
Abstract
Background:
The dorsal anterior cingulate cortex (dACC) is a key network hub for cognitive control and environmental adaptation. Previous studies have shown that task-based functional activity in this area is constrained by individual differences in sulcal pattern, a morphologic feature of cortex anatomy determined during fetal life and stable throughout development.
Methods:
By using anatomical magnetic resonance imaging and seed-based resting-state functional connectivity (rsFC), we explored the influence of sulcal pattern variability on the functional architecture of the dACC in a sample of healthy adults aged 20–80 years (n = 173).
Results:
Overall, rsFC was associated with individual differences in sulcal pattern. Furthermore, rsFC was modulated by the age–sulcal pattern interaction.
Conclusion:
Our results suggest a relationship between brain structure and function that partly traces back to early stages of brain development. The modulation of rsFC by the age–sulcal pattern interaction indicates that the effects of sulcal pattern variability on the functional architecture of the dACC may change over adulthood, with potential repercussions for brain network efficiency and cognitive function in aging.
Impact statement
We investigated for the first time the relationship between anatomical variability of the dorsal anterior cingulate cortex (dACC) and resting-state functional connectivity (rsFC) in young and older adult individuals. Seed-to-voxel rsFC analyses revealed that distinct dACC sulcal patterns are associated with specific profiles of connectivity strength and trajectories, and that this relationship changes as age increases. These findings provide novel insights into the relationship between early-determined neuroanatomical variants and brain functional architecture over adulthood.
Introduction
The dorsal anterior cingulate cortex (dACC) includes portions of the anterior and middle cingulate cortices on the brain's medial wall (Vogt and Palomero-Gallagher, 2012) and is known to mediate a set of regulatory functions critical for cognitive control and environmental adaptation (Botvinick et al., 2001; Bush et al., 2002; Sheth et al., 2012). The functional heterogeneity of the dACC is paralleled by an extensive variability in sulcal pattern, the set of fissures and folds that gives the cortical ribbon its characteristic convoluted appearance. The deepest, primary sulci emerge early during fetal life (14th–26th week of gestation) and show remarkable similarity between individuals, suggesting a deterministic imprinting (Borrell, 2018). Secondary and tertiary sulci develop as side-branches of primary sulci between the 30th and 36th week of gestation, yielding variable patterns that are likely influenced by in-womb and perinatal environmental factors (Amiez et al., 2018; Lohmann et al., 1999). Different from quantitative measures of cortex structure (e.g., cortical thickness, surface area, gyrification index), the dACC sulcal pattern is a morphologic feature determined prenatally and stable throughout development (Cachia et al., 2016; Tissier et al., 2018). The longitudinal stability of the dACC sulcal pattern and its resilience to neuroplastic changes* provide a clue to the study of individual differences in structure–function mappings, with the possibility to identify early neuroanatomical constraints on subsequent cognitive abilities (Borst et al., 2014; Fornito et al., 2004; Tissier et al., 2018). Although variability in dACC sulcation has been shown to affect the spatial distribution of local task-related functional activity (Amiez et al., 2013; Crosson et al., 1999; Jahn et al., 2016), it is largely unknown whether differences in sulcal pattern might alter the functional organization of the dACC in resting state, that is, in absence of overt task performance or stimulation. In the present study, we use structural magnetic resonance imaging (MRI) and seed-based resting-state functional connectivity (rsFC) of healthy adults (n = 173; age range = 20–80) to explore the influence of sulcal pattern variability on the functional architecture of the dACC. Moreover, since the brain's functional connectome encounters substantial changes with increasing age (Chen et al., 2016; Damoiseaux, 2017), we investigate the interaction between dACC sulcal pattern and age on rsFC to inspect for age-related changes in the relationship between local cortical morphology and dACC networks. Finally, we examine whether potential differences in connectivity profiles associated with sulcal pattern variability affect participants' performance on several cognitive test batteries.
Based on previous accounts (Jahn et al., 2016; Lopez-Persem et al., 2019), we hypothesize that, irrespective of age, seed-based rsFC will be associated with a distinct dACC sulcal pattern. In addition, although current evidence is insufficient to substantiate this prediction, the functional consequences of sulcal pattern variability may vary as a function of age, due to specific interactions between local cortical morphology and dACC networks in young and older participants. The physiologic mechanisms underlying the reported association between sulcal pattern variability and cognitive control performance are poorly understood (Cachia et al., 2014; Fornito et al., 2004; Huster et al., 2007). Here, we test the hypothesis that the association between dACC sulcal pattern and executive cognitive functions is mediated by the functional architecture of the dACC.
Materials and Methods
Participants
Participants' data were drawn from the publicly available Max Planck Institute “Leipzig Study for Mind-Body-Emotion Interactions” (LEMON) data set, which includes neuroimaging and behavioral data from 227 subjects (mean age 38.95; 82 F) (
Participants' Frequencies for Each Age Range Split by Gender
F, female; M, male.
Participants also underwent an extensive set of test batteries for cognitive assessment. To investigate whether the relationship between dACC sulcation and cognitive performance is modulated by functional connectivity, we selected tests tapping verbal and nonverbal cognitive processes that engage the dACC: California Verbal Learning Task (CVLT), Test of Attentional Performance (TAP), Trail Making Test (TMT), Wortschatztest (WST), Leistungsprüfsystem 2 (LPS 2), and Regensburger Word Fluency Test (RWT). For a detailed description of the test batteries, see Supplementary Data.
MRI acquisition
MP2RAGE sequences were acquired with a 3 Tesla scanner (MAGNETOM Verio; Siemens Healthcare GmbH, Erlangen, Germany) with the following parameters: repetition time (TR) = 5000 ms, echo time (TE) = 2.92 ms; flip angle 1 = 4° flip angle 2 = 5°, field of view (FOV) = 256 mm, number of slices = 176, voxel size = 1 mm3 isotropic, GRAPPA acceleration factor = 3, slice order = interleaved. T2*-weighted gradient echo planar imaging multiband BOLD scans were acquired with the following parameters: TR = 1400 ms, TE = 30 ms, flip angle = 69°, FOV = 202 mm, number of slices = 64, voxel size = 2.3 mm3 isotropic, imaging matrix = 88 × 88, slice thickness = 2.3 mm, echo spacing = 0.67 ms, bandwidth = 1776 Hz/pixel, partial Fourier 7/8, multiband acceleration factor = 4, volumes = 657, slice order = interleaved. For further details on MRI acquisition parameters, see Babayan and colleagues (2018).
dACC sulcal pattern classification
Differences in dACC sulcation are primarily assessed by quantifying the occurrence and characteristics of the paracingulate sulcus (PCS), a variable secondary sulcus that runs dorsal and parallel to the cingulate sulcus (CS) on the medial surface of the ACC (Ono et al., 1990; Paus et al., 1996; Yücel et al., 2001). Here, following Ono and colleagues' nomenclature (Ono et al., 1990), as in our previous studies (Cachia et al., 2017; Del Maschio et al., 2019), two types of dACC sulcation were identified depending on the PCS occurrence in each hemisphere: a “single” type (CS only) and a “double parallel” type (CS and additional PCS) (Fig. 1a, b). An asymmetry index based on this classification was used to assess the interhemispheric distribution of the PCS in each brain, yielding four sulcal patterns: (i) “single/single” (SS) (CS only in both hemispheres); (ii) “double parallel/double parallel” (DD) (CS and additional PCS in both hemispheres); (iii) “leftward asymmetry” (LA) (PCS present in the left hemisphere only); and (iv) “rightward asymmetry” (RA) (PCS present in the right hemisphere only).

The two hemispheres of the skull-stripped brain of the same individual (sub-032304) are shown.
All structural images were visually inspected for the presence of artifacts (e.g., excessive head movement, ghosting, field inhomogeneities or distortions) and six participants were excluded due to this reason. Moreover, two participants were excluded due to the presence of a calcified meningioma as assessed by an expert neurologist. Overall, T1-weighted MR images were obtained from 177 participants. The origin was set on all individual structural images to match the anterior commissure–posterior commissure (bicommissural) line; subsequently, segmentation and skull-stripping were performed by means of Statistical Parametric Mapping (SPM)12 (version 6906).
Following Garrison's PCS measurement protocol (Garrison, 2017), first reported in Garrison and colleagues (2015), all T1 images were imported into MANGO (Multi-image Analysis GUI, v 4.0,
PCS measurement and dACC sulcal pattern classification, incorporating both visual inspection and automated components, were carried by three expert blind raters (inter-rater agreement = 97%). Disagreement among raters due to ambiguous dACC sulcation (Leonard et al., 2009) was resolved by a fourth blind rater. Since ambiguity persisted after general inter-rater discussion for 4 participants (3% of the whole data set), those participants were excluded from subsequent analyses, yielding a final sample of 173 participants.
Seed-to-voxel rsFC
Data preprocessing and rsFC analyses were run with CONN toolbox (version 18.b) (Whitfield-Gabrieli and Nieto-Castanon, 2012) in SPM12. Using the default preprocessing pipeline implemented in CONN, functional images were slice-timing corrected, realigned to the first image, unwarped, and centered to (0, 0, 0) coordinates. Outlier volumes with excessive head motion were detected and scrubbed using the Artifact Rejection Toolbox (ART). Acquisitions with framewise displacement above 0.9 mm or global BOLD signal changes above 5 s.d. (over 95th percentile) were flagged as potential outliers. We allowed for scrubbing of up to 20% of functional volumes per subject. However, no subject in the final sample had to be discarded because of this reason. Structural images were segmented and normalized with the unified segmentation–normalization approach with default parameters (Ashburner and Friston, 2005). Functional images were normalized to average template in MNI space and smoothed with a full-width at half-maximum 8 mm Gaussian kernel. Since noise removal increases the reliability of rs-fMRI data, the anatomical component-based noise correction method (aCompCor) was applied to extract a representative noise signal from white matter (WM) and cerebrospinal fluid (CSF) components on a voxel-by-voxel level (Chai et al., 2012; Behzadi et al., 2007).
For each participant, realignment, scrubbing parameters, noise components from WM and CSF, and “effect of rest” were entered as nuisance covariates in the first-level analysis (Nieto-Castañón, 2020). Realignment parameters are defined as 12 components estimated from the subject motion parameters (3 translation, 3 rotation, and their first-order temporal derivatives). Scrubbing parameters consist in subject-specific dummy variables indicating outlier volumes identified with ART (Power et al., 2012). Physiological noise components from WM and CSF are defined as the significant principal components derived from nongray-matter “noise” regions in which the signal is supposed not to be modulated by the local neural activity (Behzadi et al., 2007). At first, WM and CSF masks are computed subject-wise by creating eroded-by-one-voxel binary masks of all the voxels with values greater than 50% within WM and CSF tissue probability maps. Afterward, five noise components within each mask are estimated. The first consisting the average BOLD signal and the other four as the result of a principal component analysis (PCA) following the aCompCor method as implemented by Chai and colleagues (2012) (see also Nieto-Castañón, 2020). In addition, a small trend at the very beginning of each participant's functional session (defined as “effect of rest” or “session effect”) is automatically estimated by CONN. This regressor corresponds to a step function convolved with the hemodynamic response function that allows to control for potential initial magnetization transient effects (or “ramping effects”) at the very beginning of the session, before the BOLD signal acquisition reaches the steady-state behavior. Subsequently to the linear regression of potential confounding effects, functional images were bandpass-filtered (0.01–0.1 Hz) to remove low-frequency drifts and physiological high-frequency noise.
A seed-to-voxel approach was implemented in CONN toolbox. Following Margulies and colleagues (2007), three parallel lines of spherical seeds (radius = 4 mm) were defined along the dorsal-to-caudal gradient of the dACC: (i) the ventral seed line lies 5 mm above the ACC callosal curve in the ACC; (ii) the middle seed line lies 15 mm above the callosal curve in the ACC, extending to the inferior part of the paracingulate gyrus (PCG), as defined by the anatomical mask of the probabilistic Harvard–Oxford atlas (distributed with the FMRIB Software Library FSL, Smith et al., 2004); and (iii) the dorsal seed line lies 25 mm above the callosal curve in the cingulo-frontal transition area, corresponding to the superior part of the PCG of the Harvard–Oxford atlas. † One set of seeds, comprising the three parallel lines of spherical seeds, was created for each hemisphere at x = ± 5 mm (see Fig. 1c, d, where also the MNI coordinates of the seeds are reported). A seed-to-voxel analysis with spherical seeds was preferred to selecting seed regions of interest (ROIs) based on anatomical parcellation. Indeed, anatomical variability (with specific reference to the cingulate cortex sulcation patterns) represents a source of large interpackage differences in brain parcellation schemes implemented in popular neuroimaging software, which may consequently jeopardize result generalizability and compatibility (Mikhael et al., 2018).
A whole-brain bivariate correlation analysis was carried out between the BOLD time-series of each seed (i.e., signals average across all voxels within the seed) and the BOLD time-series of all other voxels of the brain. The Pearson's correlation coefficients obtained were then converted to normally distributed scores using Fisher's R to Z transform to allow for second-level general linear model analysis. Significance threshold was set at voxel-p-uncorrected <0.001 and cluster-p-family-wise error (FWE)-corrected for multiple comparisons <0.05.
Statistical analyses
Statistical analyses were conducted to (i) explore the distribution of the dACC sulcal pattern in our sample; (ii) investigate the effects of dACC sulcal pattern variability on seed-based rsFC; (iii) assess whether the relationship between dACC sulcal pattern and seed-based rsFC changes with increasing age; and (iv) estimate correlations between connectivity profiles associated with distinct sulcal patterns and cognitive performance. Analyses were performed with CONN toolbox and SPSS software (version 21; IBM Corp. Released 2012).
dACC sulcal pattern distribution
Incidence rates (%) of dACC sulcal pattern were calculated for all participants and compared with equal frequency distribution by means of chi-square (χ2) goodness of fit. Since evidence of gender effects on sulcal pattern distribution is mixed (cf. Del Maschio et al., 2019; Leonard et al., 2009; Yücel et al., 2001), an ordinal logistic regression model was run to explore the distribution of the dACC sulcal pattern with gender as predictor (reference level: male) and dACC sulcal pattern as dependent variable (“single/single,” “double parallel/double parallel,” “leftward asymmetry,” “rightward asymmetry”).
Effects of dACC sulcal pattern variability on seed-based rsFC
To investigate the effects of individual differences in dACC sulcation on dACC connectivity, a general linear model was run with dACC sulcal pattern as regressors (“single/single,” “double parallel/double parallel,” “leftward asymmetry,” “rightward asymmetry”); age, gender, and education as nuisance covariates; and seed-based rsFC as a dependent variable. For each dACC seed (nine per hemisphere), the following paired differences t-contrasts were computed: (i) single/single>double parallel/double parallel; (ii) single/single>leftward asymmetry; (iii) single/single>rightward asymmetry; (iv) double parallel/double parallel>leftward asymmetry; (v) double parallel/double parallel>rightward asymmetry; and (vi) leftward asymmetry>rightward asymmetry. When each contrast was performed, other seeds were not covaried out. Connectivity differences between dACC sulcal patterns were tested each time for all the included seeds. Given the high number of performed univariate tests (i.e., 9 seeds*2 hemispheres*6 comparisons = 108 t-tests), the cluster-level FWE-corrected p-value threshold was additionally divided by the number of performed t-tests (i.e., 0.05/108 = 0.00046).
Task-based fMRI studies have indicated topological modulations of clusters of brain activity based on the sulcal morphology of the dACC. When interpreting results from an fMRI decision-making task, Amiez and colleagues (2013) have suggested that the CS (when a PCS is absent) and the PCS (when present) are functionally homologous. To test this hypothesis on rsFC, we have performed an additional analysis by comparing dorsal (D) and middle (M) seeds in individuals with and without a PCS. Unthresholded connectivity maps were generated to inspect spatial overlaps between M seeds and D seeds in the double parallel/double parallel and single/single groups. Connectivity differences were investigated by means of pairwise t-tests selecting middle seeds for the single/single group (i.e., M1, M2, M3) and dorsal seeds for the double parallel/double parallel group (i.e., D1, D2, D3) (Supplementary Data, Supplementary Table S1 and Supplementary Fig. S1).
Effects of age–sulcal pattern interaction on seed-based rsFC
To inspect for age-dependent changes in the relationship between dACC sulcal pattern and seed-based rsFC, a general linear model was run with dACC sulcal pattern*age as regressors (single/single*age; double parallel/double parallel*age; leftward asymmetry*age; rightward asymmetry*age), gender and education as nuisance covariates, and seed-based rsFC as a dependent variable. Contrasts were the same as those computed to investigate the effects of dACC sulcal pattern variability on seed-based rsFC. To further explore the interaction effects, significant connectivity changes that emerged from the age–sulcal pattern interaction analyses were investigated by splitting our sample into younger (age <35) and older adults (age >60) (Supplementary Data and Supplementary Table S2).
Correlations between dACC connectivity and cognitive performance
To test for cognitive effects of sulcal-related differences in dACC connectivity, correlational analyses were performed between dACC connectivity values associated with distinct sulcal patterns and performance at the following test batteries: CVLT, TAP, TMT, WST, LPS 2, and RWT. For each significant cluster resulting from the seed-to-voxel rsFC analyses, individual connectivity values were extracted. Correlations between cognitive scores and individual connectivity values were performed group-wise (e.g., for a cluster resulting from SS>DD contrast, connectivity scores of individuals with SS pattern and DD pattern were separately correlated with each of the selected cognitive tests). In case of significant correlations resulting from the same cluster with the same cognitive test, correlations coefficients were compared between groups. Following guidelines for improving brain–behavior correlations (Rousselet and Pernet, 2012), all analyses were performed with the freely available MATLAB toolbox “Robust Correlation Toolbox” (
Results
dACC sulcal pattern distribution
Absolute numbers and proportions of dACC sulcal pattern across participants and split by gender are reported in Table 2. The “leftward asymmetry” pattern occurred more frequently than others in our sample [LA = 32.37%; SS = 27.75%; DD = 20.81%; RA = 19.7%; χ2 (3) = 7.9249, p = 0.04759] confirming a well-known leftward bias in the general population (Fornito et al., 2004; Huster et al., 2007; Paus et al., 1996; Wei et al., 2017). No significant effect of gender was detected on dACC sulcal pattern distribution (β = −0.280, st.err. = 0.301, p > 0.3).
Sulcal Pattern Distribution in the Sample (N = 173)
Incidence rates (%) are calculated within groups and reported in brackets.
CS, cingulate sulcus; dACC, dorsal anterior cingulate cortex; DD, double parallel/double parallel’ (CS and additional PCS in both hemispheres); F, female; LA, leftward asymmetry (PCS present in the left hemisphere only); M, male; PCS, paracingulate sulcus; RA, rightward PCS asymmetry (PCS present in the right hemisphere only); SS, single/single (CS only in both hemispheres).
Effects of dACC sulcal pattern variability on seed-based rsFC
Individual differences in sulcal pattern were associated with distinct profiles of functional organization of the dACC. Results for each sulcal pattern are described below, reported in Table 3, and illustrated in Figure 2.

Effects of dACC sulcal pattern variability on seed-based resting-state functional connectivity. Results are reported as highlighted clusters overlayed onto unthresholded T-maps; color map indicates raw t-values (Pernet and Madan, 2020). Significant results are shown at cluster-level FWE-corrected for multiple comparisons p-value <0.00046, and voxel-level-uncorrected p-value <0.001. Axial slices are shown with Z MNI coordinates corresponding to the contrast local maxima with whole-brain height reference. Ventral seeds are labeled as V1, V2, V3; middle seeds are labeled as M1, M2, M3; dorsal seeds are labeled as D1, D2, D3. Images are in neurological display convention. dACC, dorsal anterior cingulate cortex; DD, double parallel/double parallel’ (CS and additional PCS in both hemispheres); FWE, family-wise error; L, left hemisphere; LA, leftward asymmetry (PCS present in the left hemisphere only); R, right hemisphere; RA, rightward PCS asymmetry (PCS present in the right hemisphere only); SS, single/single (CS only in both hemispheres). Color images are available online.
Paired Differences t-Contrast Results for the Main Effect of Dorsal Anterior Cingulate Cortex Sulcal Pattern on Seed-Based Functional Connectivity
Significance threshold is set at voxel-p-uncorrected <0.001 and cluster-p-FWE-corrected <0.00046. Voxel-p-FWE <0.05 corrected values are reported for additional information. Only one local maximum per significant cluster is reported.
FWE, family-wise error.
Single/single (CS only in both hemispheres)
The single/single>double parallel/double parallel contrast showed reduced rsFC of a ventral seed (V2 L) with the left insular cortex. Enhanced long-distance, intrahemispheric rsFC was found between a left middle (M3 L) seed and the cerebellum. The single/single>leftward asymmetry contrast was not associated with any significant difference. The single/single>rightward asymmetry contrast revealed extensive patterns of reduced long-distance rsFC between a right ventral seed (V1 R) and bilateral occipital cortices, as well as right temporo-occipital and cerebellar regions.
Double parallel/double parallel (CS and PCS in both hemispheres)
The double parallel/double parallel>single/single contrast showed enhanced rsFC of a ventral seed (V2 L) with the left insula. Reduced long-distance, intrahemispheric rsFC was found between a left middle (M3 L) seed and the cerebellum. The double parallel/double parallel>leftward asymmetry contrast showed enhanced intrahemispheric rsFC between a right ventral seed (V3 R) and the right angular gyrus, as well as between a left dorsal seed (D3 L) and bilateral insulo-opercular cortices. No difference in rsFC was found for the double parallel/double parallel>rightward asymmetry contrast.
Leftward asymmetry (PCS in the left hemisphere only)
The leftward asymmetry>single/single contrast showed no significant differences for any of the considered seeds. The leftward asymmetry>double parallel/double parallel contrast resulted in reduced intrahemispheric rsFC between a right ventral seed (V3 R) and the right angular gyrus, as well as between a left dorsal seed (D3 L) and bilateral insulo-opercular cortices. The leftward asymmetry>rightward asymmetry contrast showed no significant result.
Rightward asymmetry (PCS in the right hemisphere only)
The rightward asymmetry>single/single contrast revealed extensive patterns of enhanced long-distance rsFC between a right ventral (V1 R) seed and bilateral occipital cortices, as well as right temporo-occipital and cerebellar regions. No difference in rsFC was found for the rightward asymmetry>double parallel/double parallel contrast. The rightward asymmetry>leftward asymmetry contrast showed no significant difference as well.
Effects of age–sulcal pattern interaction on seed-based rsFC
The relationship between dACC sulcal pattern and dACC connectivity changed with increasing age, although to varying degree depending on the specific sulcal pattern considered. Results for each sulcal pattern are described below, reported in Table 4, and represented in Figure 3.
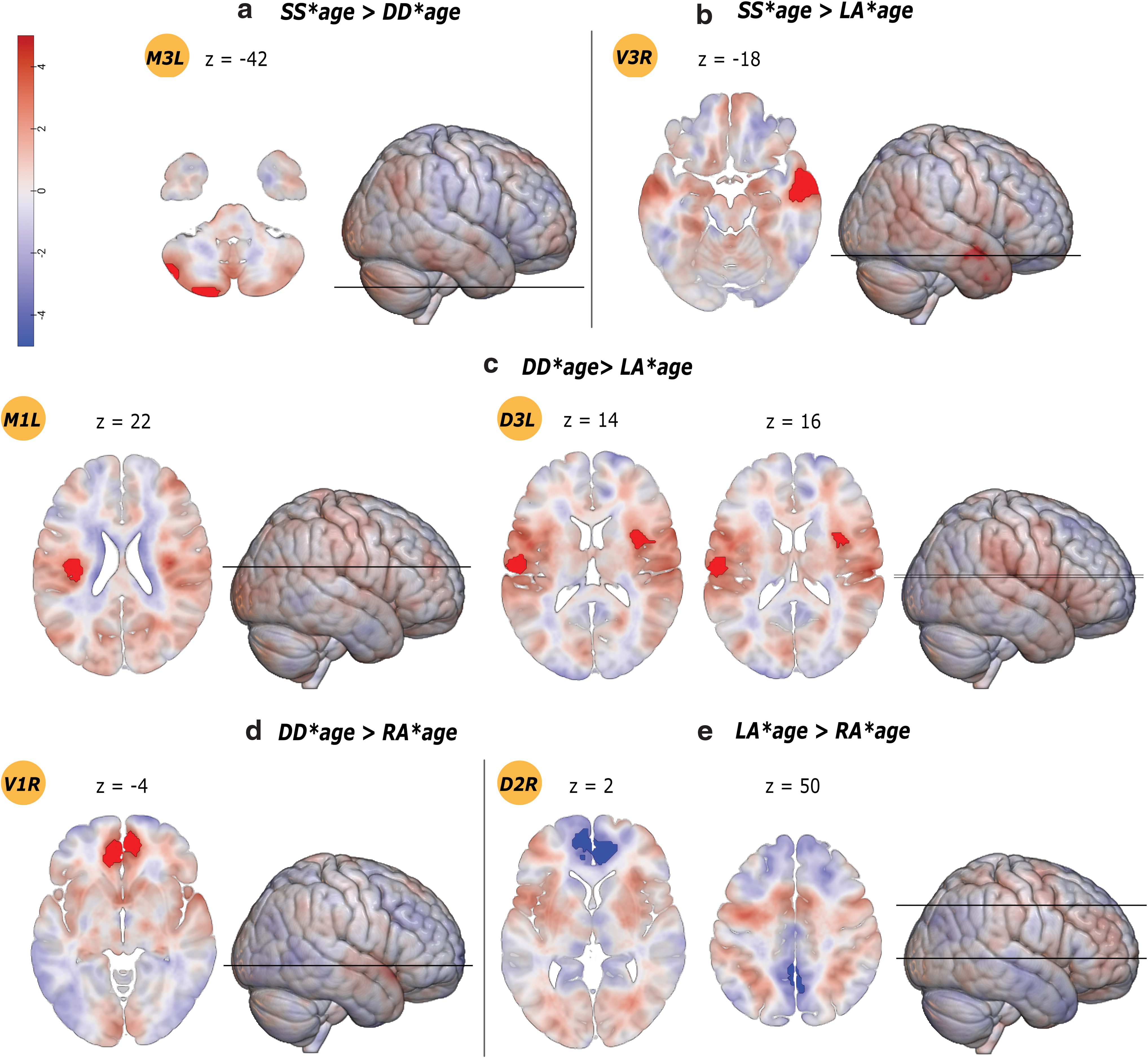
Effects of age–sulcal pattern interaction on seed-based resting-state functional connectivity. Results are reported as highlighted clusters overlayed onto unthresholded T-maps; color map indicates raw t-values (Pernet and Madan, 2018). Significant results are shown at cluster-level FWE-corrected for multiple comparisons p-value <0.00046, and voxel-level-uncorrected p-value <0.001. Axial slices are shown with Z MNI coordinates corresponding to the contrast local maxima with whole-brain height reference. Ventral seeds are labeled as V1, V2, V3; middle seeds are labeled as M1, M2, M3; dorsal seeds are labeled as D1, D2, D3. Images are in neurological display convention. Color images are available online.
Paired Differences t-Contrast Results of the Interaction Between Age and Anterior Cingulate Cortex Sulcal Pattern
Significance threshold is set at voxel-p-uncorrected <0.001 and cluster-p-FWE-corrected <0.00046. Voxel-p-FWE <0.05 corrected values are reported for additional information. Only one local maximum per significant cluster is reported.
Single/single*age
The single/single*age>double parallel/double parallel*age contrast showed increased long-distance, intrahemispheric rsFC between a left middle (M3 L) seed and the cerebellum. The single/single*age>leftward asymmetry*age contrast showed increased rsFC between a right ventral (V3 R) seed with the temporal cortex. The single/single*age>rightward asymmetry*age contrast showed no significant result.
Double parallel/double parallel*age
The double parallel/double parallel*age>single/single*age contrast showed decreased long-distance, intrahemispheric rsFC between a left middle seed (M3 L) and the cerebellum. The double parallel/double parallel*age>leftward asymmetry*age contrast revealed increased rsFC of middle (M1 L) and dorsal (D3 L) seeds with the bilateral central opercular cortex. The double parallel/double parallel*age>rightward asymmetry*age contrast showed increased rsFC between a right ventral seed (V1 R) and the ipsilateral PCG.
Leftward asymmetry*age
The leftward asymmetry*age>single/single*age contrast showed decreased rsFC with increasing age between a right ventral seed (V3 R) and the temporal cortex. The leftward asymmetry*age>double parallel/double parallel*age contrast revealed decreased rsFC of middle (M1 L) and dorsal (D3 L) seeds with the bilateral central opercular cortex. The leftward asymmetry*age>rightward asymmetry*age contrast showed decreased rsFC between a dorsal seed (D2 R) and the ACC and precuneus cortex.
Rightward asymmetry*age
The rightward asymmetry*age>single/single*age contrast showed no significant result. The rightward asymmetry*age>double parallel/double parallel*age contrast showed decreased rsFC between a right ventral seed (V1 R) and the ipsilateral PCG. The rightward asymmetry*age>leftward asymmetry*age contrast showed increased rsFC between a dorsal (D2 R) seed and the ACC and precuneus cortex.
Additional analyses with our sample split into younger (age <35) and older adults (age >60) are available in the Supplementary Data.
Correlations between dACC connectivity and cognitive performance
When testing for intergroup differences in correlations between connectivity values extracted from significant clusters and the scores from cognitive tests, no significant result emerged.
Discussion
We used a multimodal neuroimaging approach to investigate the influence of sulcal pattern variability on the functional organization of the dACC in a sample of healthy adults aged 20–80 years. Our results indicate that the effects of individual differences in local morphology can alter the functional architecture of the dACC at rest, with distinct sulcal patterns associated with specific profiles of functional connectivity strength and seed-based trajectories.
When compared with the “single/single” pattern (i.e., PCS absent in both hemispheres), patterns with a PCS in the right hemisphere (i.e., “rightward asymmetry” and “double parallel/double parallel” patterns) were associated with enhanced rsFC between ventral aspects of the dACC and the rest of the brain. The “double-parallel/double-parallel” pattern (i.e., PCS present in both hemispheres) was also associated with greater bilateral connectivity between ventral and dorsal seeds with the right angular gyrus and bilateral insulo-opercular cortices when compared with the “leftward asymmetry” pattern (i.e., PCS in the left hemisphere only), thus resulting the most connected pattern. On the contrary, the opposite “single/single” pattern was consistently linked with reduced rsFC, with the exception of a greater connectivity with the cerebellum relative to the “double parallel/double parallel” pattern. No differences were found between patterns with and without a PCS on the right hemisphere (i.e., “rightward asymmetry” and “double parallel/double parallel” vs. “leftward asymmetry” and “single/single”). Asymmetric patterns (i.e., “leftward asymmetry” and “rightward asymmetry”) were not different in terms of connectivity from each other. These results suggest profound differences between symmetric patterns (i.e., “double parallel/double parallel” vs. “single/single”) and a greater incidence of enhanced connectivity for the sulcal patterns with a PCS in the right hemisphere (i.e., “double parallel/double parallel” and “rightward asymmetry”).
In terms of seed-based trajectories, most target regions that showed significant differences associated with distinct sulcal patterns are key components of the salience network (SN) and default-mode network (DMN) (e.g., insular, opercular, and temporo-occipital cortices). This result is consistent with the central role played by the dACC as a connectivity hub in these intrinsic networks. The SN, a paralimbic–limbic network anchored in the dACC and the anterior insula with prominent subcortical nodes, plays a central role in social behavior and contributes to the processing of relevant stimuli by integrating sensory, emotional, and cognitive information (Menon, 2015; Seeley et al., 2007). The DMN is typically activated during passive rest states or internally oriented mental processes (e.g., mind wandering, self-referential processing) (Raichle, 2015). Differences in connectivity trajectories between closely placed seeds arguably reflect fine-graded changes in the cortical cytology of the ACC (Palomero-Gallagher et al., 2008, 2019). Finally, a cross-seed investigation between SS and DD patterns revealed that intergroup differences seem to reflect more a change in terms of connectivity strength than a change in functional topology (Amiez et al., 2013).
The neurobiological mechanisms underlying the association between cortical folding morphology and rsFC are not straightforward. The regional fissurization of the cortical surface has been related to variations in intra- and inter-regional structural connectivity (Hilgetag and Barbas, 2005; Toro and Burnod, 2005; Van Essen, 1997). Therefore, it would be tempting to speculate that differences in functional connectivity profiles associated with sulcal patterns are mediated by variations in the structural connectome. However, although brain regions that are well interconnected tend to display strong patterns of functional connectivity (Segall et al., 2012; Van Den Heuvel et al., 2009), the notion of a one-to-one relationship between structural and functional brain architecture has been challenged (Lim et al., 2019; Messé et al., 2014). We thus restrict ourselves to reporting that the functional connectivity of the dACC may be associated with individual differences in local cortical morphology, suggesting a relationship that partly arises from the fetal stages of brain development when sulcal patterns are determined.
When we tested for changes of the sulcal pattern-rsFC relationship dependent on aging, we found that the effects of sulcal pattern on rsFC changed from young to older adulthood, although to a varying, pattern-specific degree. For instance, when compared with all other sulcal patterns, the “leftward asymmetry” pattern was associated with extensively diminished seed-based rsFC with increasing age. By contrast, the functional organization of individuals with a symmetric sulcal pattern (i.e., double parallel/double parallel and single/single) was remarkably conserved when compared with the other patterns. The rs-fMRI literature reports substantive changes in rsFC over the adult life span, with decreased connectivity in association networks (e.g., DMN) and increased connectivity between functionally segregated networks (e.g., DMN/frontoparietal control network/somatosensory network) (Betzel et al., 2014; Geerligs et al., 2015; Grady et al., 2016). These patterns of effects have been associated with a progressive decrease in specialization of functional systems with potential detriment to information processing in older age groups. Age-related alterations in rsFC have been associated with a number of factors, including amyloid-β deposition (Ferreira and Busatto, 2013), neurovascular dysregulation (Galiano et al., 2020), and integrity across gray and white matter (Vidal-Piñeiro et al., 2014). Our findings suggest that sulcal pattern variability interacts with age-associated effects in shaping the brain's interregional connectivity shedding light on the long-term contribution of longitudinally stable neurodevelopmental markers on resting-state networks. Future longitudinal studies could investigate whether the double presence or double absence of a PCS may help individuals reach stable dACC connectivity patterns at an earlier age.
Increasing evidence shows that individual differences in dACC sulcal pattern relate to cognitive efficiency across several higher order functions that draw on cognitive control (Cachia et al., 2014; Del Maschio et al., 2019; Fornito et al., 2004). Compared with individuals with symmetric sulcal patterns, both children and adults with a leftward asymmetry pattern tend to exhibit a performance advantage on inhibitory control and working memory tasks (Borst et al., 2014; Cachia et al., 2017; Fornito et al., 2004; Huster et al., 2009; Tissier et al., 2018). The modulatory effect of sulcal pattern variability on cognitive control performance has been proposed to be mediated by differences in structural interhemispheric connectivity (Van Essen, 1997; see also Tissier et al., 2018) or functional brain activity, as hinted by task-based fMRI studies that show a topological modulation of functional responses associated with sulcation differences (Amiez et al., 2013; Crosson et al., 1999). We did not find evidence that changes in brain connectivity related to sulcal variability are also associated with differences in cognitive functions. However, since evidence for an association between sulcal variability and executive differences has been provided by other studies using different tests, a differentially modulated cognitive assessment may be required to obtain significant results.
The neurobiological interpretability of the data here presented may suffer from the lack of a univocal theory about the mechanisms regulating cortical expansion and folding (Fernández et al., 2016), representing a potential limitation to this study. The lack of a sufficient understanding of such mechanisms jeopardizes the possibility to provide fine-grained mechanistic explanations for the complex interplay between early neuroanatomical constraints, functional brain activity, and cognitive performance. What the present work highlights, however, is that variability in the dACC sulcal pattern—a longitudinally stable neuroanatomical fingerprint—is associated with the functional interdependence of the dACC with the rest of the brain, and interacts with age-associated effects in shaping connectivity changes over adulthood. Additional research utilizing multimodal imaging and behavioral data is needed to confirm these results and further investigate the cognitive effects of structure–function dynamics.
Conclusion
Taken together, our findings indicate that: (i) rsFC is associated with sulcal pattern variability, with distinct sulcal patterns associated with specific profiles of connectivity strength and trajectories (e.g., greater dACC connectivity associated with the presence of a PCS in the right hemisphere); (ii) the relationship between sulcal pattern and rsFC changes with increasing age, although to a varying, pattern-specific degree (e.g., more stable connectivity with increasing age for symmetric patterns and diminished connectivity with increasing age for the leftward asymmetry pattern); and (iii) cognitive effects associated with differences in structure–function relationships have not been detected on the performed test batteries.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.
Supplementary Material
Supplementary Data
Supplementary Figure S1
Supplementary Table S1
Supplementary Table S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.