
Abstract
Background:
The fornix is the primary axonal tract of the hippocampus, connecting it to modulatory subcortical structures. This review reveals that fornix damage causes cognitive deficits that closely mirror those resulting from hippocampal lesions.
Methods:
We reviewed the literature on the fornix, spanning non-human animal lesion research, clinical case studies of human patients with fornix damage, as well as diffusion-weighted imaging (DWI) work that evaluates fornix microstructure in vivo.
Results:
The fornix is essential for memory formation because it serves as the conduit for theta rhythms and acetylcholine, as well as providing mnemonic representations to deep brain structures that guide motivated behavior, such as when and where to eat. In rodents and non-human primates, fornix lesions lead to deficits in conditioning, reversal learning, and navigation. In humans, damage to the fornix manifests as anterograde amnesia. DWI research reveals that the fornix plays a key role in mild cognitive impairment and Alzheimer's Disease, and can potentially predict conversion from the former to the latter. Emerging DWI findings link perturbations in this structure to schizophrenia, mood disorders, and eating disorders. Cutting-edge research has investigated how deep brain stimulation of the fornix can potentially attenuate memory loss, control epileptic seizures, and even improve mood.
Conclusions:
The fornix is essential to a fully functioning memory system and is implicated in nearly all neurological functions that rely on the hippocampus. Future research needs to use optimized DWI methods to study the fornix in vivo, which we discuss, given the difficult nature of fornix reconstruction.
Impact Statement
The fornix is a white matter tract that connects the hippocampus to several subcortical brain regions and is pivotal for episodic memory functioning. Functionally, the fornix transmits essential neurotransmitters, as well as theta rhythms, to the hippocampus. In addition, it is the conduit by which memories guide decisions. The fornix is biomedically important because lesions to this tract result in irreversible anterograde amnesia. Research using in vivo imaging methods has linked fornix pathology to cognitive aging, mild cognitive impairment, psychosis, epilepsy, and, importantly, Alzheimer's Disease.
Introduction
Seminal work showing that bilateral damage to the hippocampus causes anterograde amnesia (Milner and Scoville, 1957) focused the lens of episodic memory research on the hippocampus, often to the exclusion of other connected brain regions. However, anterograde amnesia can arise from focal damage to other parts of the brain. For instance, prolonged deficiencies in thiamine can cause diencephalic amnesia, which is associated with destruction of the mammillary bodies and/or portions of the thalamus (Gudden, 1896). Anterograde amnesia can also appear after discrete lesions to the basal forebrain (Abe et al., 1998). All of these brain regions are linked together by a C-shaped white matter pathway called the fornix. Importantly, fornix damage alone can cause severe and unremitting anterograde amnesia.
However, fornix damage in humans is exceedingly rare. Until recently, it was difficult to know whether the wealth of findings about the fornix from rodents and nonhuman primates extended to humans. We can now use noninvasive imaging techniques—for example, diffusion weighted imaging (DWI)—to gain insight into the functional properties and pathological processes that are associated with various white matter tracts. The goal of this review is to provide a detailed summary of the functions and processes supported by the fornix to help researchers interpret their results and plan hypothesis-driven DWI research. To achieve this goal, we describe the anatomy of this tract and detail the effects of fornix lesions on memory and other cognitive domains in both humans and animals. In addition, we review the small diffusion imaging literature (keywords used to find studies: fornix, structural connectivity, white matter, white matter connectivity, diffusion imaging, diffusion tensor imaging, DWI, memory, episodic memory, amnesia, Alzheimer's disease [AD], mild cognitive impairment [MCI]), discuss the potential information transmission properties of this white matter tract in episodic memory, and point out future directions.
Anatomy of the Fornix
The fornix, illustrated in Figure 1, constitutes the primary efferent and afferent pathway of the hippocampus and is a central component of the Papez Circuit (Amaral and Lavenex, 2007). As an efferent pathway, the fornix begins as the fimbria, the attached white matter of the hippocampus; then, it becomes a detached bundle, forming the fornical crus. The two crura join beneath the splenium of the corpus collosum to form the body of the fornix (Catani et al., 2002; Douet and Chang, 2015). These fibers descend into the forebrain to become the columns of the fornix, which split around the anterior commissure. The columns then split further into the precommissural fibers—which innervate the septal nuclei, diagonal band of Broca, and the nucleus accumbens—and the postcommissural fibers—which extend to the mammillary bodies of the diencephalon. The postcommissural fibers divide into two smaller bundles—one that innervates several anterior hypothalamic areas, and one that carries fibers to the anterior thalamic nuclei (Amaral and Lavenex, 2007).

Anatomy of the human fornix.
The fornix is bidirectional—it also contains fibers leading from these subcortical regions to the hippocampus. Most external modulators of the products of hippocampal processing—namely the septal nuclei/diagonal band of Broca, locus coeruleus, and raphe nuclei (the latter two regions synapse onto the septal nuclei before progressing to the hippocampus)—enter the hippocampal formation via the fornix (Amaral and Lavenex, 2007).
There is some disagreement about whether the hippocampus projects to the medial prefrontal cortex (mPFC) via the fornix. Studies using histological techniques, which are superior to magnetic resonance imaging (MRI) techniques in their level of granularity and should be considered the “ground truth,” report that axons from CA1/subiculum project directly to the orbitofrontal cortex/mPFC through the fimbria/fornix, but these axons do not constitute the fornix proper. Instead, these axons constitute a structure called the hippocampal-to-prefrontal cortex (PFC) pathway (Godsil et al., 2013). This pathway has primarily been studied in rodents; it awaits further elucidation in humans. We note that a recent study by Aggleton and colleagues (2015) performed histology and reported findings that differ from the older histology findings. Nevertheless, the classic findings and definitions of the fornix, as well as the atlases, do not provide any evidence that the fornix connects the hippocampus to the mPFC.
Putative Functions of the Fornix
Episodic memory
The literature on the fornix consistently and overwhelmingly links it to long-term forms of memory. In the next sections, we review the relevant literature on nonhuman animals and humans.
Lesions of the fornix in nonhuman animals
There is a long history of creating lesions in the fornix of rodents and nonhuman primates to investigate the causal role of the fornix in long-term forms of memory. In these studies, animals are first trained on a task to a learning criterion, after which they undergo a surgical procedure to sever the fornix, while leaving the hippocampal formation intact. After a period of recovery, the animals are sometimes retested on the previously learned task to test for retrograde amnesia. They are then taught a new task, after which they are tested to determine whether they were able to acquire new learning, or whether they are inhibited by anterograde amnesia. The memory tasks used in nonhuman primates and rodents are generally spatial or navigational in nature, although some are simply visual recognition tasks. Spatial and navigational tasks require the encoding and retrieval of spatiotemporal context and, therefore, serve as a proxy for episodic memory.
Do fornix lesions in nonhuman animals cause deficits on traditional “hippocampally-dependent” tasks? A rich intellectual history links trace conditioning, contextual fear conditioning, reversal learning, and spatial tasks to hippocampal processing. Performance on all of these tasks is severely affected by lesions to the fornix (detailed in Table 1). For instance, one study found that fornix lesions disrupted rabbits' ability to learn a conditioned-unconditioned stimulus association when there was a delay in a trace conditioning paradigm (Oswald et al., 2007). Another study used a contextual fear conditioning paradigm and found that rats with fornix lesions do not exhibit freezing behavior on being returned to the cage in which the conditioned and unconditioned stimuli were initially paired (Maren and Fanselow, 1997; Phillips and LeDoux, 1995) nor they do exhibit context-dependent freezing behavior that was previously extinguished (Ji and Maren, 2008).
Relationship Between the Fornix and Cognitive Performance
This table includes findings from nonhuman animals and lesion studies in humans. For findings from diffusion imaging studies in humans, see Table 2.
CVLT, California Verbal Learning Task; WMS, Weschler Memory Scale.
Animals with fornix transections can learn simple associations when they are repeated many times, but struggle to learn the reversal or reassignments, taking far longer to reach a criterion than control animals (Hirsh and Segal, 1972; Wiig et al., 1996; Wilson et al., 2007). The largest literature of all shows that fornix transection consistently leads to robust impairments in learning new routes and spatial locations, both in nonhuman primates (Kwok and Buckley, 2006; Murray et al., 1989) and in rodents (De Bruin et al., 2001; Ennaceur and Aggleton, 1997; Hirsh and Segal, 1972; O'Keefe et al., 1975; Walker and Olton, 1979; Whishaw and Tomie, 1997) (Fig. 2).
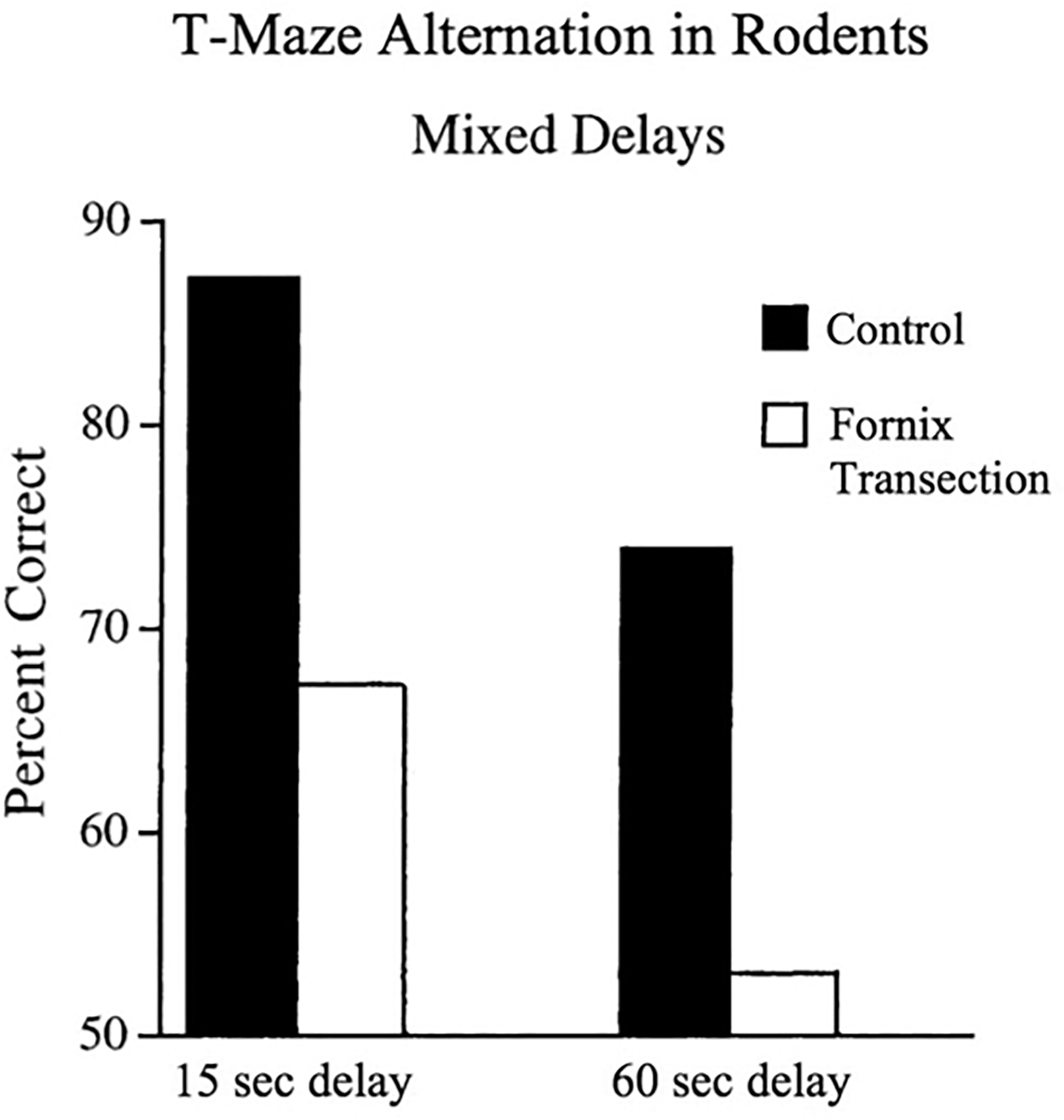
The effect of fornix transection on navigation in nonhuman primates. Compared with controls, rodents with fornix lesions perform significantly worse on navigational tasks at various delays. Reproduced with permission from Bussey et al. (2000).
Crucially, there is robust evidence that fornix lesions do not lead to deficits in nonspatial memory tasks such as object recognition in animals (Bussey et al., 2000; Ennaceur and Aggleton, 1997; Ennaceur et al., 1996; Gaffan, 1992; Markowska et al., 1989; Wilson et al., 2007). Simple object recognition is believed to be subserved by the perirhinal cortex rather than the hippocampus; however, there is some evidence to the contrary (Bachevalier et al., 1985; Browning et al., 2010; Gaffan, 1994), although the authors in these cases admit to “small and unreliable” effects, or use complex stimuli more akin to “scenes” than objects. Generally, simple object recognition is not impaired by fornix lesions, but once a spatial or contextual component is added to the task, there are consistent and robust deficits in performance.
Lesions of the human fornix
Humans rarely incur damage exclusive to the fornix. When it does occur, it is typically in the context of a colloid cyst of the third ventricle (Gaffan et al., 1991; Hodges and Carpenter, 1991; Poreh et al., 2006; Tsivilis et al., 2008), bilateral infarct of the subcallosal artery (Moudgil et al., 2000; Park et al., 2000), or traumatic brain injury (D'Esposito et al., 1995). Findings from this small clinical literature have documented that fornix lesions give rise to a persistent anterograde amnesia that is best described as a disconnection syndrome (Table 1). This memory deficit occurs against the backdrop of intact intellect and the absence of anosognosia. Working memory, procedural memory, and executive and language functioning (except for slightly reduced verbal IQ) (Hodges and Carpenter, 1991; Park et al., 2000) are generally in the normal range.
When the fornix is completely transected, memory deficits are stimulus invariant, such that they are observed for all types of visual and verbal materials. Even partial damage results in severe decrements, especially if the insult is incurred to the left fornix, given the lateralization of language abilities and the import of language to adequate functioning. There is some material specificity in the expected direction, with left fornix damage tending to lead to verbal memory deficits and right fornix damage being more associated with visual memory deficits (Gaffan et al., 1991; McMackin et al., 1995). However, it is worth noting that the extent of damage is more predictive of deficits than the lesion location for both verbal and visual tasks (Korematsu et al., 2010; Sara et al., 2004 for an animal model).
In fornix lesion cases, the episodic memory deficits are generally more profound for recall than for recognition (Calabrese et al., 1995; Tsivilis et al., 2008), although there are deficits in recognition memory as well (Calabrese et al., 1995; D'Esposito et al., 1995; Gaffan et al., 1991). The clinical profile does not typically include confabulation, distinguishing this form of amnesia from the diencephalic variant. Nevertheless, the episodic memory deficits after fornix lesions in humans are profound, unremitting, and bear an uncanny resemblance to the episodic memory deficits observed after focal bilateral hippocampal lesions.
The multi-trace theory of memory proposes that retrieving past experiences, even remote ones from childhood, necessitates the involvement of the hippocampus (Nadel et al., 2000). Interestingly, there is some evidence that fornix damage causes problems recollecting the far past—deficits in recalling preoperative learning in animals (Squire et al., 2001; Wiig et al., 1996; Winocur et al., 2001) and retrograde amnesia in humans (D'Esposito et al., 1995; Hodges and Carpenter, 1991; Poreh et al., 2006). We note that a few studies have tested this in humans, but those that have examined it have reported that the retrograde amnesia is primarily for episodic aspects of remote memory, not semantic aspects—thus, it is most commonly observed in autobiographical recollection (Hodges and Carpenter, 1991; Poreh et al., 2006). However, other studies find normal levels of preoperative memory in animals (Buckley et al., 2004, 2008; Gaffan et al., 2001) or no retrograde amnesia in humans (Park et al., 2000). In sum, the role of the fornix in accessing remote memories is under-developed, but potentially quite interesting.
How to measure the fornix in humans: DWI and volumetry
Lesion evidence provides strong inferential power about a region's function. However, because fornix lesions are so rare in humans, other means must be sought after to elucidate the role of the fornix in cognitive function. DWI has emerged as a powerful tool to measure the microstructure of brain white matter. This technique uses Brownian motion to create contrast in MRI images—in other words, it tracks the rate and direction that water molecules move in the brain. There are several indices of water diffusivity, with two of the most commonly used being fractional anisotropy (FA) and mean diffusivity (MD). The interpretation of these indices is not always straightforward, and there is not a one-to-one correspondence between any measure and any particular characteristic of white matter.
FA is a composite measure of the degree of anisotropy—asymmetry of water movement—in a given voxel, and it ranges from 0 (totally isotropic) to 1 (totally anisotropic). It is taken to indicate the degree of directional restriction of water diffusivity within a voxel. MD, on the other hand, is a measure of overall diffusivity (mean of three eigenvalues of the diffusion tensor) in a given voxel, with lower values indicating less diffusivity.
DWI has some known limitations. It is inferior to invasive histology methods used in nonhuman animals, because it cannot reveal the direction of information flow, and it cannot distinguish between the different fibers that constitute a pathway (e.g., excitatory vs. inhibitory, originating in the lateral septum vs. medial septum, etc.). DWI also cannot tell you the biological reason underlying differences in microstructure ( Filley and Fields, 2016; see Assaf et al., 2017; Jones et al., 2013; O'Donnell and Pasternak, 2015).
Another way to quantify the fornix is to use high-resolution T1 and T2 images and manually extract the macrostructure—or volume—of the fornix. A recent study used this method and found that fornix volume was associated with spatial memory and olfactory identification in young adults (Dahmani et al., 2020). Volumetric measurements are a less popular method than DWI, although they can provide useful insights, especially in certain clinical populations such as those with AD, where there is usually cell loss in the hippocampus and concomitant volume loss in the fornix. Researchers conducting hippocampal segmentation studies may wish to consider conducting volumetry of the greater fornix, given that the same MRI data and conceptually similar methods are used as in DWI. Mapping methods are described by Amaral and colleagues (2016)—the authors hand-segmented hippocampal subfields as well as the fornix and other neighboring structures on a subset of participants and then generated an atlas to use for automated segmentation of the entire dataset.
Much of the fornix is located in the third ventricle, inferior (and perpendicular) to the corpus callosum, and, as such, is completely surrounded by cerebrospinal fluid (CSF). This location makes the fornix particularly difficult to image due to ever-present susceptibility artifacts (Gunbey et al., 2014; Hodgetts et al., 2017, 2020; Oishi and Lyketsos, 2014; Sullivan et al., 2010). We discuss this issue and potential solutions in Box 1.
How to Measure the Fornix with Diffusion-Weighted Imaging
DWI of the fornix in development of episodic memory
The fornix is one of the first major white matter tracts to reach maturity. It can be identified in the fetal brain within the second trimester, and the entire fornical trajectory—the topology—is developed at 19 gestational weeks (Huang, 2010), setting the stage for its developmental course in the first years of life.
A small literature describes the development of the fornix and the relationship of DWI metrics to memory performance. Although it has been shown that fornix macrostructure is established by the time a human fetus reaches 19 gestational weeks (Huang, 2010), it is less clear when the microstructure reaches maturity. When looking at children beginning at 2 years of age, researchers found a relationship between age and increased FA in the fornix (Reynolds et al., 2019), suggesting that fornix microstructure is still developing in these very young children (Fig. 3). However, no relationship between age and fornix FA was found in studies of children in whom the youngest participants were 4 years old (Ngo et al., 2017), 5.6 years old (Lebel et al., 2008), and 7 years old (Wendelken et al., 2015), which suggests that fornix microstructure reaches maturity before age 4.
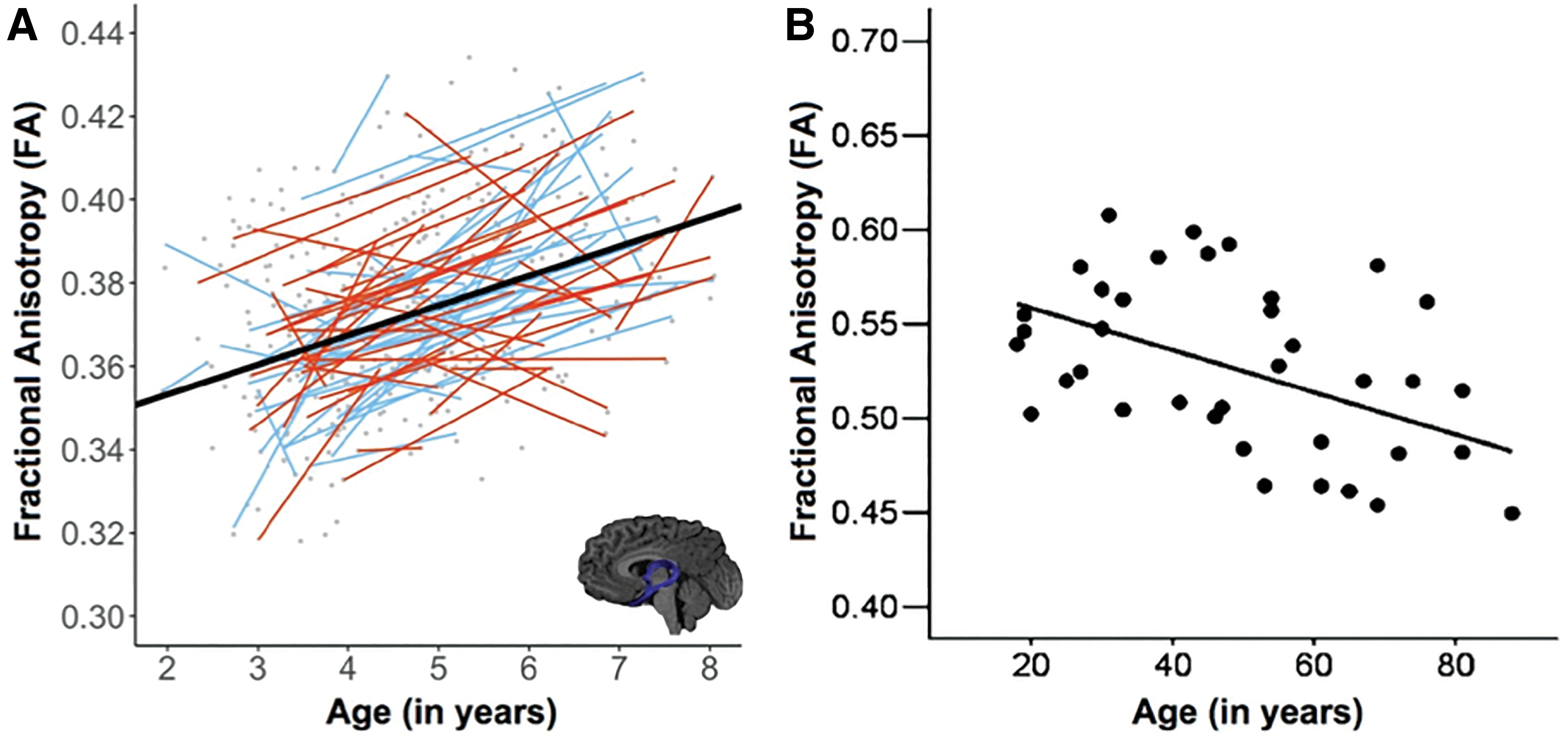
Fornix microstructure in development and aging.
Relationships between age and fornix MD during development are conflicting, with some evidence suggesting decreased MD in the fornix in children with increasing age (Lebel et al., 2008), whereas other studies show no relationship between age and MD (Reynolds et al., 2019). This discrepancy could be due to the different age ranges and developmental stages of children included in these studies. Wendelken and colleagues (2015) also found a relationship between fornix FA and memory performance in adults, but not in children. This finding is surprising but potentially explained by issues surrounding imaging of the fornix (Box 1).
DWI of the fornix in adult episodic memory
The small diffusion imaging literature in healthy young adults has consistently reported that lower fornix FA and/or higher MD correlates with worse episodic memory performance on scene memory and virtual navigation tasks (Hodgetts et al., 2015, 2020; Rudebeck et al., 2009) and worse recall of autobiographical memories (Hodgetts et al., 2017) (Table 2). In some studies, only the hippocampal portion of the fornix—the fimbria—was examined.
Empirical Papers Using Diffusion-Weighted Imaging Methods that Reported Effects in the Fornix
Papers are grouped by population(s) tested, then listed alphabetically. Disorders for which there were less than three studies are listed under the subheading “Other.” We identified relevant publications by searching PubMed with the following combinations of search terms: (1) fornix and memory and diffusion; (2) fornix and TBSS; (3) fornix and tractography; and (4) fornix and structural connectivity. Publications that were reviews, methods papers, written in a language other than English, or used a neonatal sample were excluded.
The primary analysis method is listed for each paper, as well as the white matter indices are evaluated. The type of episodic memory assessed is listed, if applicable. An asterisk (*) denotes studies in which episodic memory was significantly associated with one or more diffusion metrics.
AxD, axial diffusivity; ADC, apparent diffusion coefficient; FA, fractional anisotropy; MD, mean diffusivity; RD, radial diffusivity; ROI, region of interest; TBI, traumatic brain injury; TBSS, tract-based spatial statistics.
For instance, Wendelken et al. (2015) had young adult participants attend to some scenes but not others; then, they later tested old/new recognition and confidence ratings. They reported a correlation between the fimbria and memory performance. In another study, participants were required to remember the location of a particular face; performance on this task correlated with FA in the fimbria (Schwarb et al., 2019). One study that examined participants aged 18–88 found decreases in performance on item and associative memory tasks across the lifespan, as well as decreased FA in the fornix across the lifespan that accelerated in old age (Henson et al., 2016).
A larger literature has accrued in healthy older adults. There is a reliable relationship between chronological age and decreased FA and/or increased MD in the fornix, indicating that the fornix is generally compromised in older adults (Bennett et al., 2015; Bennett and Stark, 2016; Gunbey et al., 2014; Henson et al., 2016; Jang et al., 2011; Metzler-Baddeley et al., 2011; Metzler-Baddeley et al., 2011; Pelletier et al., 2013; Stadlbauer et al., 2008; Zahr et al., 2009; Zhuang et al., 2012, 2013) (Fig. 3).
Importantly, most studies that have tested memory have found that reduced episodic memory performance in older adults correlates with decreased fornix microstructure across a range of tasks and procedures; for instance, object-location relational tasks (Antonenko et al., 2016), “pattern separation” tasks (old-new-similar object recognition task, word list recall) (Bennett et al., 2015), the mnemonic similarity task (which is an old-new-similar object recognition task) (Bennett and Stark, 2016), verbal and visual recognition as measured by the Mayo Clinic neuropsychological task battery (Kantarci et al., 2014), verbal and visual recall (Doors and People, Free and Cued Selective Reminding Test) (Metzler-Baddeley et al., 2011; Metzler-Baddeley et al., 2011; Ray et al., 2015), and delayed verbal recall (Logical Memory delayed recall, the Rey Auditory Verbal Learning Test) (Zhuang et al., 2013).
Increases in MD have also been linked to decreases in episodic memory performance in older adults in some of the studies cited earlier (Antonenko et al., 2016; Bennett et al., 2015; Zhuang et al., 2013). There are also reduced fornix fiber counts found with increasing age (Stadlbauer et al., 2008). However, a few investigators (all using tract-based spatial statistics [TBSS] methods) did not find a relationship between FA/MD and episodic memory (Table 2).
Summary of findings on episodic memory
Findings from the reviewed studies consistently and overwhelmingly converge on the idea that the fornix is essential for memory tasks that are traditionally thought of as “hippocampal.” Fornix damage in rodents or monkeys causes deficits across a range of tasks, including trace conditioning, contextual fear conditioning, spatial navigation, object-location pairing, and other visuospatial tasks. Moreover, fornix lesions in nonhuman primates lead to deficits that mirror the temporality of amnesia observed in humans suffering from bilateral hippocampal lesions—anterograde, but limited retrograde, amnesia.
After transection of the fornix, primates demonstrate accurate memory for preoperative learning in many studies, but they show an inability to learn new tasks postoperatively (Buckley et al., 2004, 2008; Gaffan et al., 2001). In humans, fornix damage causes anterograde amnesia that is worse for recall as compared with recognition. The clinical profile is more similar to medial temporal lobe (MTL) amnesia than it is to diencephalic amnesia, due to the absence of confabulation. Are the deficits after fornix lesions of the same duration and of the same magnitude as that observed after hippocampal lesions in nonhuman animals? Most studies suggest that the deficits are virtually identical (e.g., Aggleton et al., 1992; Maren and Fanselow, 1997).
Recent findings using noninvasive methods to examine the fornix such as DWI converge on a similar narrative, showing a correlation between diffusion metrics and episodic memory performance across a range of age groups and tasks. The tasks range from well-validated standardized neuropsychological tests of memory such as Doors and People (Baddeley et al., 1994), to novel laboratory tasks that require remembering lists of words or similar-looking objects. What is not known is whether the fornix is involved in a broader array of tasks, since most studies only examine episodic memory. In addition, plasticity of the fornix is relatively unexplored but of great importance (see Plasticity section), given that fornix microstructure degrades with age (Fig. 3).
The DWI methods have greatly improved since the inception of this technique more than 20 years ago. Therefore, older studies, which use only a small number of gradient directions and rarely control for the effects of CSF on diffusion metrics, are likely less reliable than those conducted more recently. Further studies employing state-of-the art diffusion imaging techniques are recommended to get more reliable and specific measures of diffusion properties in the fornix going forward (Table 2 and Box 1).
Is the role of the fornix in humans exclusive to spatial memory?
A putative role for the fornix in cognition is that it plays a key role in spatial memory (e.g., navigation) and spatial perception, with decades of evidence from the animal literature supporting this theory (see Lesions of the Fornix in Nonhuman Animals section). This view has led to a controversial model that the hippocampus plays a material-specific role in cognition: that it is essential for perceiving and remembering scenes and locations; whereas another MTL region—the perirhinal cortex—plays an essential role in face/object perception (reviewed in Graham et al., 2010). Because the fornix innervates the hippocampus, but not perirhinal cortex, these findings would suggest that fornix transection could cause spatial memory and perception deficits, but not object/face memory and perception deficits.
Supporting this view, the literature shows that in nonhuman primates, memory for objects and faces is usually spared, whereas spatial memory is impaired, after fornix transection (Browning et al., 2010; Bussey et al., 2000; Ennaceur and Aggleton, 1997; Ennaceur et al., 1996; Gaffan, 1992; Kwok and Buckley, 2009, 2010; Markowska et al., 1989; Wilson et al., 2007; but see Bachevalier et al., 1985; Gaffan, 1994). In addition, Graham and colleagues used DWI in young healthy participants and found that higher fornix FA correlated with better performance on spatial memory measures but did not correlate with performance on object memory measures (Hodgetts et al., 2015, 2017; Rudebeck et al., 2009).
As compelling as these findings are, it must be remembered that humans differ from rats and monkeys in several small ways, but also in one major way: We have language. Language, much like flight, is a species game-changer. Although the only way to tap into “episodic-like” memory in nonhuman animals is through spatial tasks, tasks such as verbal recall are regularly used in human studies as an index of episodic memory—because humans have language, we can simply ask study participants what they remember. The ability to remember someone's name, to communicate instructions, or to follow the narrative of a story is essential for human interactions. Humans with fornix damage fail to remember words and conversations, consistent with global amnesia (Calabrese et al., 1995; D'Esposito et al., 1995). A quick look at Table 1 shows that nearly every study of individuals with fornix damage reported verbal memory impairments. Thus, evidence from humans does not support the claim that the fornix is solely involved in spatial memory.
As for the claim regarding the role of the fornix in visual perception (as opposed to memory), there is evidence from nonhuman primates that fornix transection impairs the ability to discriminate between pictures of complex scenes, while leaving the ability to discriminate between pictures of objects intact (Browning et al., 2010; Gaffan, 1992, 1994; Easton et al., 2002; Wilson et al., 2008). The tiny literature from humans with fornix lesions is mixed (D'Esposito et al., 1995; Gaffan et al., 1991), whereas the only DWI study found evidence for a material-specific role such that fornix microstructure correlated with the perception of scenes (Postans et al., 2014).
Working memory, curiosity, and other aspects of cognition
Some researchers have conducted diffusion imaging studies on healthy participants and found associations between working memory performance and higher FA (Zahr et al., 2009) and lower longitudinal and transverse diffusivity in the fornix (measures similar to MD) (Sullivan et al., 2010). Higher fornix FA is related to better performance on measures of working memory and motor control among older adults (Zahr et al., 2009). However, studies of patients with fornix lesions find that they have typical working memory capacities (Calabrese et al., 1995; Gaffan et al., 1991; Hodges and Carpenter, 1991; Park et al., 2000; Poreh et al., 2006; Tsivilis et al., 2008). One case report detailed a patient with deficits in working memory, but this patient sustained a gunshot wound, and the authors speculate that her working memory deficit was a result of damage by the bullet to the left posterior parietal lobe (D'Esposito et al., 1995). In sum, evidence for the fornix playing an essential role in working memory is weak.
Recently, a DWI study linked variation in fornix microstructure to the traits of novelty seeking and curiosity (Valji et al., 2019). These findings make sense in light of a large literature showing that the hippocampus is extremely sensitive to novelty, containing a comparator mechanism that allows incoming information to be compared with stored representations (Lisman and Grace, 2005).
Summary
To summarize, the fornix is critical for all forms of episodic memory, not just spatial memory. In addition, the literature linking the fornix to spatial perception is small and findings are mixed, and these are potentially confounded by demands placed on transsaccadic memory [e.g., the limited capacity visual memory system that knits together the visual world over multiple saccades (Irwin and Gordon, 1998)]. Future research should investigate whether transsaccadic memory as well as other forms of working and short-term memory rely on the fornix. Other aspects of cognition—reasoning, attention, language, mathematical abilities, and conceptual knowledge, for instance—do not appear to rely on the fornix, although, admittedly, the literature on this is almost nonexistent.
Mechanistic Role of the Fornix
The reviewed empirical literature strongly indicates that the fornix plays an essential role in episodic forms of memory. Here we ask, what is the mechanistic role of the fornix? Rigorous investigations in rodents have identified several types of information transmitted through the fornix, which are reviewed next.
Transmission from basal forebrain to the hippocampus: acetylcholine and theta
A key role of the fornix is to transmit an essential neurotransmitter—acetylcholine—from the basal forebrain to the hippocampus. Cells in the medial septum (a region in the basal forebrain, which, in turn, receives input from the hypothalamus and several brainstem areas) (Khakpai et al., 2013) are the brain's primary source for acetylcholine, which is transmitted to the hippocampus via the fornix. Thus, damage or deterioration of the fornix diminishes acetylcholine delivery to the hippocampus (Blaker et al., 1988; Rawlins et al., 1979). We know that acetylcholine plays an essential role in memory based on decades of findings showing, for instance, that depletion of basal forebrain acetylcholine precedes AD (Muir, 1997), that blocking acetylcholine in rodents using scopolamine inhibits episodic memory formation (Dixon et al., 1996), and that supplementing acetylcholine can improve episodic memory (Oswald et al., 2007).
Mechanistically, acetylcholine upregulates memory encoding while simultaneously downregulating memory retrieval/consolidation (Hasselmo, 1999). In other words, acetylcholine acts like a switch, shifting the function of hippocampal circuits from an internal, auto-associative phase to an externally oriented encoding phase. This helps to explain why older adults, who commonly have diminished cholinergic tone, show a “retrieval bias” on memory tasks (Wilson et al., 2006). We note that these findings predict that altered fornix macro- or microstructure, regardless of age, should correlate with poor cholinergic tone, and relatively poor memory encoding, especially on pattern separation tasks. It is, thus, possible that the observed memory impairments after fornix damage can be completely explained by harkening to essential neurotransmitter depletion. However, there is an orthogonal explanation regarding theta rhythms.
Theta rhythms are found throughout the brain and dominate the local field potential in the hippocampus. There is mounting evidence that hippocampal theta rhythms (4–12 Hz) play an important role in episodic memory (e.g., Hsieh and Ranganath, 2014) and spatial navigation (Buzsáki, 2005). Theta power at encoding correlates with subsequent word recall (Klimesch et al., 1996) and memory for word-video associations (Staudigl and Hanslmayr, 2013). Further, greater theta band coherence compared with baseline for MTL-PFC interaction has been found during memory recall (Anderson et al., 2010).
Theta may also play some role in memory binding—remembering the co-occurrence of multiple elements within an event (Buzsáki and Draguhn, 2004; Hanslmayr et al., 2016). Corroborating this view, several studies have found that hippocampal theta-gamma coupling is linked to item-context binding processes (Backus et al., 2016; Lega et al., 2012; Staudigl and Hanslmayr, 2013; Tort et al., 2009). The GABAergic neurons in the medial septum serve as a generator and pacemaker for theta. These cells burst at theta frequencies in vivo, and destruction of the medial septum halts whole-brain theta rhythms, whereas destruction of the fornix halts hippocampal theta rhythms (Rawlins et al., 1979), resulting in episodic memory impairments (Cassel, 1997; Lipponen et al., 2012). There is also evidence that acetylcholine modulates theta power (Zhang et al., 2010).
Transmission from the hippocampus to the basal forebrain
The mechanisms we just described only explain one direction of fornix information flow; they do not explain how hippocampal outflow via the fornix modulates subcortical structures. The literature relevant to this question is surprisingly small. However, a few recent studies suggest that this flow of information may be essential for modulating motivated behavior. For instance, a recent study reported that the hippocampus regulates social attack via a circuit from CA2→lateral septum→ventromedial hypothalamus. It is possible that a decision to engage in social attack requires retrieval of memories of past social encounters that allow a prediction to be made about the possible win–lose outcome of aggressive behavior (Pagani et al., 2015). Using similar logic, we speculate that the hippocampus, via the fornix, also regulates the decision to engage in sexual behavior, based on older findings showing that direct electrical stimulation of the human septum elicited subjective pleasure and sexual arousal (Heath, 1964, 1972; Olds and Milner, 1954)
Other studies have reported that a circuit from CA3→lateral septum modulates food intake by regulating learned or emotional aspects of food intake, such as how much time has elapsed since the last meal (Sweeney and Yang, 2015). This finding harkens to studies showing that individuals with anterograde amnesia are unable to recall recent eating and will consume multiple meals in quick succession (Hebben et al., 1985; Higgs et al., 2008). It also helps to explain new DWI findings linking fornix microstructure to body mass index (BMI) (Metzler-Baddeley et al., 2013).
The fornix also provides a direct connection between the hippocampus and the nucleus accumbens and there is evidence that this circuit allows memories of reward-predicting environments to guide where animals look for food (Trouche et al., 2019). Taken together, these findings suggest that the fornix transmits mnemonic information that shapes and guides motivated behaviors. Evidence about this role is only beginning to emerge in the literature. It is entirely possible that the third role is broader than our description; the fornix may transmit mnemonic information that guides decision making in all shapes and forms.
Summary
The reviewed findings suggest three mechanistic functions of the fornix in episodic memory. One role is in transmitting acetylcholine, which switches the hippocampus from a baseline consolidation state to an encoding state. A second potential role is in transmitting theta, which may serve a binding role in episodic memory. A third role is in transmitting mnemonic information that shapes decisions about where to forage, when to feed, and who to fight. Although we have highlighted these three functions, it is entirely possible that the fornix is involved in the transmission of mnemonic information that guides reasoning, creativity, and other aspects of human cognition that are shaped by our experience.
The reviewed findings and mechanisms allow us to conjecture which brain disorders might be linked to fornix dysfunction. Disorders in which there is a prominent episodic memory component such as MCI and AD should show fornix pathology. In addition, there are disorders with subtle, but consistent episodic memory impairments, such as schizophrenia and major depression, which may involve the fornix (along with other white matter tracts). Last, disorders linked to aberrant reward-seeking behavior, such as obesity or substance abuse, may involve the fornix. These disorders are reviewed next. Studies are listed in Table 2.
The Involvement of the Fornix in Neuropsychiatric Disorders
The DWI literature linking the fornix to various clinical disorders has been reviewed in some detail recently by Douet and Chang (2015). These authors reviewed studies linking fornix changes to Parkinson's disease, multiple sclerosis, frontotemporal dementia, and MTL epilepsy (see Douet and Chang, 2015). Other authors have reported changes in the fornix in rare disorders such as idiopathic normal pressure hydrocephalus (Hattori et al., 2011; Oishi and Lyketsos, 2014). Most of these literatures are very small, and findings tend to be inconsistent. In the next sections, we review larger literatures linking changes in the fornix to MCI, AD, schizophrenia and psychosis, mood disorders, and eating disorders.
The fornix in MCI and AD
The “MCI” is a term used to conceptualize the cognitive abilities of older adults whose mental faculties are compromised beyond what would be expected in normal aging. It can be divided into two subtypes: nonamnestic (naMCI) and amnestic (aMCI), with the latter including a memory impairment that exceeds what would be expected in typical aging, but that does not reach the level of dementia (reviewed in Yu et al., 2017a). The DWI is particularly useful for examining changes in the early stages of cognitive impairment, as it is sensitive to subtle white matter alterations that would not be evident by using traditional MRI (Moseley, 2002). Of particular relevance to pathology of the fornix is the amnestic variant of MCI, as the memory impairments specific to aMCI should be related to fornix microstructural changes.
Individuals with aMCI have relatively decreased FA in the fornix compared with healthy, age-matched controls (Bozoki et al., 2012; Kantarci et al., 2014; Rémy et al., 2015; Zhuang et al., 2012, 2013). Changes in fornix FA or MD appear to precede conversion to AD, making this a potent biomarker for pathological memory profile (Fletcher et al., 2013, 2014; Yu et al., 2017a; Zhuang et al., 2013), especially since around 40 percent of patients with aMCI will eventually convert to AD (reviewed in Douet and Chang, 2015). It has even been proposed that the “fornix sign,” a discoloration of the fornix in color-coded FA maps of the fornix, can be used to predict conversion to AD (Oishi et al., 2012) (Fig. 4). Several findings indicate that white matter alterations in limbic tracts may precede gray matter degeneration in the hippocampus as well as secondary loss of white matter volume due to neuronal loss (Fletcher et al., 2013; Zhuang et al., 2013). Some researchers have suggested that using models that incorporate both gray matter volume changes and white matter microstructural changes together are optimal for detecting the early signs of MCI (Douaud et al., 2013). Importantly, in a study of older adults with MCI, including fornical crus FA in a logistic regression improved the ability of the model to distinguish aMCI from naMCI participants (Gyebnár et al., 2018). These findings underscore the importance of fornix microstructure in understanding the degradation of memory capacity in senescence.
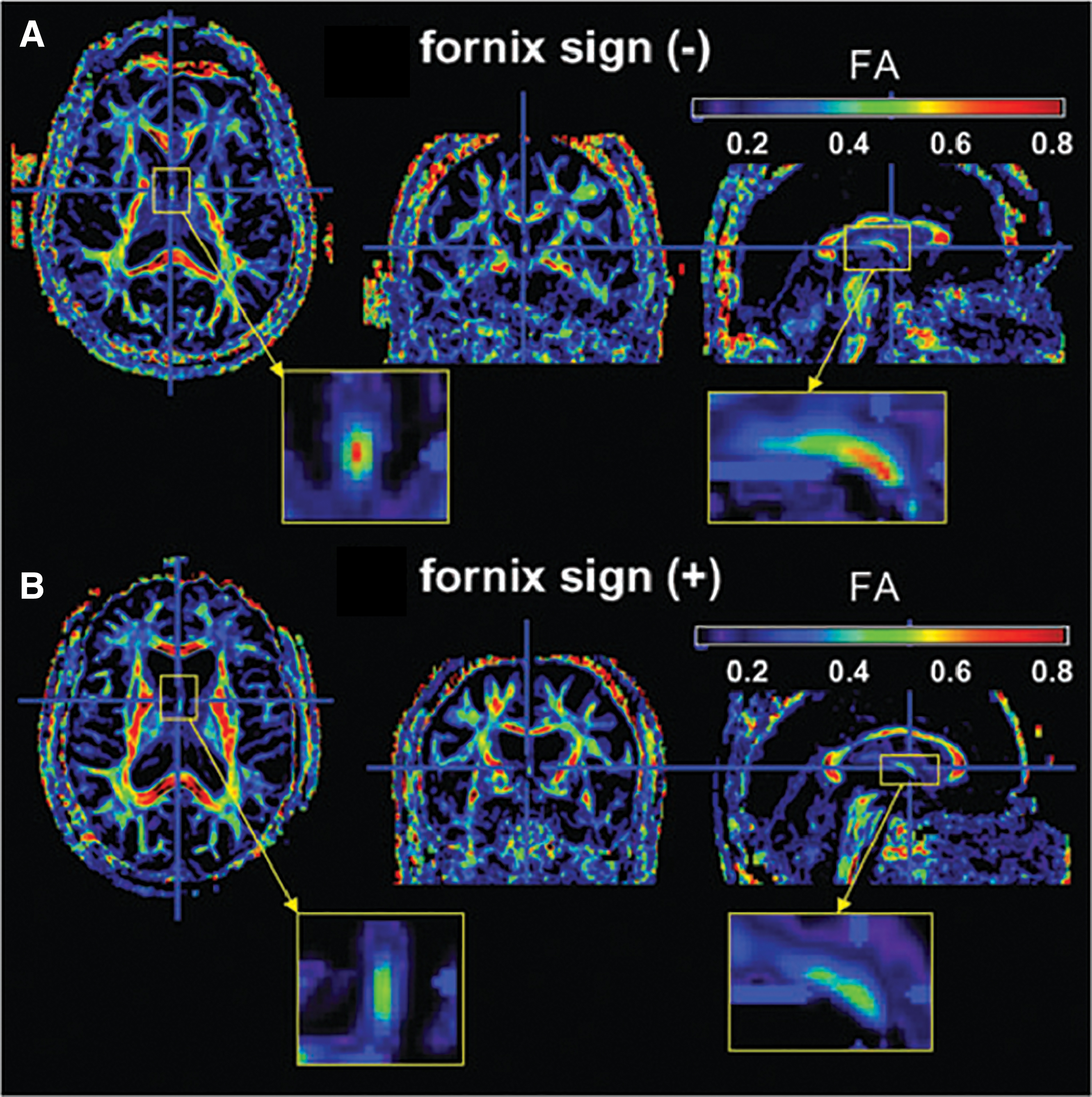
The “fornix sign.” The colors on FA maps in people with white matter degeneration indicate decreased FA in the fornix, one of the early signs of cognitive decline.
A meta-analysis by Yu et al. (2017a) indicated that the white matter tracts most consistently linked to aMCI were the three major limbic pathways: the fornix, uncinate, and parahippocampal portions of the cingulum bundle. Further, their region of interest (ROI)-based analyses showed a consistent relationship between aMCI and fornix FA, but not MD. The authors note that the inconsistency of free water correction across studies not only might explain the heterogeneity of results for MD and a lack of significant findings for this metric, but also might lead to an overestimation of the association between fornix FA and aMCI pathology (Yu et al., 2017a).
Indeed, other studies have found the opposite pattern—that increased fornix MD is more strongly associated with MCI pathology than is FA (Nir et al., 2013; Nowrangi et al., 2013; Tang et al., 2017), whereas still another found that both fornix FA and MD are effective individual predictors of longitudinal cognitive decline in MCI patients (Mielke et al., 2012). It is worth noting, however, that many of these studies do not distinguish between aMCI and naMCI, which may affect the results, as individuals with naMCI do not always have the memory impairments associated with AD and fornix degradation.
Although this review is focused on the fornix, it is worth mentioning that it is not the only tract implicated in mnemonic impairment and cognitive decline. The literature on MCI indicates that changes in the microstructure of the fornix as well as the cingulum bundle can be important predictors of memory impairment and of eventual conversion to AD (Bozoki et al., 2012; Gyebnár et al., 2018; Nir et al., 2013; Nowrangi et al., 2013; Yu et al., 2017a). Future diffusion imaging studies should focus on tracking older adults longitudinally to better elucidate which diffusion measures are most sensitive to white matter changes and, as such, are the best predictors of cognitive decline.
The AD is characterized by the presence of beta-amyloid plaques and neurofibrillary tangles in the brain that often appear first in the MTL before spreading to the cortex. An early symptom is significant decline in episodic memory performance, and hippocampal atrophy is commonly linked to this condition (Apostolova et al., 2012). Widespread white matter changes are observed in AD, but changes in the fornix are especially interesting since such changes accord with essential limbic pathology and are consistently observed across different samples and methodologies (Oishi and Lyketsos, 2014).
Fornix pathology will necessarily impair cholinergic transmission to the hippocampus. Indeed, it has been known for decades that AD is accompanied by loss of this transmission (Coyle et al., 1983). Similar to MCI, AD pathology is associated with decreased FA/increased MD in the fornix (Agosta et al., 2011; Bozoki et al., 2012; Kantarci et al., 2017; Mayo et al., 2016; Mielke et al., 2012; Nir et al., 2013; Nowrangi et al., 2013; Tang et al., 2017). There is some inconsistency in whether any given study finds a change in FA or MD; however, the fornix is consistently implicated, regardless of the diffusion parameter showing changes. In addition, fornix diffusivity metrics are consistently linked to worse performance on memory tasks for patients with both MCI and AD (reviewed in Douet and Chang, 2015), highlighting the specific connection between fornix pathology and memory impairment.
One of the most important findings regarding the fornix is that white matter abnormalities may precede the gray matter degradation in the MTL that is typical of Alzheimer's pathology (Agosta et al., 2011; Fletcher et al., 2013, 2014; Zhuang et al., 2013; reviewed in Douet and Chang, 2015), and they are often present in patients with aMCI before they convert to AD. This has led to the controversial suggestion that early white matter disruption (especially in the fornix) triggers an adaptive response of deposition of amyloid and tau along with deficits in cholinergic transmission (Bartzokis, 2011). The change in cholinergic transmission leads to the behavioral manifestation—memory problems—whereas the accumulation of amyloid and tau potentially causes the secondary deterioration of hippocampal gray matter. This would explain why traumatic brain injury and cerebro-vascular disease, which are both associated with diffuse axonal injury and damage to myelin, are also strong risk factors for AD (Filley and Fields, 2016).
Clinicians are interested in predicting AD in vivo, so tracking changes in white matter diffusion properties could be an effective method for projecting future amnestic symptoms and cognitive decline in older adults. Volume loss and microstructural changes in the fornix have been found to be predictive of conversion from normal cognition to MCI and AD (Fletcher et al., 2014). In addition, there is some evidence that acetylcholinesterase inhibitors, which are commonly prescribed for the treatment of AD, can have a positive effect on myelination (Bartzokis, 2007).
In sum, there is strong evidence that fornix pathology is tightly associated with aMCI and the development of AD. Tracking fornix pathology by using diffusion imaging can be an effective way to predict cognitive decline and memory impairment in older adults at risk for developing AD.
The fornix in schizophrenia, psychosis, and mood disorders
The diffusion imaging literature on schizophrenia is quite large due to long-standing theories about disrupted circuitry in this disorder (Andreasen et al., 1998). In addition, schizophrenia is often accompanied by widespread cognitive alterations, including changes in relational memory (Jung and Lee, 2016), which provides a theoretical justification for looking at the fornix in this disorder. Some studies have reported changes in fornix microstructure in schizophrenia, whereas others have not (reviewed by Douet and Chang, 2015; see also Baumann et al., 2016). However, postmortem studies of individuals with schizophrenia have not found that the fornix is altered (reviewed by Douet and Chang, 2015) and case reports on humans with fornix damage do not report that they suffer from any of the cardinal position or negative symptoms of schizophrenia.
Explanations for inconsistent findings include enlarged ventricles in individuals with schizophrenia (Sayo et al., 2012), which can bias the diffusion metrics in the fornix (see Box 1), and heterogeneous etiologies in the sample population. Nevertheless, this remains an interesting tract to target in this population, by using optimized imaging methods.
A stronger case can be made for the fornix in mood disorders. It has been hypothesized that the fornix plays an important role in the mediation of fear and anxiety (Degroot and Treit, 2004) as well as daily stress, as the conduit of glucocorticoids to the hippocampus (Savransky et al., 2017). Alterations to the fornix microstructure may cause affective dysregulation, which is observed in many mood disorders. For example, variations in the microstructural properties of the fornix body have been associated with early trauma and with symptom severity in individuals with panic disorder (Yu et al., 2017b).
Adolescents with bipolar disorder had lower fornix FA compared with age-matched controls, suggesting that alterations in this white matter pathway are present early in the course of the disease (Barnea-Goraly et al., 2009). Another study reported that unmedicated patients with bipolar disorder had reduced fimbria volume compared with medicated patients and controls (Elvsåshagen et al., 2012). A recent pilot study reported that deep brain stimulation (DBS) of the fornix improved mood in individuals with AD (Mao et al., 2018). These results corroborate findings in healthy controls, showing a correlation between individual differences in sub-clinical anxiety and fornix microstructure (Modi et al., 2013).
Although the findings we described are a mixed lot of clinical disorders and methods (Table 2), they hint that the fornix may play a role in affect regulation, which would not be surprising since the subcortical nuclei at the anterior end of the fornix are involved in many essential limbic functions, including reward signaling. Future research should examine whether the fornix is demyelinated or atrophied in stress-related disorders such as post-traumatic stress disorder.
The fornix in aberrant reward-seeking: eating disorders and substance abuse
Decisions about when to eat and where to optimally forage for food rely on long-term memories (see Transmission from the Hippocampus to the Basal Forebrain section). Thus, it is possible that alterations in fornix microstructure account for pathology in eating behavior. Some studies report that in humans, variation in fornix microstructure correlates with BMI (Metzler-Baddeley et al., 2013; Xu et al., 2013; reviewed in Kullmann et al., 2015). These studies generally show that higher BMI is associated with lower FA, higher MD, or both. In addition to degraded fornix microstructure in those who are overweight or obese, there is a similar pattern in those with eating disorders of food restriction, such as anorexia nervosa and bulimia nervosa. Several studies have investigated this relationship and found decreased FA values, increased MD values, or both in participants with anorexia nervosa (Frank et al., 2013; Kazlouski et al., 2011; Monzon et al., 2016; Phillipou et al., 2018) and bulimia nervosa (Kazlouski et al., 2011; Mettler et al., 2013).
This pattern across participants who are both over- and underweight suggests that the fornix may be integral in circuitry related to eating behaviors. However, many of these studies found microstructural changes in tracts other than the fornix, indicating that very high or low BMI (or perhaps frequent vacillations in body weight related to disordered eating patterns) may lead to widespread white matter changes across the brain that are not constrained to the fornix. This is complemented by findings that indicate no group differences in white matter microstructure between women who have recovered from anorexia nervosa and normal controls (Bang et al., 2018), suggesting that the white matter abnormalities found in patients with anorexia nervosa may be a result of eating disorder pathology rather than a cause.
One review suggests that such white matter degradation in the brain is related to increased inflammation in the body due to poor physical health, which can, in turn, impact neurons' myelin sheaths (Porter et al., 2018). More work is needed to elucidate whether there is a particular role of the fornix in eating behaviors or if, instead, having a BMI outside what is considered the normal range is related to nonspecific white matter degradation across the brain. Since abnormal eating behaviors are often related to restriction, binging, or both, there is a component of cognitive control and reward seeking inherently tied to disordered eating. Perhaps it is these behaviors that are tied to the altered fornix microstructure in people who are over- or underweight.
Substance abuse is a classic model of reward seeking gone awry. We recently reviewed the diffusion imaging literature on substance abuse (Hampton et al., 2019) and found that the literature was highly inconsistent, even when restricted to a particular substance such as cocaine. The fornix was mentioned as being affected in only a single study of adolescents with alcohol use disorder (Cardenas et al., 2013). The absence of findings should not be weighted too heavily, as this literature suffers from many methodological problems such as imaging methods with poor signal-to-noise ratios, reliance on whole-brain TBSS analyses, and absence of hypotheses that might point toward a specific tract or brain region. Future research should closely examine the fornix, especially in substance use disorders where pleasurable memories may be especially important in driving behavior (e.g., opioids).
At the Cutting Edge: DBS of the Fornix
There is currently great interest in electrically stimulating the fornix to potentially improve memory in MCI and AD. This idea has its genesis in a now-classic study by Hamani and colleagues, in which the goal was to use DBS of the hypothalamus to help with weight control in a patient who was morbidly obese (Hamani et al., 2008). The DBS functions therapeutically in humans at frequencies between 80 and 185 Hz and with a stimulation current between 1 and 10 mA (Vedam-Mai et al., 2012). The effects of DBS on neuronal function can be excitatory or inhibitory, depending on the stimulation site. In Hamani and colleagues' study (2008), they inadvertently stimulated the fornix (due to the proximity of the fornix to the hypothalamus), which caused the patient to recount extremely detailed autobiographical memories during the surgery, as well as leading to increased activity in the hippocampus and parahippocampal cortex. The patient continued to demonstrate increased performance on verbal recognition and spatial memory assessments after 3 weeks of continuous stimulation (Hamani et al., 2008).
This compelling finding helped launch a small industry of fornix DBS studies. Studies in rats showed that DBS of the fornix can activate the hippocampus (Gondard et al., 2015) and can reset hippocampal theta, promoting optimal memory encoding (Williams and Givens, 2003). The DBS of the fornix in rats also improves performance on hippocampal-dependent spatial memory tasks (Hescham et al., 2013; Zhang et al., 2015). In addition, it can reverse Alzheimer's-type memory impairments in these rats (Hescham et al., 2013), but it does not appear to have an effect on the presence of Alzheimer's-related proteins such as beta-amyloid or tau (Gondard et al., 2015).
The benefits to memory in rats with impairments mimicking Alzheimer's deficits raise the question of whether this would translate to humans. Several studies have revealed optimistic findings: Fornix DBS in humans can drive neural activity in the entorhinal cortex and hippocampus to reverse impaired glucose utilization in the temporal and parietal lobes (Laxton et al., 2010) and increase metabolism in the MTLs (Fontaine et al., 2013). In addition, patients with AD who were receiving DBS to the fornix showed slowed hippocampal atrophy relative to AD controls who did not receive stimulation (Sankar et al., 2015).
Case studies have also shown that patients whose memory was declining before fornix stimulation stabilized after DBS (Fontaine et al., 2013; Laxton et al., 2010), and that even patients with severe AD showed some improvements in mood, memory, and language ability, as reported by caregivers (Mao et al., 2018). Intriguingly, a recent report described how 48% of AD patients in a clinical trial who received DBS to the fornix experienced vivid flashbacks. Most of these flashbacks had elaborate spatiotemporal details and the details and clarity of the recollection increased as the voltage was increased (Deeb et al., 2019).
In general, the memory improvements in DBS studies are modest, and it has been proposed that more robust results would be obtained if studies employed closed-loop stimulation rather than open-loop stimulation, as closed-loop formats allow stimulation to optimally target memory-related activity in the brain (Senova et al., 2018). Open-loop stimulation refers to the most common type of DBS in which a constant stimulation, at a constant frequency, is applied. In closed-loop protocols, stimulation is done in targeted intervals at specific frequencies that align with brain activity occurring during either encoding or retrieval periods. Another possibility that can be explored is to use rhythmic DBS of the fornix to pace or re-pace potentially aberrant hippocampal theta rhythms.
Although the benefits of DBS of the fornix on memory in MCI and AD remain inconclusive, there is some evidence that fornix DBS may have an unexpected benefit for epilepsy. Koubeissi and colleagues (2013) found modest benefits of fornix stimulation on memory recall in epilepsy. More robust was the 92 percent decrease in seizure occurrence, suggesting clinical benefits of fornix stimulation beyond memory improvements (Koubeissi et al., 2013).
Plasticity
Only a few studies have looked at plasticity in the fornix, but those that have done so suggest that interventions may have an effect on fornix microstructure. There is some evidence, in both humans and rats, that only 2 h of training gives rise to measurable microstructure changes in the adult fornix (Hofstetter et al., 2013). In one study, a group of healthy older adults trained on an object-location task designed to index episodic memory. The FA increased and MD decreased in the fornix from pre- to post-training, and increased FA across training was related to better recall—but not recognition—performance at post-test (Antonenko et al., 2016).
A range of environmental factors can also have longlasting effects on episodic memory. Chronic sleep disturbances, long-term use of anti-cholinergic medications (e.g., Benadryl), and hypoxia (while mountain climbing, deep sea diving, or even from being on a ventilator) can all impair episodic memory. In contrast, socializing and aerobic exercise improve episodic memory, especially in aged populations (Blondell et al., 2014; James et al., 2011; but see also Sabia et al., 2017). Burzynska and colleagues (2017) found that older adults randomized to participate in a dance intervention (as compared with walking, walking and nutrition, and active control groups) showed increased FA and decreased MD and RD in the fornix, whereas the other groups showed the opposite pattern across the intervention period. This improvement in the dance intervention group was specific to the fornix—in other brain regions; WM indices reflected patterns of decline regardless of group. These results suggest that for older adults, participating in physical and social activities may help to alleviate the detrimental effects of aging on the fornix. Future studies should assess the longevity of these effects, whether they extend to other sample populations, and whether microstructural changes in white matter and gray matter happen in conjunction.
Conclusions
Our review of the fornix literature reveals that this white matter tract plays an essential role in episodic forms of memory across species and tasks. We were surprised to find that the effects of fornix damage on memory are nearly indistinguishable from the deficits observed after focal hippocampal lesions. Classic linkages and functional divisions in the literature on the hippocampus such as episodic versus semantic memory, recall versus recognition, and explicit versus implicit are mimicked in the literature on the fornix, supporting an older idea that damage to any part of the “extended hippocampal memory system” would cause similar memory deficits (e.g., Aggleton and Brown, 1999; Warrington and Weiskrantz, 1982).
The mechanistic role of the fornix is, of course, quite different than the computational role of the hippocampus in episodic memory. The fornix is the conduit by which the neurotransmitter acetylcholine—which is crucial for memory encoding—is sent from the medial septum/diagonal band of Broca to the hippocampus. In addition, the GABAergic neurons in the septal nuclei generate theta rhythms that are transmitted through the fornix to the hippocampus. In the absence of these external modulators, the hippocampus is radically dysfunctional.
Without an intact fornix, hippocampal neurons do not have a direct pathway to the basal forebrain or diencephalon, although it retains intact communication with MTL regions such as the entorhinal cortex, amygdala, and retrosplenial cortex (Aggleton et al., 2015). It has been suggested that fornix pathology in MCI and AD should lead to a greater reliance on alternative MTL pathways, perhaps via the cingulum bundle (Metzler-Baddeley et al., 2012) or uncinate fasciculus. The uncinate fasciculus clearly plays a role in learning and memory (Alm et al., 2015, 2016; Von Der Heide et al., 2013), whereas the cingulum bundle does not appear to play an essential role in episodic memory (Bubb et al., 2018).
The findings in the lesion literature are mirrored in the newer diffusion imaging literature. Microstructural properties of the fornix, especially FA, decrease with advanced age, correlate with age-related memory impairments, and decrease in disease states defined clinically by episodic memory problems (aMCI and AD). In fact, it has been suggested that fornix pathology precedes gray matter degeneration in the hippocampus as well as secondary loss of white matter volume due to neuronal loss (Fletcher et al., 2013; Zhuang et al., 2013). The “fornix sign,” a discoloration of the fornix in color-coded FA maps of the fornix, has been used to predict conversion from MCI to AD (Oishi et al., 2012) (Fig. 4).
It is increasingly recognized that brain rhythms are essential for normal cognition. A simple way to perturb a brain rhythm is to alter myelination of the connecting fibers. There are many avenues to myelin disruption—exposure to toxic reagents, metabolic problems, inflammation, infection, and the list goes on (Filley and Fields, 2016). It is possible that myelin disturbances may cause network disruptions that ultimately lead to neuropsychiatric disorders. The DWI literature has linked changes in the fornix to different psychiatric disorders, including schizophrenia and psychosis, bipolar disorder, and eating disorders. We note that in all of these cases, the fornix is not selectively altered, but rather is one of the many affected white matter tracts.
Unlike the episodic memory literature, the psychosis literature is decidedly mixed on whether or not the fornix plays an important role in the pathophysiology of this disease, potentially due to small and heterogeneous sample populations, and difficulties encountered in imaging the fornix. The literature on bipolar disorder is more promising, and the literature on eating disorders is very small but is beginning to show interesting relationships between fornix microstructure and eating behavior. We suggest that future researchers interested in psychosis and/or bipolar disorder should measure episodic memory performance (preferably using a task with relational and spatial processing demands) as well as stress and allostatic load, and we correlate these measures with fornix microstructure, rather than clinical diagnosis per se. In addition, they should use optimized imaging methods as described in Box 1.
Our review exposed several gaps in the literature that we detail in Box 2. We note that the human literature on the fornix would benefit by drawing from the rich literature on the rodent fornix. For instance, lesion and DWI studies of humans should use tasks that depend on an intact fornix in rodents and monkeys (Table 1). Using tasks such as reversal learning or trace conditioning in humans would allow for greater cross-species comparison. In addition, delineating the contributions of each MTL subregion has been the primary focus of most memory models in terms of their distinct computational properties (Montaldi and Mayes, 2010; Norman and O'Reilly, 2003) and representation-specific processes (Davachi, 2006; Ranganath, 2010).
Big Questions for Future Research
Much controversy surrounds the functional divisions among the MTL gray matter subregions, most notably the hippocampus and MTL cortex including the perirhinal cortex, parahippocampal cortex, and entorhinal cortex. Given the crucial role of the fornix in the multitude of mnemonic processes outlined in this review, integrating this major white matter pathway in these existing theories would generate novel predictions about complex memory capacities expressed as products of an inter-connected circuitry.
In the future, quantitative DWI of the fornix may be able to reliably diagnose AD and predict conversion from MCI to AD. In addition, imaging of the fornix may be able to provide a differential diagnosis of AD, which is important, given that disorders such as depression can have a clinical presentation of memory impairments in the elderly. Apart from imaging the fornix, the effectiveness of treating neuropsychiatric disorders with DBS of the fornix has been modest thus far, but it has the potential for added benefit with modifications, such as employing closed-loop formats. Closed-loop stimulation of the fornix could even be used to pace or replace potentially aberrant hippocampal theta rhythms in AD (see optogenetic study by Giovannetti et al., 2018) and potentially improve mood, not only in AD but also in other disorders such as unipolar depression (see Mao et al., 2018). There is emerging evidence that scalp EEG can record theta oscillations (Crivelli-Decker et al., 2018), opening the door to future research that uses a noninvasive form of brain stimulation, rather than DBS, to modulate theta rhythms.
More research is needed to uncover the specific role of the fornix in neuropsychiatric disorders and to discover how best to target this tract for potentially alleviating symptoms or delaying onset of the disorders.
Footnotes
Acknowledgments
The authors would like to thank David Waxler for helpful comments. The content is solely the responsibility of the authors and does not necessarily represent the official views of the National Institute of Mental Health or the National Institutes of Health.
Author Disclosure Statement
No competing financial interests exist for any of the authors of this article.
Funding Information
This work was supported by National Institute of Health grants to I. Olson [R01 MH091113 and R21 HD098509].