
Abstract
Aim:
The present study aims at investigating the possible correlation between peripheral markers of inflammation and brain networks.
Introduction:
Amyotrophic lateral sclerosis (ALS) is a neurodegenerative disease dominated by progressive motor impairment. Among the complex mechanisms contributing to the pathogenesis of the disease, neuroinflammation, which is associated with altered circulating cytokine levels, is suggested to play a prominent role.
Methods:
Based on magnetoencephalography data, we estimated topological properties of the brain networks in ALS patients and healthy controls. Subsequently, the blood levels of a subset of cytokines were assayed. Finally, we modeled the brain topological features in the function of the cytokine levels.
Results:
Significant differences were found in the levels of the cytokines interleukin (IL)-4, IL-1β, and interferon-gamma (IFN-γ) between patients and controls. In particular, IL-4 and IL-1β levels increased in ALS patients, while the IFN-γ level was higher in healthy controls. We also detected modifications in brain global topological parameters in terms of hyperconnectedness. Despite both blood cytokines and brain topology being altered in ALS patients, such changes do not appear to be in a direct relationship.
Conclusion:
Our results would be in line with the idea that topological changes relate to neurodegenerative processes. However, the absence of correlation between blood cytokines and topological parameters of brain networks does not preclude that inflammatory processes contribute to the alterations of the brain networks.
Impact statement
The progression of amyotrophic lateral sclerosis entails both neurodegenerative and inflammatory processes. Furthermore, disease progression induces global modifications of the brain networks, with advanced stages showing a more compact, hyperconnected network topology. The pathophysiological processes underlying topological changes are unknown. In this article, we hypothesized that the global inflammatory profile would relate to the topological alterations. Our results showed that this is not the case, as modeling the topological properties as a function of the inflammatory state did not yield good predictions. Hence, our results suggest that topological changes might directly relate to neurodegenerative processes instead.
Introduction
Amyotrophic lateral sclerosis (ALS) is a neurodegenerative disease primarily characterized by the progressive death of motor neurons, resulting in the phased paralysis of skeletal muscles and respiratory failure within 2–5 years after disease onset (Rusconi et al., 2017).
Many mechanisms have been proposed to explain motor neuron degeneration. These include glutamate toxicity, oxidative stress, aberrant protein aggregation, defective axonal transport, mitochondrial dysfunction, and altered RNA metabolism (Al-Chalabi and Leigh, 2000; Barber and Shaw, 2010; Smith et al., 2019). Although there is little doubt that these mechanisms are involved at some point in the disease pathogenesis, none alone accounts for the death of motor neurons. It is commonly accepted that the disease is the consequence of a combination of pathogenic processes, which are not only limited to the death of motor neurons but also entail neuroinflammation, with microglial activation, astrogliosis, and infiltration of monocytes, neutrophils, and T lymphocytes into the brain parenchyma (McCauley and Baloh, 2019; Philips and Robberecht, 2011; Prado et al., 2018; Rusconi et al., 2017). Such process involves both the innate and the adaptative immune response (Lyon et al., 2019; Rusconi et al., 2017).
In ALS, the involvement of the two types of immune response is confirmed by autoptic tissues, where the presence of both T cells (Engelhardt et al., 1993; Kawamata et al., 1992) and cells from the innate response (Graves et al., 2004; Henkel et al., 2004) is demonstrated. However, these features are not distinguishable from different neurodegenerative diseases, or from other conditions such as stroke or traumatic injury. Given this lack of specificity, one can suppose that the inflammatory reaction is secondary to neuronal death. In contrast, recent research has proposed an active role of the neuroinflammatory chain in neurodegenerative diseases (Komine and Yamanaka, 2015). For istance, autopsy-based brain studies have shown that neuroinflammation might promote progression of these disorders and therefore plays a primary role in the pathophysiology of the disease (McGeer et al., 1988).
Increasing evidence suggests that neuroinflammation in ALS is associated with systemic inflammation involving the innate immune system (Keizman et al., 2009; Lunetta et al., 2017; Won et al., 2016). The inflammatory response is characterized not only by altered circulating lymphocytes and monocytes (Murdock et al., 2017), but also by the alteration of inflammatory cytokine levels (McCombe and Henderson, 2011). Robelin and Gonzalez De Aguilar (2014) suggested that the blood cytokines may be followed in the periphery as potential biomarkers of neuroinflammation.
Cytokines are the main modulators of cell proliferation and differentiation, and depending on their function in the inflammatory processes, they are divided into interleukins (ILs), tumor necrosis factors (TNFs), and interferons (IFNs).
Interleukines may be proinflammatory or anti-inflammatory and are mainly synthesized by T cells, macrophages, and endothelial cells, promoting the development and differentiation of both T and B cells (Moreno-Martinez et al., 2019). It is known that the levels of several interleukins, such as IL-1β, IL-2, and IL-4, are increased in both the cerebrospinal fluid (CSF) and the blood of ALS patients compared with healthy controls (Blasco et al., 2017; Lu et al., 2016; Mitchell et al., 2009).
Among the major proinflammatory cytokines, the TNF-α is secreted by activated macrophages and it is involved in the production of cytokines, in the activation of phagocytes, and in the expression of adhesion molecules (Turner et al., 2014). Its role in ALS pathogenesis is still unclear because of its pleiotropic actions. In fact, in some cases, the level of TNF-α in the blood and CSF of ALS patients increases compared with healthy controls (Chen et al., 2018; Hu et al., 2017), while in other cases it decreases (Andrés-Benito et al., 2017) or does not vary at all (Vlam et al., 2015).
Finally, IFNs regulate several processes, such as pathogen recognition, antigen processing and presentation, inhibition of viral or cellular proliferation, apoptosis, immunomodulation, and leukocyte trafficking (Schroder et al., 2004). IFN-γ is the only IFN that was found to be altered in ALS (Moreno-Martinez et al., 2019). It is known that high levels of IFN-γ in the blood and CSF of ALS patients are associated with faster disease progression and shorter survival (Liu et al., 2015). However, its levels may also be lower in the blood and the CSF of ALS patients (Lu et al., 2016), or not vary at all (Hu et al., 2017).
The traditional view of ALS as a pathological condition affecting only the motor areas has been largely overcome (Rusconi et al., 2017; Smith et al., 2019). Studies of diffusion-based magnetic resonance imaging (MRI) tractography have demonstrated that ALS is a multisystem disorder that exhibits alterations in both motor and extramotor brain areas (Tu et al., 2018), involving cognitive domains from the early stages of the disease (Trojsi et al., 2017). Accordingly, several studies of functional magnetic resonance imaging (fMRI), electroencephalography, and magnetoencephalography (MEG) have shown that functional modification in ALS is widespread throughout the brain, and not limited to motor areas. However, fMRI studies on ALS have shown partially contradictory results, indicating both an increase and a decrease in connectivity depending on the different brain areas (Agosta et al., 2013; Chiò et al., 2014; Trojsi et al., 2017).
Recently, we conducted a study on ALS patients where we evaluated functional brain connectivity using MEG, demonstrating that the pathology is associated with a global brain hyperconnectivity, with an increasingly compact, less flexible, and more vulnearble network as ALS progresses (Sorrentino et al., 2018). However, while neuroimaging studies have helped elucidating the widespread nature of ALS, they did not shed light on the underlying pathogenic mechanisms.
Numerous evidences suggest that brain networks are highly sensitive to molecular changes occuring peripherally in both physiological and pathological conditions. For instance, the serum concentration of a systemic molecule such as IGF-1 relates to the (global) rearrangements of the brain network topology (Sorrentino et al., 2017). A broad spectrum of structural, functional, and metabolic neuroimaging findings is documented in response to dysmetabolic diseases such as liver or kidney disease, diabetes, and obesity (for recent reviews, see Guzzardi and Iozzo, 2019; Tagliapietra and Monaco, 2020; Viggiano et al., 2020).
Even more so, brain network dynamics is constrained by molecular mechanisms that occur primarily in the brain. As an example, misfolded and aggregated proteins that represent the molecular basis of different neurodegenerative conditions, such as Alzheimer's disease (AD) or Parkinson's disease (PD), determine changes in brain networks. In AD and in PD, the altered metabolism of tau and β-amyloid or α-synuclein causes numerous structural and functional brain changes. Importantly, the changes in the networks are often associated with the clinical picture, such as the cognitive decline in AD (Jacini et al., 2018) or the motor impairment in PD (Sorrentino et al., 2019). In conclusion, identifying associations between molecular processes and network rearrangements can be relevant not only on theoretical grounds, by improving the understanding of the pathophysiology, but also on practical grounds, as it might help the diagnostic management, the disease monitoring, and possibly therapeutic approaches.
The inflammatory processes, and even more so the neuroinflammatory ones, are extremely complex especially in terms of molecular pathways that can express dual neuroprotective/neurotoxic mechanisms. For example, the early stages of the ALS are characterized by the presence of neuroprotective anti-inflammatory mediators, whereas during later stages, cytotoxic proinflammatory mediators are largely predominant (Beers et al., 2011). Anti-inflammatory microglia isolated from early-stage ALS mouse spinal cords protect motor neurons in culture, while microglia isolated from late-stage ALS mouse spinal cords exhibit a proinflammatory phenotype and are toxic to motor neurons. This complex picture has made it difficult to identify a single marker of neuroinflammation and hence to study the relationship between inflammatory molecules and rearrangements in large-scale brain networks.
Our work aimed at verifying if blood cytokine levels could be a suitable peripheral biomarker of the complex neurodegenerative mechanisms of ALS, which reflect themselves into changes in brain topological parameters. To this aim, we evaluated blood cytokine levels (IL-2, IL-4, IL-1β, TNF-α, and IFN-γ) in 35 ALS patients and 30 matched controls. Furthermore, topological parameters in the classical frequency bands (delta, theta, alpha, beta, and gamma) of the resting-state, source-level, MEG-based brain networks have been computed to test if such parameters correlate with cytokine levels.
Materials and Methods
Participants
Thirty-five ALS patients (26 males, 9 females; mean age ± SD, 59.61 ± 18.27) were recruited from the ALS Center of the First Division of Neurology of the University of Campania “Luigi Vanvitelli” (Naples, Italy). Patients were right-handed and native Italian speakers with diagnosis of ALS, according to the revised El-Escorial criteria of ALS (Brooks et al., 2000). None of the patients showed any mutation in the screened genes SOD1, TARDBP, FUS/TLS, and C9ORF72. Thirty age-matched healthy controls (14 males, 16 females; mean age ± SD, 59.53 ± 13.41) were also included in the study. More clinical details and descriptive information about the cohort are shown in Table 1. Inclusion criteria were as follows: (1) no major medical illness and no use of substances that could interfere with MEG signals; (2) no other major systemic, psychiatric, or neurological diseases; and (3) no focal or diffuse brain damage at routine MRI.
Demographic and Clinical Characteristics of Amyotrophic Lateral Sclerosis Patients and Healthy Controls Recruited in the Study
p < 0.01.
ALS, amyotrophic lateral sclerosis; ALSFRS-R, Amyotrophic Lateral Sclerosis Functional Rating Scale-Revised; ECAS, Edinburgh Cognitive and Behavioural ALS Screen; LMN, lower motor neuron; SD, standard deviation; UMN, upper motor neuron.
The study protocol was approved by the local ethics committee and all participants provided written informed consent in accordance with the Declaration of Helsinki.
Cytokine assay
Blood sample collection
All patients and controls enrolled in the study were subjected to venous blood sampling. Whole blood samples were collected in S-Monovette tubes (Sarstedt), containing gel with clotting activator to facilitate the separation of serum from the cellular fraction. To this aim, samples were centrifuged at 4000 rpm for 10 min, then the serum was collected, aliquoted in 1.5 mL tubes (Sarstedt), and stored at −20°C.
Enzyme-linked immunosorbent assays
Serum concentrations of the cytokines IL-2, IL-1β, TNF-α, IFN-γ, and IL-4 were determined by enzyme-linked immunosorbent assay (ELISA) commercial kits (Diaclone SAS, Besançon cedex, France) according to the manufacturer's instructions. All tests were performed in duplicate. The ranges of the sensitivity standard curve of the ELISA kits were 31.2–1000 pg/mL for IL-2, 15.6–500 pg/mL for IL-1β, 25–800 pg/mL for TNF-α, 12.5–400 pg/mL for IFN-γ, and 1.1–35 pg/mL for IL-4. Standard diluents, capture antibody, and detection antibody were obtained as a kit for each cytokine. Standard diluent–serum sample was used to obtain a Cedex standard curve. The samples in the multiwell plate were mixed by repeated aspirations and ejections, taking care not to scratch the inner surfaces. Freeze-dried control vials were also reconstituted adding the most appropriate standard diluent to the sample. Biotinylated anticytokines, biotinylated secondary antibody, and streptavidin-HRP also were prepared according to the manufacturer's protocol. The absorbance value of each well was read on a spectrophotometer Thermo Scientific Multiskan GO, using 450 nm as the primary wavelength and optionally 620 nm as the reference wavelength (610–650 nm as acceptable).
Network analysis
MRI acquisition
MRI images of 31 ALS patients and 24 controls were acquired on a 3-T scanner equipped with an 8-channel parallel head coil (General Electric Healthcare, Milwaukee, WI). MR scans were acquired after the MEG recording or at least a month before. In particular, three-dimensional T1-weighted images (gradient-echo sequence inversion recovery prepared fast spoiled gradient recalled-echo, time repetition = 6988 ms, TI = 1100 ms, TE = 3.9 ms, flip angle = 10, voxel size = 1 × 1 × 1.2 mm3) were acquired. The remaining participants (four patients and six controls) did not complete the MRI because of the difficulty in lying down or because they refused to perform the MRI scan. In this case, we used a standard MRI template.
MEG acquisition
The MEG system was developed at the Institute of Applied Sciences and Intelligent Systems “E. Caianiello” of the National Research Council of Pozzuoli, Naples. The system is equipped with 163 magnetometers (Lardone et al., 2018) and it is placed at the Institute of Diagnosis and Treatment Hermitage Capodimonte (Naples), in a magnetically shielded room (ATB, Ulm, Germany) to reduce background noise. Acquisition, preprocessing, and source reconstruction were performed as previously described (Jacini et al., 2018). In short, before each acquisition, four reference positions (nasion, right, and left preauricolar and apex) were digitalized on the subject's head using Fastrak (Polhemus®). Electrocardiographic and electro-oculographic signals were corecorded to remove artifacts arising from heartbeat and blinking (Gross et al., 2013).
The brain activity of each subject was recorded in four segmentes of 2.5 min each, alternating closed eyes and opened eyes (Fig. 1a). Data from MEG acquisitions were acquired with a sampling frequency of 1024 Hz. Subsequently, the signal was filtered using a fourth-order Butterworth IIR band-pass filter in the 0.5–48 Hz band (Rucco et al., 2019).
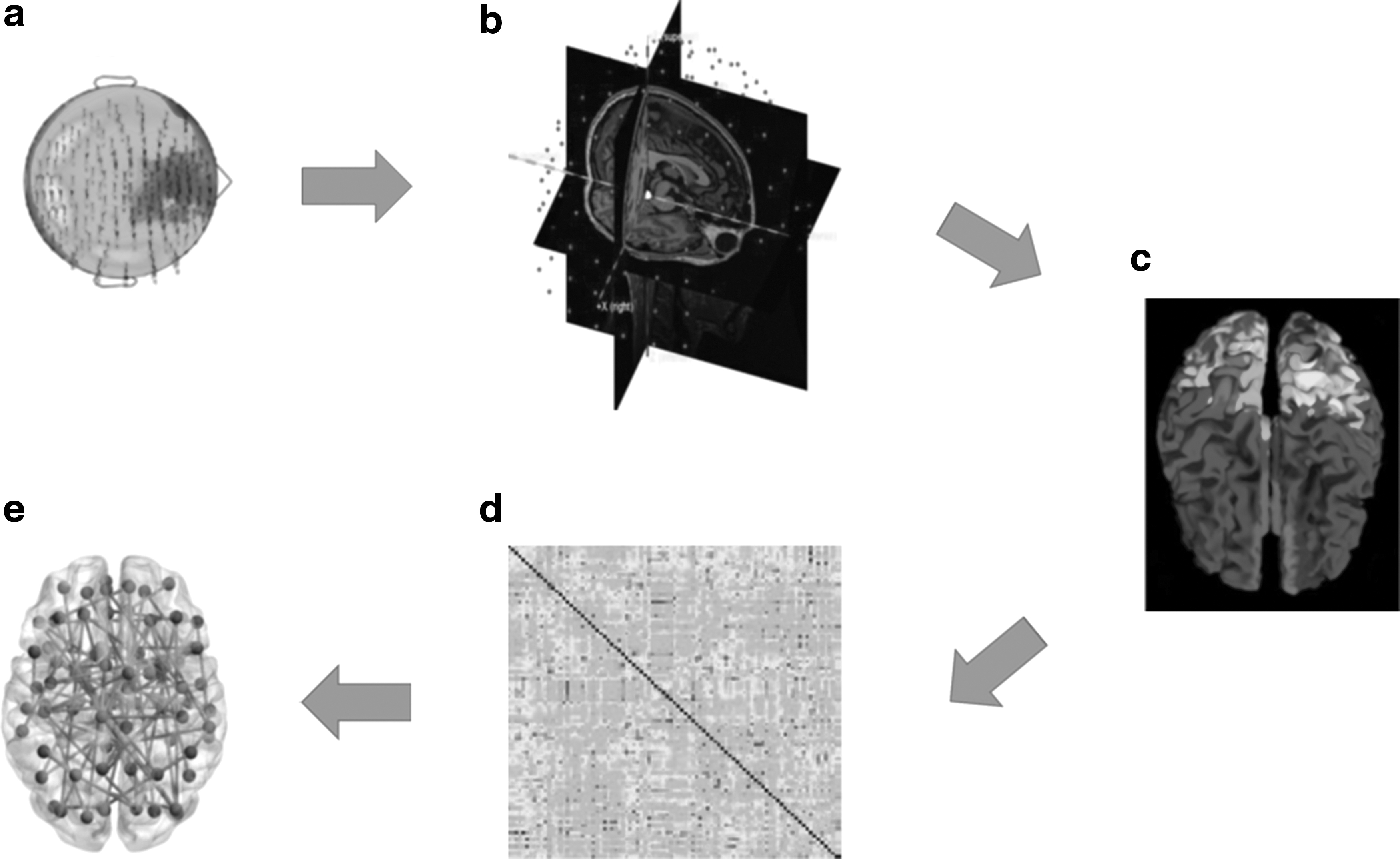
Schematic illustration of MEG data reconstruction.
Data preprocessing
At this stage, the principal component analysis and independent component analysis were used to remove environmental noise and physiological noise. The first clean 10 epochs of 8 sec were selected. The length of 8 sec is a good compromise to have enough cleaned epochs and avoid drowsiness (Gross et al., 2013) while obtaining a reliable estimate of the connectivity measure (Fraschini et al., 2016).
Source reconstruction
The beamforming procedure has been performed using the Fieldtrip toolbox (Oostenveld et al., 2011), similarly as in Sorrentino and colleagues (2018). In particular, the brain volume conduction model proposed by Nolte (2003) was used in a linearly constrained minimum variance beamformer (Van Veen et al., 1997), to reconstruct the time series related to the centroids (Hillebrand et al., 2016) of 116 regions-of-interest (ROIs), derived from the Automated Anatomical Labeling (AAL) atlas (Gong et al., 2009; Tzourio-Mazoyer et al., 2002) (Fig. 1b). We decided to exclude cerebellar areas (hence considering 90 instead of 116 ROIs) given their lower reliability (Brookes et al., 2011). For each source, we projected the time series along the dipole direction that explains the most variance by means of singular value decomposition (Fig. 1c). Source time series were resampled at 512 Hz.
Connectivity and network analysis
The phase lag index (PLI) was used to estimate functional connectivity (Stam et al., 2007), using BrainWave software (CJS, version 09.152.1.23). The PLI is based on the distribution of the differences of the instantaneous phases [ΔΦ (t)] (derived from the Hilbert transform of the time series) computed as follows:

where “< >” indicates the mean value, “sign” stands for the signum function, “||” denotes the absolute value, and “tk” are the samples. The phase difference is defined in the [−π, π] range. PLI values range between 0 and 1, where 1 indicates perfect synchronization and 0 indicates nonsynchronous activity. By computing the PLI for each couple of brain regions, we obtained a 90 × 90 weighted adjacency matrix (Fig. 1d) for each epoch and for each subject, in all the frequency bands: delta (0.5–4.0 Hz), theta (4.0–8.0 Hz), alpha (8.0–13.0 Hz), beta (13.0–30.0 Hz), and gamma (30.0–48.0 Hz). The weighted adjacency matrix was used to reconstruct a network (Stam, 2014), where the 90 areas of the AAL atlas are represented as nodes and the PLI values form the weighted edges.
A frequency-specific minimum spanning tree (MST) was calculated for each epoch (Fig. 1e). Since we were interested in the strongest connections, for the construction of the MST, the edge weight was defined as 1/PLI. In fact, Kruskal's algorithm (Kruskal, 1956) first ranks the links in ascending order and then constructs the network by adding one link at a time, discarding links that would form a loop. The algorithm proceeds until all nodes are connected resulting in a loopless graph with N nodes and M = N − 1 links.
We used the MST to avoid some of the biases in traditional network analyses. In fact, graph Theoretical measures are affected by the number of nodes and the average degree, in a way that depends on the underlying network topology (Van Wijk et al., 2010). Even correcting using random surrogates might not be an optimal solution (Van Wijk et al., 2010). We used the MST as a way to obtain comparable networks in terms of size and density, at the cost of possibily discarding some information (Tewarie et al., 2015).
Based on the MST, we calculated both global (degree divergence, degree correlation/disassortativity, leaf fraction, tree hierarchy, and diameter) and nodal parameters (degree, betweenness centrality, and eccentricity) (Boersma et al., 2013; Tewarie et al., 2015).
To understand their role in the interpretation of our connectivity data, it will be useful to define each global parameter of interest: (1) the leaf fraction is defined as the fraction of nodes with degree 1 (Boersma et al., 2013), providing an indication of the integration of the network. In particular, higher leaf fraction implies that the network tends toward a star-like topology, where the nodes are closer to each other (Rucco et al., 2019). In line-like networks, going from one node to another takes on average more steps compared with more star-like networks. However, the star-like topology shows less resilience to targeted attacks to the most central nodes. Presumably, the healthy brain reaches an optimal trade-off between these two extreme conditions; (2) the tree hierarchy is a parameter specifically designed to capture such trade-off; (3) the diameter is defined as the longest among the shortest paths of an MST and it is related to the leaf fraction. In fact, when the leaf number increases (star-like network), the value of the diameter decreases (Tewarie et al., 2015); (4) the degree divergence is a measure of the broadness of the degree distribution, related to the presence of specifically important nodes (hubs), and hence to the resilience against attacks, epidemic spreading, and the synchronizability of complex networks (Tewarie et al., 2015).
Statistical analyses
The ELISA results are all reported in pg/mL. For each of the five cytokines, the average values and the standard deviations (SDs) were calculated, for both patients and healthy controls. After verifying that the data were normally distributed, we performed a Student's t-test to evaluate possible significant differences between the two groups, for each cytokine, using a significance level of p ≤ 0.05. We used the false discovery rate (FDR) (Benjamini and Hochberg, 1995) to correct for multiple comparisons, with a significance level of p ≤ 0.05.
All statistical analyses on topological data were performed in MATLAB (MathWorks®, version R2013a). To compare data between ALS patients and healthy controls, for each frequency band and for each topological parameter, we used permutation testing (Nichols and Holmes, 2002) where the null distribution for between-group differences is derived from the data. In details, assuming no group differences, the labels of the subjects were permuted 104 times. Each time, the difference between the averages of the two groups was computed, obtaining the null distribution for between-group differences. Such distribution was used to define the statistical significance of the observed difference between the two groups. We used the FDR with a significance level of p ≤ 0.05.
Finally, to explore the relationship between biochemical results (cytokines) and topological data, we built a multivariate linear model. Colinearities among the predictors were explored as a preliminary step, following which a forward stepwise procedure was used to add the predictors. Age, gender, education, ALSFRS, and the cytokines were added as predictors. The adjusted R 2 was used to evaluate the performance of the model.
Results
ELISAs performed on serum samples to evaluate the level of IL-2, IL-4, IL-1β, IFN-γ, and TNF-α of both ALS patients and healthy controls show that there are significant differences in the serum levels of IL-1β (Fig. 2a, p = 0.01, *pFDR = 0.03), IL-4 (Fig. 2b, p = 2 × 10−6, ***pFDR = 1 × 10−5), and IFN-γ (Fig. 2c, p = 0.03, *pFDR = 0.05) between patients and controls. In particular, the levels of interleukines IL-1β and IL-4 are increased in ALS patients compared with controls, while the levels of IFN-γ are higher in healthy controls.
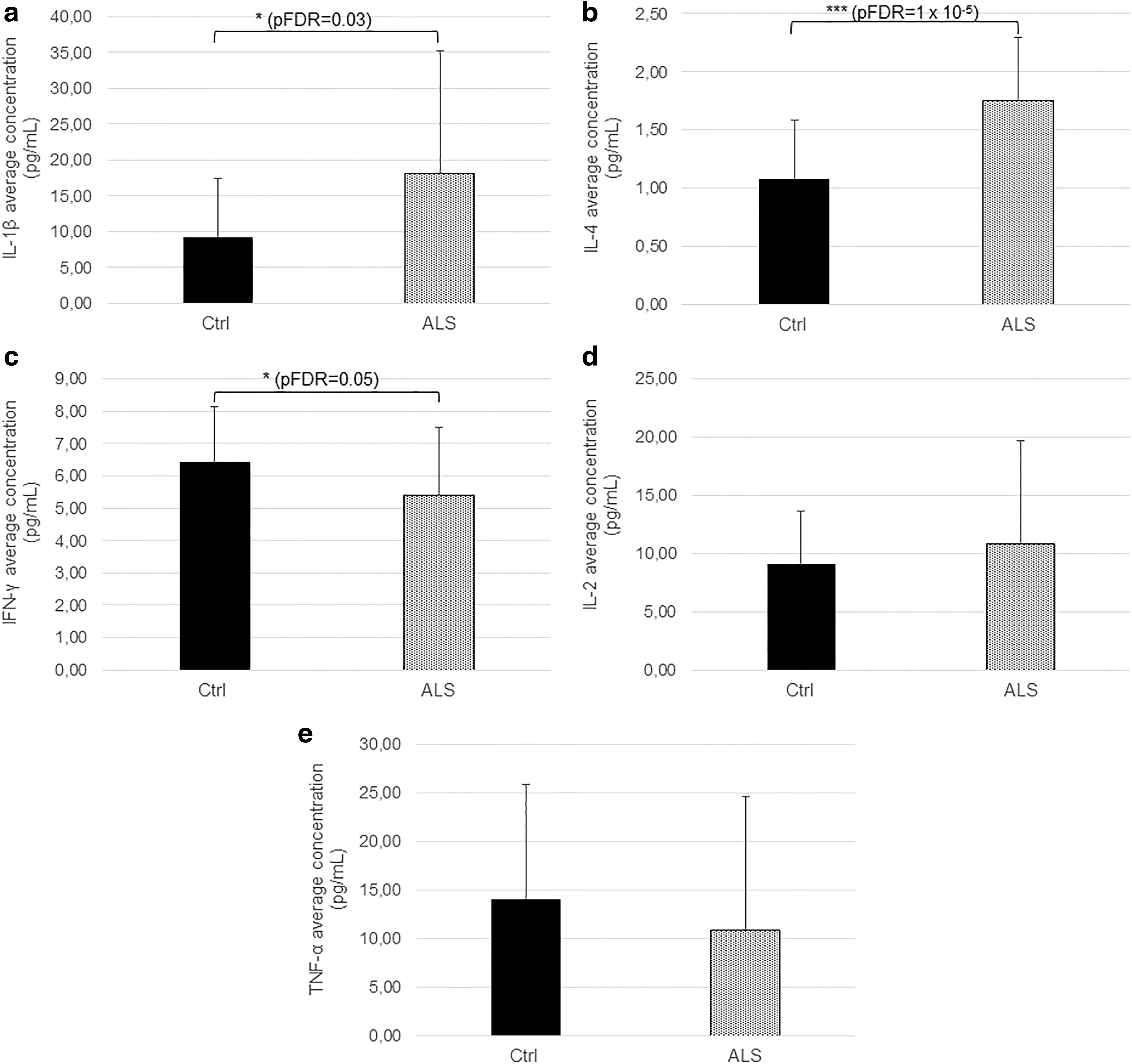
Evaluation of the serum level of cytokines in ALS patients (ALS) and healthy controls (Ctrl). Histograms show that the levels of IL-1β
Conversely, there are no significant differences between the two groups with respect to levels of cytokines IL-2 (Fig. 2d, p = 0.33, pFDR = 0.33) and TNF-α (Fig. 2e, p = 0.33, pFDR = 0.33).
With regard to the brain network analysis, after building a PLI-based adjacency matrix, we used an MST-based network approach to compare global (noncentrality) topological parameters in ALS patients and healthy controls.
In particular, in the theta frequency band, the two groups differ in the noncentrality parameters degree divergence (p = 0.01, *pFDR = 0.03), leaf fraction (p = 0.02, *pFDR = 0.03), and diameter (p = 0.02, *pFDR = 0.03) (Fig. 3a). In particular, the degree divergence and the leaf fraction increase in patients, while the diameter is lower compared with controls.
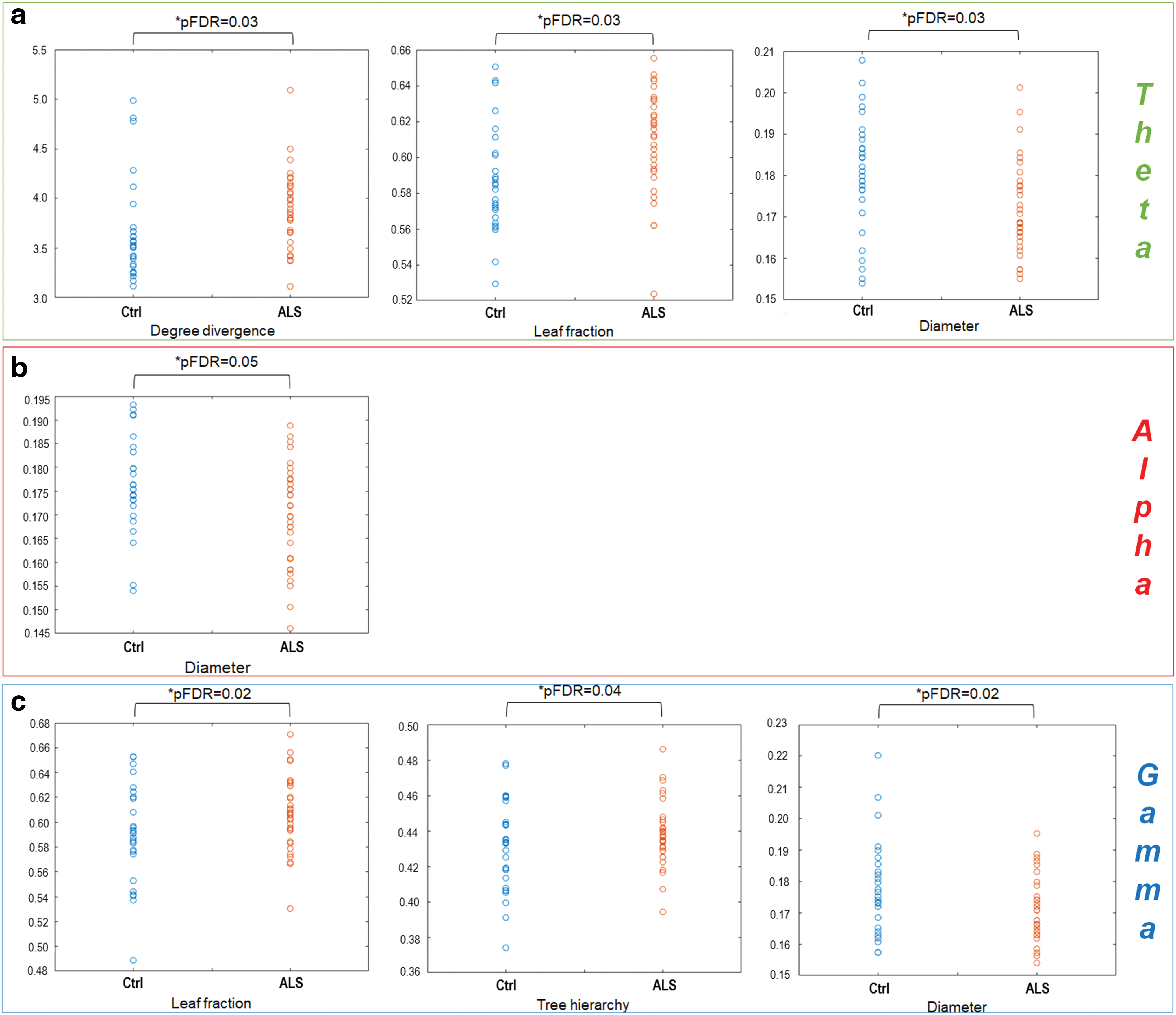
Network parameters in ALS patients (red circles, ALS) and healthy controls (blue circles, Ctrl).
In the alpha band, we observed significant differences between the two groups with respect to the diameter, a global parameter, which decreases in ALS patients compared with healthy controls (p = 0.01, *pFDR = 0.05). These data are shown in Figure 3b.
Significant differences also appeared with respect to the leaf fraction (p = 0.01, *pFDR = 0.02), tree hierarchy (p = 0.02, *pFDR = 0.04), and the diameter (p = 0.01, *pFDR = 0.02) in the gamma band (Fig. 3c). In particular, we registered an increase in the leaf fraction and tree hierarchy of ALS patients, while the diameter decreases in the pathological condition.
Despite the significant differences observed between the two groups in both cytokine levels and topological parameters, the linear model showed that cytokines fail to predict the topological properties of the brain networks with the adjusted R 2 invariably below 0.2 (data not shown). Finally, we did not find any significant correlation between blood cytokines or topological features and cognitive profile as evaluated by the Edinburgh Cognitive and Behavioural ALS Screen (ECAS).
Discussion
The objective of this study was to verify if neuroinflammatory mechanisms observed in ALS reflected themselves in alterated levels of blood cytokines and if these humoral modifications were related to changes in the brain connectivity. To this aim, the serum levels of five cytokines (IL-2, TNF-α, IL-1β, IL-4, and IFN-γ) were assessed as potential biomarkers of neuroinflammation (McCombe and Henderson, 2011; Robelin and Gonzalez De Aguilar, 2014). Furthermore, we used MEG data to evaluate changes in the topological parameters of the brain networks. The relationship between the cytokines and the topological parameters that were altered in ALS patients compared with healthy controls was investigated.
Comparing the cytokine levels, we found significant differences in the serum levels of IL-1β, IL-4, and IFN-γ between patients and healthy controls.
In particular, we observed increased IL-1β and IL-4 levels in ALS patients, which is in line with previous studies. In fact, high blood levels of IL-1β (Hu et al., 2017) result in decreased neurogenesis and increased gliogenesis (Borsini et al., 2015), leading to amplified inflammatory processes. This is true in both ALS and other neurodegenerative diseases, such as AD and PD (Prado et al., 2018), or neuropsychiatric diseases, such as depression (Borsini et al., 2015). This aspecificity would suggest that although the inflammatory process in ALS is demonstrated by autoptic tissues (Engelhardt et al., 1993; Graves et al., 2004; Henkel et al., 2004; Kawamata et al., 1992), it is not a peculiarity of the disease. Such finding might support the idea that the inflammatory reaction is not a causative event of neuronal death. On the contrary, high levels of IL-4 in the blood or CSF are associated in ALS with disease progression (Moreno-Martinez et al., 2019), and can be produced and overexpressed in relation to muscle hyperexcitability (Peake et al., 2015).
With regard to IFN-γ, we found a decrease in ALS patients, and this may be related to its complex and pleiotropic role. In fact, since IFN-γ participates in both the activation of macrophages and the adaptative T cell immune response (and to several other processes, such as pathogen recognition, antigen processing and presentation, inhibition of viral or cellular proliferation, apoptosis, immunomodulation, and leukocyte trafficking) (Schroder et al., 2004), its serum levels could increase (Liu et al., 2015), decrease (Lu et al., 2016), or not vary at all (Hu et al., 2017) in the blood and CSF of ALS patients. These controversial results could be due to the fact that each process is involved in different stages of the disease, with serum levels of IFN-γ varying as a function of disease stage.
We did not find any significant differences in IL-2 and TNF-α serum levels between the two groups. Our results are partially in accordance with previous studies. In fact, high plasma levels of IL-2 seem to have a “protective role” in the incidence of ALS (Prado et al., 2018) in some cases, while in other cases, they seem to correlate with a lower survival rate (Moreno-Martinez et al., 2019). These controversial results may be due to the fact that this cytokine does not appear to have a direct pathogenic role in ALS, but probably modulates the disease process by activation of natural killer cells, known to be cytotoxic for a wide range of neurons (Lu et al., 2016).
The role of TNF-α in ALS pathogenesis is also still unclear because of its pleiotropic actions (Moreno-Martinez et al., 2019). In some cases, the TNF-α level increases in the CSF or blood of ALS patients compared with controls (Chen et al., 2018; Hu et al., 2017), while in other cases it decreases (Andrés-Benito et al., 2017) or does not vary at all (Vlam et al., 2015). The difficulty in interpreting TNF-α results partly lies in the cellular pathways in which it is involved and in those that it can activate (Tortarolo et al., 2017). In fact, TNF-α has a crucial role in regulating macrophage biology in many processes of ALS and several other diseases, including rheumatoid arthritis, Crohn's disease, atherosclerosis, psoriasis, sepsis, diabetes, and obesity (Parameswaran and Patial, 2010), and its serum levels may vary depending on the biological processes involved.
We applied network analysis to source-level MEG data obtained from a cohort of ALS patients and healthy controls in resting state. We found significant changes in global metrics of ALS patients, which capture widespread reorganization in the brain activity. In particular, topological parameters are altered in processes causing diffuse damage to the brain, such as neurodegenerative diseases, including ALS (Sorrentino et al., 2018; Trojsi et al., 2017; Yu et al., 2016).
In this study, we registered significant changes in the degree divergence, leaf fraction, and diameter in the theta band, in the diameter in the alpha band, and in the leaf fraction, tree hierarchy, and the diameter in the gamma band.
In particular, the leaf fraction increases in ALS patients (in gamma and theta bands) compared with healthy controls, which implies a trend toward a star-like topology, and consequently a more centralized organization of the brain network (Boersma et al., 2013) with overload of central nodes.
The decrease in the diameter (in alpha, gamma, and theta bands) observed in patients compared with controls is in line with the increase in the leaf fraction, and also points toward a more compact and therefore a more vulnerable network (Tewarie et al., 2015).
Furthermore, the degree divergence (broadness of the degree distribution) increases in patients in the theta band. Higher values of degree divergence imply the presence of high-degree nodes (Boersma et al., 2013), and this is once again in line with a more centralized network. The high-degree nodes allow a more rapid synchronization while making the network more vulnerable to targeted attacks (Tewarie et al., 2015).
Last, the increased tree hierarchy in patients (in the gamma band) suggests that the disease leads to a suboptimal trade-off between having a well-integrated network and while preventing node overload. This condition might be due to a more centralized network, as described above, which is more susceptible to node overload.
All together, these topological data suggest that the brain in ALS shows functional hyperconnectedness. Such hyperconnectivity might be deleterious as it would not allow the network to display flexible dynamics, as the activity of any area would be too tightly constrained by the activity of the other brain areas.
To verify if the blood cytokine level could be a suitable peripheral biomarker acting as a surrogate of neurodegeneration in ALS, we evaluated its possible correlation with modifications of the brain network that were previously shown to be altered as a function of ALS progression (Sorrentino et al., 2018).
The present work demonstrates that circulating cytokines are altered but do not correlate with brain topological parameters in ALS, also when adjusted for possible confounders such as gender, education, age, and clinical progression. This can be explained with some limitations regarding the use of peripheral biomarkers to study neurodegenerative diseases. Some biological events associated with cerebral inflammation may not be detectable in peripheral blood (D'Ambrosio et al., 2015), as the alteration of peripheral cytokines is not specific to a specific body district but rather it is an index of systemic inflammation. Therefore, they could be not specific enough to track the inflammation in the central nervous system.
Furthermore, as we have previously seen, cytokines display a pleiotropic role (Lai et al., 2017) and it is likely that the combined action of more cytokines, and not a single one, is associated with clinical outcomes (Cherbuin et al., 2019).
Finally, the lack of correlation between blood cytokine levels and cognitive (ECAS) or functional (ALSFR-S) parameters suggests that the changes observed in patients compared with controls do not reflect directly the disease progression or the cognitive impairment.
However, the lack of correlation does not rule out the possibility that inflammatory processes might contribute to brain changes as recently reviewed by Albrecht and colleagues (2016). Hereafter, the integration of the diffusion MRI, a powerful approach in studying network topology in health (Gong et al., 2009) and disease (Mishra et al., 2020), with MEG may be able to further investigate the possible effects of inflammation on the brain topological rearrangements.
Conclusions
In this article, we evaluated blood cytokine levels and estimated the connectivity among brain areas of ALS patients through MEG, finding a widespread topological reorganization of the brain toward a more integrated and more vulnerable network in the pathological condition. Furthermore, we found that although the levels of specific blood cytokines differ between ALS patients and healthy controls, they do not correlate with the changes observed in the functional brain networks. Our results cast doubt on the hypothesis that peripheral cytokines could be used as biomarkers of neuroinflammation. However, the absence of correlations between serum levels of cytokines and functional connectivity data does not rule out the hypothesis that inflammatory processes might contribute to the alteration of the brain network in ALS.
Footnotes
Authors' Contributions
A.P. and R.R. contributed equally to this work as cofirst authors, performing all data collection and analysis and writing the article. A.P. contributed to the collection and storage of blood samples. I.S., M.G., and A.M.D. contributed to designing and performing cytokine assays. R.M., M.P., and P.S. contributed to the collection and analysis of MEG data. S.B. and F.T. contributed to the article revision. G.S. and P.S. contributed to MEG data interpretation and article design and revision. All authors read and approved the submitted version.
Author Disclosure Statement
The authors declare that they have no conflicts of interest and that no competing financial interests exist. All participants included in this study provided written informed consent in accordance with the Declaration of Helsinki.
Funding Information
This study was funded by University of Naples Parthenope within the Project “Bando Ricerca Competitiva 2017” (D.R. 289/2017). We thank Enrico Amico for the valuable discussion on the statistical analysis.