oscillations of mPFC and vHPC can affect gamma power of the mPFC (Tamura et al., 2017).
According to recent studies, the hippocampus can be indirectly linked to the mPFC via nRE, which is a midline thalamic nucleus. Hence, one of the crucial circuits between the hippocampus and the PFC is the PFC-nRE-Hip circuit, which can play a role in emotional, episodic, and spatial memory (Jin and Maren, 2015). Whenever rats are trained to perform delayed nonmatch-to-sample spatial alternation in T-maze, the injection of procaine to the nRE through a cerebral cannula results in long-term delays. However, following the injection of muscimol, it is difficult to perform all delays, and animals suffer from spatial perseveration. Thereby, during the correction steps, they enter the wrong arm repeatedly (Viena et al., 2018).
Generally, PFC-nRE-Hip circuit is fundamental in SWM tasks, in which (1) the dHPC affects the PFC through nRE, such that whenever the animal performs the SWM task correctly, a group of neurons in the PFC under the influence of the hippocampus begin to produce theta rhythm; (2) in an SWM situation, hippocampal theta rhythm in the hippocampus is coupled with theta as well as the gamma oscillations of the PFC; (3) if the nRE is deactivated by muscimol, the circuit will be disrupted, and the animal will not be able to perform the SWM task. In addition, following muscimol infusions into nRE, mPFC spike entrainment to the hippocampal theta oscillation diminishes during the task (Hallock et al., 2016).
These findings exhibit evidence for the theta rhythm importance in coordinating the mPFC with vHPC and dHPC during SWM, mental flexibility, and decision-making. The thalamic nRE plays a vital role in the hippocampal frontal pathway through the PFC-nRE-Hip circuit (Fig. 4).
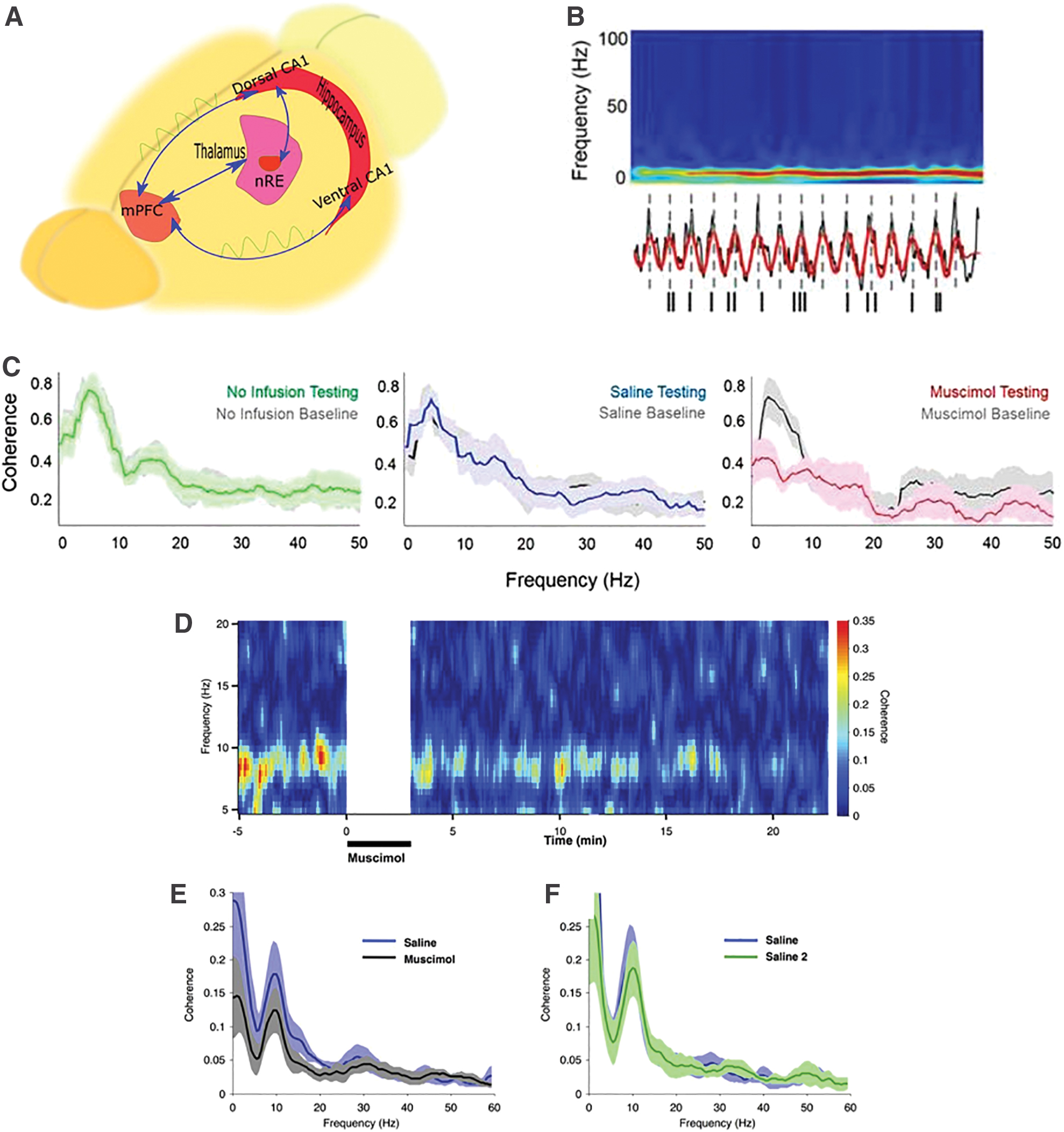
Role of hippocampal/prefrontal synchronization in the integration of working memory.
Fear learning and anxiety behaviors
Insight into the anxiety circuits and pathways has shed light on the mechanism of this complex behavior. mPFC, vHPC, and BLA are involved in anxiety-like behaviors (Felix-Ortiz et al., 2013; Kjelstrup et al., 2002; Maren and Holt, 2004; Sierra-Mercado et al., 2011). In conditioned fear learning tasks, the pharmacological blockade of BLA before the task decreases the neuron firing rate that projects from the mPFC to the BLA and also decreases the tonic responses of single-neuron spiking in the mPFC (Sotres-Bayon et al., 2012). In addition, anxiolytic drugs weaken the power of evoked theta frequency in the posterior hippocampus, but anxiogenic drugs conversely potentiate it (Yeung et al., 2012, 2013a,b).
In contrast to the dHPC, considering vast connections with mPFC as well as close and bidirectional communication with the amygdala, the vHPC has a more critical role in the discovery of theta rhythm mechanisms in anxiety-like behavior (Fanselow and Dong, 2010). According to the genetic model of anxiety-like behavior (serotonin 1A receptor knockout mice), theta power increases in the mPFC as a predictor of aversive avoidance, indicating that the hippocampus is in synchrony with downstream targets such as mPFC during anxiety. Moreover, based on single unit and LFP recordings along with optogenetic manipulations, unilateral inhibition of the vHPC projections to the mPFC resulted in the loss of anxiety and lowered the firing rate of mPFC and theta power of the pathway, but did not alter power correlations of the delta, beta, and slow gamma frequencies (Padilla-Coreano et al., 2016). Blockade of gap junctions in the vHPC and mPFC depresses anxiety-like behaviors, plus the change of theta rhythm fluctuations (Schoenfeld et al., 2014).
Neural signals conducting the memory consolidation and extinction originate from various brain regions, including the lateral amygdala (LA), hippocampal CA1 region, and infralimbic area of the mPFC (Ji and Maren, 2007; Maren and Quirk, 2004; Sehlmeyer et al., 2009). Based on local field potential and multiunit recordings, the theta rhythm can connect areas mentioned above in terms of time/frequency coupling. It has been shown that during fear retrieval, theta rhythm coupling in the CA1-LA-mPFC circuit increases, but declines with the extinction of learning (Lesting et al., 2011). The conditioned avoidance reflex is associated with high frequencies of theta rhythm (8–11 Hz) and evanescence of delta following the entrance of the animal into the safe half of the chamber (Serkov et al., 2016). The mPFC cortex is essential for detecting the safety and fearful condition, so placing the mouse in the position of innate and learned anxiety enhances the theta synchrony between the prefrontal and amygdala. Also, once the mouse feels safe, the amygdala mimics the mPFC theta frequency. Based on phase/amplitude coupling results, slow (40–70 Hz) and fast (70–120 Hz) gamma frequencies are strongly coupled with low theta (4–8 Hz) frequency that is provoked by the fear (Likhtik et al., 2014). The mPFC–vHPC–BLA circuit has been suggested to have a role in risk assessment behaviors. Thus, simultaneous local field potential recordings from vHPC, BLA, and mPFC indicate a harmonic pattern of theta power variations that predicts the exploration of open-arm or returning to the closed safe arm in the elevated plus maze (EPM). Likewise, similar patterns of theta variations in the chronic unpredictable stress are observed, thereby with approaching to the open arm theta coherence in the vHPC-BLA, and BLA-mPFC circuits augment over the time from baseline (Jacinto et al., 2016). According to the EPM results, single-unit recordings from mPFC neurons in addition to local field potentials of the hippocampus have shown that the exploratory action of animals in the maze increases the activity of mPFC neurons coupling with the hippocampal theta rhythm (Adhikari et al., 2011). Although theta synchrony and power in the vHPC-BLA and BLA-mPFC circuits increase during innate anxiety, it is noteworthy that fast gamma power does not follow theta and decreases in both circuits (Lesting et al., 2011; Likhtik et al., 2014; Stujenske et al., 2014).
Generally, the mPFC controls fear-related emotions by integrating diverse inputs projecting from different cerebral areas, particularly the vHPC and BLA. In this scene, mPFC acts as a gateway to control fear and anxiety through the coupling of theta oscillations with the firing activity of neurons (Fig. 5).
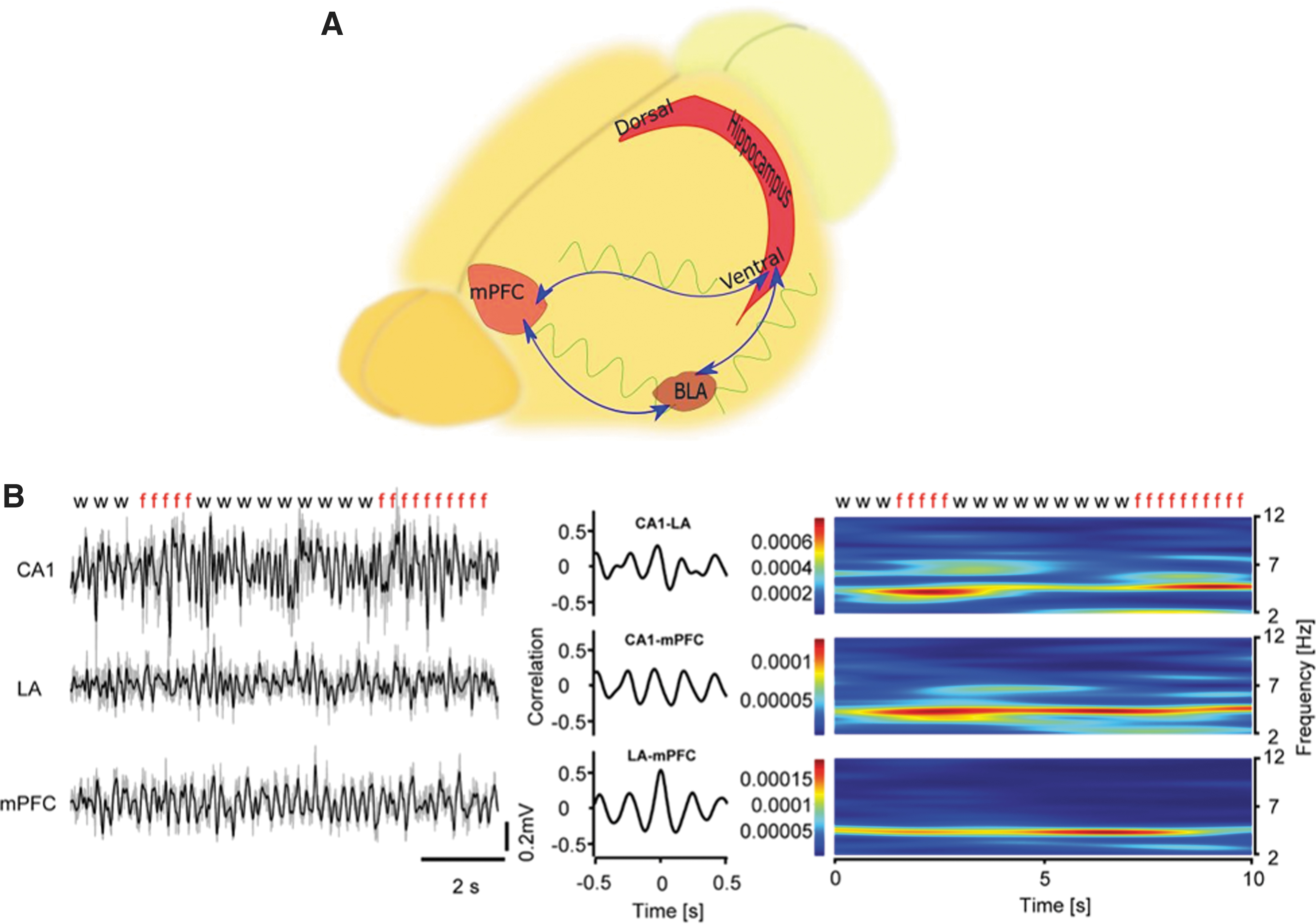
Role of the hippocampal/prefrontal synchronization in the fear learning behavior.
Associative learning
According to various studies, both the hippocampus and PFC are involved in associative learning, and raised theta phase synchrony in the hippocampal/prefrontal pathway is observed throughout the retrieval of object/context associations. On the authority of the theta-amplitude cross-correlation in the hippocampal/prefrontal pathway, the correlation invigorates following an accurate performance under a context learning task (Place et al., 2016).
As previously mentioned, the hippocampal/prefrontal pathway is important in learning and spatial memory, but recent studies have shown that this pathway is also critical for nonspatial learning. Caudal mPFC plays a role in the acquisition of hippocampal-related eye-blink conditioning trace, associating two separate stimuli: a trace paradigm with an air puff as unconditioned stimulus (US) and tone as conditioned stimulus (CS), whereas the rostral mPFC is more critical in the extinction of the acquired conditioned response (Weible et al., 2000). Classical eye-blink conditioning is accompanied by some changes in the synaptic strength of reuniens-mPFC and the reuniens-CA1 pathways. With a single stimulus pulse applied to the nRE at intervals between conditional and unconditional stimulation, no significant synaptic variations were observed in the reuniens-CA1 and reuniens-mPFC pathways during the eye-blink acquisition, considered a conditional response (CR). However, HFS diminished the number of CRs without changing the LTP. As the percentage of success in the discrimination of objects depletes, the number of facilitated paired-pulses declines (Eleore et al., 2011). It can be concluded that the nRE can promote associative learning through the formation of a circuit between the hippocampus and PFC.
In the cognitive disorder models, whenever animals underwent the eye-blink conditioning trace, theta and gamma phase/phase or amplitude/amplitude coupling in the hippocampal/prefrontal pathway rose during the two separate stimuli interval. Despite typical memory acquisition, this coupling was weakened, and theta/gamma cross-frequency coupling (phase/amplitude) in the mPFC increased (Tanninen et al., 2017).
In monkeys learning pair association of objects, the hippocampus reflects the correct or incorrect response in each trial; the firing of neurons in the PFC changes along with the animal's learning and task performance. In the presence of errors, synchronization of the hippocampal/prefrontal pathway is intensified through the theta range, in which the hippocampus is initially affected by the PFC. Conversely, following the occurrence of correct trials, the hippocampus affects the PFC, followed by strengthened alpha/beta synchrony. As learning is ongoing, the frequency of hippocampal/prefrontal synchrony shifts from low frequency to a high frequency over time (Brincat and Miller, 2015).
Evidence from pre-clinical literature shows that rapid associative learning initially occurs in the PFC, but the hippocampus plays a crucial regulatory role by discriminating against the mistake and correct trial; thus it alters the coordination of the hippocampal/prefrontal rhythms. It must be noted that the results of clinical studies verify the role of hippocampal/prefrontal synchrony in the processes of associative learning too. As claimed by the weighted phase lag index (a functional connectivity measure), throughout the encoding of contextually expected and unexpected visual stimuli, theta phase synchronization (during time-frequency bins) between the hippocampus and PFC was higher for unexpected items (Gruber et al., 2017).
Theta Oscillation Alteration in the Hippocampal/Prefrontal Pathway Following Neuropsychological Disorders
In addition to the pivotal role of hippocampal/prefrontal theta oscillations in normal conditions such as learning and memory formation, changes in these oscillations have been observed in several pathologic conditions. In the following, we review frequently reported diseases in which theta rhythms have undergone alterations in the hippocampal/prefrontal pathway in accompanying with cognitive deficits.
Depression
Chronic depression can lead to volumetric and structural variations in the hippocampal formation, and considering the prevalence of memory disorders in people with depression, structural and physiological modifications in these areas are expected (Hastings et al., 2004; Sauer et al., 2015). According to fMRI studies, following depression, connections between the hippocampus and the PFC interrupt, and it is expected that disturbances of the hippocampal/prefrontal pathway affect its oscillations, and thereby disarrange the processing and transmission of cognitive information (Liu et al., 2017; Sigurdsson and Duvarci, 2016). Following LTP induction, depressed rats showed reduced theta coupling in the hippocampal/prefrontal pathway compared with healthy ones. Based on this evidence, there is a positive relationship between either region coupling with their plasticity. Also, after HFS, theta/slow gamma, phase/amplitude coupling increases in the CA1 region in a long-term manner (Zheng and Zhang, 2015). Taking advantage of the knowledge discussed above, a new method of treatment for depression calling theta-burst stimulation has been established recently. This method is based on a repetitive form of transcranial magnetic stimulation, which is a noninvasive and safe approach to change brain activity and alter cortical neural function (Chung et al., 2015).
Alzheimer's disease
Alzheimer's disease (AD) is a type of dementia that causes problems with memory, thinking, and behavior (Zangbar et al., 2016). The primary pathologic findings of this disease are the formation of neurofibrillary tangles and senile plaques, induced by the accumulation of amyloid-β (Aβ) peptide and tau, leading to loss of synapses, as well as neurons (Kumar and Singh, 2015). Eventual neurodegeneration in areas affected by these pathological features will cause modifications in brain rhythms and change in the memory and cognitive processing.
In the early stages of AD, based on magnetoencephalography (MEG) assessments, there is strongly attenuated intensity of alpha-band oscillation bursts over temporoparietal areas in the long-life and waiting-times with significantly low autocorrelations on long time scales. Conversely, theta oscillations in the waiting- and life-times have shown remarkable elevation over the mPFC. Both mPFC theta and temporoparietal alpha oscillations are positively correlated with retrieval and retention of memory and cognitive information. Given that synaptic and structural deficiencies are observed mainly in temporoparietal regions in the early stages of AD, increased theta oscillations in the mPFC can represent a compensatory mechanism (Montez et al., 2009). There is a negative correlation between the Mini-Mental State Examination and hippocampal theta power, but a positive correlation exists with respect to the cortical peak frequency. Theta power could be a good diagnostic discriminator between AD and healthy persons (Engels et al., 2016).
It is worth mentioning that the Aβ aggregation changes the response of the mPFC to the stimulation of the hippocampus, both in the single unit and local field activity levels (Flores-Martínez and Peña-Ortega, 2017). Besides, tau hyperphosphorylation in the early stage of AD correlates with attenuation of neuronal bursts and remodeling of theta oscillations in hippocampal pyramidal neurons and some parvalbumin-positive interneurons (Mondragón-Rodríguez et al., 2018). APP-KO is a knockout mouse model of amyloid precursor protein (APP) that exhibits significant cognitive impairments along with lower theta/gamma coupling through LFP recordings of the hippocampus, but not in the PFC (Zhang et al., 2016). Coherent activity and theta/gamma, phase/amplitude/cross-frequency coupling in the P301L mice, a tau model of AD, rise in the PFC-CA1-CA3 pathway. Early reduction of theta oscillations, as well as coherent activity, is manifested in this pathway, followed by theta/gamma cross-frequency coupling (Ahnaou et al., 2017). Overall, these observations imply variations of the hippocampal/prefrontal theta rhythm in AD and even its importance in early diagnosis of it.
Schizophrenia
Schizophrenia is a neurodevelopmental psychiatric disorder affecting the social life of persons, and the severity of symptoms are linked to structural and synaptic anomalies of the hippocampus and PFC (Qiu et al., 2010). These disturbances influence the functional coupling between the hippocampus and PFC, consequently affecting the synchrony between these two regions (Meyer-Lindenberg et al., 2005). In the course of working memory tasks in rats with maternal immune activation, a neurodevelopmental model of schizophrenia, the theta coherence between the hippocampus and mPFC is reduced (Dickerson et al., 2012). Connections between the hippocampus and PFC can be affected by the polymorphism in the ZNF804A gene, a specific gene linked to schizophrenia. Using the imaging approach, this gene's effects on the network and rhythm performance of the hippocampus have been studied. The dual regression approach was used to examine the coherence between the hippocampus and other areas of the brain; as a result, MEG analyses showed a significant reduction in the hippocampal theta; in contrast, fMRI results indicated high coordination of the superior frontal gyrus with the hippocampus in high-risk homozygotes (Cousijn et al., 2015). In a mouse model of schizophrenia, with a microdeletion in the 22 (22q11.2), the hippocampal/prefrontal synchrony was reduced during working memory tasks (Sigurdsson et al., 2010).
Other disorders
The neural dynamics could be affected in chromosomal disorders such as Down syndrome (DS), that cognitive dysfunction is one of the main features of this disorder (Real et al., 2018). In the trisomic mouse model of DS (Dp1Tyb), theta frequency is lower in the DS animal compared with the wild-type (WT) animal in both mPFC and hippocampus areas during spontaneous alternation of T-maze, but there was no significant difference in terms of theta power. Also, there was a considerable difference between DS and WT animals in the phase/amplitude coupling of theta and gamma rhythms (Chang et al., 2020).
There is an abnormal connection between the hippocampus and mPFC in the temporal lobe epilepsy (TLE) (Kemmotsu et al., 2014). In the rat model of TLE, theta/delta power raise in the hippocampus before the seizure onset, and theta coherence may enhance in the hippocampal/prefrontal pathway in the pre-ictal phase that instantly reduces following seizure attack (Broggini et al., 2016).
Conclusion
Anatomical, electrophysiological, and imaging studies verify the crucial role of the hippocampal/prefrontal pathway in the control of cognitive function. In this review, we detailed the evidence that illustrates the importance of theta rhythm oscillations in the proper function of this pathway in terms of cognitive behaviors. The synchrony of theta rhythm between these areas is evident while performing complex cognitive tasks such as spatial memory, associative learning, and anxiety behaviors.
Various neural circuits are involved in accomplishing these cognitive tasks: (1) PFC-nRE-Hip circuit, which is mainly involved in working memory and associative learning. In this pathway, the hippocampus affects the PFC through the nRE nucleus, followed by the prefrontal theta oscillations. (2) CA1-LA-mPFC circuit, which is vital in fear learning and anxiety behaviors. During fear retrieval, theta rhythm coupling increases in this circuit but declines in the extinction of learning (Table 1).
Overview of Hippocampal/Prefrontal Theta Oscillations Through Cognitive Performances
BLA, basolateral amygdala; dHPC, dorsal hippocampus; EPM, elevated plus maze; mPFC, medial prefrontal cortex; nRE, nucleus reuniens; OF, open field; vHPC, ventral hippocampus.
In the light of hippocampal/prefrontal oscillations' importance in the normal cognitive execution, damage to this pathway will affect the coherence and phase-locking of theta in this route, and along with cognitive disorders such as depression, Alzheimer's, and schizophrenia, oscillatory representation of this pathway such as theta oscillations changes. It seems that understanding the regulatory mechanisms of the current pathway can offer a promising therapeutic approach, based on the recovery of normal oscillatory patterns.
Footnotes
Authors' Contributions
H.S.Z. and P.S. devised the main conceptual idea. H.S.Z. and P.S. wrote the article with support from T.G. S.F. and M.G. helped to prepare the figures. T.G., A.E.K., and M.S.V. helped to edit the article.
Acknowledgment
The authors thank the Neuroscience Research Center (NSRC) for providing the conditions and research space.
Ethical Approval
This study (
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This article was supported by Iran National Science Foundation (INSF) (No. 97017102).